
Using Sentinel-2 Images to Map the Populus euphratica Distribution Based on the Spectral Difference Acquired at the Key Phenological Stage


Abstract
:1. Introduction
2. Study Site and Data
2.1. Study Site
2.2. Data
2.2.1. Sentinel-2 Imagery
2.2.2. Reference Data
2.2.3. Ground Phenological Observation Data
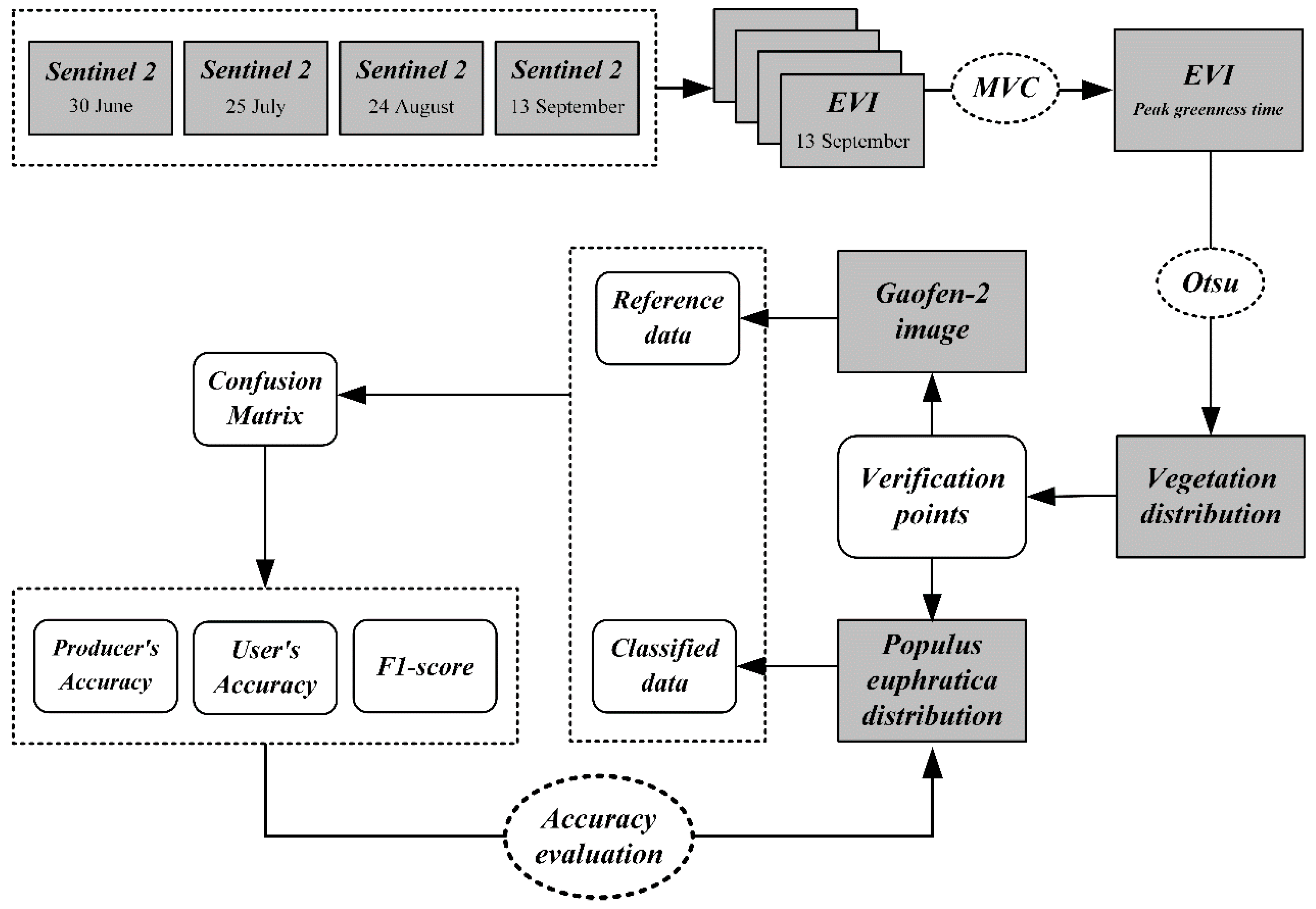
3. Methods
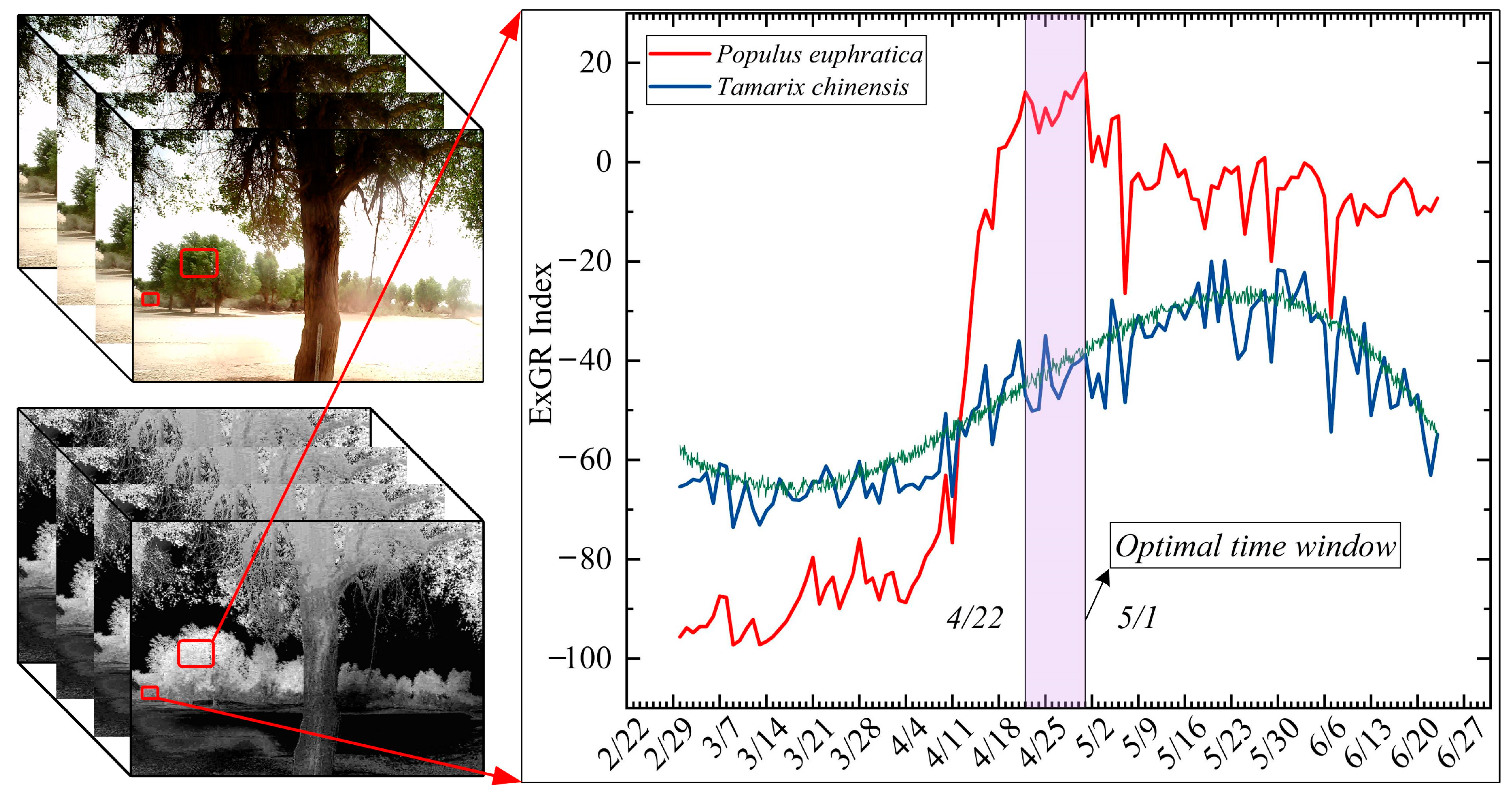
3.1. Determination of the Optimal Time Window
3.2. Screening the Appropriate Vegetation Index
3.3. Automatic Threshold Determination Method
3.4. Accuracy Evaluation
4. Results
4.1. Spectral Reflectance Difference
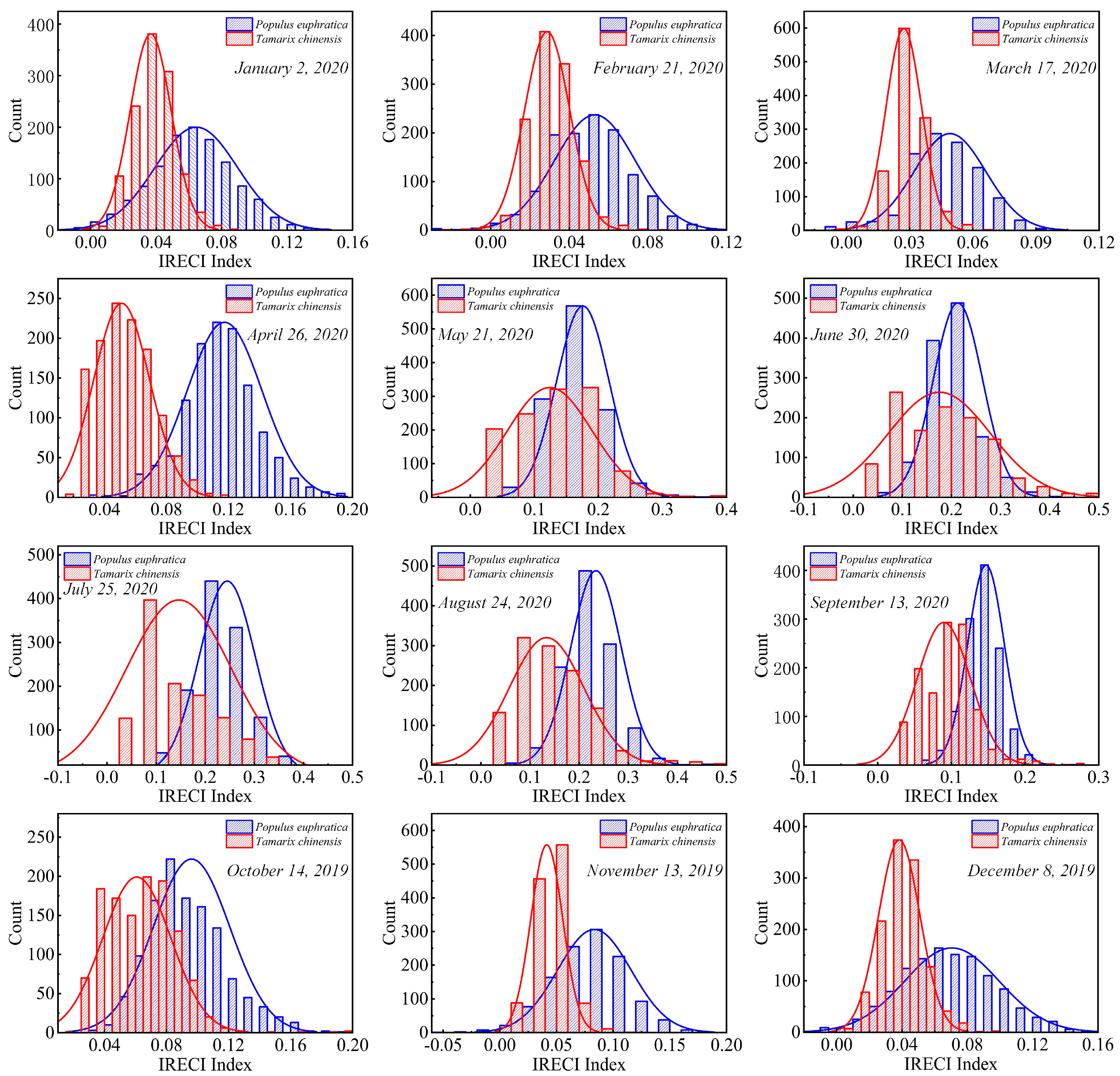
4.2. Difference in Frequency Distribution of IRECI
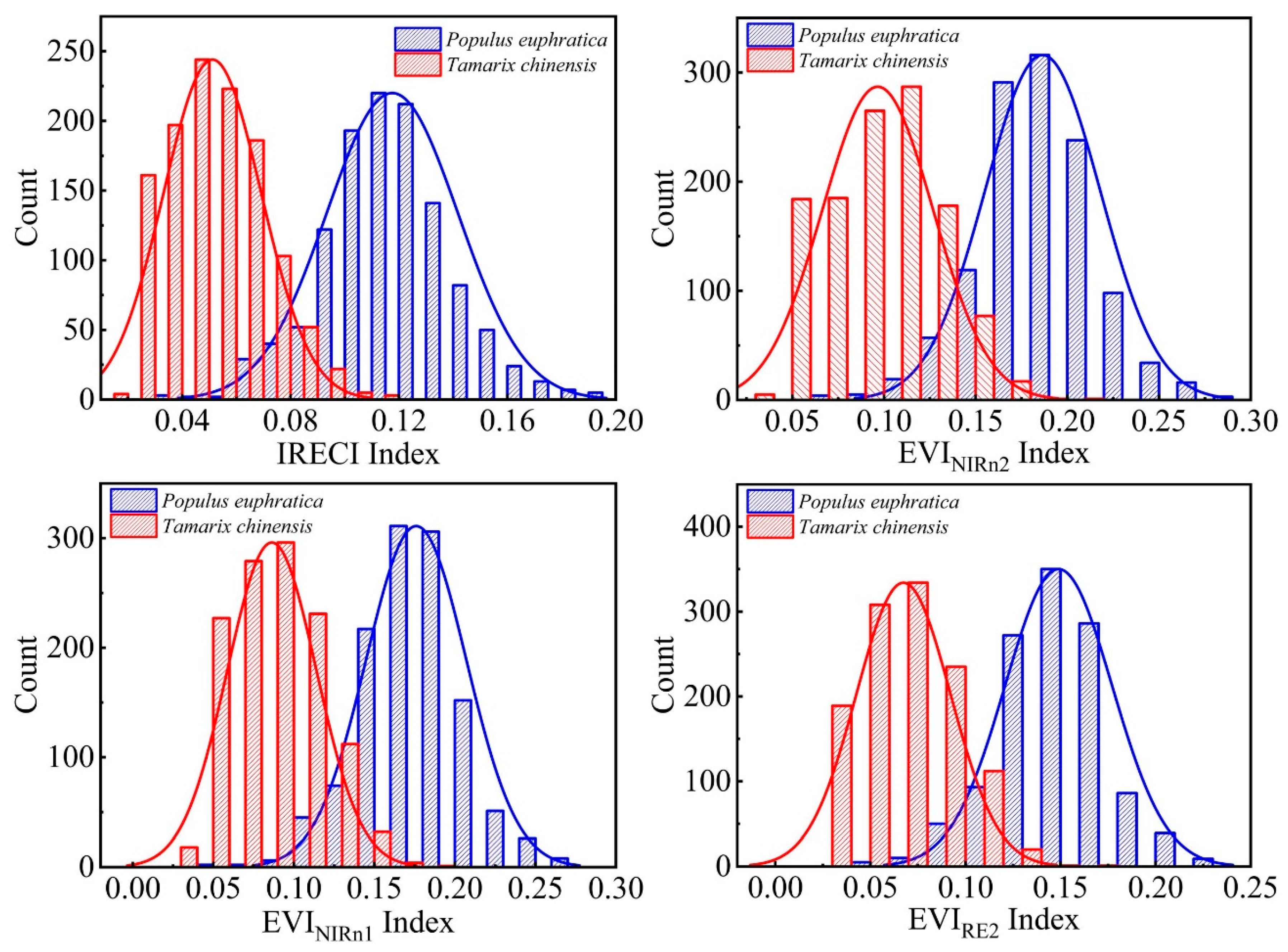
4.3. Appropriate Vegetation Index and Threshold Determination Method Selection
4.4. Populus Euphratica Distribution Mapping Results
5. Discussion
5.1. Ground Verification for Optimal Time Window
5.2. Main Factors Affecting the Extraction Accuracy of Populus euphratica
5.3. Advantages of Sentinel-2 Image in Mapping Populus euphratica Distribution
5.4. Result Applications and Limitations
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lang, P.; Jeschke, M.; Wommelsdorf, T.; Backes, T.; Lv, C.; Zhang, X.; Thomas, F.M. Wood harvest by pollarding exerts long-term effects on Populus euphratica stands in riparian forests at the Tarim River, NW China. For. Ecol. Manag. 2015, 353, 87–96. [Google Scholar] [CrossRef]
- Aishan, T.; Halik, Ü.; Betz, F.; Gärtner, P.; Cyffka, B. Modeling height–diameter relationship for Populus euphratica in the Tarim riparian forest ecosystem, Northwest China. J. For. Res. 2016, 27, 889–900. [Google Scholar] [CrossRef]
- Chen, Y.; Chen, Y.; Xu, C.; Ye, Z.; Li, Z.; Zhu, C.; Ma, X. Effects of ecological water conveyance on groundwater dynamics and riparian vegetation in the lower reaches of Tarim River, China. Hydrol. Process. 2010, 24, 170–177. [Google Scholar] [CrossRef]
- Ling, H.; Zhang, P.; Xu, H.; Zhao, X. How to Regenerate and Protect Desert Riparian Populus euphratica Forest in Arid Areas. Sci. Rep. 2015, 5, 15418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halik, Ü.; Aishan, T.; Betz, F.; Kurban, A.; Rouzi, A. Effectiveness and challenges of ecological engineering for desert riparian forest restoration along China’s largest inland river. Ecol. Eng. 2019, 127, 11–22. [Google Scholar] [CrossRef]
- Immitzer, M.; Böck, S.; Einzmann, K.; Vuolo, F.; Pinnel, N.; Wallner, A.; Atzberger, C. Fractional cover mapping of spruce and pine at 1ha resolution combining very high and medium spatial resolution satellite imagery. Remote Sens. Environ. 2018, 204, 690–703. [Google Scholar] [CrossRef] [Green Version]
- Persson, M.; Lindberg, E.; Reese, H. Tree Species Classification with Multi-Temporal Sentinel-2 Data. Remote Sens. 2018, 10, 1794. [Google Scholar] [CrossRef] [Green Version]
- Immitzer, M.; Atzberger, C.; Koukal, T. Tree Species Classification with Random Forest Using Very High Spatial Resolution 8-Band WorldView-2 Satellite Data. Remote Sens. 2012, 4, 2661–2693. [Google Scholar] [CrossRef] [Green Version]
- Diao, C.; Wang, L. Incorporating plant phenological trajectory in exotic saltcedar detection with monthly time series of Landsat imagery. Remote Sens. Environ. 2016, 182, 60–71. [Google Scholar] [CrossRef]
- Ji, W.; Wang, L. Phenology-guided saltcedar (Tamarix spp.) mapping using Landsat TM images in western U.S. Remote Sens. Environ. 2016, 173, 29–38. [Google Scholar] [CrossRef]
- Pu, R.; Landry, S. A comparative analysis of high spatial resolution IKONOS and WorldView-2 imagery for mapping urban tree species. Remote Sens. Environ. 2012, 124, 516–533. [Google Scholar] [CrossRef]
- Xun, L.; Wang, L. An object-based SVM method incorporating optimal segmentation scale estimation using Bhattacharyya Distance for mapping salt cedar (Tamarisk spp.) with QuickBird imagery. GIScience Remote Sens. 2015, 52, 257–273. [Google Scholar] [CrossRef]
- Daryaei, A.; Sohrabi, H.; Atzberger, C.; Immitzer, M. Fine-scale detection of vegetation in semi-arid mountainous areas with focus on riparian landscapes using Sentinel-2 and UAV data. Comput. Electron. Agric. 2020, 177, 105686. [Google Scholar] [CrossRef]
- Immitzer, M.; Neuwirth, M.; Böck, S.; Brenner, H.; Vuolo, F.; Atzberger, C. Optimal Input Features for Tree Species Classification in Central Europe Based on Multi-Temporal Sentinel-2 Data. Remote Sens. 2019, 11, 2599. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.; Liu, D. Accurate mapping of forest types using dense seasonal Landsat time-series. ISPRS J. Photogramm. Remote Sens. 2014, 96, 1–11. [Google Scholar] [CrossRef]
- Ji, W.; Wang, L. Discriminating Saltcedar (Tamarix ramosissima) from Sparsely Distributed Cottonwood (Populus euphratica) Using a Summer Season Satellite Image. Photogramm. Eng. Remote Sens. 2015, 81, 795–806. [Google Scholar] [CrossRef]
- Hill, R.A.; Wilson, A.K.; George, M.; Hinsley, S.A. Mapping tree species in temperate deciduous woodland using time-series multi-spectral data. Appl. Veg. Sci. 2010, 13, 86–99. [Google Scholar] [CrossRef]
- Dennison, P.E.; Roberts, D.A. The effects of vegetation phenology on endmember selection and species mapping in southern California chaparral. Remote Sens. Environ. 2003, 87, 295–309. [Google Scholar] [CrossRef]
- Kollert, A.; Bremer, M.; Löw, M.; Rutzinger, M. Exploring the potential of land surface phenology and seasonal cloud free composites of one year of Sentinel-2 imagery for tree species mapping in a mountainous region. Int. J. Appl. Earth Obs. Geoinf. 2021, 94. [Google Scholar] [CrossRef]
- Schriever, J.R.; Congalton, R. Evaluating seasonal variability as an aid to cover-type mapping from Landsat Thematic Mapper data in the Northeast. Photogramm. Eng. Remote Sens. 1995, 61, 321–327. [Google Scholar]
- Hu, Q.; Sulla-Menashe, D.; Xu, B.; Yin, H.; Tang, H.; Yang, P.; Wu, W. A phenology-based spectral and temporal feature selection method for crop mapping from satellite time series. Int. J. Appl. Earth Obs. Geoinf. 2019, 80, 218–229. [Google Scholar] [CrossRef]
- Weil, G.; Lensky, I.M.; Levin, N. Using ground observations of a digital camera in the VIS-NIR range for quantifying the phenology of Mediterranean woody species. Int. J. Appl. Earth Obs. Geoinf. 2017, 62, 88–101. [Google Scholar] [CrossRef]
- Cheng, Y.; Vrieling, A.; Fava, F.; Meroni, M.; Marshall, M.; Gachoki, S. Phenology of short vegetation cycles in a Kenyan rangeland from PlanetScope and Sentinel-2. Remote Sens. Environ. 2020, 248, 112004. [Google Scholar] [CrossRef]
- Chen, J.; Jönsson, P.; Tamura, M.; Gu, Z.; Matsushita, B.; Eklundh, L. A simple method for reconstructing a high-quality NDVI time-series data set based on the Savitzky–Golay filter. Remote Sens. Environ. 2004, 91, 332–344. [Google Scholar] [CrossRef]
- Ma, M.; Veroustraete, F. Reconstructing pathfinder AVHRR land NDVI time-series data for the Northwest of China. Adv. Space Res. 2006, 37, 835–840. [Google Scholar] [CrossRef]
- Cai, Z.; Jönsson, P.; Jin, H.; Eklundh, L. Performance of Smoothing Methods for Reconstructing NDVI Time-Series and Estimating Vegetation Phenology from MODIS Data. Remote Sens. 2017, 9, 1271. [Google Scholar] [CrossRef] [Green Version]
- Wulder, M.A.; Loveland, T.R.; Roy, D.P.; Crawford, C.J.; Masek, J.G.; Woodcock, C.E.; Allen, R.G.; Anderson, M.C.; Belward, A.S.; Cohen, W.B.; et al. Current status of Landsat program, science, and applications. Remote Sens. Environ. 2019, 225, 127–147. [Google Scholar] [CrossRef]
- Grabska, E.; Frantz, D.; Ostapowicz, K. Evaluation of machine learning algorithms for forest stand species mapping using Sentinel-2 imagery and environmental data in the Polish Carpathians. Remote Sens. Environ. 2020, 251, 112103. [Google Scholar] [CrossRef]
- Kowalski, K.; Senf, C.; Hostert, P.; Pflugmacher, D. Characterizing spring phenology of temperate broadleaf forests using Landsat and Sentinel-2 time series. Int. J. Appl. Earth Obs. Geoinf. 2020, 92, 102172. [Google Scholar] [CrossRef]
- Jr, J.G.; Civco, D.; Silander, J. Delineating forest canopy species in the Northeastern United States using multi-temporal TM imagery. Photogramm. Eng. Remote Sens. 1998, 64, 891–904. [Google Scholar]
- Wang, L.; Silván-Cárdenas, J.L.; Yang, J.; Frazier, A.E. Invasive Saltcedar (Tamarisk spp.) Distribution Mapping Using Multiresolution Remote Sensing Imagery. Prof. Geogr. 2013, 65, 1–15. [Google Scholar] [CrossRef]
- Rundquist, B.C.; Brookman, D.A. Spectral characterization of the invasive shrub saltcedar (Tamarix spp.) in North Dakota. Geocarto Int. 2007, 22, 63–72. [Google Scholar] [CrossRef]
- Hao, L.; Qingdong, S.; Imin, B.; Kasim, N. Methodology for optimizing quadrat size in sparse vegetation surveys: A desert case study from the Tarim Basin. PLoS ONE 2020, 15, e0235469. [Google Scholar] [CrossRef] [PubMed]
- Masemola, C.; Cho, M.A.; Ramoelo, A. Sentinel-2 time series based optimal features and time window for mapping invasive Australian native Acacia species in KwaZulu Natal, South Africa. Int. J. Appl. Earth Obs. Geoinf. 2020, 93, 102207. [Google Scholar] [CrossRef]
- Wai-Tim, N.; Purity, R.; Kathrin, E.; Markus, I.; Clement, A.; Sandra, E.J.R.S. Assessing the Potential of Sentinel-2 and Pléiades Data for the Detection of Prosopis and Vachellia spp. in Kenya. Remote Sens. 2017, 9, 74. [Google Scholar]
- Chenghai, Y.; James, H.E.; Reginald, S.F. Evaluating airborne hyperspectral imagery for mapping saltcedar infestations in west Texas. J. Appl. Remote Sens. 2013, 7, 1. [Google Scholar] [CrossRef]
- Aragones, D.; Rodriguez-Galiano, V.F.; Caparros-Santiago, J.A.; Navarro-Cerrillo, R.M. Could land surface phenology be used to discriminate Mediterranean pine species? Int. J. Appl. Earth Obs. Geoinf. 2019, 78, 281–294. [Google Scholar] [CrossRef]
- Rodriguez-Galiano, V.; Dash, J.; Atkinson, P. Characterising the Land Surface Phenology of Europe Using Decadal MERIS Data. Remote Sens. 2015, 7, 9390–9409. [Google Scholar] [CrossRef] [Green Version]
- Shi, Q.; Guo, Y.; Zhou, X. Machanism of the influence of surface water and groundwater on vegetation pattern in Daliyaboyi oasis at the tail of Keriya river in Taklamakan deser. J. Xinjiang Univ. 2019, 36, 53–259. [Google Scholar] [CrossRef]
- Zhang, F.; Wang, T.; Yimit, H.; Shi, Q.; Ruan, Q.; Sun, Z.; Li, F. Hydrological changes and settlement migrations in the Keriya River delta in central Tarim Basin ca. 2.7–1.6 ka BP: Inferred from 14C and OSL chronology. Sci. China Earth Sci. 2011, 54, 1971–1980. [Google Scholar] [CrossRef]
- Foody, G.M. Status of land cover classification accuracy assessment. Remote Sens. Environ. 2002, 80, 185–201. [Google Scholar] [CrossRef]
- Frampton, W.J.; Dash, J.; Watmough, G.; Milton, E.J. Evaluating the capabilities of Sentinel-2 for quantitative estimation of biophysical variables in vegetation. ISPRS J. Photogramm. Remote Sens. 2013, 82, 83–92. [Google Scholar] [CrossRef] [Green Version]
- Majasalmi, T. The potential of Sentinel-2 data for estimating biophysical variables in a boreal forest: A simulation study. Remote Sens. Lett. 2016, 7. [Google Scholar] [CrossRef]
- Huete, A.; Didan, K.; Miura, T.; Rodriguez, E.P.; Gao, X.; Ferreira, L.G. Overview of the radiometric and biophysical performance of the MODIS vegetation indices. Remote Sens. Environ. 2002, 83, 195–213. [Google Scholar] [CrossRef]
- Meyer, G.E.; Camargo Neto, J.; Jones, D.D.; Hindman, T.W. Intensified fuzzy clusters for classifying plant, soil, and residue regions of interest from color images. Comput. Electron. Agric. 2004, 42, 161–180. [Google Scholar] [CrossRef] [Green Version]
- Kapur, J.N.; Sahoo, P.K.; Wong, A.K.C. A new method for gray-level picture thresholding using the entropy of the histogram. Comput. Vis. Graph. Image Process. 1985, 29, 273–285. [Google Scholar] [CrossRef]
- Otsu, N. A Threshold Selection Method from Gray-Level Histograms. IEEE Trans. Syst. Man Cybern. 1979, 9, 62–66. [Google Scholar] [CrossRef] [Green Version]
- Tsai, W.-H. Moment-preserving thresolding: A new approach. Comput. Vis. Graph. Image Process. 1985, 29, 377–393. [Google Scholar] [CrossRef]
- Ridler, T.W.; Calvard, S. Picture Thresholding Using an Iterative Selection Method. IEEE Trans. Syst. Man Cybern. 1978, 8, 630–632. [Google Scholar] [CrossRef]
- Kittler, J.; Illingworth, J. Minimum error thresholding. Pattern Recognit. 1986, 19, 41–47. [Google Scholar] [CrossRef]
- Glasbey, C.A. An Analysis of Histogram-Based Thresholding Algorithms. CVGIP Graph. Models Image Process. 1993, 55, 532–537. [Google Scholar] [CrossRef]
- Carbonneau, P.E.; Dugdale, S.J.; Breckon, T.P.; Dietrich, J.T.; Fonstad, M.A.; Miyamoto, H.; Woodget, A.S. Adopting deep learning methods for airborne RGB fluvial scene classification. Remote Sens. Environ. 2020, 251, 112107. [Google Scholar] [CrossRef]
- Hmimina, G.; Dufrêne, E.; Pontailler, J.Y.; Delpierre, N.; Aubinet, M.; Caquet, B.; de Grandcourt, A.; Burban, B.; Flechard, C.; Granier, A.; et al. Evaluation of the potential of MODIS satellite data to predict vegetation phenology in different biomes: An investigation using ground-based NDVI measurements. Remote Sens.Environ. 2013, 132, 145–158. [Google Scholar] [CrossRef]
- Zheng, Y.; Feng, M. Investigation of bud burst, shoot growth and leaf expansion in Populus euphratica of different ages. Acta Ecol. Sin. 2015, 35, 1198–1207. [Google Scholar] [CrossRef] [Green Version]
- Aishan, T.; Halik, Ü.; Kurban, A.; Cyffka, B.; Kuba, M.; Betz, F.; Keyimu, M. Eco-morphological response of floodplain forests (Populus euphratica Oliv.) to water diversion in the lower Tarim River, northwest China. Environ. Earth Sci. 2015, 73, 533–545. [Google Scholar] [CrossRef]
- Immitzer, M.; Vuolo, F.; Atzberger, C. First Experience with Sentinel-2 Data for Crop and Tree Species Classifications in Central Europe. Remote Sens. 2016, 8, 166. [Google Scholar] [CrossRef]
- Bajocco, S.; Ferrara, C.; Alivernini, A.; Bascietto, M.; Ricotta, C. Remotely-sensed phenology of Italian forests: Going beyond the species. Int. J. Appl. Earth Obs. Geoinf. 2019, 74, 314–321. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, X.; Zheng, Y.; Qiu, A.; Zhang, L. Effects of temperature on flowering phenological traits of Populus euphratica Oliv. and Populus pruinosa Schrenk populations, Xinjiang, China. J. Arid Land 2019, 11, 754–763. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Data Type | Acquisition Date | Bands | Spatial Resolution (m) | ||
|---|---|---|---|---|---|
| Day | Month | Year | |||
| Sentinel-2 | 2 | January | 2020 | Blue Red Red-edge 1 Red-edge 2 Near-infrared-1 Near-infrared-2 | 10 |
| 21 | February | ||||
| 17 | March | ||||
| 26 | April | ||||
| 21 | May | ||||
| 30 | June | ||||
| 25 | July | ||||
| 24 | August | ||||
| 13 | September | ||||
| 14 | October | 2019 | |||
| 13 | November | ||||
| 8 | December | ||||
| Gaofen-2 | 17 | September | 2018 | Blue/Green/Red/Near-infrared | 1 |
| UAV | 28 | August | 2018 | Blue/Green/Red | 0.33 |
| Name | Abbrev | Formula | Reference |
|---|---|---|---|
| Inverted Red-Edge Chlorophyll Index | IRECI | (NIRn1 − R)/(RE1/RE2) | [42,43] |
| Enhanced Vegetation Index | EVIRE2 | 2.5 × (RE2 − R)/(1 + RE2 + 6R − 7.5B) | [44] |
| EVINIRn1 | 2.5 × (NIRn1 − R)/(1 + NIRn1 + 6R − 7.5B) | ||
| EVINIRn2 | 2.5 × (NIRn2 − R)/(1 + NIRn2 + 6R − 7.5B) | ||
| EVI | 2.5 × (NIR − R)/(1 + NIR + 6R − 7.5B) | ||
| Excess Green minus Excess Red Index | ExGR | 3G − 2.4R − B | [45] |
| Month | Species | Threshold | PA (%) | UA (%) | OA (%) |
|---|---|---|---|---|---|
| January | Populus euphratica | 0.052 | 70.75 | 86.81 | 80.00 |
| Tamarix chinensis | 89.25 | 75.32 | |||
| February | Populus euphratica | 0.042 | 68.92 | 85.61 | 73.99 |
| Tamarix chinensis | 88.42 | 73.99 | |||
| March | Populus euphratica | 0.039 | 75.00 | 90.82 | 83.71 |
| Tamarix chinensis | 92.42 | 78.71 | |||
| April | Populus euphratica | 0.08 | 93.67 | 93.20 | 93.42 |
| Tamarix chinensis | 93.17 | 93.63 | |||
| May | Populus euphratica | 0.132 | 86.33 | 64.43 | 69.33 |
| Tamarix chinensis | 52.33 | 79.29 | |||
| June | Populus euphratica | 0.157 | 89.58 | 62.46 | 67.88 |
| Tamarix chinensis | 46.17 | 81.59 | |||
| July | Populus euphratica | 0.204 | 77.17 | 76.78 | 76.92 |
| Tamarix chinensis | 76.67 | 77.05 | |||
| August | Populus euphratica | 0.178 | 87.58 | 77.62 | 81.17 |
| Tamarix chinensis | 74.75 | 85.76 | |||
| September | Populus euphratica | 0.118 | 89.00 | 84.16 | 86.13 |
| Tamarix chinensis | 83.25 | 88.33 | |||
| October | Populus euphratica | 0.076 | 79.17 | 76.43 | 77.38 |
| Tamarix chinensis | 75.58 | 78.39 | |||
| November | Populus euphratica | 0.06 | 77.25 | 90.35 | 84.50 |
| Tamarix chinensis | 91.75 | 80.13 | |||
| December | Populus euphratica | 0.056 | 68.58 | 89.95 | 80.46 |
| Tamarix chinensis | 92.33 | 74.61 |
| Vegetation Index | ||||
|---|---|---|---|---|
| Overall accuracy | IRECI | EVIRE2 | EVINIRn1 | EVINIRn2 |
| 0.934 | 0.936 | 0.904 | 0.931 | |
| Threshold Method | Vegetation Index | |||
|---|---|---|---|---|
| IRECI | EVIRE2 | EVINIRn1 | EVINIRn2 | |
| Maximum entropy | 0.923 | 0.500 | 0.500 | 0.500 |
| Otsu | 0.645 | 0.604 | 0.589 | 0.563 |
| Moments | 0.592 | 0.550 | 0.555 | 0.528 |
| Isodata | 0.500 | 0.500 | 0.500 | 0.500 |
| Minimum error | 0.592 | 0.550 | 0.542 | 0.510 |
| Mean | 0.539 | 0.509 | 0.505 | 0.500 |
| Classified Data | Reference Data | |
|---|---|---|
| Populus euphratica | Other | |
| Populus euphratica | 158 | 32 |
| other | 62 | 1181 |
| PA | 158/(158 + 62) = 0.72 | |
| UA | 158/(158 + 32) = 0.83 | |
| F1-score | 2/(1/PA + 1/UA) = 0.77 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, H.; Shi, Q.; Wan, Y.; Shi, H.; Imin, B. Using Sentinel-2 Images to Map the Populus euphratica Distribution Based on the Spectral Difference Acquired at the Key Phenological Stage. Forests 2021, 12, 147. https://doi.org/10.3390/f12020147
Li H, Shi Q, Wan Y, Shi H, Imin B. Using Sentinel-2 Images to Map the Populus euphratica Distribution Based on the Spectral Difference Acquired at the Key Phenological Stage. Forests. 2021; 12(2):147. https://doi.org/10.3390/f12020147
Chicago/Turabian StyleLi, Hao, Qingdong Shi, Yanbo Wan, Haobo Shi, and Bilal Imin. 2021. "Using Sentinel-2 Images to Map the Populus euphratica Distribution Based on the Spectral Difference Acquired at the Key Phenological Stage" Forests 12, no. 2: 147. https://doi.org/10.3390/f12020147