
Rock-Solubilizing Microbial Inoculums Have Enormous Potential as Ecological Remediation Agents to Promote Plant Growth

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Seed Test Material, Microbial Strains, and Soil
2.3. Pot Experiment Setup
2.4. Plant Root Nodule Parameters
2.5. Analysis of Soil Properties
2.6. Data Analysis
3. Results
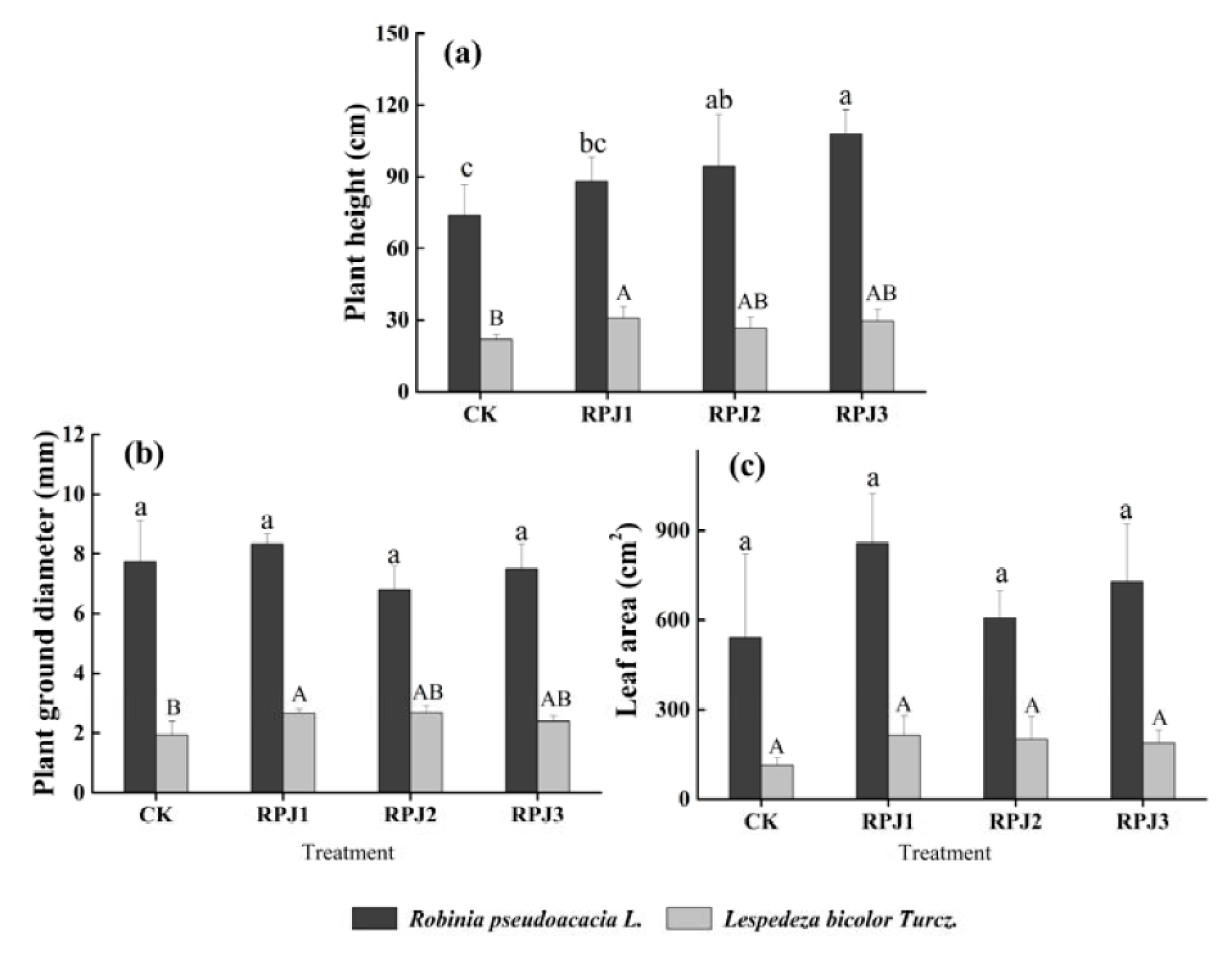
3.1. Plant Growth Influenced by the Supply of Rock-Solubilizing Microbes
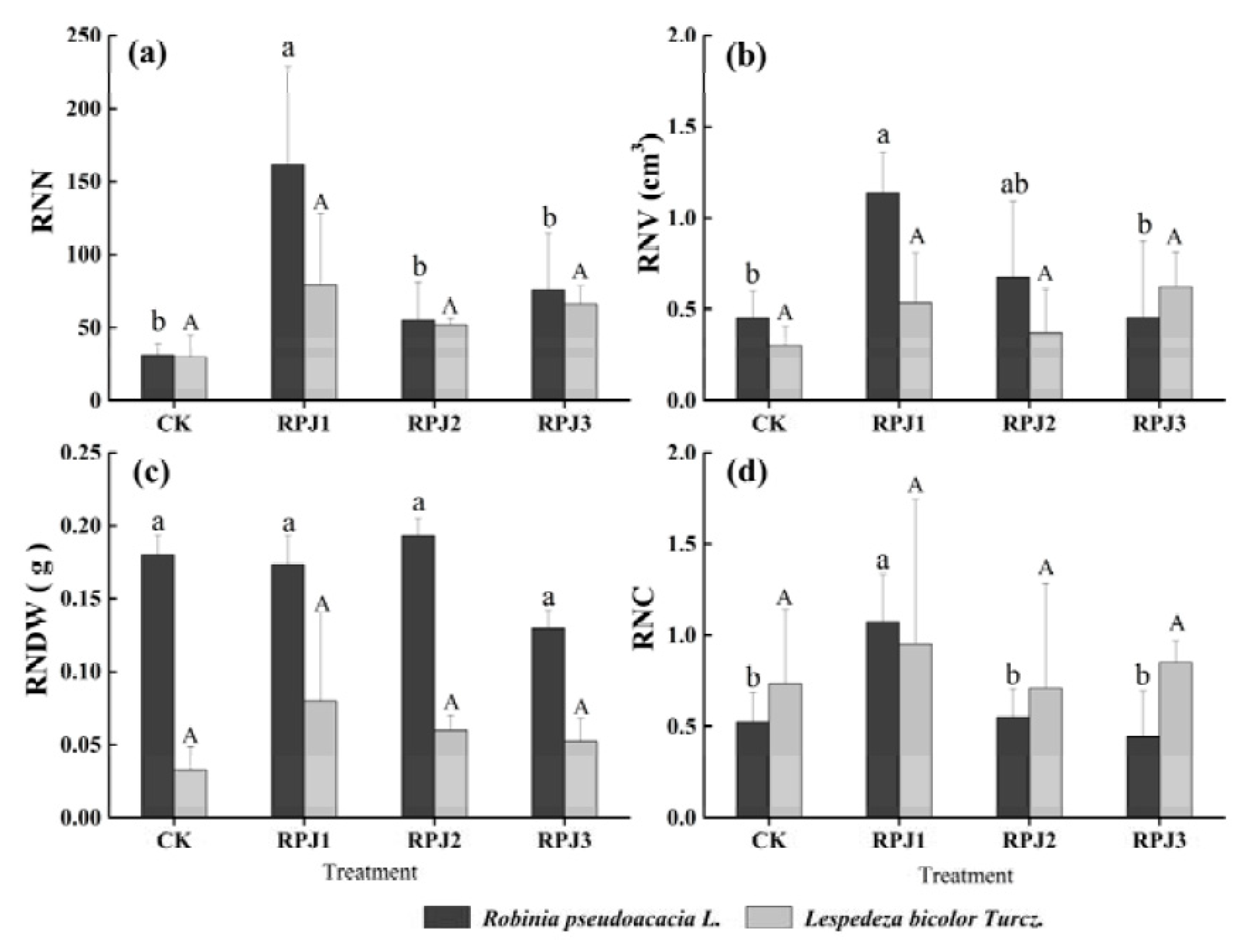
3.2. Nodule Formation under the Addition of Rock-Solubilizing Microbes
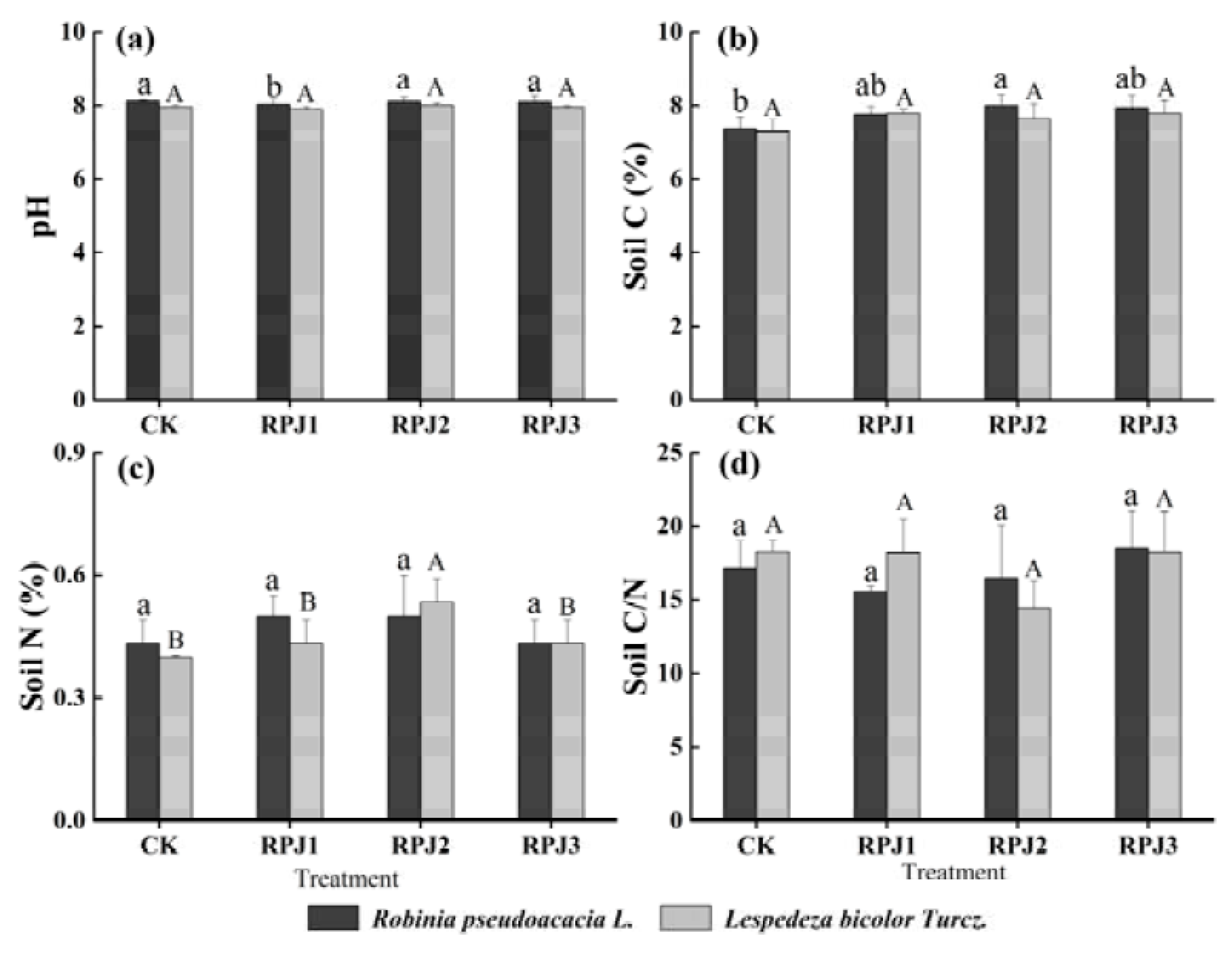
3.3. Effects of Rock-Solubilizing Microbes on Soil Properties
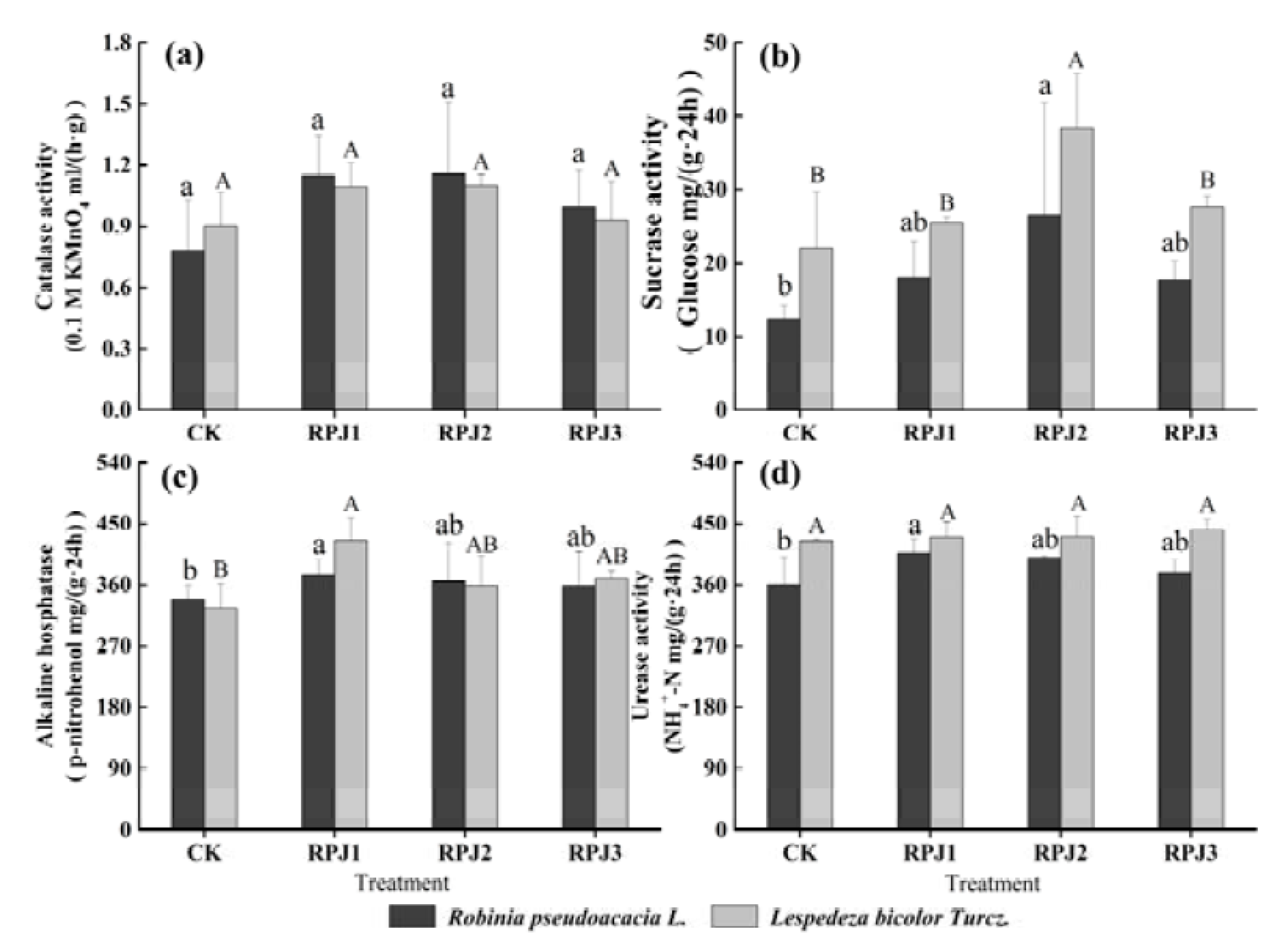
3.4. Effects of Rock-Solubilizing Microbes on Soil Enzymes
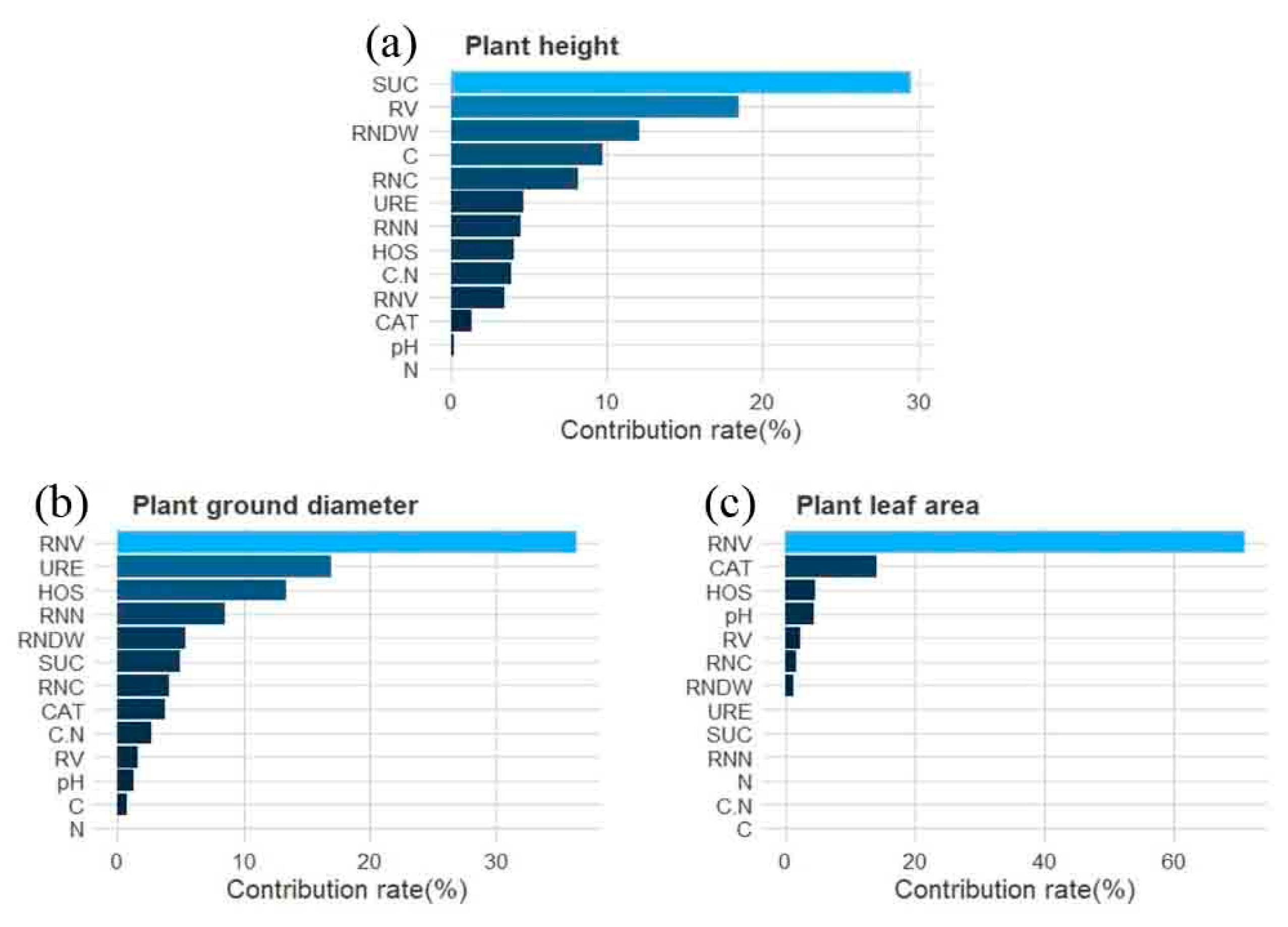
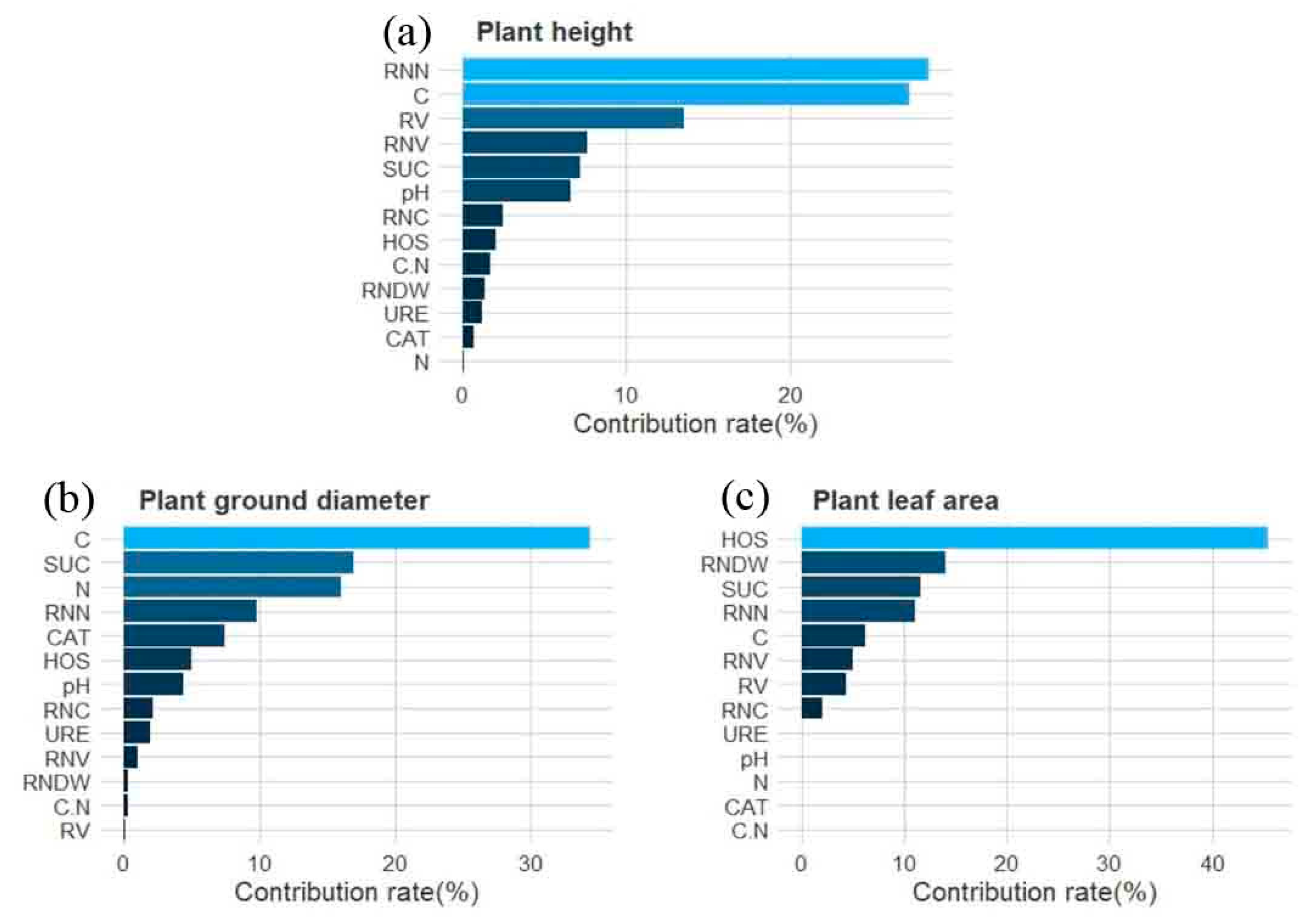
3.5. Boosted Regression Tree (BRT) Model Analysis
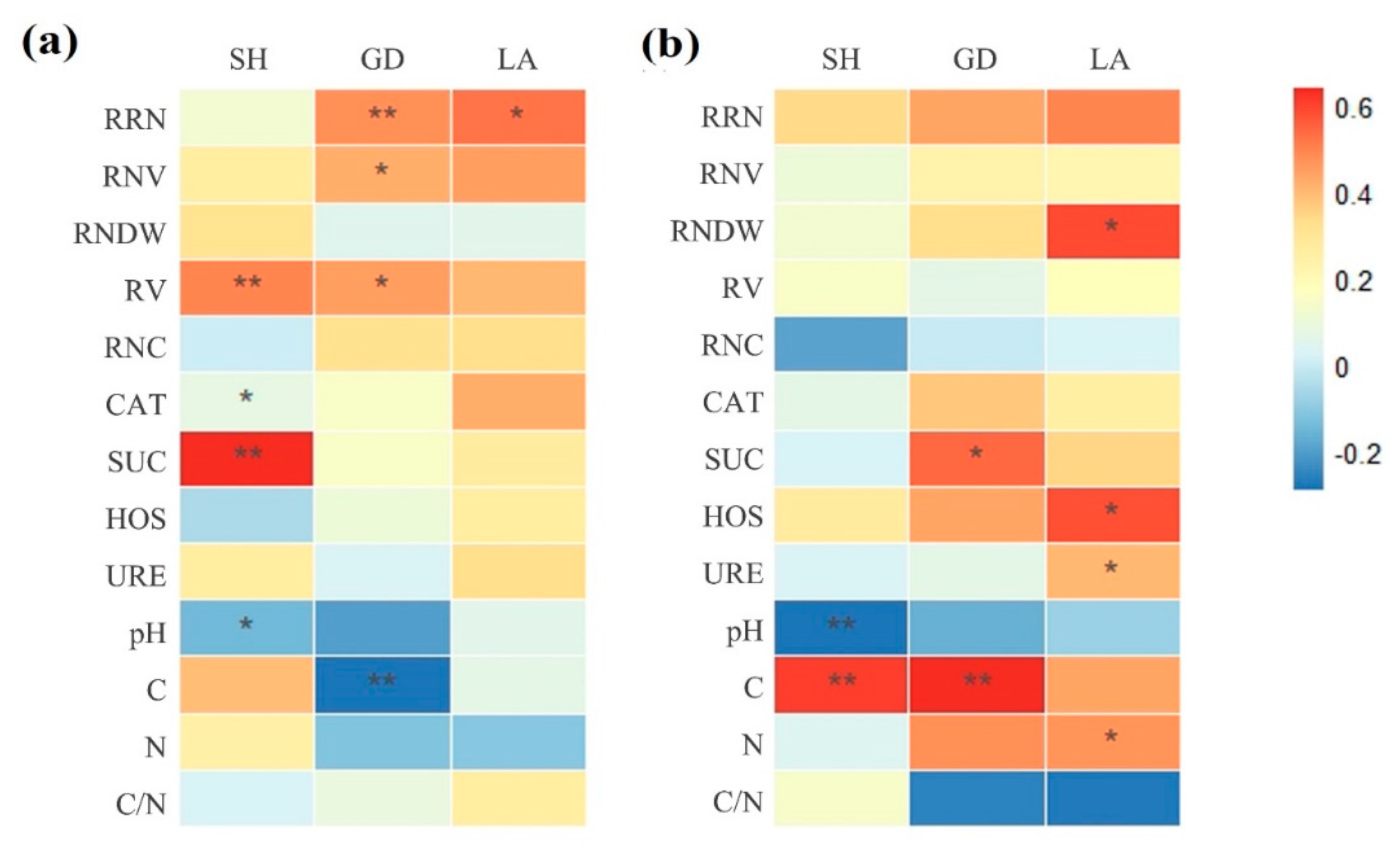
3.6. Correlation Coefficients between Plant Growth and Different Parameters
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vangsnes, G.F. The meanings of mining: A perspective on the regulation of artisanal and small-scale gold mining in southern Ecuador. Extr. Ind. Soc. 2018, 5, 317–326. [Google Scholar] [CrossRef]
- Lemly, A.D. Environmental hazard assessment of Benga Mining’s proposed Grassy Mountain Coal Project. Environ. Sci. Policy 2019, 96, 105–113. [Google Scholar] [CrossRef]
- Lv, Y.; Jiang, Y.; Hu, W.; Cao, M.; Mao, Y. A review of the effects of tunnel excavation on the hydrology, ecology, and environment in karst areas: Current status, challenges, and perspectives. J. Hydrol. 2020, 586, 124891. [Google Scholar] [CrossRef]
- Yang, Z.; Li, W.; Li, X.; Wang, Q.; He, J. Assessment of eco-geo-environment quality using multivariate data: A case study in a coal mining area of Western China. Ecol. Indic. 2019, 107, 105651. [Google Scholar] [CrossRef]
- Yang, Z.; Yang, L.; Zhang, B. Soil erosion and its basic characteristics at karst rocky-desertified land consolidation area: A case study at Muzhe Village of Xichou County in Southeast Yunnan, China. J. Mt. Sci. 2010, 7, 55–72. [Google Scholar] [CrossRef]
- Martins, W.B.R.; Lima, M.D.R.; Junior, U.D.O.B.; Amorim, L.S.V.-B.; Oliveira, F.D.A.; Schwartz, G. Ecological methods and indicators for recovering and monitoring ecosystems after mining: A global literature review. Ecol. Eng. 2020, 145, 105707. [Google Scholar] [CrossRef]
- Hou, X.; Liu, S.; Zhao, S.; Zhang, Y.; Wu, X.; Cheng, F.; Dong, S. Interaction mechanism between floristic quality and environmental factors during ecological restoration in a mine area based on structural equation modeling. Ecol. Eng. 2018, 124, 23–30. [Google Scholar] [CrossRef]
- Sapkota, R.; Stahl, P.; Rijal, K. Restoration governance: An integrated approach towards sustainably restoring degraded ecosystems. Environ. Dev. 2018, 27, 83–94. [Google Scholar] [CrossRef]
- Li, S.; Li, Y.; Shi, J.; Zhao, T.; Yang, J. Optimizing the formulation of external-soil spray seeding with sludge using the orthogonal test method for slope ecological protection. Ecol. Eng. 2017, 102, 527–535. [Google Scholar] [CrossRef]
- Feng, Y.; Wang, J.; Bai, Z.; Reading, L. Effects of surface coal mining and land reclamation on soil properties: A review. Earth-Sci. Rev. 2019, 191, 12–25. [Google Scholar] [CrossRef]
- Li, M. Ecological restoration of mineland with particular reference to the metalliferous mine wasteland in China: A review of research and practice. Sci. Total Environ. 2006, 357, 38–53. [Google Scholar] [CrossRef] [PubMed]
- Maiti, S.K.; Ahirwal, J. Chapter 3—Ecological Restoration of Coal Mine Degraded Lands: Topsoil Management, Pedogenesis, Carbon Sequestration, and Mine Pit Limnology. In Phytomanagement of Polluted Sites; Elsevier: Kidlington, Oxford, UK, 2019; pp. 83–111. [Google Scholar] [CrossRef]
- Guimarães, B.C.; Arends, J.B.; van der Ha, D.; Van de Wiele, T.; Boon, N.; Verstraete, W. Microbial services and their management: Recent progresses in soil bioremediation technology. Appl. Soil Ecol. 2010, 46, 157–167. [Google Scholar] [CrossRef] [Green Version]
- Thavamani, P.; Samkumar, R.A.; Satheesh, V.; Subashchandrabose, S.R.; Ramadass, K.; Naidu, R.; Venkateswarlu, K.; Megharaj, M. Microbes from mined sites: Harnessing their potential for reclamation of derelict mine sites. Environ. Pollut. 2017, 230, 495–505. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Zhang, J.; Guo, X. An Indigenous Soil Bacterium Facilitates the Mitigation of Rocky Desertification in Carbonate Mining Areas. Land Degrad. Dev. 2017, 28, 2222–2233. [Google Scholar] [CrossRef]
- Bruneel, O.; Mghazli, N.; Sbabou, L.; Héry, M.; Casiot, C.; Filali-Maltouf, A. Role of microorganisms in rehabilitation of mining sites, focus on Sub Saharan African countries. J. Geochem. Explor. 2019, 205, 106327. [Google Scholar] [CrossRef]
- Wu, Y.-W.; Zhang, J.-C.; Wang, L.-J.; Wang, Y.-X. A rock-weathering bacterium isolated from rock surface and its role in ecological restoration on exposed carbonate rocks. Ecol. Eng. 2017, 101, 162–169. [Google Scholar] [CrossRef]
- Jacob, J.M.; Karthik, C.; Saratale, R.G.; Kumar, S.S.; Prabakar, D.; Kadirvelu, K.; Pugazhendhi, A. Biological approaches to tackle heavy metal pollution: A survey of literature. J. Environ. Manag. 2018, 217, 56–70. [Google Scholar] [CrossRef] [PubMed]
- Rashid, M.I.; Mujawar, L.H.; Shahzad, T.; Almeelbi, T.; Ismail, I.M.; Oves, M. Bacteria and fungi can contribute to nutrients bioavailability and aggregate formation in degraded soils. Microbiol. Res. 2016, 183, 26–41. [Google Scholar] [CrossRef] [PubMed]
- Carabassa, V.; Ortiz, O.; Alcañiz, J.M. RESTOQUARRY: Indicators for self-evaluation of ecological restoration in open-pit mines. Ecol. Indic. 2019, 102, 437–445. [Google Scholar] [CrossRef]
- Li, C.; Jia, Z.; Yuan, Y.; Cheng, X.; Shi, J.; Tang, X.; Wang, Y.; Peng, X.; Dong, Y.; Ma, S.; et al. Effects of mineral-solubilizing microbial strains on the mechanical responses of roots and root-reinforced soil in external-soil spray seeding substrate. Sci. Total Environ. 2020, 723, 138079. [Google Scholar] [CrossRef] [PubMed]
- Xue, S.; Yang, X.; Liu, G.; Gai, L.; Zhang, C.; Ritsema, C.J.; Geissen, V. Effects of elevated CO2 and drought on the microbial biomass and enzymatic activities in the rhizospheres of two grass species in Chinese loess soil. Geoderma 2017, 286, 25–34. [Google Scholar] [CrossRef]
- Mehrabian, H.; Myrehaug, S.; Soliman, H.; Sahgal, A.; Stanisz, G.J. Quantitative Magnetization Transfer in Monitoring Glioblastoma (GBM) Response to Therapy. Sci. Rep. 2018, 8, 2475. [Google Scholar] [CrossRef] [Green Version]
- Wickham, H. ggplot2. Wiley Interdiscip. Rev. Comput. Stat. 2011, 3, 180–185. [Google Scholar] [CrossRef]
- Singh, B.; Boukhris, I.; Kumar, V.; Yadav, A.N.; Farhat-Khemakhem, A.; Kumar, A.; Singh, D.; Blibech, M.; Chouayekh, H.; Alghamdi, O.A. Contribution of microbial phytases to the improvement of plant growth and nutrition: A review. Pedosphere 2020, 30, 295–313. [Google Scholar] [CrossRef]
- Ortega, R.; Domene, M.; Soriano, M.; Sánchez-Marañón, M.; Asensio, C.; Miralles, I. Improving the fertility of degraded soils from a limestone quarry with organic and inorganic amendments to support vegetation restoration with semiarid Mediterranean plants. Soil Tillage Res. 2020, 204, 104718. [Google Scholar] [CrossRef]
- Legay, N.; Clément, J.; Grassein, F.; Lavorel, S.; Lemauviel-Lavenant, S.; Personeni, E.; Poly, F.; Pommier, T.; Robson, T.; Mouhamadou, B.; et al. Plant growth drives soil nitrogen cycling and N-related microbial activity through changing root traits. Fungal Ecol. 2020, 44, 100910. [Google Scholar] [CrossRef]
- Chaudhary, D.R.; Rathore, A.P.; Sharma, S. Effect of halotolerant plant growth promoting rhizobacteria inoculation on soil microbial community structure and nutrients. Appl. Soil Ecol. 2020, 150, 103461. [Google Scholar] [CrossRef]
- Kong, Z.; Wu, Z.; Glick, B.R.; He, S.; Huang, C.; Wu, L. Co-occurrence patterns of microbial communities affected by inoculants of plant growth-promoting bacteria during phytoremediation of heavy metal-contaminated soils. Ecotoxicol. Environ. Saf. 2019, 183, 109504. [Google Scholar] [CrossRef]
- Larsen, J.; Pineda-Sánchez, H.; Delgado-Arellano, I.; Castellano-Morales, V.; Carreto-Montoya, L.; Villegas-Moreno, J. Interactions between microbial plant growth promoters and their effects on maize growth performance in different mineral and organic fertilization scenarios. Rhizosphere 2017, 3, 75–81. [Google Scholar] [CrossRef]
- Raheem, A.; Sajid, M.; Iqbal, M.S.; Aslam, H.; Bilal, M.; Rafiq, F. Microbial inhabitants of agricultural land have potential to promote plant growth but they are liable to traditional practice of wheat (T. aestivum L.) straw burning. Biocatal. Agric. Biotechnol. 2019, 18, 101060. [Google Scholar] [CrossRef]
- Di Salvo, L.P.; Cellucci, G.C.; Carlino, M.E.; de Salamone, I.E.G. Plant growth-promoting rhizobacteria inoculation and nitrogen fertilization increase maize (Zea mays L.) grain yield and modified rhizosphere microbial communities. Appl. Soil Ecol. 2018, 126, 113–120. [Google Scholar] [CrossRef]
- Wood, J.L.; Tang, C.; Franks, A.E. Microbial associated plant growth and heavy metal accumulation to improve phytoextraction of contaminated soils. Soil Biol. Biochem. 2016, 103, 131–137. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Liang, C.; Wang, Y.; Cheng, H.; An, S.; Chang, S.X. Soil extracellular enzyme stoichiometry reflects the shift from P- to N-limitation of microorganisms with grassland restoration. Soil Biol. Biochem. 2020, 149, 107928. [Google Scholar] [CrossRef]
- Tripathi, P.; Na, C.-I.; Kim, Y. Effect of silicon fertilizer treatment on nodule formation and yield in soybean (Glycine max L.). Eur. J. Agron. 2021, 122, 126172. [Google Scholar] [CrossRef]
- Iturralde, E.T.; Stocco, M.C.; Faura, A.; Mónaco, C.I.; Cordo, C.; Pérez-Giménez, J.; Lodeiro, A.R. Coinoculation of soybean plants with Bradyrhizobium japonicum and Trichoderma harzianum: Coexistence of both microbes and relief of nitrate inhibition of nodulation. Biotechnol. Rep. 2020, 26, e00461. [Google Scholar] [CrossRef] [PubMed]
- Etesami, H.; Adl, S.M. Can interaction between silicon and non–rhizobial bacteria help in improving nodulation and nitrogen fixation in salinity–stressed legumes? A review. Rhizosphere 2020, 15, 100229. [Google Scholar] [CrossRef]
- Ferreira, L.D.V.M.; De Carvalho, F.; Andrade, J.F.C.; Oliveira, D.P.; De Medeiros, F.H.V.; Moreira, F.M.D.S. Co-inoculation of selected nodule endophytic rhizobacterial strains with Rhizobium tropici promotes plant growth and controls damping off in common bean. Pedosphere 2020, 30, 98–108. [Google Scholar] [CrossRef]
- Cardoso, P.; Alves, A.; Silveira, P.; Sá, C.; Fidalgo, C.; Freitas, R.; Figueira, E. Bacteria from nodules of wild legume species: Phylogenetic diversity, plant growth promotion abilities and osmotolerance. Sci. Total Environ. 2018, 645, 1094–1102. [Google Scholar] [CrossRef]
- Nascimento, F.X.; Tavares, M.J.; Rossi, M.J.; Glick, B.R. The modulation of leguminous plant ethylene levels by symbiotic rhizobia played a role in the evolution of the nodulation process. Heliyon 2018, 4, e01068. [Google Scholar] [CrossRef] [Green Version]
- Güereña, D.T.; Lehmann, J.; Thies, J.E.; Vanek, S.; Karanja, N.; Neufeldt, H. Nodulation of beans with inoculant carriers from pyrolyzed and non-pyrolyzed sugarcane bagasse in response to different pre-planting water availability. Appl. Soil Ecol. 2019, 143, 126–133. [Google Scholar] [CrossRef]
- Lehmann, J.; Rillig, M.C.; Thies, J.; Masiello, C.A.; Hockaday, W.C.; Crowley, D. Biochar effects on soil biota—A review. Soil Biol. Biochem. 2011, 43, 1812–1836. [Google Scholar] [CrossRef]
- Naveed, M.; Duan, J.; Uddin, S.; Suleman, M.; Hui, Y.; Li, H. Application of microbially induced calcium carbonate precipitation with urea hydrolysis to improve the mechanical properties of soil. Ecol. Eng. 2020, 153, 105885. [Google Scholar] [CrossRef]
- Xiao, X.-Y.; Wang, M.-W.; Zhu, H.-W.; Guo, Z.-H.; Han, X.-Q.; Zeng, P. Response of soil microbial activities and microbial community structure to vanadium stress. Ecotoxicol. Environ. Saf. 2017, 142, 200–206. [Google Scholar] [CrossRef] [PubMed]
- Tosi, M.; Brown, S.; Machado, P.V.F.; Wagner-Riddle, C.; Dunfield, K. Short-term response of soil N-cycling genes and transcripts to fertilization with nitrification and urease inhibitors, and relationship with field-scale N2O emissions. Soil Biol. Biochem. 2020, 142, 107703. [Google Scholar] [CrossRef]
- Pietrzykowski, M. Tree species selection and reaction to mine soil reconstructed at reforested post-mine sites: Central and eastern European experiences. Ecol. Eng. X 2019, 3, 100012. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jia, Z.; Meng, M.; Li, C.; Zhang, B.; Zhai, L.; Liu, X.; Ma, S.; Cheng, X.; Zhang, J. Rock-Solubilizing Microbial Inoculums Have Enormous Potential as Ecological Remediation Agents to Promote Plant Growth. Forests 2021, 12, 357. https://doi.org/10.3390/f12030357
Jia Z, Meng M, Li C, Zhang B, Zhai L, Liu X, Ma S, Cheng X, Zhang J. Rock-Solubilizing Microbial Inoculums Have Enormous Potential as Ecological Remediation Agents to Promote Plant Growth. Forests. 2021; 12(3):357. https://doi.org/10.3390/f12030357
Chicago/Turabian StyleJia, Zhaohui, Miaojing Meng, Chong Li, Bo Zhang, Lu Zhai, Xin Liu, Shilin Ma, Xuefei Cheng, and Jinchi Zhang. 2021. "Rock-Solubilizing Microbial Inoculums Have Enormous Potential as Ecological Remediation Agents to Promote Plant Growth" Forests 12, no. 3: 357. https://doi.org/10.3390/f12030357