
First-Decade Biomass and Carbon Accumulation, and Woody Community Change after Severe Wind Damage in a Hemlock-White Pine Forest Remnant


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site and Disturbance Event
2.2. Field and Statistical Methods
3. Results
3.1. Pre-Disturbance Canopy Structure and Composition
3.2. Structural Damage, Mortality, and Sprouting
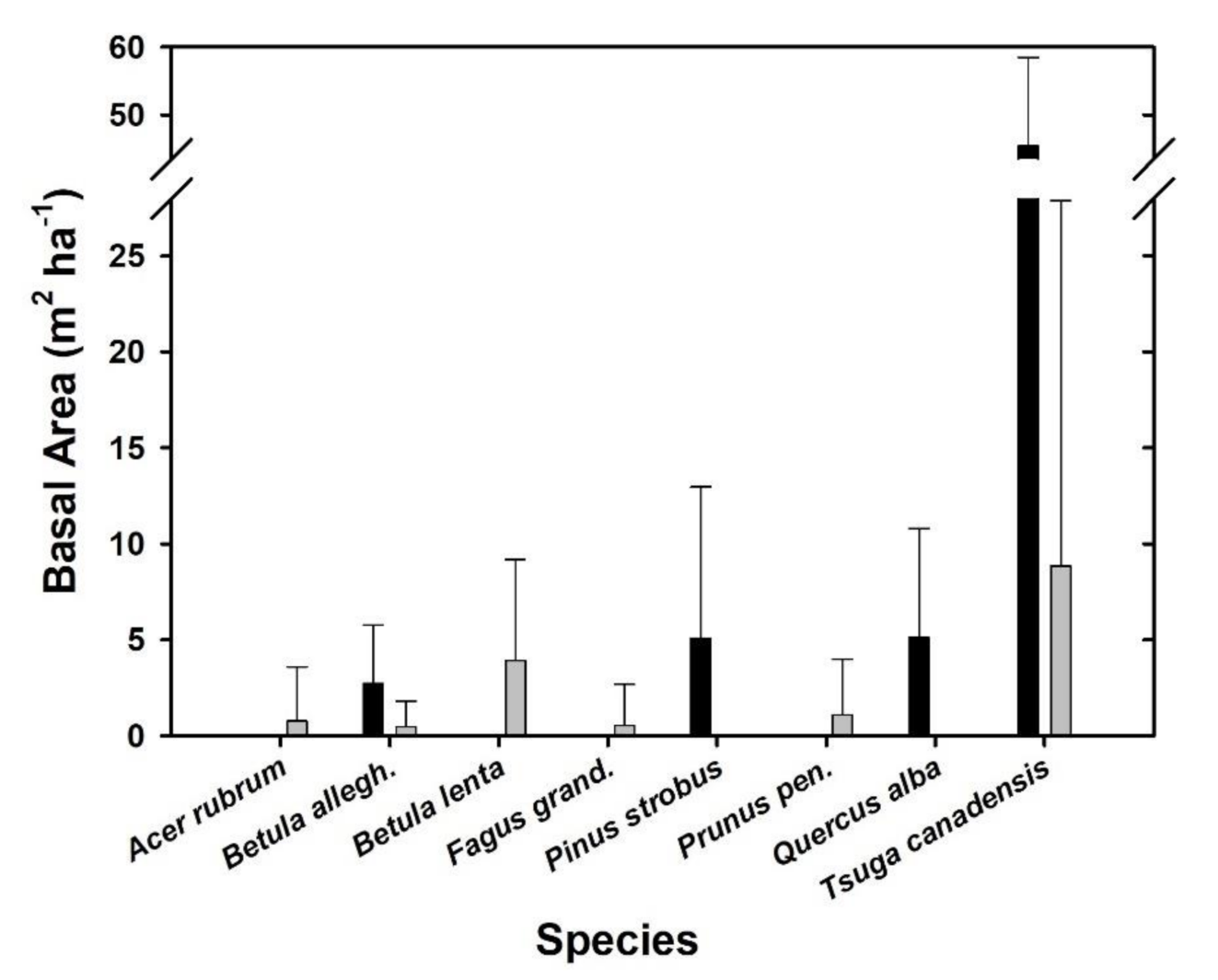
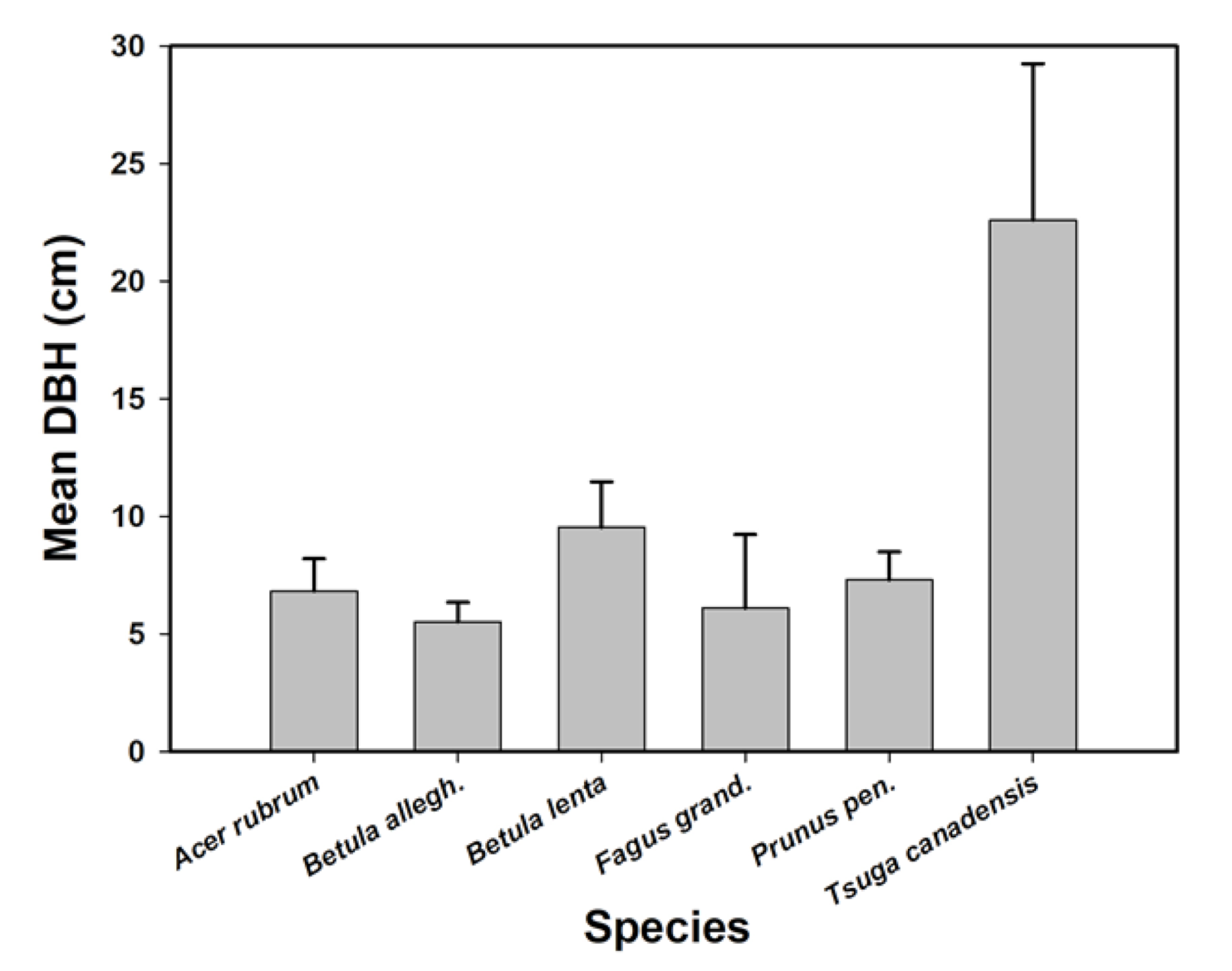
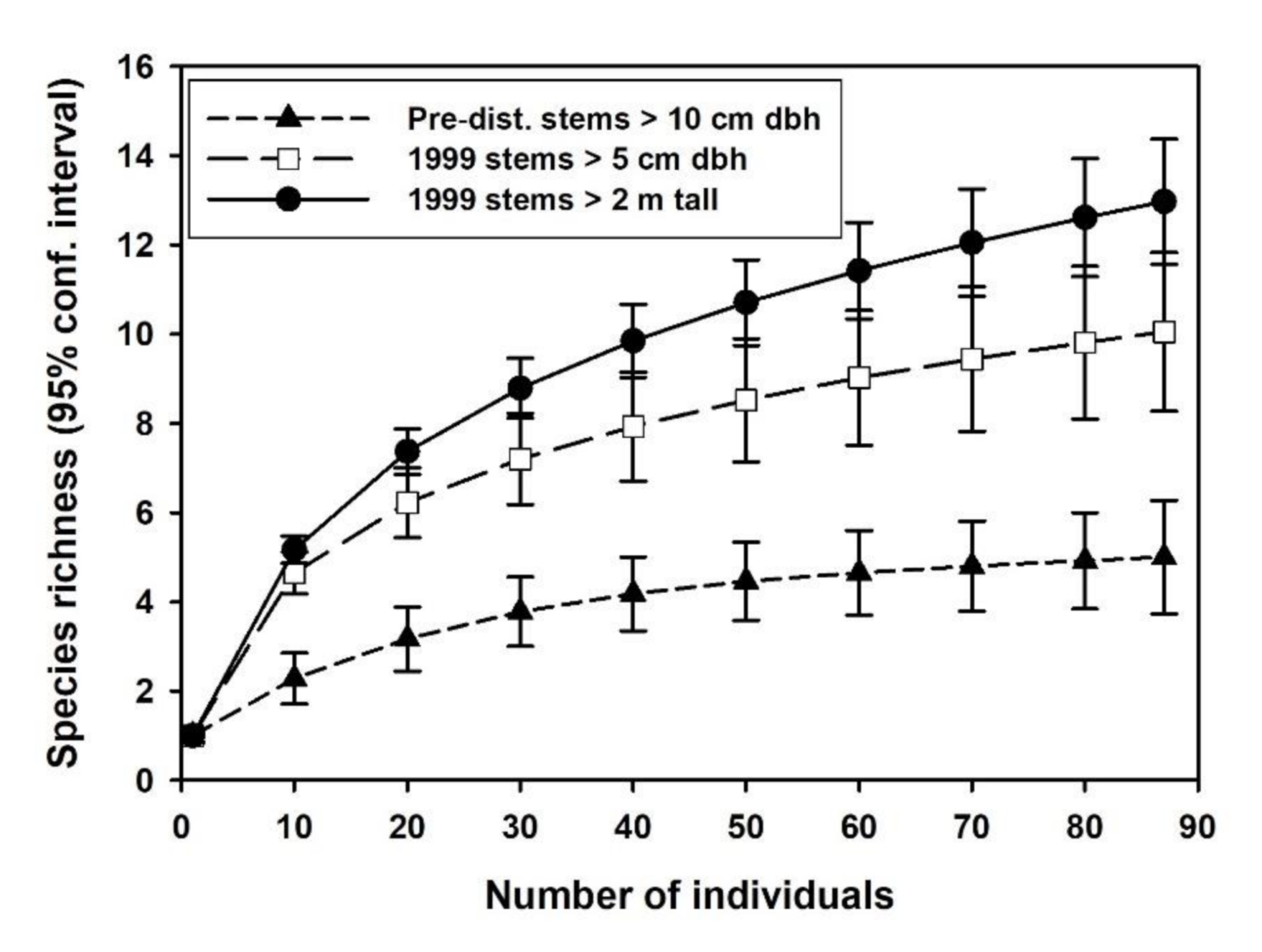
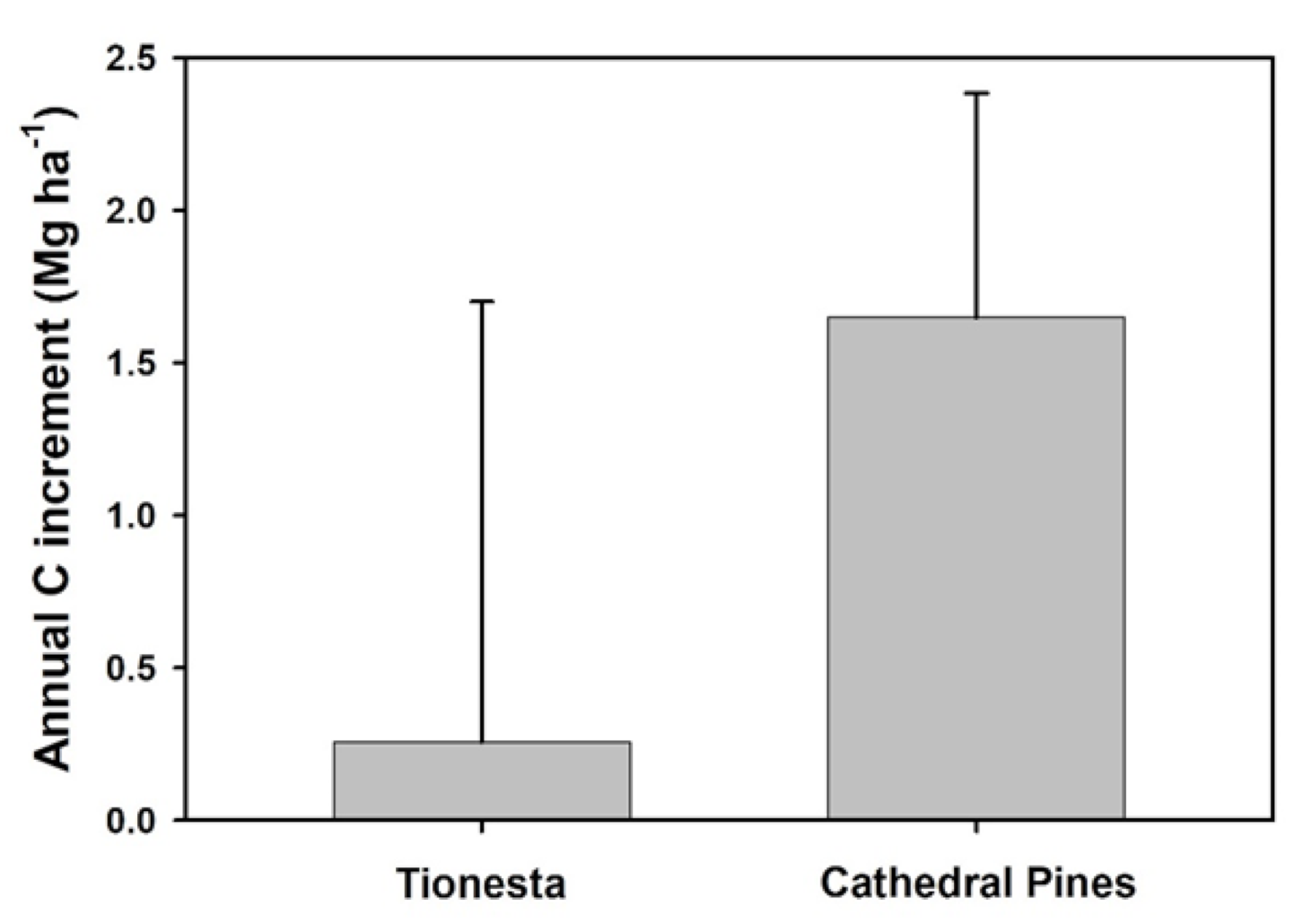
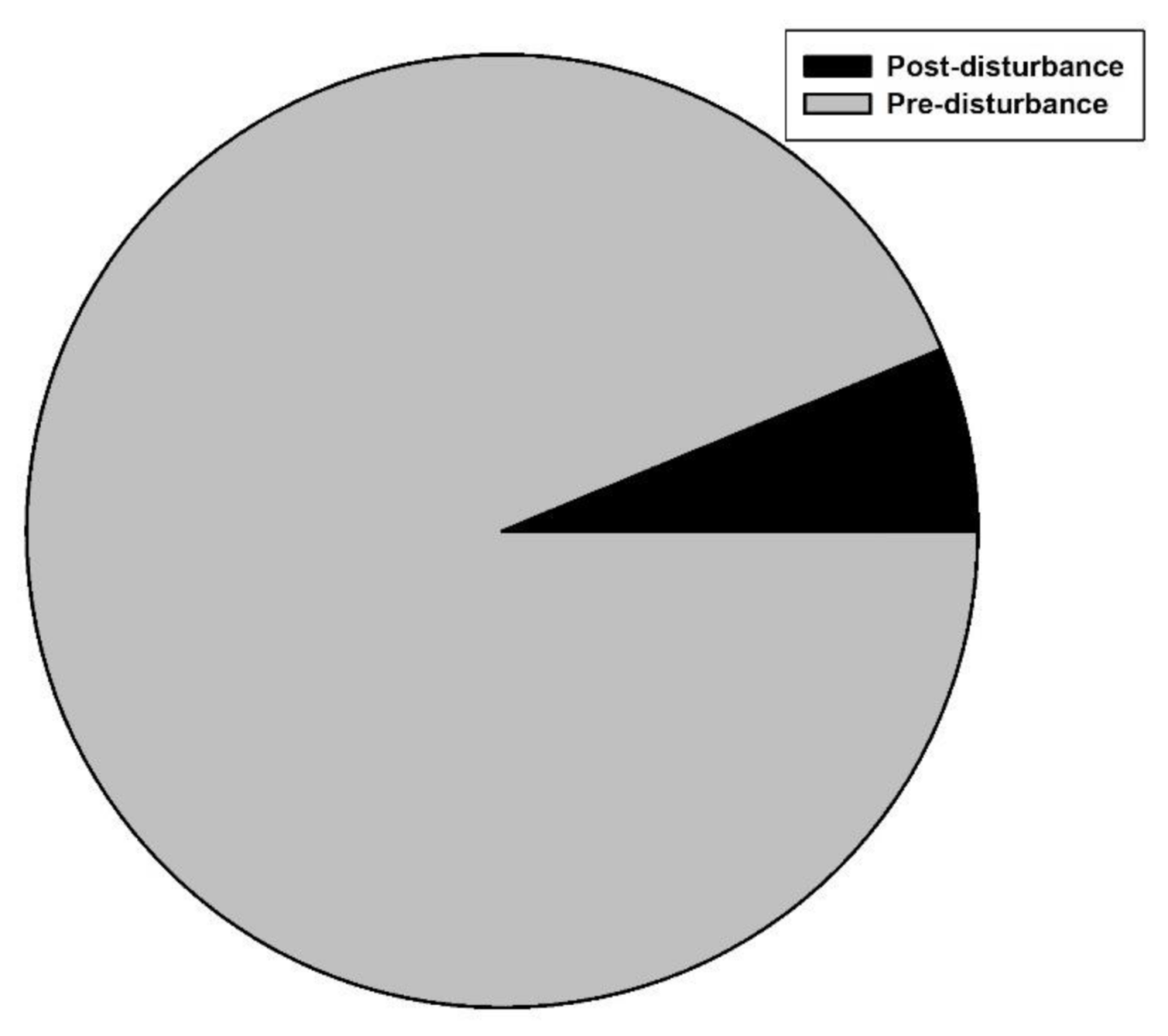
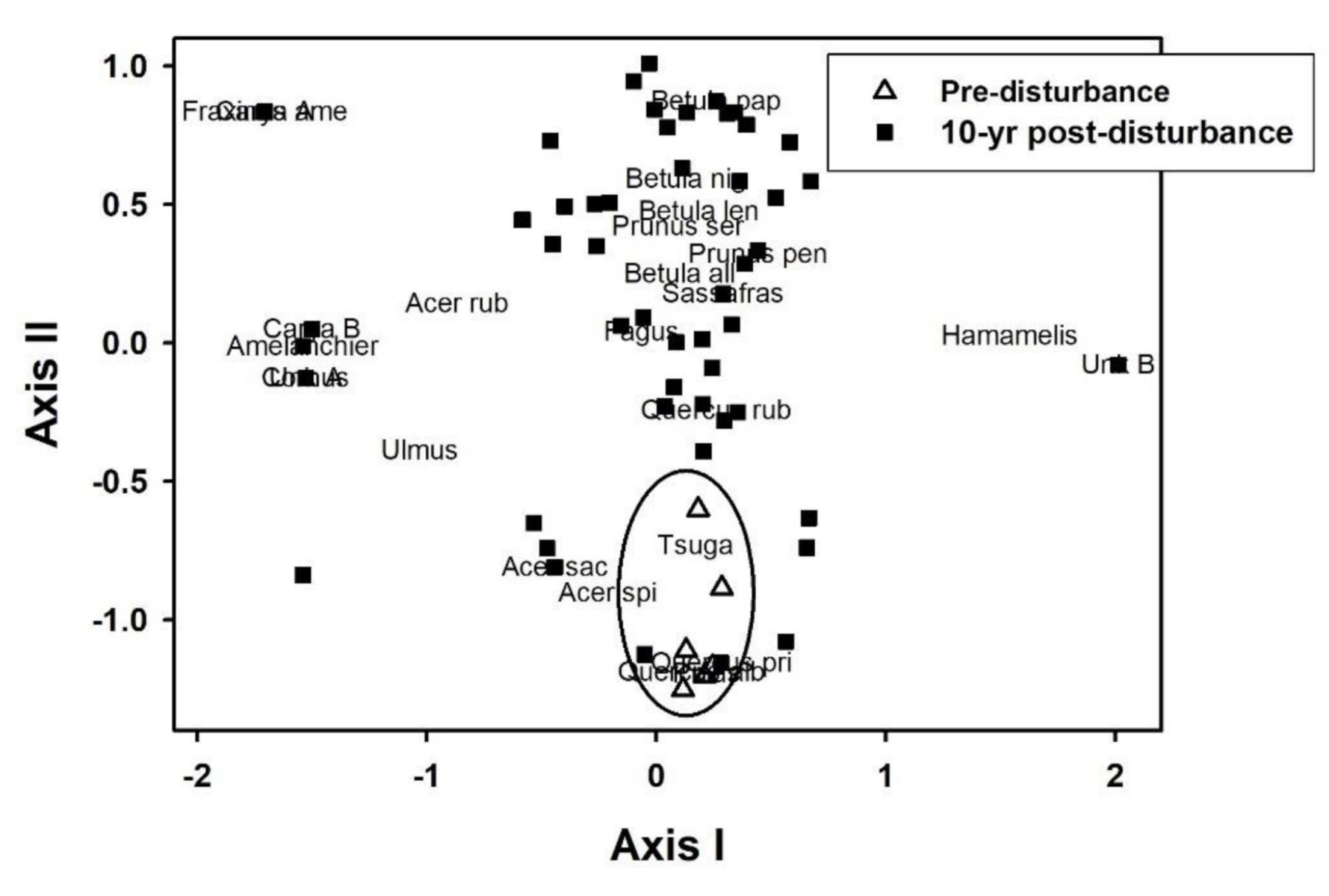
3.3. Revegetation
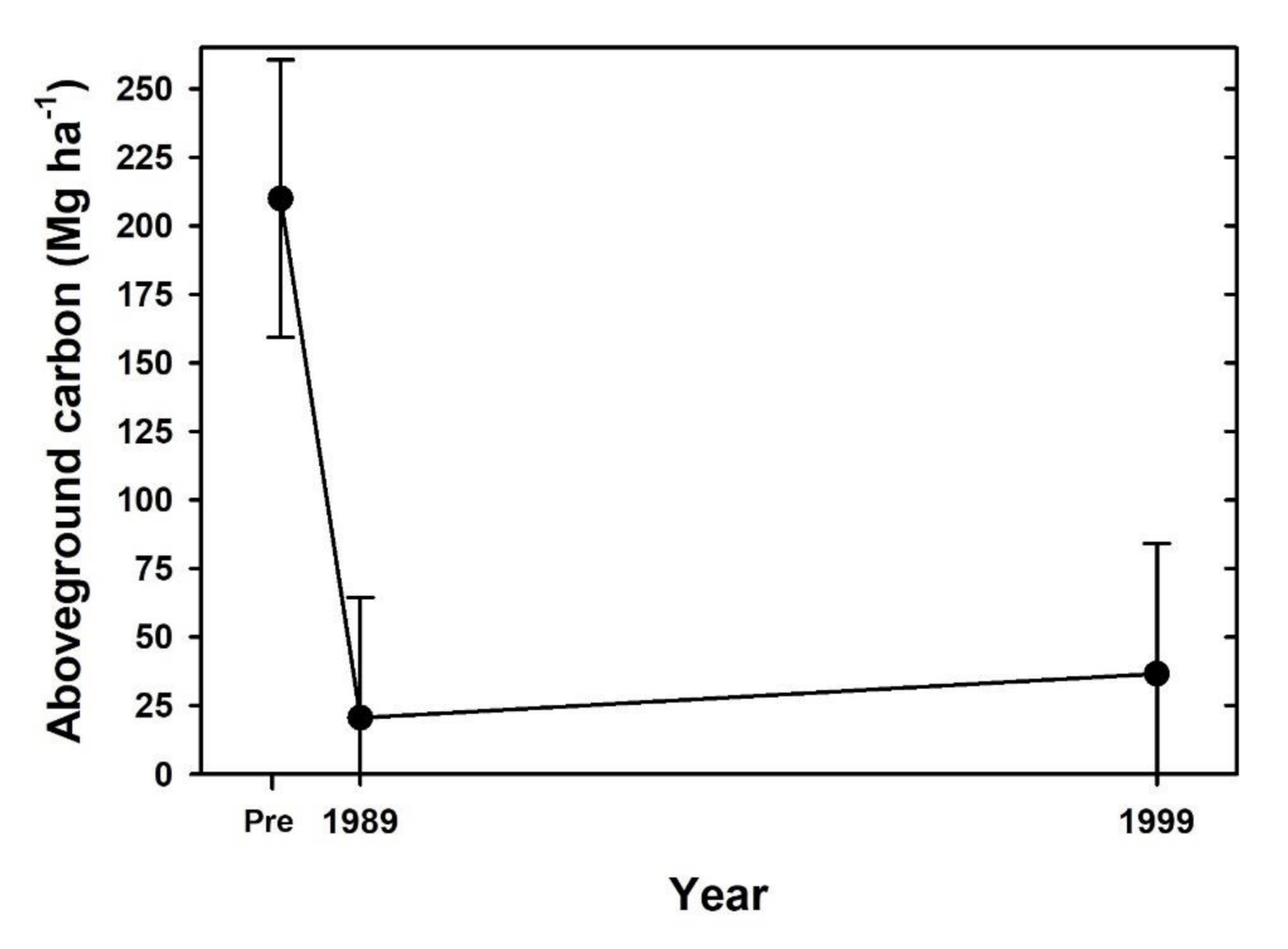
4. Discussion
5. Conclusions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bradford, J.B.; Fraver, S.; Milo, A.M.; D’Amato, A.W.; Palik, B.; Shinneman, D.J. Effects of multiple interacting disturbances and salvage logging on forest carbon stocks. For. Ecol. Manag. 2012, 267, 209–214. [Google Scholar] [CrossRef]
- Kasischke, E.S.; Amiro, B.D.; Barger, N.N.; French, N.H.F.; Goetz, S.J.; Grosse, G.; Harmon, M.E.; Hicke, J.A.; Liu, S.; Masek, J.G. Impacts of disturbance on the terrestrial carbon budget of North America. J. Geophys. Res. Biogeosciences 2013, 118, 303–316. [Google Scholar] [CrossRef]
- Williams, C.A.; Gu, H.; MacLean, R.; Masek, J.G.; Collatz, G.J. Disturbance and the carbon balance of US forests: A quantitative review of impacts from harvests, fires, insects, and droughts. Glob. Planet. Chang. 2016, 143, 66–80. [Google Scholar] [CrossRef]
- Ghimire, B.; Williams, C.A.; Collatz, G.J.; Vanderhoof, M. Fire-induced carbon emissions and regrowth uptake in western U.S. forests: Documenting variation across forest types, fire severity, and climate regions. J. Geophys. Res. 2012, 117, G03036. [Google Scholar] [CrossRef] [Green Version]
- Mascaro, J.; Perfecto, I.; Barros, O.; Boucher, D.H.; Granzow de la Cerda, I.; Ruiz, J.; Vandermeer, J. Aboveground biomass accumulation in a tropical wet forest in Nicaragua following a catastrophic hurricane disturbance. Biotropica 2005, 37, 600–608. [Google Scholar] [CrossRef] [Green Version]
- Meigs, G.W.; Keeton, W.S. Intermediate-severity wind disturbance in mature temperate forests: Legacy structure, carbon storage, and stand dynamics. Ecol. Appl. 2018, 28, 798–815. [Google Scholar] [CrossRef]
- Magnabosco Marra, D.; Trumbore, S.E.; Higuchi, N.; Ribeiro, G.H.P.M.; Negron-Juarez, R.I.; Holzwarth, F.; Rifai, S.W.; dos Santos, J.; Lima, A.J.N.; Kinupp, V.F.; et al. Windthrows control biomass patterns and functional composition of Amazon forests. Glob. Chang. Biol. 2018, 2018, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Bond-Lamberty, B.; Hicke, J.A.; Vargas, R.; Zhao, S.; Chen, J.; Edburg, S.L.; Hu, Y.; Liu, J.; McGuire, A.D. Simulating the impacts of disturbances on forest carbon cycling in North America: Processes, data, models, and challenges. J. Geophys. Res. 2011, 116, G00K08. [Google Scholar] [CrossRef] [Green Version]
- Everham, E.M.; Brokaw, N.V.L. Forest damage and recovery from catastrophic wind. Bot. Rev. 1996, 62, 113–185. [Google Scholar] [CrossRef]
- Mitchell, S.J. Wind as a natural disturbance agent in forests—A synthesis. Forestry 2013, 86, 147–157. [Google Scholar] [CrossRef] [Green Version]
- Plotkin, A.B.; Foster, D.; Carlson, J.; Magill, A. Survivors, not invaders, control forest development following simulated hurricane. Ecology 2013, 94, 414–423. [Google Scholar] [CrossRef] [Green Version]
- Webb, S.L.; Scanga, S.E. Windstorm disturbance without patch dynamics: Twelve years of change in a Minnesota forest. Ecology 2001, 82, 893–897. [Google Scholar] [CrossRef]
- Lain, E.J.; Haney, A.; Burris, J.M.; Burton, J. Response of vegetation and birds to severe wind disturbance and salvage logging in a southern boreal forest. For. Ecol. Manag. 2008, 256, 863–871. [Google Scholar] [CrossRef]
- Holzmueller, E.J.; Gibson, D.J.; Suchecki, P. Accelerated succession following an intense wind storm in an oak-dominated forest. For. Ecol. Manag. 2012, 279, 141–146. [Google Scholar] [CrossRef]
- Peterson, C.J.; Pickett, S.T.A. Forest reorganization: A case study in an old-growth forest catastrophic blowdown. Ecology 1995, 76, 763–774. [Google Scholar] [CrossRef]
- Lang, K.D.; Shulte, L.A.; Guntenspergen, G.R. Windthrow and salvage logging in an old-growth hemlock-northern hardwoods forest. For. Ecol. Manag. 2009, 259, 56–64. [Google Scholar] [CrossRef] [Green Version]
- Peterson, C.J. Twenty-five years of aboveground biomass and carbon accumulation following extreme wind damage in an old-growth forest. Forests 2019, 10, 289. [Google Scholar] [CrossRef] [Green Version]
- D’Amato, A.W.; Orwig, D.A.; Foster, D.R.; Plotkin, A.B.; Schoonmaker, P.K.; Wagner, M.R. Long-term structural and biomass dynamics of virgin Tsuga canadensis-Pinus strobus forests after hurricane disturbance. Ecology 2017, 98, 721–733. [Google Scholar] [CrossRef] [PubMed]
- Webb, S.L. Contrasting windstorm consequences in two forests, Itasca State Park, Minnesota. Ecology 1989, 70, 1167–1180. [Google Scholar] [CrossRef]
- Webb, S.L. Disturbance by wind in temperate-zone forests. In Ecosystems of Disturbed Ground; Walker, L.R., Ed.; Elsevierz: Amsterdam, The Netherland, 1999; pp. 187–222. [Google Scholar]
- Gonick, W.N.; Shearin, A.E.; Hill, D.E. Soil Survey, Litchfield County, Connecticut; Soil Conservation Service: Washington, DC, USA, 1970.
- National Climatic Data Center, U.S. Department of Commerce. Climate Norms for Litchfield County, CT; National Climatic Data Center, U.S. Department of Commerce: Asheville, NC, USA, 1981.
- Patterson, W.A., III; Foster, D.R. “Tabernacle Pines”—The rest of the story. J. For. 1990, 88, 23–25. [Google Scholar]
- Siccama, T.; Kim, B. Cathedral Pines 1989—An update. Interim Rep. Nat. Conserv. 1989. Unpublished. [Google Scholar]
- Navitsky, S. Pines in A Pasture: The History of Cathedral Pines, Northwestern Connecticut and the Blowdown of July 1989. Unpublished B. S. Honors Thesis, Harvard University Cambridge, Cambridge, MA, USA, 1990. [Google Scholar]
- Cobleigh, H.N., Jr. Vegetation and Soils of the Cathedral Pines. M.F. Thesis, Yale School of Forestry, New Haven, CT, USA, 1957. [Google Scholar]
- Gleason, H.A.; Cronquist, A. Manual of Vascular Plants of Northeastern United States and Adjacent Canada; New York Botanical Garden: Bronx, NY, USA, 1991; p. 910. [Google Scholar]
- National Climatic Data Center, U.S. Department of Commerce. Storm Data; Department of Commerce: Asheville, NC, USA, 1989; Volume 31.
- Tritschler, J.M.; Elliott, M. Assessment of Disturbance Patterns in the Wind Damaged Cathedral Pines of Cornwall, CT; Interim Report to The Nature Conservancy; 1989; Unpublished. [Google Scholar]
- Canham, C.D.; Papaik, M.J.; Uriarte, M.; McWilliams, W.H.; Jenkins, J.; Twery, M.J. Neighborhood analyses of canopy tree competition along environmental gradients in New England forests. Ecol. Appl. 2006, 16, 540–554. [Google Scholar] [CrossRef] [Green Version]
- Jenkins, J.C.; Chojnacky, D.C.; Heath, L.S.; Birdsey, R.A. National-scale biomass estimators for United States tree species. For. Sci. 2003, 49, 12–35. [Google Scholar]
- Fatemi, F.D.; Hanai, R.D.; Hamburg, S.P.; Vadeboncoeur, M.A.; Arthur, M.A.; Briggs, R.D.; Levine, C.R. Allometric equations for young northern hardwoods: The importance of age-specific equations for estimating aboveground biomass. Can. J. For. Res. 2011, 41, 881–891. [Google Scholar] [CrossRef]
- Brown, S.; Schroeder, P.; Birdsey, R. Aboveground biomass distribution of US eastern hardwood forests and the use of large trees as an indicator of forest development. For. Ecol. Manag. 1997, 96, 37–47. [Google Scholar] [CrossRef]
- Hoover, C.M.; Leak, W.B.; Keel, B.G. Benchmark carbon stocks from old-growth forests in northern New England, USA. For. Ecol. Manag. 2012, 266, 108–114. [Google Scholar] [CrossRef]
- Brokaw, N.V.L.; Walker, L.R. Summary of the effects of Caribbean hurricanes on vegetation. Biotropica 1991, 23, 442–447. [Google Scholar] [CrossRef]
- Peterson, C.J. Consistent influence of tree diameter and species on damage in nine eastern North America tornado blowdowns. For. Ecol. Manag. 2007, 250, 96–108. [Google Scholar] [CrossRef]
- Foster, D.R. Species and stand response to catastrophic wind in central New England. J. Ecol. 1988, 76, 135–151. [Google Scholar] [CrossRef]
- Cooper-Ellis, S.M.; Foster, D.R.; Carlton, G.; Lezberg, A. Vegetation and ecosystem response to catastrophic wind; evaluation of an experimental hurricane. Ecology 1999, 80, 2683–2696. [Google Scholar] [CrossRef]
- Peterson, C.J.; Carson, W.P.; McCarthy, B.C.; Pickett, S.T.A. Microsite variation and soil dynamics within newly created treefall pits and mounds. Oikos 1990, 58, 39–46. [Google Scholar] [CrossRef]
- Carlton, G.C.; Bazzaz, F.A. Regeneration of three sympatric birch species on experimental hurricane blowdown microsites. Ecol. Monogr. 1998, 68, 99–120. [Google Scholar] [CrossRef]
- Nakashizuka, T. Regeneration of beech (Fagus crenata) after the simultaneous death of undergrowing dwarf bamboo (Sasa kurilensis). Ecol. Res. 1988, 3, 21–35. [Google Scholar] [CrossRef]
- Walker, L.R.; Landau, F.H.; Velazquez, E.; Shiels, A.B.; Sparrow, A.D. Early successional woody plants facilitate and ferns inhibit forest development on Puerto Rican landslides. J. Ecol. 2010, 98, 625–635. [Google Scholar] [CrossRef]
- Frelich, L.E.; Calcote, R.R.; Davis, M.B.; Pastor, J. Patch formation and maintenance in an old-growth hemlock-hardwood forest. Ecology 1993, 74, 513–527. [Google Scholar] [CrossRef]
- Catovsky, S.; Bazzaz, F.A. Feedbacks between canopy composition and seedling regeneration in mixed conifer-broad-leaved forests. Oikos 2008, 98, 403–420. [Google Scholar] [CrossRef] [Green Version]
- Hibbs, D.E. Forty years of forest succession in central New England. Ecology 1983, 64, 1394–1401. [Google Scholar] [CrossRef] [Green Version]
- Mabry, C.; Korsgren, T. A permanent plot study of vegetation and vegetation-site factors fifty-three years following disturbance in central New England, U.S.A. Ecoscience 1998, 5, 232–240. [Google Scholar] [CrossRef]
- Frelich, L.E.; Reich, P.B. Neighborhood effects, disturbance severity, and community stability in forests. Ecosystems 1999, 2, 151–166. [Google Scholar] [CrossRef] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peterson, C.J. First-Decade Biomass and Carbon Accumulation, and Woody Community Change after Severe Wind Damage in a Hemlock-White Pine Forest Remnant. Forests 2021, 12, 231. https://doi.org/10.3390/f12020231
Peterson CJ. First-Decade Biomass and Carbon Accumulation, and Woody Community Change after Severe Wind Damage in a Hemlock-White Pine Forest Remnant. Forests. 2021; 12(2):231. https://doi.org/10.3390/f12020231
Chicago/Turabian StylePeterson, Chris J. 2021. "First-Decade Biomass and Carbon Accumulation, and Woody Community Change after Severe Wind Damage in a Hemlock-White Pine Forest Remnant" Forests 12, no. 2: 231. https://doi.org/10.3390/f12020231