
Trade-Offs among Release Treatments in Jack Pine Plantations: Twenty-Five Year Responses

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
- No competition control (C), in which, aside from site preparation, post-logging vegetation was left undisturbed.
- Motor-manual brushsaw cutting (BS), with non-crop vegetation cut at ground level with a motor-manual brushsaw in June 1993 (Bending Lake) or with non-crop vegetation cut at 25 cm above groundline with a Husqvarna 165 clearing saw in October 1994 (E.B. Eddy).
- Operational single aerial herbicide application (AS), with 4 L ha−1 of Vision® (1.5 hg acid equivalent [a.e.] ha−1) applied in a total spray volume of 35 L ha−1 using a G47T helicopter in late August 1992 (Bending Lake), or with 4 L ha−1 of Vision® (356 g L−1 a.e. glyphosate as a water soluble liquid) applied in a total spray volume of 34 L ha−1 using a Bell 206 helicopter in late August 1993 (E.B. Eddy).
- Complete competition removal (complete removal; CR), with annual backpack spray applications of a 2% solution (400 mL glyphosate in 20 L water) of Vision® applied in August 1993, June 1995, and June 1996 (Bending Lake) or September 1993 and August 1994 (E.B. Eddy). The amount of Vision® sprayed via backpack application varied among plots depending on how much vegetation survived previous applications; however, comparing the efficacy of different Vision® rates was beyond the scope of this study. Each complete removal treatment plot was randomly located in a single aerial spray plot.
2.2. Vegetation Assessment
2.3. Statistical Analyses
3. Results
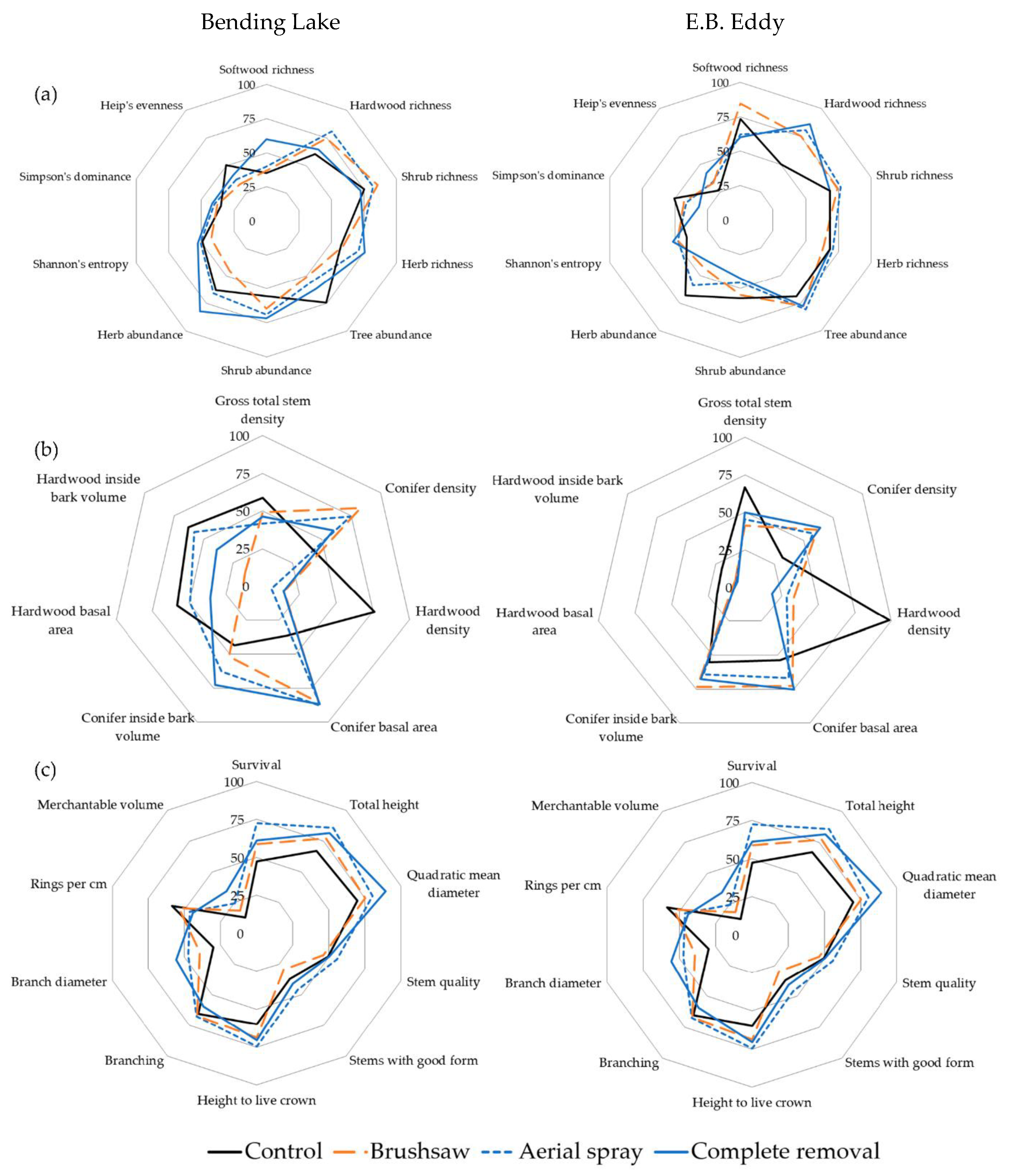
3.1. Plant Diversity
3.2. Stand-Scale Yield
3.3. Individual Jack Pine Characteristics
4. Discussion
4.1. Effect of Site and Species Ecology
4.2. Treatment Efficacy
4.3. Treatment Effects on Diversity
4.4. Treatment Effects on Stand-Scale Yield
4.5. Treatment Effects on Individual Jack Pine Characteristics
4.5.1. Survival, Height, and Diameter
4.5.2. Stem Form, Knot Size, and Branch Diameter
5. Conclusions and Recommendations
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gauthier, S.; Bernier, P.; Kuuluvainen, T.; Shvidenko, A.Z.; Schepaschenko, D.G. Boreal forest health and global change. Science 2015, 349, 819–822. [Google Scholar] [CrossRef]
- Natural Resources Canada. The State of Canada’s Forests: Annual Report 2019; Natural Resources Canada, Canadian Forest Service: Ottawa, ON, Canada, 2020; p. 84. Available online: https://cfs.nrcan.gc.ca/publications?id=40084 (accessed on 1 December 2020).
- FAO; UNEP. The State of the World’s Forests: Forests, Biodiversity and People; Food and Agriculture Organization of the United Nations: Rome, Italy, 2020; p. 214. [Google Scholar]
- Carle, J.; Holmgren, P. Wood from planted forest: A global outlook 2005–2030. For. Prod. J. 2008, 58, 6–18. [Google Scholar]
- Hearnden, K.W.; Millson, S.V.; Wilson, W.C. A Report on the Status of Forest Regeneration: Ontario Independent Forest Audit; Ontario Ministry of Natural Resources, Ontario Independent Forest Audit Committee: Sault Ste. Marie, ON, Canada, 1992; p. 117. [Google Scholar]
- Wagner, R.G.; Little, K.M.; Richardson, B.; McNabb, K. The role of vegetation management for enhancing productivity of the world’s forests. Forestry 2006, 79, 57–79. [Google Scholar] [CrossRef]
- Bell, F.W.; Dacosta, J.; Penner, M.; Morneault, A.; Stinson, A.; Towill, B.; Luckai, N.J.; Winters, J. Longer-term volume trade-offs in spruce and jack pine plantations following various conifer release treatments. For. Chron. 2011, 87, 235–250. [Google Scholar] [CrossRef] [Green Version]
- Canadian Forest Service. Criteria and Indicators for the Conservation and Sustainable Management of Temperate and Boreal Forests: The Montreal Process; Natural Resources Canada, Canadian Forest Service, Headquarters, Policy, Planning and International Affairs Branch: Ottawa, ON, Canada, 1995; p. 28. [Google Scholar]
- Liang, J.; Crowther, T.W.; Picard, N.; Wiser, S.; Zhou, M.; Alberti, G.; Schulze, E.-D.; McGuire, A.D.; Bozzato, F.; Pretzsch, H.; et al. Positive biodiversity-productivity relationship predominant in global forests. Science 2016, 6309, aaf8957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lieffers, V.J.; Armstrong, G.W.; Stadt, K.J.; Marenholtz, E. Forest regeneration standards: Are they limiting management options for Alberta’s boreal mixedwoods? For. Chron. 2008, 84, 78–82. [Google Scholar] [CrossRef] [Green Version]
- Willig, M.R. Biodiversity and productivity. Science 2011, 333, 1709–1710. [Google Scholar] [CrossRef] [PubMed]
- May, E.E. At the Cutting Edge—The Crisis in Canada’s Forests; Key Porter Books: Toronto, ON, Canada, 2005; p. 432. [Google Scholar]
- Bell, F.W.; Newmaster, S.G. The effects of silviculture disturbance on the diversity of seed-producing plants in the boreal mixedwood forests. Can. J. For. Res. 2002, 32, 1180–1191. [Google Scholar] [CrossRef]
- Mallik, A.U.; Bell, F.W.; Gong, Y. Effectiveness of delayed brush cutting and herbicide treatments for vegetation control in a seven-year-old jack pine plantation in north-western Ontario, Canada. Silva Fenn. 2002, 35, 505–519. [Google Scholar] [CrossRef] [Green Version]
- Dampier, J.E.E.; Luckai, N.; Bell, F.W.; Towill, W.D. Do tree-level monocultures develop following Canadian boreal silviculture? Tree-level diversity tested using a new method. Biodivers. Conserv. 2007, 16, 2933–2948. [Google Scholar] [CrossRef]
- Zheng, J.; Blanco, J.A.; Wei, X.; Liu, C. Sustainable management of Metasequoia glyptostroboides plantation forests in Shanghai. Forests 2018, 9, 64. [Google Scholar] [CrossRef] [Green Version]
- Fahlvik, N.; Ekö, P.-M.; Pettersson, N. Influence of precommercial thinning grade on branch diameter and crown ratio in Pinus sylvestris in southern Sweden. Scand. J. For. Res. 2005, 20, 243–251. [Google Scholar] [CrossRef]
- Sharma, M.; Zhang, S.Y. Stand density management diagram for jack pine stands in Eastern Canada. North J. Appl. For. 2007, 24, 22–29. [Google Scholar] [CrossRef] [Green Version]
- Briggs, D.G.; Smith, W.R. Effects of silvicultural practices on wood properties in conifers: A review. In Douglas-Fir: Stand Management for the Future; Hanley, D.P., Johnson, J.A., Eds.; College of Forest Resources, University of Washington: Seattle, WA, USA, 1985; No. 55; pp. 108–117. [Google Scholar]
- Zhang, S.Y.; Chauret, G.; Ren, H.Q.; Desjardins, R. Impact of initial spacing on plantation black spruce lumber grade yield, bending properties, and MSR yield. Wood Fiber Sci. 2002, 34, 460–475. [Google Scholar]
- Beniušiene, L.; Šilinskas, B.; Beniušis, R.; Aleinikovas, M.; Petrauskas, E.; Varnagiryte-Kabašinkiene, I. Scots pine stem parameters in sites with different stand densities in Lithuania. Forests 2020, 11, 716. [Google Scholar] [CrossRef]
- Wagner, R.G.; Buse, L.J.; Lautenschlager, R.A.; Bell, F.W.; Hollstedt, C.; Strobl, S.; Morneault, A.; Lewis, W.; Ter-Mikaelian, M.T. Vegetation Management Alternatives Program: 1994–1995 Annual Report; Ontario Ministry of Natural Resources, Ontario Forest Research Institute: Sault Ste. Marie, ON, Canada, 1995; p. 99. [Google Scholar]
- Pitt, D.G.; Morneault, A.E.; Bunce, P.; Bell, F.W. Five years of vegetation succession following vegetation management treatments in a jack pine ecosystem. North J. Appl. For. 2000, 17, 100–109. [Google Scholar] [CrossRef] [Green Version]
- Newmaster, S.G.; Bell, F.W. The effects of silvicultural disturbances on cryptogram diversity in the boreal-mixedwood forest. Can. J. For. Res. 2002, 32, 38–51. [Google Scholar] [CrossRef]
- Forest Co-op Growth and Yield Science Unit. Annual Report; The Forest Ecosystem Science Co-operative Inc.: Thunder Bay, ON, Canada, 2009. [Google Scholar]
- Sims, R.A.; Towill, W.D.; Baldwin, K.A.; Wickware, G.M. Field Guide to the Forest Ecosystem Classification for Northwestern Ontario; Canadian Forest Service, Ontario Ministry of Natural Resources: Sault Ste. Marie, ON, Canada, 1989; p. 191. ISBN 0777859904. [Google Scholar]
- Chambers, B.; Naylor, B.; Nieppola, J.; Merchant, B.; Uhlig, P. Field Guide to Forest Ecosystems of Central Ontario; SCSS FG-01; Ontario Ministry of Natural Resources, Southcentral Science Centre: North Bay, ON, Canada, 1997; p. 22. [Google Scholar]
- Bell, F.W.; Hunt, S.; Dacosta, J.; Sharma, M.; Larocque, G.R.; Winters, J.A.; Newmaster, S.G. Effects of silviculture intensity on plant diversity response patterns in young managed northern temperate and boreal forests. Écoscience 2014, 21, 327–339. [Google Scholar] [CrossRef]
- Honer, T.G.; Ker, M.F.; Alemdag, L.S. Metric Timber Tables for Commercial Tree Species of Central and Eastern Canada; Info. Rep. M-X-140; Environment Canada, Canadian Forest Service, Maritimes Forest Research Centre: Fredericton, NB, Canada, 1983; p. 139. [Google Scholar]
- Sharma, M. Comparing height-diameter relationships of boreal tree species grown in plantations and natural stands. For. Sci. 2016, 62, 70–77. [Google Scholar] [CrossRef]
- Sharma, M.; Parton, J. Height-diameter equations for boreal tree species in Ontario using a mixed-effects modeling approach. For. Ecol. Manag. 2007, 249, 187–198. [Google Scholar] [CrossRef]
- Sharma, M. Inside and outside bark volume models for jack pine (Pinus banksiana) and black spruce (Picea mariana) plantations in Ontario, Canada. For. Chron. 2019, 95, 50–57. [Google Scholar] [CrossRef]
- Sharma, M. Increasing volumetric prediction accuracy—An essential prerequisite for end-product forecasting in red pine. Forests 2020, 11, 1050. [Google Scholar] [CrossRef]
- Honer, T.G. Standard Volume Tables and Merchantable Conversion Factors for the Commercial Tree Species of Central and Eastern Canada; Info. Rep. FMR-X-5; Canadian Department of Forestry and Rural Development, Forest Management Research and Services Institute: Ottawa, ON, Canada, 1967; p. 21. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. Available online: https://www.R-project.org/ (accessed on 9 January 2021).
- Homagain, K.; Shahi, C.K.; Leitch, M.; Luckai, N.J.; Bell, F.W. Differences in extrinsic tree quality and value of fibre production following alternative vegetation management treatments in northwestern Ontario. For. Chron. 2011, 87, 251–259. [Google Scholar] [CrossRef] [Green Version]
- Thompson, I.D.; Guariguata, M.B.; Okabe, K.; Bahamondez, C.; Nasi, R.; Heymell, V.; Sabogal, C. An operational framework for defining and monitoring forest degradation. Ecol. Soc. 2013, 18, 20. [Google Scholar] [CrossRef]
- Condit, R.; Hubbell, S.P.; Lafrankie, J.V.; Sukumar, R.; Manokaran, N.; Foster, R.B.; Ashton, P.S. Species-area and species-individual relationships for tropical trees: A comparison of three 50-ha plots. J. Ecol. 1996, 84, 549–562. [Google Scholar] [CrossRef] [Green Version]
- Yang, T.-R.; Lam, T.Y.; Su, S.-H. A simulation study on the effects of plot size and shape on sampling plant species composition for biodiversity management. J. Sustain. For. 2018, 38, 116–129. [Google Scholar] [CrossRef]
- Pausas, J.G.; Austin, M.P. Patterns of plant species richness in relation to different environments: An appraisal. J. Veg. Sci. 2001, 12, 153–166. [Google Scholar] [CrossRef]
- Yang, Z.; Hautier, Y.; Borer, E.T.; Zhang, C.; Du, G. Abundance- and functional-based mechanisms of plant diversity loss with fertilization in the presence and absence of herbivores. Oecologia 2015, 179, 261–270. [Google Scholar] [CrossRef]
- Lichter, J. Primary succession and forest development on coastal lake Michigan sand dunes. Ecol. Monogr. 1998, 68, 487–510. [Google Scholar] [CrossRef]
- Kemperman, J. Sucker-Root Relationships in Aspen; Forest Research Note; Paper 4773; no. 12; Ministry of Natural Resources: ON, Canada, 1978. [Google Scholar]
- Farmer, R.J. Aspen root sucker formation and apical dominance. J. For. Sci. 1962, 8, 403–410. [Google Scholar]
- Frey, B.R.; Lieffers, V.J.; Landhäusser, S.M.; Comeau, P.G.; Greenway, K.J. An analysis of sucker regeneration of trembling aspen. Can. J. For. Res. 2003, 33, 1169–1179. [Google Scholar] [CrossRef]
- Perala, D.A. Populus tremuloides Michx.-Quaking Aspen. In Silvics of North America: Volume 2, Hardwoods; Agriculture Handbook 654; Burns, R.M., Honkala, B.H., Eds.; United States Department of Agriculture, Forest Service: Washington, DC, USA, 1990; pp. 555–569. [Google Scholar]
- Wang, G.G. Early regeneration and growth dynamics of Populus tremuloides suckers in relation to fire severity. Can. J. For. Res. 2003, 33, 1998–2006. [Google Scholar] [CrossRef]
- McCavour, M.J.; Paré, D.; Messier, C.; Thiffault, N.; Thiffault, E. The role of aggregated forest harvest residue in soil fertility, plant growth, and pollination services. Soil Sci. Soc. Am. J. 2014, 78, S196–S207. [Google Scholar] [CrossRef]
- Pickett, S.T.A.; Kempf, J.S. Branching patterns in forest shrubs and understory trees in relation to habitat. New Phytol. 1980, 86, 219–228. [Google Scholar] [CrossRef]
- Gregg, B.M.; Dougherty, P.M.; Hennessey, T.C. Growth and wood quality factors in relation to stand density and climatic factors. Can. J. For. Res. 2011, 18, 851–858. [Google Scholar] [CrossRef]
- Man, R.; Rice, J.A.; MacDonald, G.B. Early effects of pre- and post-harvest herbicide application and partial cutting in regenerating aspen—Jack pine mixtures in northeastern Ontario. Can. J. For. Res. 2011, 41, 1082–1090. [Google Scholar] [CrossRef]
- Fu, S.; Chen, H.Y.H.; Bell, F.W.; Sharma, M.; Delaney, J.R.; Peterson, G. Effects of timing of glyphosate application on jack pine, black spruce, and white spruce plantations in northern Manitoba. For. Chron. 2008, 84, 37–45. [Google Scholar] [CrossRef] [Green Version]
- Homagain, K.; Shahi, C.K.; Luckai, N.J.; Leitch, M.; Bell, F.W. Benefit-cost analysis of vegetation management alternatives: An Ontario case study. For. Chron. 2011, 87, 260–273. [Google Scholar] [CrossRef] [Green Version]
- Jobidon, R. Stump height effects on sprouting of mountain maple, paper birch and pin cherry—10 year results. For Chron. 1997, 73, 590–595. [Google Scholar] [CrossRef]
- Bell, F.W.; Morneault, A.E.; Pickering, S.M. Response of immature trembling aspen to season and height of cut. North J. Appl. For. 1999, 16, 108–114. [Google Scholar] [CrossRef] [Green Version]
- Bell, F.W.; Pitt, D.G. Seasonal susceptibility of boreal plants: Red raspberry phenology as a bioindicator of optimum within-season timing of glyphosate applications. For. Chron. 2007, 83, 733–741. [Google Scholar] [CrossRef] [Green Version]
- Wagner, R.G.; Robinson, A.P. Critical period of interspecific competition for four northern conifers: 10-year growth response and associated vegetation dynamics. Can. J. For. Res. 2006, 36, 2474–2485. [Google Scholar] [CrossRef] [Green Version]
- Ilisson, T.; Chen, H.Y.H. The direct regeneration hypothesis in northern forests. J. Veg. Sci. 2009, 20, 735–744. [Google Scholar] [CrossRef]
- Taylor, A.R.; Gao, B.; Chen, H.Y.H. The effect of species diversity on tree growth varies during forest succession in the boreal forest of central Canada. For. Ecol. Manag. 2020, 455, 117641. [Google Scholar] [CrossRef]
- Man, R.; Bell, F.W. Temporal changes of understory plant community in response to pre- and post-harvesting herbicide treatments and partial cutting in aspen-dominated boreal mixedwood stands. Eur. J. For. Res. 2018, 137, 337–348. [Google Scholar] [CrossRef]
- Kumar, P.; Chen, H.Y.H.; Thomas, S.C.; Shahi, C. Linking resource availability and heterogeneity to understorey species diversity through succession in boreal forest of Canada. J. Ecol. 2018, 106, 1266–1276. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, H.Y.H.; Taylor, A.R. Positive species diversity and above-ground biomass relationships are ubiquitous across forest strata despite interference from overstorey trees. Funct. Ecol. 2017, 31, 419–426. [Google Scholar] [CrossRef]
- Hedwall, P.; Holmström, E.; Lindbladh, M.; Felton, A. Concealed by darkness: How stand density can override the biodiversity benefits of mixed forests. Ecosphere 2019, 10, e02835. [Google Scholar] [CrossRef] [Green Version]
- Wagner, R.; Flynn, J.; Mertz, C.; Slovic, P.; Gregory, R. Acceptable practices in Ontario’s forests: Differences between the public and forestry professionals. New For. (Dordr.) 1998, 16, 139–154. [Google Scholar]
- Jobidon, R.; Cyr, G.; Thiffault, N. Plant species diversity and composition along an experimental gradient of northern hardwood abundance in Picea mariana plantations. For. Ecol. Manag. 2004, 198, 209–221. [Google Scholar] [CrossRef]
- Connell, J. Diversity in tropical rainforests and coral reefs. Science 1978, 199, 1302–1310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collins, S.; Glenn, S.; Gibson, D. Experimental analysis of intermediate disturbance and initial floristic composition: Decoupling cause effect. Ecology 1995, 76, 486–492. [Google Scholar] [CrossRef]
- Haeussler, S.; Bedford, L.; Leduc, A.; Bergeron, Y.; Kranabetter, M. Silvicultural disturbance severity and plant communities of the southern Canadian boreal forest. Silva Fenn. 2002, 76, 307–327. [Google Scholar] [CrossRef] [Green Version]
- Magnussen, S.; Smith, V.G.; Yeatman, C.W. Tree size, biomass, and volume growth of twelve 34-year-old Ontario jack pine provenances. Can. J. For. Res. 1985, 15, 1129–1136. [Google Scholar] [CrossRef]
- ESSA Technologies Ltd. FVSOntario User Guide: Forest Stand Growth Projection Modelling System (Version 3.08); ESSA Technologies Ltd.: Vancouver, BC, Canada, 2008. [Google Scholar]
- Guo, J.; Wang, J.R. Comparison of height growth and growth intercept models of jack pine plantations and natural stands in northern Ontario. Can. J. For. Res. 2006, 36, 2179–2188. [Google Scholar] [CrossRef]
- Zhang, L.; Peng, C.; Dang, Q. Individual-tree basal area growth models for jack pine and black spruce in northern Ontario. For. Chron. 2004, 80, 366–374. [Google Scholar] [CrossRef] [Green Version]
- Morris, D.; Reid, D.; Kwiaton, M.; Hunt, S.; Gordon, A. Comparing growth patterns of jack pine and black spruce in mixed natural stands and plantations. Écoscience 2014, 21, 1–10. [Google Scholar] [CrossRef]
- Barry, K.E.; Mommer, L.; van Ruijven, J.; Wirth, C.; Wright, A.J.; Bai, Y.; Connolly, J.; De Deyn, G.B.; de Kroon, H.; Isbell, F.; et al. The future of complementarity: Disentangling causes from consequences. Trends Ecol. Evol. 2019, 34, 167–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pretzch, H.; Schütze, G. Crown allometry and growing space efficiency of Norway spruce (Picea abies [L.] Karst.) and European beech (Fagus sylvatica L.) in pure and mixed stands. Plant Biol. 2005, 7, 628–639. [Google Scholar] [CrossRef] [Green Version]
- Kelty, M. Comparative productivity of monocultures and mixed-species stands. In The Ecology and Silviculture of Mixed-Species Forests; Kelty, M.J., Larson, B.C., Oliver, C.D., Eds.; Springer: Dordrecht, The Netherlands, 1992; Volume 40, pp. 125–141. ISBN 978-90-481-4135-7. [Google Scholar]
- Zhang, Y.; Chen, H.Y.H.; Reich, P.B. Forest productivity increases with evenness, species richness and trait variation: A global meta-analysis. J. Ecol. 2012, 100, 742–749. [Google Scholar] [CrossRef]
- Searle, E.B.; Chen, H.Y.Y. Complementarity effects are strengthened by competition intensity and global environmental change in the central boreal forests of Canada. Ecol. Lett. 2020, 23, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Reich, P.B.; Tilman, D.; Isbell, F.; Mueller, K.; Hobby, S.E.; Flynn, D.F.B.; Eisenhauer, N. Impacts of biodiversity loss escalate through time as redundancy fades. Science 2012, 336, 589–592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Chen, H.Y.H.; Searle, E.B.; Chen, C.; Reich, P.B. Negative to positive shifts in diversity effects on soil nitrogen over time. Nat. Sustain. 2020, 1–8. [Google Scholar] [CrossRef]
- Jucker, T.; Koricheva, J.; Finér, L.; Bouriaud, O.; Iacopetti, G.; Coomes, D. Good things take time—Diversity effects on tree growth shift from negative to positive during stand development in boreal forests. J. Ecol. 2020, 108, 2198–2211. [Google Scholar] [CrossRef]
- Brassard, B.W.; Chen, H.Y.H.; Cavard, X.; Laganière, J.; Reich, P.B.; Bergeron, Y.; Pare, D.; Yuan, Z. Tree species diversity increases fine root productivity through increased soil volume filling. J. Ecol. 2013, 101, 210–219. [Google Scholar] [CrossRef]
- Urli, M.; Thiffault, N.; Houle, D.; Gauthier, S.; Bergeron, Y. Role of green alder in boreal conifer growth: Competitor or facilitator? FACETS 2020, 5, 166–181. [Google Scholar] [CrossRef]
- Griess, V.; Knoke, T. Growth performance, windthrow, and insects: Meta-analyses of parameters influencing performance of mixed-species stands in boreal and northern temperate biomes. Can. J. For. Res. 2011, 41, 1141–1158. [Google Scholar] [CrossRef]
- Wagner, R.G. Competition and critical-period thresholds for vegetation management decisions in young conifer stands. For. Chron. 2000, 76, 961–968. [Google Scholar] [CrossRef] [Green Version]
- Kenkel, N.; Walker, D.; Watson, P.; Caners, R.; Lastra, R. Vegetation dynamics in boreal forest ecosystems. Coenoses 1997, 12, 97–108. [Google Scholar]
- Logan, K.T. Growth of Tree Seedlings as Affected by Light Intensity. II. Red Pine, White Pine, Jack Pine, and Eastern Larch; Publication 1160; Canadian Department of Forestry: Ottawa, ON, Canada, 1966; p. 19. [Google Scholar]
- Groot, A.; Adhikary, S.; Sharma, M.; Luckai, N.; Bell, F.; Larocque, G. Effect of species composition on the production rate and efficiency of young Picea glauca–Populus tremuloides forests. For. Ecol. Manag. 2014, 315, 1–11. [Google Scholar] [CrossRef]
- Jobidon, R.; Trottier, F.; Charette, L. Dégagement chimique ou manuel de plantations d’épinette noire? Étude de cas dans le domaine de la sapinière à bouleau blanc au Québec. For. Chron. 1999, 76, 973–979. [Google Scholar] [CrossRef] [Green Version]
- Bella, I.E.; DeFranceschi, J.P. Spacing Effects 15 Years after Planting Three Conifers in Manitoba; Information Report NOR-X-223; Canadian Forestry Service, Northern Forest Research Centre: Edmonton, AB, Canada, 1980; p. 10. [Google Scholar]
- Bella, I.E. Spacing Effects 20 Years after Planting Three Conifers in Manitoba; Forest Management Note 39; Canadian Forestry Service, Northern Forestry Centre: Edmonton, AB, Canada, 1987; p. 12. [Google Scholar]
- Cremer, K.W.; Borough, C.J.; McKinnell, F.H.; Carter, P.R. Effects of stocking and thinning on wind damage in plantations. N. Z. J. For. 1982, 12, 244–268. [Google Scholar]
- Tong, Q.J.; Zhang, S.Y. Impact of initial spacing and precommercial thinning on jack pine tree growth and stem quality. For. Chron. 2005, 81, 418–427. [Google Scholar] [CrossRef] [Green Version]
- Archibald, D.J.; Bowling, C. Jack Pine Density Management Diagram for Boreal Ontario; TN-005 (NWST TN-31); Ontario Ministry of Natural Resources, Northeast and Northwest Science and Technology: Thunder Bay, ON, Canada, 1995; p. 20. [Google Scholar]
- Lu, P.; Bell, F.; Charrette, P.; Thompson, M. Performance of jack pine (Pinus banksiana) rooted cuttings from proliferated dwarf shoots versus seedlings 8 years after planting. Can. J. For. Res. 2012, 42, 1–6. [Google Scholar] [CrossRef]
- Lautenschlager, R.; Sullivan, T. Effects of herbicide treatments on biotic components in regenerating northern forests. For. Chron. 2002, 78, 695–731. [Google Scholar] [CrossRef] [Green Version]
- Swift, K.I.; Bell, F.W. What are the environmental consequences of using silviculturally effective vegetation management alternatives? For. Chron. 2011, 87, 201–216. [Google Scholar] [CrossRef]
- Venier, L.A.; Thompson, I.D.; Fleming, R.; Malcolm, J.; Aubin, I.; Trofymow, J.A.; Langor, D.; Sturrock, R.; Patry, C.; Outerbridge, R.O.; et al. Effects of natural resource development on the terrestrial biodiversity of Canadian boreal forests. Environ. Rev. 2014, 22, 457–490. [Google Scholar] [CrossRef]
- Bell, F.W.; Thiffault, N.; Szuba, K.; Luckai, N.J.; Stinson, A. Synthesis of silviculture options, costs, and consequences of alternative vegetation management practices relevant to boreal and temperate conifer forests: Introduction. For. Chron. 2011, 87, 155–160. [Google Scholar] [CrossRef] [Green Version]
- Thiffault, N.; Roy, V. Living without herbicides in Québec (Canada): Historical context, current strategy, research and challenges in forest vegetation management. Eur. J. For. Res. 2011, 130, 117–133. [Google Scholar] [CrossRef]
- Wyatt, S.; Rousseau, M.H.; Nadeau, S.; Thiffault, N.; Guay, L. Social concerns, risk and the acceptability of forest vegetation management alternatives: Insights for managers. For. Chron. 2011, 87, 274–289. [Google Scholar] [CrossRef]
- Moffat, K.; Lacey, J.; Zhang, A.; Leipold, S. The social licence to operate: A critical review. Forestry 2015, 89, 477–488. [Google Scholar] [CrossRef] [Green Version]
- Kayahara, G.J.; Armstrong, C.L. Understanding First Nations rights and perspectives on the use of herbicides in forestry: A case study from northeastern Ontario. For. Chron. 2015, 91, 126–140. [Google Scholar] [CrossRef] [Green Version]
- Schneider, R.; Zhang, S.Y.; Swift, D.E.; Bgin, J.; Lussier, J.-M. Predicting selected wood properties of jack pine following commercial thinning. Can. J. For. Res. 2008, 38, 2030–2043. [Google Scholar] [CrossRef]
- Pielou, E.C. Ecological Diversity; John Wiley and Sons: New York, NY, USA, 1975; p. 165. [Google Scholar] [CrossRef]
- Simpson, E.H. Measurement of diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Heip, C. A new index measuring evenness. J. Mar. Biolog. Assoc. UK 1974, 54, 555–557. [Google Scholar] [CrossRef]


{kind=link}
| Site | Block | Geology | Preharvest Stand Conditions | Harvest | Site Preparation | Planting |
|---|---|---|---|---|---|---|
| Bending Lake | 1–4 (48°57′ N, 92°02′ W) | Flat to gently rolling; 1–10% bedrock outcrops; rapidly drained, coarse loamy to fine sand | V-17 ‡: jack pine mixedwood/shrub rich forest | Clearcut; tree length, conventional cut and skid, 1986–1987 | Mechanical; heavy drags (barrels and chains), 1987 | Block 1: Jack pine; Cerkon shelter cone seeding, 3000 seedlings ha−1 (1.8 × 1.8 m spacing), 1988 Blocks 2–4: Jack pine; overwintered Spencer-Lemaire 6 container stock; 3000 seedlings ha−1 (1.8 × 1.8 m spacing), 1988 |
| E.B. Eddy | 1 (46°47′ N, 82°08′ W) | Glacial outwash plain; fine sands to silty loams (ES13.1) * | FRI 1974 ±: Pw5Pj3Bw2 Site Class 2; age 150; height 24 m; stocking 0.7 | Clearcut; tree-length, conventional cut and skid, 1988 * | Young’s teeth, 1989 * | Jack pine; current 408 paper pot, planted with Potti Putki, June 27–29, 1989 † |
| 2 (46°47′ N, 82°10′ W) | Glacial outwash plain; silty loam (ES15.1) † | FRI 1974±: Pj7Sb2Po1 Site Class 2; age 85; height 20 m; stocking 0.8 | Clearcut; tree-length, conventional cut and skid, 1989 † | Young’s teeth, 1989 † | Jack pine; overwintered FH408 paper pot, planted with Potti Putki, May 13–16, 1991 † | |
| 3 (46°46′ N, 82°04′ W) | Glacial outwash plain; silty, very fine sands to silty loams (ES15.1) † | FRI 1974±: Pj6Sb2Po2 Site Class 2; age 90; height 21 m; stocking 0.6 | Clearcut; tree-length, conventional cut and skid, 1989 † | Young’s teeth, 1989 † | Jack pine; current 165 Jiffy pot, planted with Potti Putki, June 14, 1991† |
| Site | Treatment | Softwood Richness (Number of Species) | Hardwood Richness (Number of Species) | Shrub Layer Richness (Number of Species) | Herb Layer Richness (Number of Species) | Tree Abundance (Percent Cover) | Shrub Layer Abundance (Percent Cover) | Herb layer Abundance (Percent Cover) | Understory Diversity (H’) | Understory Dominance (D) | Understory Evenness (EHeip) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Bending Lake | Control | 1.8 * (0.2) | 2.4 (0.3) | 9.0 (0.5) | 11.5a (0.6) | 52.0 (3.7) | 41.9 (5.0) | 47.2 (4.2) | 1.48 (0.18) | 0.34 (0.06) | 0.41 * (0.05) |
| Brushsaw | 1.8 * (0.2) | 3.0 (0.3) | 10.3 (0.3) | 11.8a (0.6) | 35.4 (3.7) * | 48.8 (6.8) | 34.2 (4.8) | 1.29 (0.23) | 0.36 (0.08) | 0.31 (0.04) | |
| Aerial spray | 2.0 * (0.2) | 3.3 (0.2) | 9.8 (0.4) | 14.2b (0.8) | 39.0 (4.0) * | 52.2 * (7.1) | 49.5 (6.5) | 1.52 (0.22) | 0.38 (0.08) | 0.35 (0.07) | |
| Complete removal | 2.6 (0.3) | 3.0 (0.4) | 8.7 (0.5) | 15.1b (0.7) | 43.2 (4.0) * | 54.1 * (7.8) | 61.8 * (10.5) | 1.59 (0.22) | 0.40 (0.07) | 0.39 (0.05) | |
| E.B. Eddy | Control | 3.7 (0.3) | 2.0 (0.4) | 8.2 (0.4) | 13.7 (0.5) | 48.3 (2.9) | 43.2 (4.3) | 51.1 (10.0) | 1.23 (0.28) | 0.48 (0.12) | 0.22 (0.04) |
| Brushsaw | 4.2 (0.3) | 3.0 (0.2) | 8.9 (0.9) | 12.7 (1.3) | 54.4 (3.8) | 41.2 (5.6) | 33.0 (7.0) | 1.45 (0.30) | 0.41 (0.11) | 0.31 (0.05) | |
| Aerial spray | 3.1 (0.5) | 3.3 (0.4) | 9.2 (0.6) | 14.0 (0.8) | 56.4 (2.6) | 34.5 (6.4) | 44.2 (7.7) | 1.44 (0.29) | 0.40 (0.10) | 0.32 (0.05) | |
| Complete removal | 3.0 (0.4) | 4.0 (0.2) | 8.2 (0.9) | 13.7 (0.7) | 54.3 (3.8) | 32.1 (5.7) | 28.5 (3.8) | 1.56 (0.23) | 0.30 (0.08) | 0.39 (0.05) |
| Site | Treatment | Gross Total Stem Density (Stems ha−1) | Conifer Density (Stems ha−1) | Hardwood Density (Stems ha−1) | Conifer Basal Area (m2 ha−1) | Conifer Inside Bark Volume (m3 ha−1) | Hardwood Basal Area (m2 ha−1) | Hardwood Inside Bark Volume (m3 ha−1) |
|---|---|---|---|---|---|---|---|---|
| (n = 151) | (n = 81) | (n = 70) | (n = 81) | (n = 81) | (n = 70) | (n = 70) | ||
| Bending Lake | Control | 3518c (228) | 1233a (121) | 2285b * (287) | 11.8a (1.5) | 63.6a (8.1) | 10.6b * (1.3) | 57.0b * (6.9) |
| Brushsaw | 2910bc * (183) | 2498c * (157) | 452a * (73) | 27.7b (0.7) | 148b (5.2) | 0.69a (0.2) | 3.53a (0.9) | |
| Aerial spray | 2402ab (177) | 2236bc * (145) | 180a * (53) | 28.4b * (0.8) | 160b * (4.7) | 0.50a (0.2) | 2.90a (1.2) | |
| Complete removal | 2045a * (102) | 1794b (71) | 430a (166) | 28.2b (0.8) | 154b (5.5) | 0.60a (0.2) | 3.27a (1.0) | |
| E.B. Eddy | Control | 4011a (344) | 958a (163) | 3053b (353) | 10.8a (2.1) | 63.0a (10.9) | 13.8b (2.2) | 89.1b (19.0) |
| Brushsaw | 2331b (224) | 1842b (114) | 989a (219) | 26.9b (0.9) | 154b (7.1) | 2.38a (0.7) | 13.6a (4.1) | |
| Aerial spray | 2561b (149) | 1736b (108) | 850a (143) | 22.7b (1.1) | 128b (4.9) | 1.65a (0.3) | 8.24a (1.5) | |
| Complete removal | 2725b (177) | 1925b (89) | 550a (173) | 27.3b (0.9) | 152b (8.8) | 1.31a (0.4) | 6.90a (2.3) |
| Source | df | Survival | Stem Quality | Branching | |||
|---|---|---|---|---|---|---|---|
| F | p-Value | F | p-Value | F | p-Value | ||
| Site * | 1 | 5.8 | 0.016 | 30.41 | <0.001 | 4.12 | 0.048 |
| Treatment * | 3 | 17.71 | <0.001 | 1.14 | 0.334 | 8.67 | <0.001 |
| CR vs. AS | 1 | 1.651 | 0.099 | NA | NA | 2.39 | 0.017 |
| CR vs. BS | 1 | 1.068 | 0.286 | NA | NA | 3.31 | 0.001 |
| AS vs. BS | 1 | 1.79 | 0.074 | NA | NA | 0.78 | 0.435 |
| C vs. treatments | 1 | 7.06 | <0.001 | NA | NA | 3.8 | <0.001 |
| Site | Treatment | Total Height (m) | Quadratic Mean Diameter (cm) | Height to Live Crown (m) | Branch Diameter (mm) | Rings (Per cm) | Merchantable Volume (m3 ha−1) |
|---|---|---|---|---|---|---|---|
| Bending Lake | Control | 9.5a * (0.4) | 11.0a (0.4) | 6.1a (1.1) | 11.5a (2.0) | 2.2b * (0.1) | 33.3a * (3.3) |
| Brushsaw | 11.4b * (0.2) | 11.9b (0.3) | 7.2b (0.8) | 16.6b * (2.4) | 2.0ab * (0.1) | 97.4b * (6.1) | |
| Aerial spray | 12.7c (0.1) | 12.7b (0.3) | 7.8b (1.2) | 38.9d * (2.5) | 1.7a * (0.1) | 114.1c * (7.4) | |
| Complete removal | 11.9bc (0.2) | 14.2c (0.2) | 7.3b (1.1) | 22.8c (2.8) | 1.6a * (0.1) | 129.2d (5.1) | |
| E.B. Eddy | Control | 10.8a (0.4) | 13.5ab (0.3) | 7.0a (0.2) | 13.3a (1.3) | 1.7b (0.1) | 32.6a (5.5) |
| Brushsaw | 12.4b (0.2) | 13.4ab (0.5) | 7.0a (0.2) | 19.8b (0.7) | 1.4a (0.1) | 93.9b (5.8) | |
| Aerial spray | 12.9b (0.2) | 12.0a (0.3) | 7.6b (0.2) | 23.5c (0.9) | 1.2a (0.1) | 118.1c (7.4) | |
| Complete removal | 12.3b (0.3) | 12.9a (0.2) | 7.2ab (0.2) | 25.4c (1.0) | 1.3a (0.1) | 127.1c (4.8) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deighton, H.D.; Bell, F.W.; Thiffault, N.; Searle, E.B.; Leitch, M.; Sharma, M.; Dacosta, J. Trade-Offs among Release Treatments in Jack Pine Plantations: Twenty-Five Year Responses. Forests 2021, 12, 370. https://doi.org/10.3390/f12030370
Deighton HD, Bell FW, Thiffault N, Searle EB, Leitch M, Sharma M, Dacosta J. Trade-Offs among Release Treatments in Jack Pine Plantations: Twenty-Five Year Responses. Forests. 2021; 12(3):370. https://doi.org/10.3390/f12030370
Chicago/Turabian StyleDeighton, Holly D., Frederick Wayne Bell, Nelson Thiffault, Eric B. Searle, Mathew Leitch, Mahadev Sharma, and Jennifer Dacosta. 2021. "Trade-Offs among Release Treatments in Jack Pine Plantations: Twenty-Five Year Responses" Forests 12, no. 3: 370. https://doi.org/10.3390/f12030370