1. Introduction
Disturbances such as tree-killing bark beetles are a driver of tree mortality in North American forests and a concern for forest managers. Across North America, bark beetle outbreaks have resulted in widespread tree mortality in multiple forest types [
1,
2]. Understanding risk factors for individual trees and stands is important to determine where outbreaks are likely to occur and which trees may be more susceptible. However, the manner in which stand dynamics may interact with the site climate to influence mortality risk in a bark beetle outbreak is poorly understood.
Tree mortality as a result of bark beetle outbreaks is often correlated with site and forest structural characteristics, as well as individual tree characteristics within a stand [
3]. Higher elevations, where extreme winter temperatures may limit the survival of bark beetle larvae, have previously been associated with lower tree mortality [
4,
5,
6]. Dense stands [
6,
7,
8,
9,
10,
11] and low species diversity [
12,
13,
14,
15,
16] may contribute to higher stand susceptibility due to a greater density of suitable host trees and a higher abundance of trees experiencing drought stress from overcrowding [
17,
18]. Similarly, trees with thinner crowns [
19,
20] and trees occupying subordinate positions in the canopy [
7,
21,
22] are often more susceptible due to decreased fitness. Large [
2,
3,
4,
7,
23,
24] and older [
3,
18,
24] trees are often more susceptible due to both decreased defenses and minimum host size requirements. Further, climate plays a role in regulating bark beetle-related tree mortality. Precipitation trends, including droughts [
6,
25,
26] and long-term regional climate averages [
27,
28], have been correlated with beetle-related mortality. Drought-stressed trees may have attenuated defenses that render them more susceptible to bark beetle-related mortality ([
29,
30] and references therein).
Consequently, site climate, as well as tree- and stand-level variables, can be helpful in determining mortality risk during a bark beetle outbreak, but interactions between these variables are rarely synthesized or compared within one study. Many stand-level variables may influence the environmental stress experienced by individual trees, which in turn, may influence beetle-related mortality. Drought stress emerges from a complex interplay between stand conditions, climate, and soil conditions. Stand conditions that produce drought-stressed trees in one location may not have the same impact in another. For instance, trees in dense stands may be more prone to drought stress due to greater competition for moisture, and trees located on south-facing slopes may be hotter and drier. Given the interplay between drought stress and host-tree susceptibility described in previous studies [
6,
25,
26,
29,
30], an expected pattern of tree–environment–beetle interactions is that thresholds of stand density resulting in heightened mortality susceptibility may be lower in relatively dry environments than in wetter environments. Large trees, which require more water than small trees, may similarly be most stressed and thus more susceptible to attack in low-precipitation environments. Aspect, or related physiographic variables such as heat load index, may be most influential in the hottest portions of a tree species’ range, where south-facing slopes are likely to reach more extreme temperatures. Still, few studies have addressed these potential interactions between stand structure, physiography, climate, and probability of beetle-driven tree mortality. While it is clear that many stand variables influence mortality risk, it is less clear whether the rate at which mortality risk increases as a function of stand conditions depends on how hot or dry a particular site may be, and if so, which stand- or tree-level variables are affected.
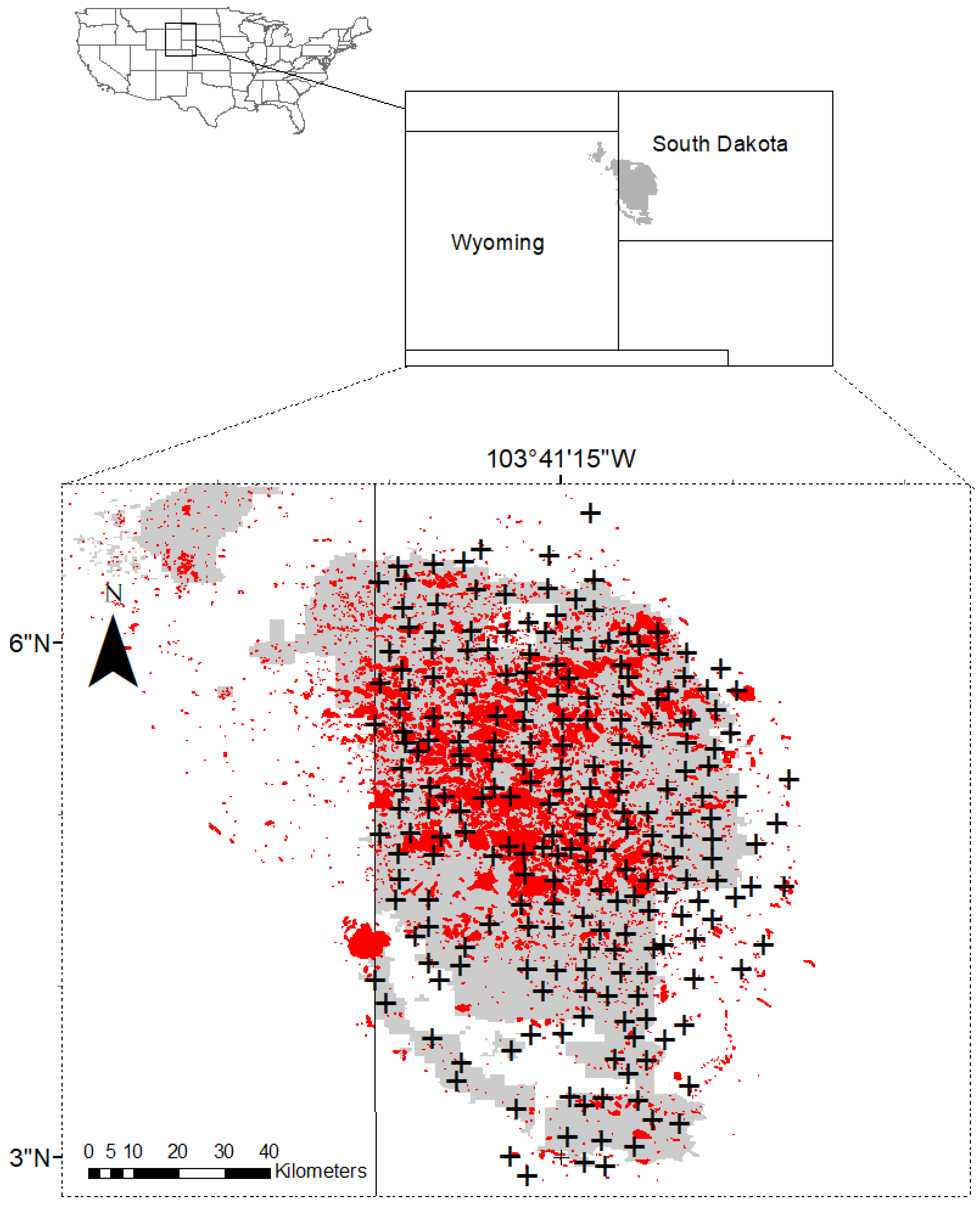
To address these gaps in understanding, this study examines ponderosa pine forests of the Black Hills, in western South Dakota, USA. The mountain pine beetle (
Dendroctonus ponderosae Hopkins) reached outbreak proportions in the Black Hills beginning in the early 2000s, and by 2016, over 180,000 hectares had been affected by beetle damage [
31]. Although the exact cause of the outbreak is undetermined, the gradual accumulation of viable host material (i.e., ponderosa pines of sufficient size to host mountain pine beetle) in relatively dense, homogenous stands may have contributed [
31]. We analyzed Forest Inventory and Analysis (FIA) data from this area to address three objectives: (1) Identify the stand- and tree-level factors influencing the insect-related mortality hazard during a bark beetle outbreak, (2) determine whether climate can influence the mortality hazard when controlling for tree and stand properties, and (3) determine how the relationship between stand properties and mortality risk is influenced by climate. Our analyses provide new information on the role of environmental controls as mediators of mortality during a bark beetle outbreak, and describe how these controls interact with tree- and site-level factors to predict tree survival. These results have consequences for forest managers concerned with managing stands for resistance to bark beetle damage.
4. Discussion
Several factors contributed to insect-related tree mortality during the recent bark beetle outbreak in the Black Hills, South Dakota. Although we did find a significant interaction between elevation and precipitation, the effect of stand density, tree size, and crown ratio on mortality was not mediated by the relative moistness of the site. This indicates that while climate may influence some of the factors that predispose trees to beetle-caused mortality, many of those factors are climate-independent.
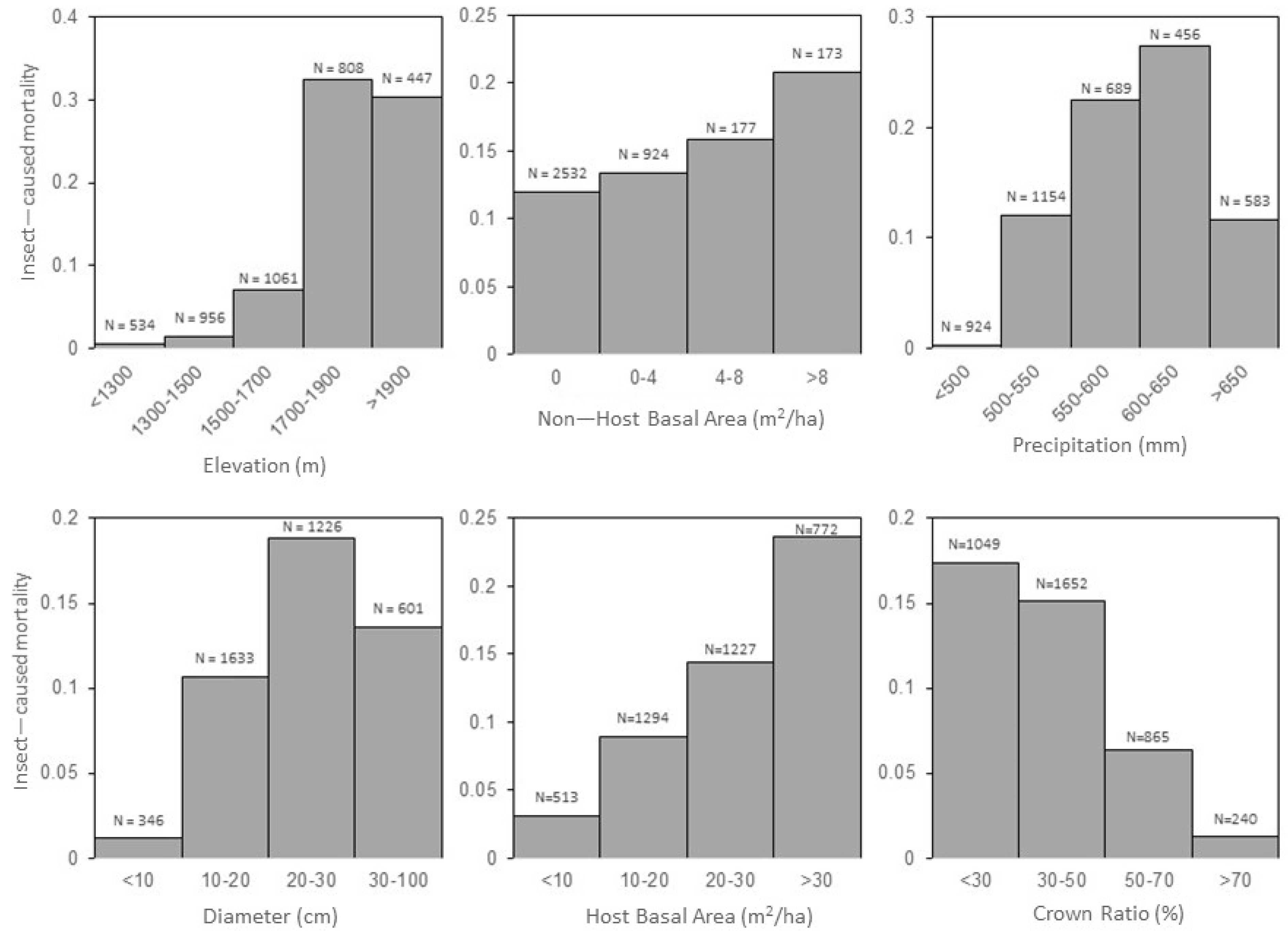
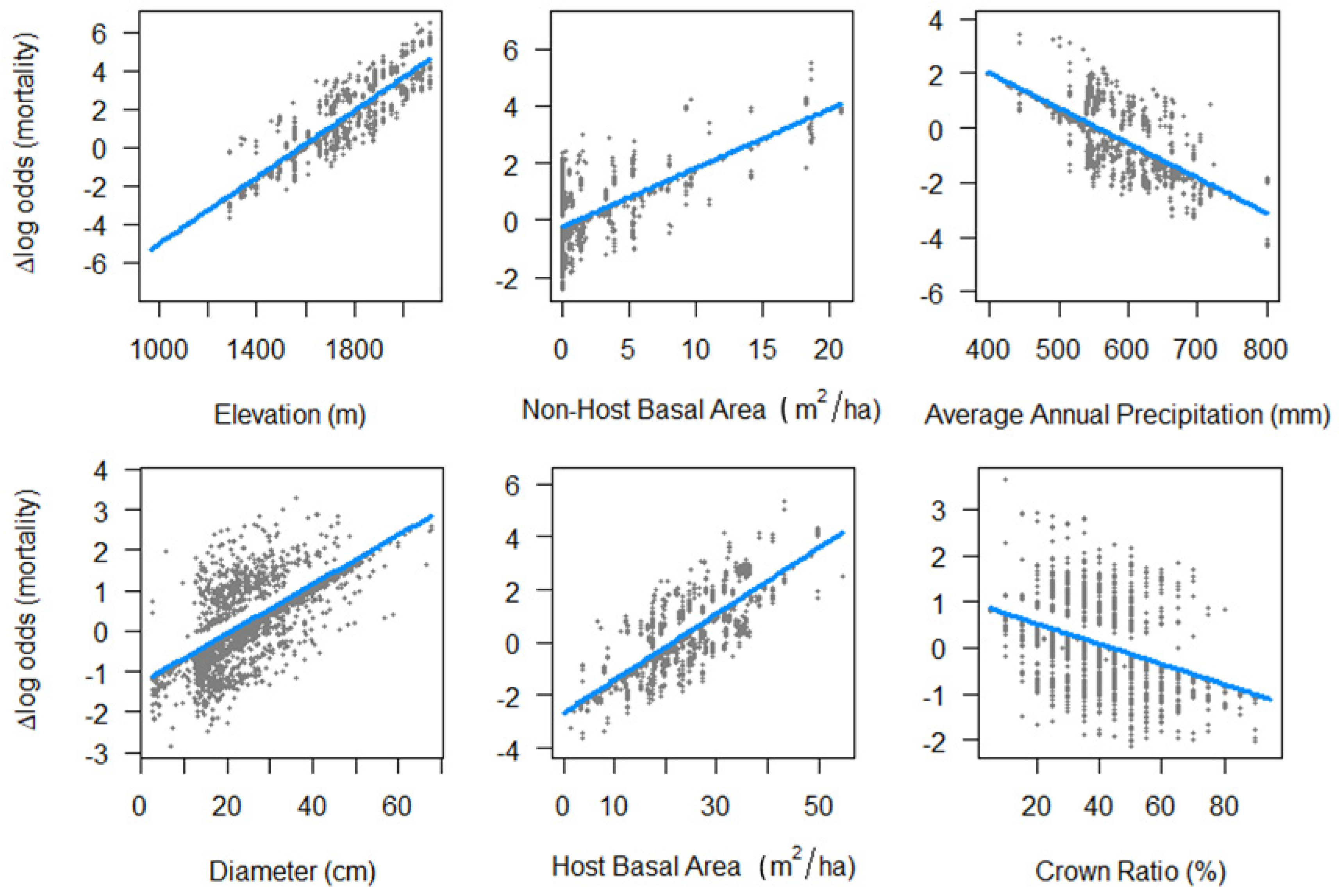
At the level of the individual tree, larger diameters and smaller crowns were associated with higher mortality rates (
Figure 2 and
Figure 3). The association between larger trees and higher mortality risk is consistent with past studies [
7,
9,
23,
45,
46] that have examined mountain pine beetle-related mortality in ponderosa pine. Although rarely examined in the context of beetle-related mortality, crown ratio is an oft-used proxy for tree vigor [
19,
20,
24] and larger crowns have been associated with lower rates of general tree mortality [
47], so the association with lower beetle-related mortality is not surprising. Despite past work associating canopy position with mortality risk [
7], we did not find a significant link, possibly because canopy position was strongly correlated with diameter (Pearson’s coefficient = 0.57), and diameter is critical in quantifying mountain pine beetle susceptibility.
At the stand level, higher elevations and denser stand conditions were associated with higher mortality rates (
Figure 2 and
Figure 3). The positive relationship observed between live basal area and mortality hazard during a bark beetle outbreak is well-established for bark beetles generally [
6,
8,
10,
48] and
D. ponderosae specifically [
7,
9,
11,
49]. Interestingly, our model revealed that it did not matter whether the live basal area was contributed by ponderosa pine or a nonhost species, such as aspen or white spruce—the addition of live basal from either group contributed equally to higher mortality risk. This indicates that individuals from pure stands were just as likely to experience mortality as individuals from relatively heterogenous, species-diverse stands of a similar density.
Prior research on the impact of nonhost trees on bark beetle outbreaks is mixed. Several studies indicate that host trees located in stands containing higher proportions of nonhost material suffer lower rates of mortality [
50,
51], an assumption that is maintained in the stand hazard rating developed for
D. ponderosae-P. ponderosa in the Black Hills for the Forest Service’s National Insect and Disease Risk Map [
15]. However, other work from ponderosa pine forests in Colorado indicates that the presence of nonhost material in the stand can enhance mortality risk for host trees [
7], which is consistent with our results. While nonhost species cannot amplify pest populations, they still compete with host individuals for moisture, which may explain why mortality risk from our study increased with nonhost basal area. However, conifer forests in the Black Hills are relatively homogeneous, so effects of species diversity on bark beetle-driven tree mortality may not be fully captured in this study.
While we identified a significant main effect of basal area, there was no evidence of a significant interaction with precipitation. This indicates that high-basal area stands are prone to beetle-related mortality across the full gradient of precipitation in the Black Hills, and that high-basal area stands are no more susceptible in dry environments than moist environments within the study region. Past research [
52] reasoned that the live basal area could be increased in productive sites without an associated increase in mortality risk from beetles, but our data do not match this prediction, though productivity is influenced by more than just precipitation. Our results may support other hypotheses that dense stands promote beetle-caused mortality through moisture-independent pathways, such as by creating cooler microenvironments conducive to larval beetle development [
8] and/or by suppressing winds that could interfere with aerial pheromones [
31]. Alternatively, the lack of a precipitation x basal area interaction could be due to the relatively high intensity of the outbreak, which may have led to stands experiencing little drought stress being targeted by bark beetles at the same intensity as drought-stressed stands. The gradient of precipitation in the Black Hills may also not have been broad enough to detect an interaction, and analyses of other regions with different precipitation regimes may produce different results.
The strong relationship observed between higher elevation and increased mortality is one that, to our knowledge, has not been previously documented for mountain pine beetle outbreaks in ponderosa pine. Below 1500 m, the observed mortality rate during the study period was ~1%, but above 1700 m, the mortality rate rose to over 30% (
Figure 2). While elevation has been shown to influence bark beetle-related mortality in other areas, the effect has typically been in the opposite direction. Past work has documented reduced bark beetle-associated mortality at higher elevations for
D. ponderosae in lodgepole pine [
4,
5] and
Ips in ponderosa pine [
6], but those studies were conducted at higher elevations than ours. Lower rates of beetle-caused tree mortality at high elevation are thought to be caused by extreme low winter temperatures, which can kill brood developing below the bark, or through increased winter precipitation inputs that improve tree access to water during summer months [
4,
5,
6]. This is unlikely to happen in the Black Hills, as the highest elevations in the Black Hills do not experience low winter temperatures needed to kill larval beetles [
5,
38]. Extreme highs—and not extreme lows—offer one alternative explanation for the relative dearth of insect-related mortality observed at low elevation in the Black Hills, as the mortality of larval bark beetles has been documented at high temperatures [
53]. Other nonclimatic factors, such as soil quality or relative topographic position (i.e., whether a stand is located at the top of a ridge vs. the bottom of a draw) could also provide mechanistic explanations for the higher mortality rates observed at high elevation.
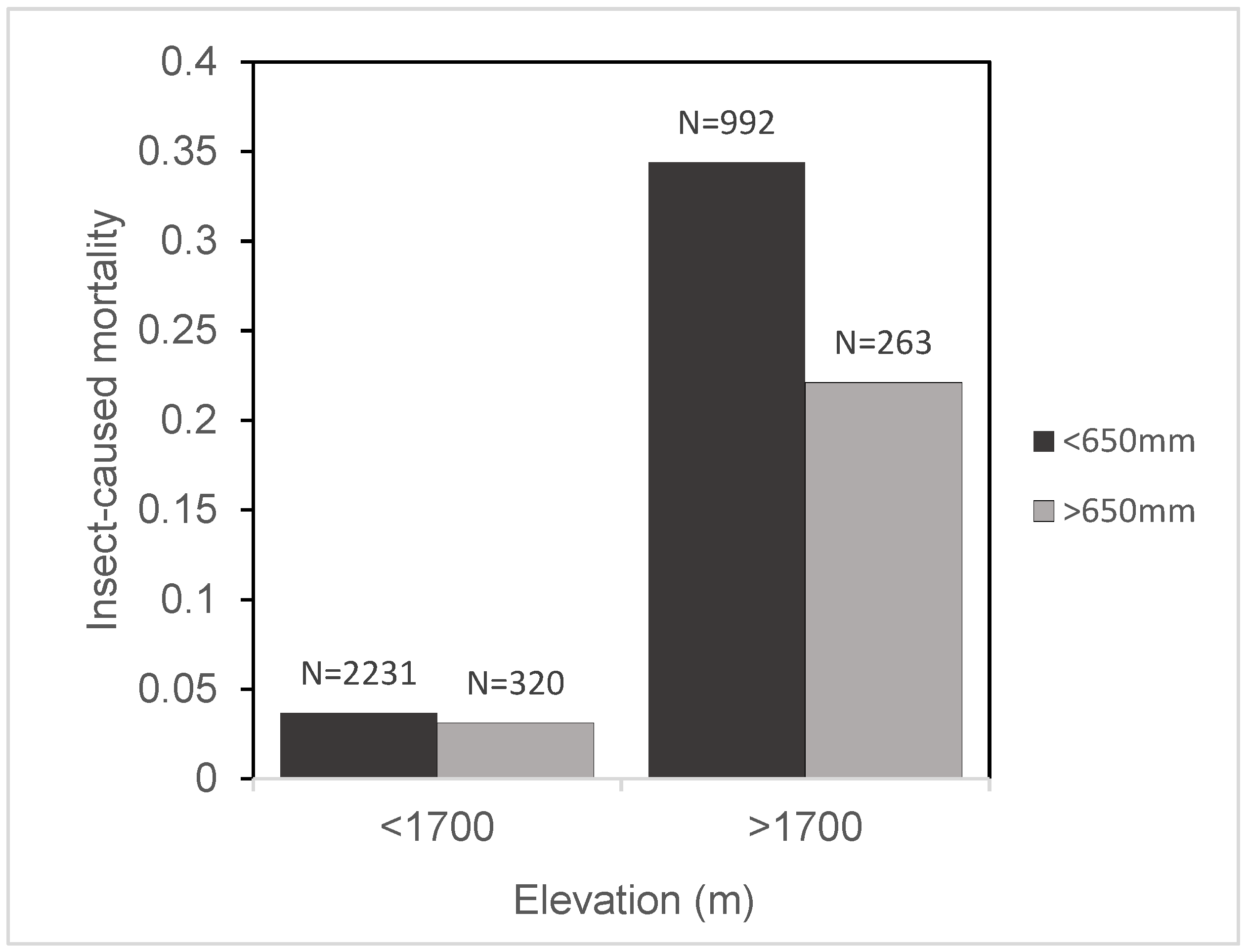
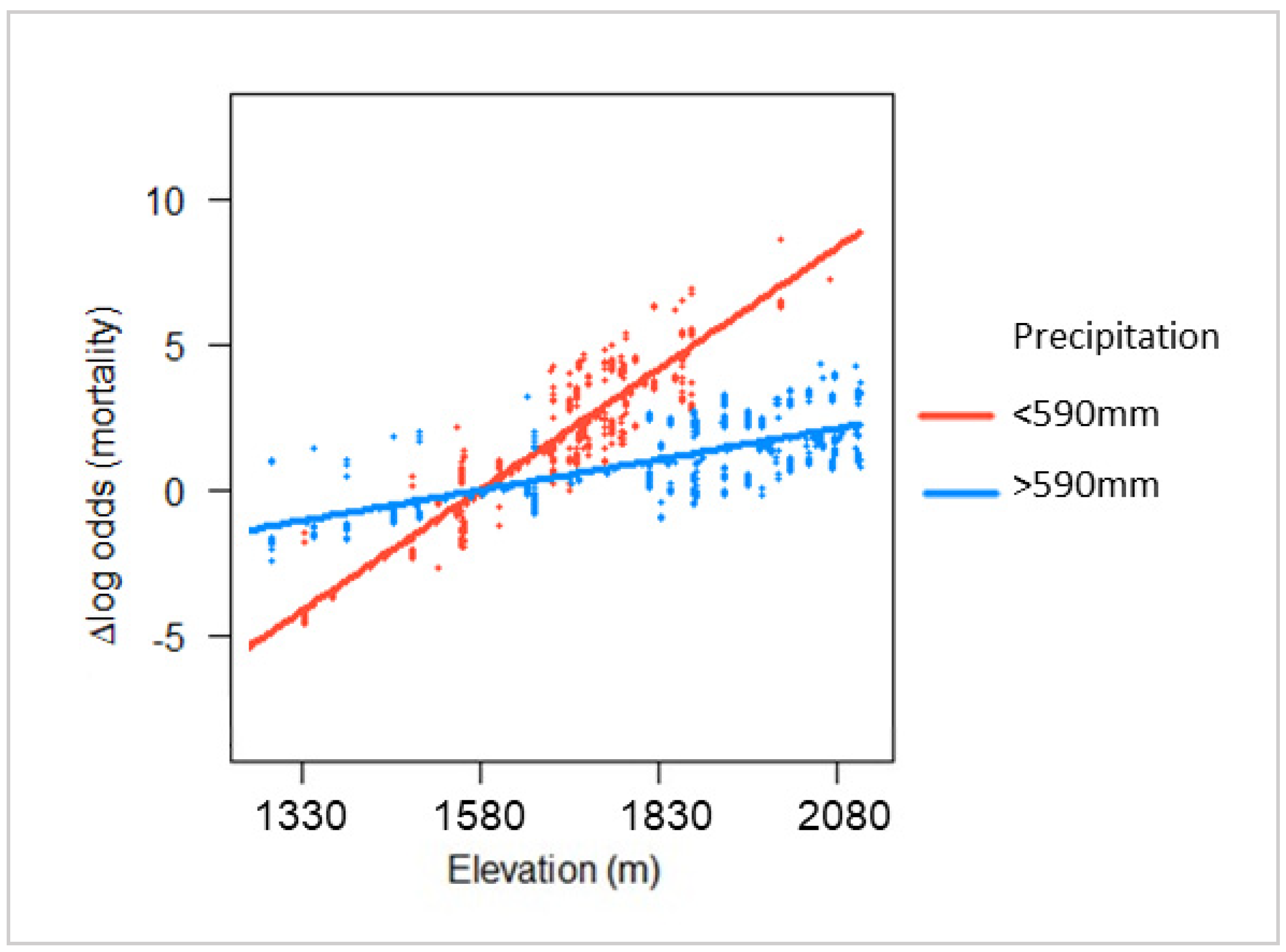
Although observed mortality rates were greatest at high elevation, this effect was buffered by a higher average annual precipitation. As elevation increased, mortality risk increased faster in dry sites than in moist sites (
Figure 4 and
Figure 5). This finding demonstrates that climate may influence the relationship between stand properties and mortality risk during a bark beetle outbreak. Stand hazard ratings have been developed for a multitude of pest–host–region combinations [
12,
13,
15,
50,
54], but few models incorporate local climatic variation into their ratings. Our results suggest that such variations may be important in understanding tree susceptibility.
Despite these potential implications, it remains that most of the significant stand- and tree-level variables were not influenced by climate. This indicates that while the inclusion of climate variables could potentially improve current stand hazard ratings, the characteristics of an individual tree or stand are likely to remain the most important determinants for mortality risk. However, the relative importance of stand vs. climate variables (and their interactions) is likely to vary according to geographic scale. The Black Hills represents a relatively small land area, and our plots were separated by no more than 140 km (
Figure 1). If the study area were broadened to include a greater range of climatic variation, climate variables could play a more important role in modulating mortality risk.
Results from this study could help determine which stands are at the highest risk of mortality during a future bark beetle outbreak. While high-value trees, such as those located in campgrounds or tourist areas, can be protected using insecticides or pheromones [
55,
56], these treatments are typically expensive, and managers must prioritize which areas to treat. Our results show that within a stand, large trees with sparse crowns were most susceptible. Meanwhile, across the landscape, stands that were dry, dense, and at high elevation were most susceptible to insect-related mortality. Thus, treatments to reduce tree density may be most effective in relatively dry stands above 1700 m. Additionally, outbreak mitigation may be most effective in areas with no recent outbreaks, as many susceptible mature trees in post-outbreak stands have presumably been selected out of the population [
57].
The scope of this study is limited by several factors. First, our observation window covers only a portion of the latest mountain pine beetle outbreak in the Black Hills. FIA plots in the Black Hills are currently transitioning from a five- to a seven-year resampling period. This means that some plots measured in 2013–2015 will not be remeasured again until 2020–2022, and are not represented in our dataset. The outbreak subsided around 2016, so additional mortality is likely to be observed as plots continue to be remeasured. Second, this is an observational study with many uncontrolled or unobserved variables potentially influencing results. While we report correlations between observed variables, it is difficult to draw causal linkages. Third, our study relies on modelled climate averages for each plot [
38], which may have varying levels of accuracy. Additionally, climate averages for each plot were provided by the Forest Service, which fuzzed the value of a subset of plots to within 20% of the actual value in order to preserve plot location confidentiality. Due to this fuzzing, the strength of correlations between climate variables and mortality risk could be underestimated.
Our study highlights the importance of examining both tree- and stand-level variables that could contribute to bark beetle-related mortality. The results from our analysis can help managers identify potentially high-risk stands if another bark beetle outbreak were to occur in the Black Hills. Additionally, our results demonstrate that correlations between stand characteristics and mortality hazard can be strongly mediated by climatic variables. The role that average annual precipitation played in our analysis underscores the potential for bark beetle-driven patterns of tree mortality to be strongly affected by alterations in regional climate, and it will be important to further determine whether these effects were driven by summer or winter precipitation inputs. As precipitation patterns across the western U.S. shift due to climate change, longstanding correlations between stand characteristics and bark beetle susceptibility may change as well.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}




