Mechanisms by Which Pre-Commercial Thinning Increases Black Spruce Growth in Different Climates and Soil Types


 and
and
Abstract
:1. Introduction
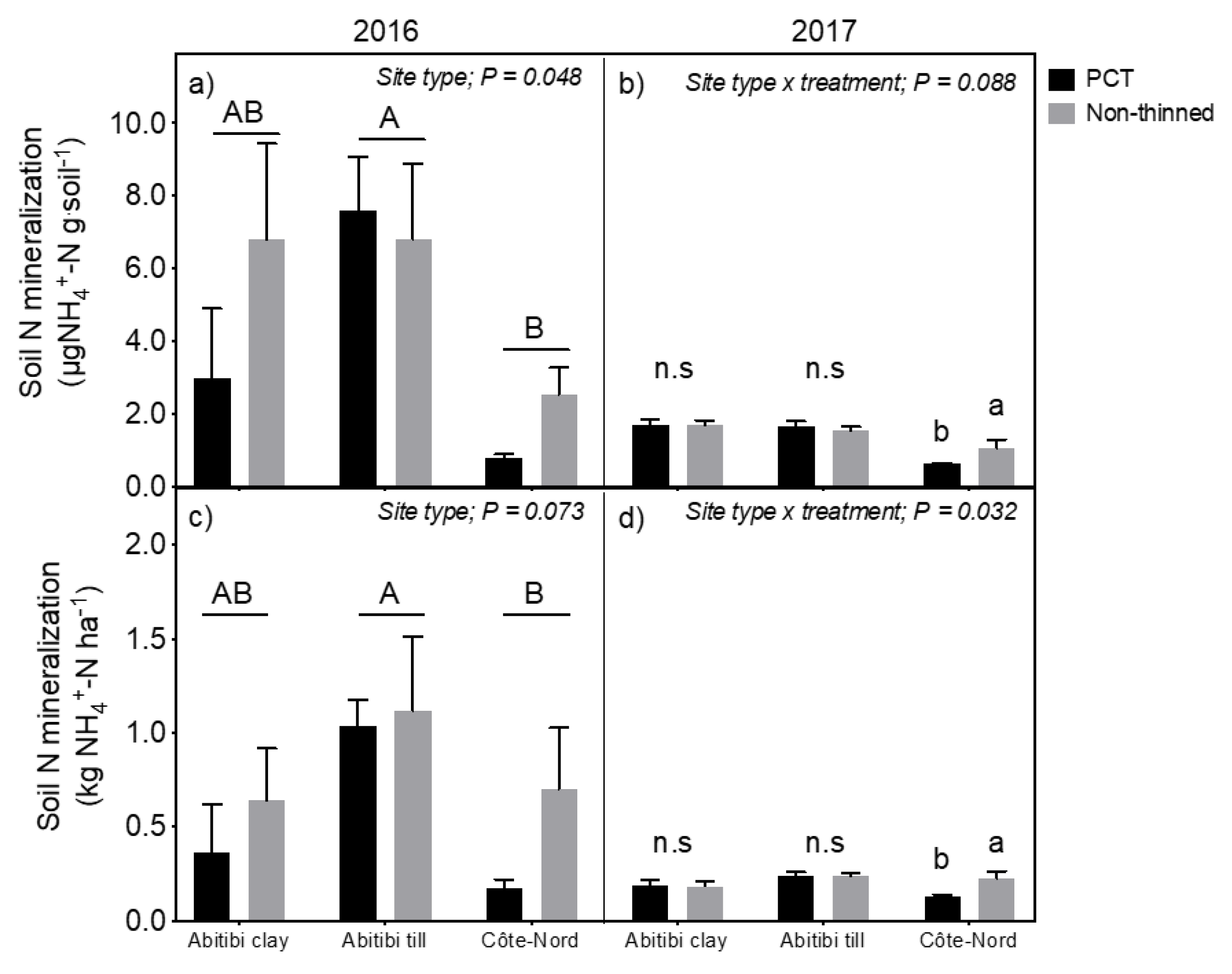
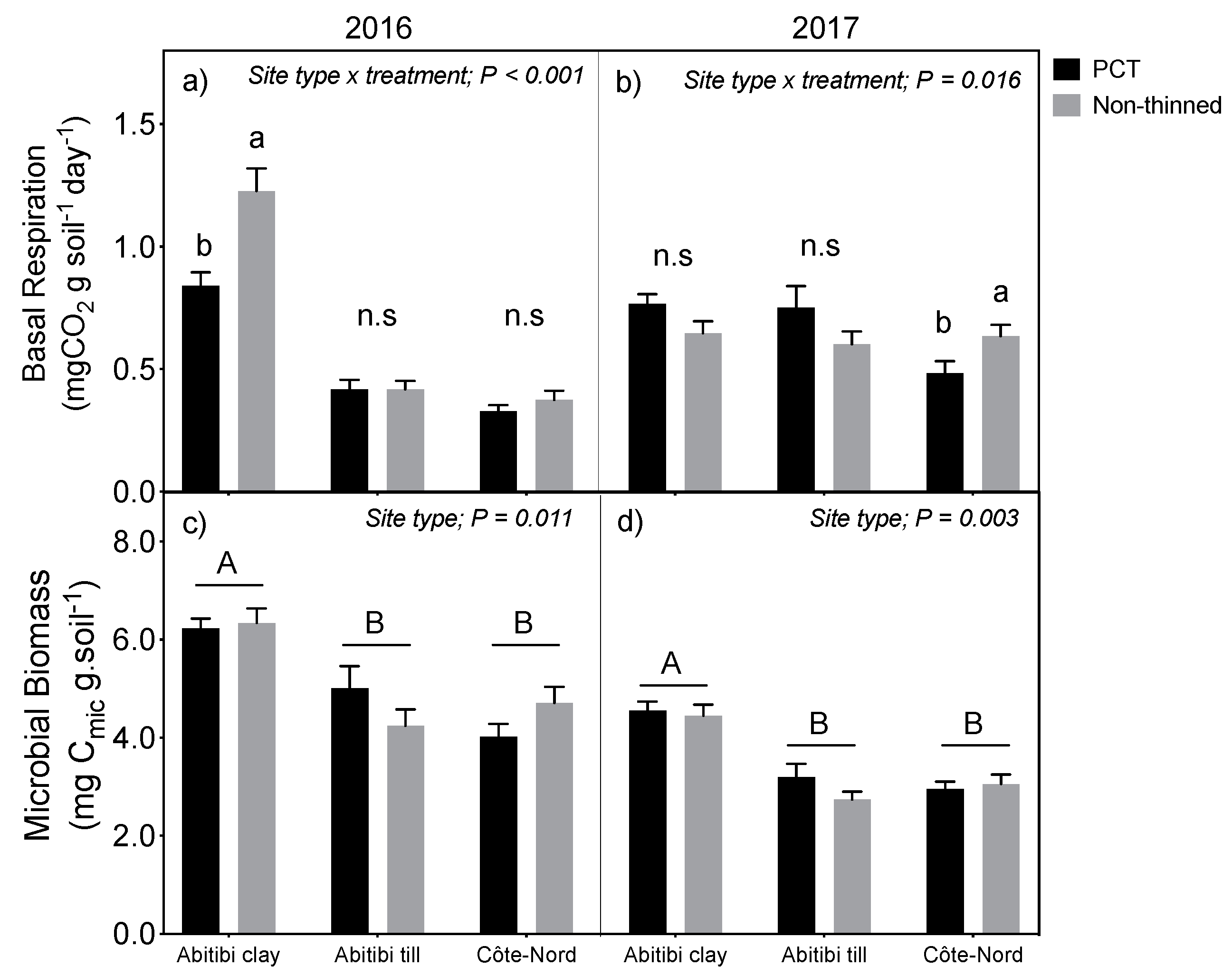
- Cuttings left on site after thinning could potentially provide a pulse of available soil nutrients, especially if decomposition is stimulated by warmer soil temperature due to canopy gaps [11,12,13]. Even if such a litter nutrient pulse was transitory, there is evidence that it may still improve long-term site fertility. For example, Bradley et al. [14] described an improvement of nutritional site quality 13 years after a single application of fertilizer in regenerating conifer cutovers on northern Vancouver Island (Canada). They proposed that fertilizer had accelerated canopy closure, which in turn improved litter quality and soil nutrition. Such a mechanism can be verified by comparing rates of soil nutrient cycling and soil microbial activity in PCT and non-thinned forest plots.
- As previously mentioned, PCT may benefit tree growth in dry regions by reducing competition for soil water [15]. By alleviating soil water limitations, PCT may reduce the ratio of net photosynthesis to stomatal conductance, better known as the water use efficiency (WUE) [16,17,18]. This can be verified either through leaf-level gas exchange measurements, or by testing for a positive relationship between δ13C and δ 18O of spruce needle tissues [19]. Conversely, PCT may be detrimental to spruce growth on poorly drained sites, if the reduction in overall evapotranspiration results in waterlogged soils.
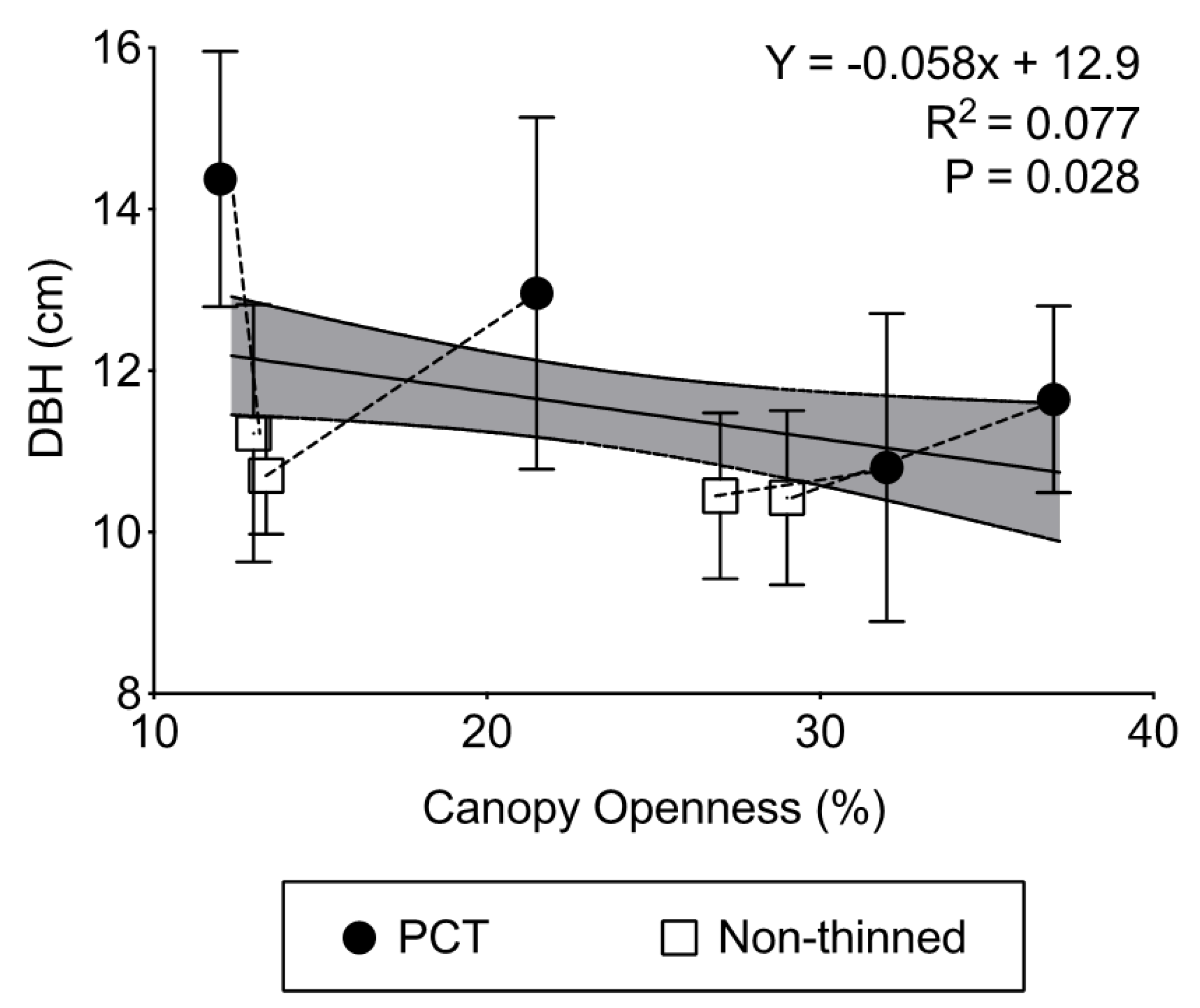
- Finally, it is unclear if PCT may increase growth rates of black spruce by increasing the capture of available light. Black spruce is a self-pruning species that regularly sheds lower branches as light intensity decreases below the canopy. By creating canopy gaps, PCT might promote the retention of subcanopy branches and needles [20], which would translate into a higher net assimilation of carbon. On the other hand, PCT plots that retain a high level of canopy openness many years after treatment may be indicative of poor growth rates resulting in slow rates of canopy closure [21]. Both of these possibilities can be verified by comparing tree growth in PCT and non-thinned plots across sites with different levels of canopy openness.
2. Materials and Methods
2.1. Study Area and Experimental Design
2.2. Tree Mensuration
2.3. Canopy Openness
2.4. Forest Floor Sampling and Analyses
2.5. Foliar Sampling and Analyses
2.6. Indices of Soil Moisture Availability
2.7. Statistical Analyses
3. Results
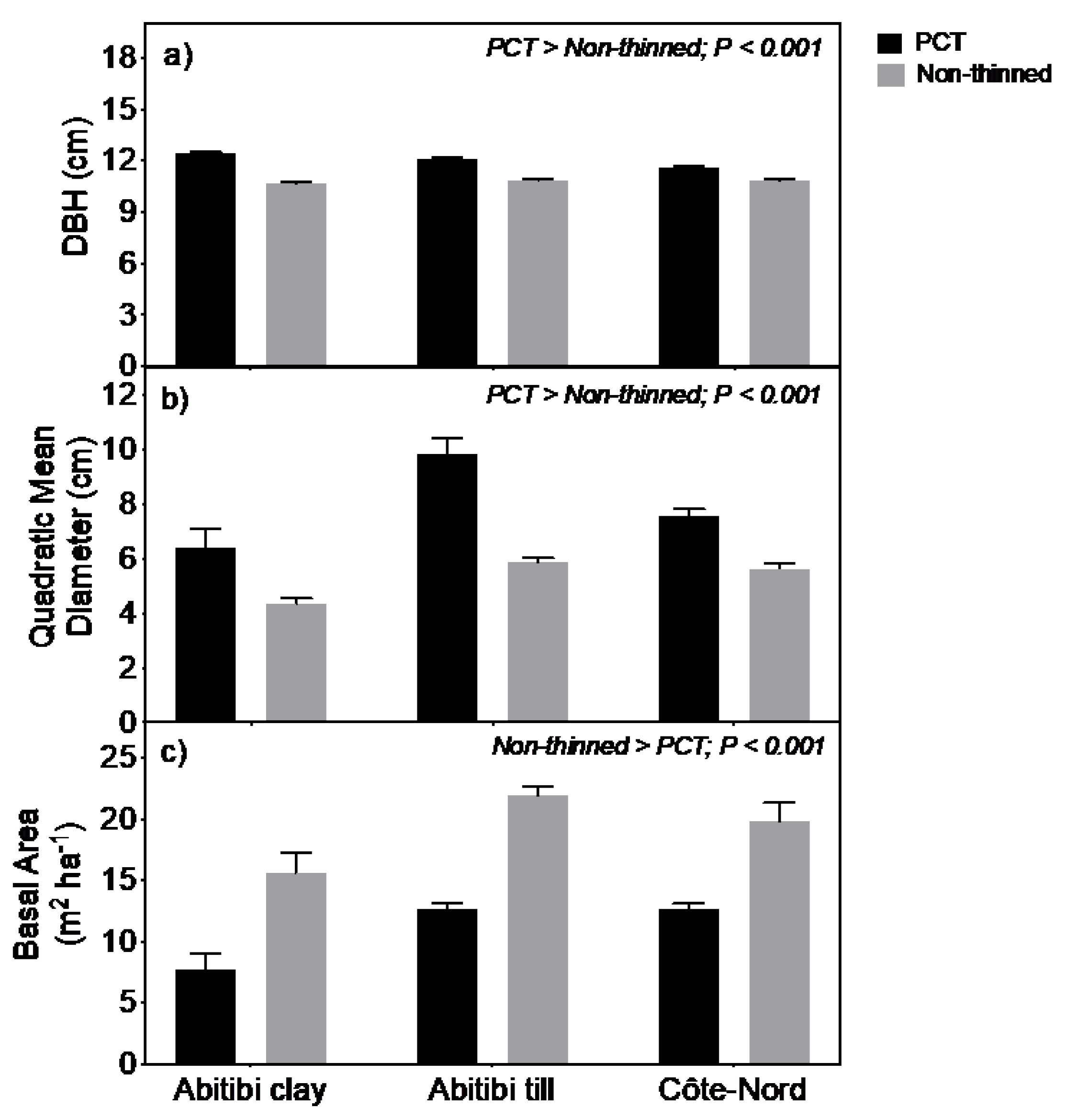
3.1. Structural Characteristics of Forest Stands
3.2. Forest Floor Nitrogen and Microbial Dynamics
3.3. Foliar Chemistry
3.4. Indices of Soil Moisture Availability
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ministère des Ressources naturelles et de la Faune. Sustainable Management in the Boreal Forest: A Real Response to Environmental Challenges; Ministère des Ressources naturelles et de la Faune, Direction de l’environnement et de la protection des forêts: Québec City, QC, Canada, 2008. [Google Scholar]
- Liu, C.; Zhang, S.Y.; Cloutier, A.; Rycabel, T. Modeling lumber bending stiffness and strength in natural black spruce stands using stand and tree characteristics. For. Ecol. Manag. 2007, 242, 648–655. [Google Scholar] [CrossRef]
- Laflèche, V.; Tremblay, S. Reésultats De Cinq Ans De La Mesure Des Effets Reéels Du Traitement d’Eéclaircie Preécommerciale De Peuplements À Dominance Reésineuse; Ministère des Ressources Naturelles et de la Faune, Direction de la Recherche Forestière: Roberval, QC, Canada, 2008. [Google Scholar]
- Bell, F.W.; Parton, J.; Stocker, N.; Joyce, D.; Reid, D.; Wester, M.; Stinson, A.; Kayahara, G.; Towill, B. Developing a silvicultural framework and definitions for use in forest management planning and practice. For. Chron. 2008, 84, 678–693. [Google Scholar] [CrossRef] [Green Version]
- Pothier, D. Twenty-year results of precommercial thinning in a balsam fir stand. For. Ecol. Manag. 2002, 168, 177–186. [Google Scholar] [CrossRef]
- Brissette, J.C.; Frank, R.M.; Stone, T.L.; Skratt, T.A. Precommercial thinning in a northern conifer stand: 18-Year results. For. Chron. 1999, 75, 967–972. [Google Scholar] [CrossRef] [Green Version]
- Poulin, J. Éducation au stade gaulis. In Bureau Du Forestier En Chef. Manuel De Détermination Des Possibilités Forestières 2013–2018; Gouvernement du Québec: Roberval, QC, Canada, 2013; pp. 83–85. [Google Scholar]
- Brockley, R.P. Effects of post-thinning density and repeated fertilization on the growth and development of young lodgepole pine. Can. J. For. Res. 2005, 35, 1952–1964. [Google Scholar] [CrossRef]
- Sohn, J.A.; Gebhardt, T.; Ammer, C.; Bauhus, J.; Häberle, K.H.; Matyssek, R.; Grams, T.E.E. Mitigation of drought by thinning: Short-term and long-term effects on growth and physiological performance of Norway spruce (Picea abies). For. Ecol. Manag. 2013, 308, 188–197. [Google Scholar] [CrossRef]
- Bureau Du Forestier En Chef. Prévisibilité, Stabilité Et Augmentation Des Possibilités Forestières; Bureau Du Forestier En Chef: Roberval, QC, Canada, 2017. [Google Scholar]
- Thibodeau, L.; Raymond, P.; Camiré, C.; Munson, A.D. Impact of precommercial thinning in balsam fir stands on soil nitrogen dynamics, microbial biomass, decomposition, and foliar nutrition. Can. J. For. Res. 2000, 30, 229–238. [Google Scholar] [CrossRef]
- Gray, A.N.; Spies, T.A.; Easter, M.J. Microclimatic and soil moisture responses to gap formation in coastal Douglas-fir forests. Can. J. For. Res. 2002, 32, 223–343. [Google Scholar] [CrossRef] [Green Version]
- Ritter, E. Litter decomposition and nitrogen mineralization in newly formed gaps in a Danish beech (Fagus sylvatica) forest. Soil Biol. Biochem. 2005, 27, 1237–1247. [Google Scholar] [CrossRef]
- Bradley, R.L.; Titus, B.D.; Preston, C.M.; Bennett, J. Improvement of nutritional site quality 13 years after single application of fertiliser N and P on regenerating cedar-hemlock cutovers on northern Vancouver Island, BC. Plant Soil 2000, 223, 195–206. [Google Scholar] [CrossRef]
- Chase, C.W.; Kimsey, M.J.; Shaw, T.M.; Coleman, M.D. The response of light, water, and nutrient availability to pre-commercial thinning in dry inland Douglas-fir forests. For. Ecol. Manag. 2016, 363, 98–109. [Google Scholar] [CrossRef]
- Kolb, T.E.; Holmberg, J.M.; Wagner, M.R.; Stone, J.E. Regulation of ponderosa pine foliar physiology and insect resistance mechanisms by basal area treatments. Tree Physiol. 1998, 18, 375–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolb, T.E.; Stone, J.E. Differences in leaf gas exchange and water relations among species and tree sizes in an Arizona pine-oak forest. Tree Physiol. 2000, 12, 1–12. [Google Scholar] [CrossRef] [PubMed]
- McDowell, N.; Brooks, J.R.; Fitzgerald, S.A.; Bond, B.J. Carbon isotope discrimination and growth response of old Pinus ponderosa trees to stand density reductions. Plant Cell Environ. 2003, 26, 631–645. [Google Scholar] [CrossRef]
- Farquhar, G.; O’Leary, M.; Berry, J. On the Relationship between Carbon Isotope Discrimination and the Intercellular Carbon Dioxide Concentration in Leaves. Aust. J. Plant Physiol. 1982, 9, 121–137. [Google Scholar] [CrossRef]
- Landsberg, J.; Sands, P. The 3-PG Process-Based Model. In Physiological Ecology of Forest Production; Terrestrial, Ecology; Landsberg, J., Sands, P., Eds.; Elsevier: Amsterdam, The Netherlands, 2011; Volume 4, pp. 241–282. [Google Scholar]
- Pacé, M.; Barrette, M.; Fenton, N.J.; Paré, D.; Bergeron, Y. Ground-Layer Composition May Limit the Positive Impact of Precommercial Thinning on Boreal Stand Productivity. For. Sci. 2017, 63, 559–568. [Google Scholar] [CrossRef] [Green Version]
- Morneau, C.; Landry, Y. Guide De Reconnaissance Des Types écologique De La Région écologique 6j—Hautes Collines Du Lac Cacaoui; Ministère des Ressources naturelles et de la Faune, Forêt Québec, Direction des inventaires forestiers, Division de l’analyse et de la diffusion des informations forestières et écologiques: Roberval, QC, Canada, 2010. [Google Scholar]
- Blouin, J.; Berger, J.-P. Guide De Reconnaissance Des Types écologiques De La Région écologique 5a—Plaine De L’Abitibi; Ministère des Ressources naturelles du Québec, Forêt Québec, Direction des inventaires forestiers, Division de la classification écologique et productivité des stations: Québec, QC, Canada, 2002. [Google Scholar]
- Vincent, J.-S.; Hardy, L. L’évolution et l’extension des lacs glaciaires Barlow et Ojibway en territoire québécois Article abstract. Géographie Phys. Quat. 1977, 31, 357–372. [Google Scholar] [CrossRef] [Green Version]
- Soil Classification Working Group. The Canadian System of Soil Classification, 3rd ed.; Agriculture and Agri-Food Canada Publication, Ed.; NRC Research Press: Ottawa, ON, Canada, 1998. [Google Scholar]
- Curtis, R.O.; Marshall, D.D. Technical note: Why quadratic mean diameter? West. J. Appl. For. 2000, 15, 137–139. [Google Scholar] [CrossRef] [Green Version]
- Frazer, G.W.; Canham, C.D.; Lertzman, K.P. Gap Light Analyzer (GLA), Version 2.0: Imaging Software to Extract Canopy Structure and Gap Light Transmission Indices from True-Colour Fisheye Photographs, Users Manual and Program Documentation; Copyright © 1999: Simon Fraser University, Burnaby, British Columbia, and the Institute of Ecosystem Studies: Millbrook, NY, USA, 1999. [Google Scholar]
- Bond-Lamberty, B.; Gower, S.T. Decomposition and fragmentation of coarse woody debris: Re-visiting a boreal black spruce chronosequence. Ecosystems 2008, 11, 831–840. [Google Scholar] [CrossRef]
- Shaw, C.H.; Bhatti, J.S.; Sabourin, K.J. An Ecosystem Carbon Database for Canadian Forests; Canadian Forest Service, Northern Forestry Centre: Edmonton, AB, Canada, 2005. [Google Scholar]
- Anderson, J.P.E.; Domsch, K.H. A physiological method for the quantitative measurement of microbial biomass in soils. Soil Biol. Biochem. 1978, 10, 215–221. [Google Scholar] [CrossRef]
- Régnière, J. Generalized Approach to Landscape-Wide Seasonal Forecasting with Temperature-Driven Simulation Models. Environ. Entomol. 1996, 25, 869–881. [Google Scholar] [CrossRef]
- Régnière, J.; St-Amant, R. Stochastic simulation of daily air temperature and precipitation from monthly normals in North America north of Mexico. Int. J. Biometeorol. 2007, 51, 415–430. [Google Scholar] [CrossRef] [PubMed]
- Ruel, J.-C.; Messier, C.; Doucet, R.; Claveau, Y.; Comeau, P. Morphological indicators of growth response of coniferous advance regeneration to overstorey removal in the boreal forest. For. Chron. 2000, 76, 633–642. [Google Scholar] [CrossRef] [Green Version]
- Linder, S. Foliar analysis for detecting and correcting nutrient imbalances in Norway spruce. Ecol. Bull. 1995, 44, 178–190. [Google Scholar]
- Hebert, F.; Thiffault, N.; Munson, A.D. Field Photosynthesis Measurements on Black Spruce (Picea mariana): Does Needle Age Matter? Commun. Soil Sci. Plant Anal. 2011, 42, 2738–2750. [Google Scholar] [CrossRef]
- Reicis, K.; Bradley, R.L.; Joanisse, G.; Houle, D.; Tremblay, S.; Barrette, M.; Wotherspoon, A. Pre-commercial thinning enhances competitive traits of boreal ericaceous shrubs and reduces soil fertility. For. Ecol. Manag. 2020, 458, 117801. [Google Scholar] [CrossRef]
- Joanisse, G.D.; Bradley, R.L.; Preston, C.M.; Bending, G.D. Sequestration of soil nitrogen as tannin-protein complexes may improve the competitive ability of sheep laurel (Kalmia angustifolia) relative to black spruce (Picea mariana). New Phytol. 2009, 181, 187–198. [Google Scholar] [CrossRef]
- Lamhamedi, M.; Bernier, P. Ecophysiology and field performance of black spruce (Picea mariana): A review. Ann. Des Sci. For. 1994, 51, 529–551. [Google Scholar] [CrossRef] [Green Version]
- Goudiaby, V.; Brais, S.; Grenier, Y.; Berninger, F. Thinning effects on jack pine and black spruce photosynthesis in Eastern boreal forests of Canada. Silva Fenn. 2011, 45, 595–609. [Google Scholar] [CrossRef] [Green Version]
- Houle, D.; Bouffard, A.; Duchesne, L.; Logan, T.; Harvey, R. Projections of Future Soil Temperature and Water Content for Three Southern Quebec Forested Sites. J. Clim. 2012, 25, 7690–7701. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Abitibi Clay | Abitibi Till | Côte-Nord | ||||
| Year of clear-cut | 1974–1978 | 1979–1982 | 1974–1979 | |||
| Year of PCT | 1995–1999 | 1996–1999 | 1997–1999 | |||
| Understory community | Kalmia angustifolia, Ledum groenlandicum, Vaccinium spp., Sphagnum spp., Alnus rugosa | Kalmia angustifolia, Vaccinium spp., Pleurozium schreberi, Diervilla lonicera | Kalmia angustifolia, Ledum groenlandicum, Pleurozium schreberi, Sphagnum spp. | |||
| PCT | Non- Thinned | PCT | Non- Thinned | PCT | Non- Thinned | |
| Stand density 15 y after treatment (stems ha−1) | 13,912 (1574) | 23,319 (373) | 8856 (1667) | 19,644 1924) | 9100 (1572) | 17,381 (3738) |
| Tree height (m) | 8.1 (0.22) | 8.3 (0.16) | 8.9 (0.30) | 8.9 (0.21) | 7.8 (0.22) | 7.7 (0.21) |
| Canopy openness (%) | 25.8 (5.6) | 20.6 (4.3) | 16.5 (2.3) | 14.5 (0.5) | 22.3 (1.5) | 20.5 (1.6) |
| Forest floor depth (cm) | 8.0 (1.2) | 7.6 (1.1) | 10.5 (0.8) | 11.3 (0.47) | 15.6 (1.4) | 17.4 (2.1) |
| Abitibi Clay | Abitibi Till | Côte-Nord | Significant Effects | ||||
|---|---|---|---|---|---|---|---|
| PCT | Non- Thinned | PCT | Non- Thinned | PCT | Non- Thinned | ||
| Foliar N (mg g−1) | 9.19 (0.17) | 9.09 (0.16) | 9.94 (0.12) | 9.95 (0.11) | 8.63 (0.11) | 8.31 (0.11) | Abitibi till > Côte Nord (p = 0.008) |
| Foliar C:N | 53.59 (1.02) | 54.01 (0.087) | 50.74 (0.60) | 49.96 (0.56) | 56.23 (0.71) | 58.60 (0.73) | Côte Nord > Abitibi till (p = 0.023) |
| δ13C | −28.31 (0.10) | −28.55 (0.07) | −28.40 (0.07) | −28.26 (0.10) | −28.56 (0.13) | −28.23 (0.08) | None |
| Abitibi Clay | Abitibi Till | Côte-Nord | |
|---|---|---|---|
| Total annual precipitation (mm) | |||
| 2016 | 664 | 664 | 1036 |
| 2017 | 684 | 684 | 1159 |
| Potential evapotranspiration (mm) | |||
| 2016 | 408 | 445 | 339 |
| 2017 | 404 | 440 | 395 |
| Climate moisture index (mm) | |||
| 2016 | 256 | 219 | 637 |
| 2017 | 280 | 244 | 764 |
| Drainage class | |||
| PCT | Imperfect | Moderate to rapid | Moderate to rapid |
| Non-thinned | Imperfect | Moderate to rapid | Moderate to rapid |
| Gravimetric moisture content (%) | |||
| PCT | 393.7 a (18.0) | 247.9 a (15.8) | 250.1 a (8.6) |
| Non-thinned | 407.1 a (24.9) | 169.9 b (11.0) | 258.0 a (8.8) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wotherspoon, A.; Bradley, R.L.; Houle, D.; Tremblay, S.; Barrette, M.; Reicis, K. Mechanisms by Which Pre-Commercial Thinning Increases Black Spruce Growth in Different Climates and Soil Types. Forests 2020, 11, 599. https://doi.org/10.3390/f11050599
Wotherspoon A, Bradley RL, Houle D, Tremblay S, Barrette M, Reicis K. Mechanisms by Which Pre-Commercial Thinning Increases Black Spruce Growth in Different Climates and Soil Types. Forests. 2020; 11(5):599. https://doi.org/10.3390/f11050599
Chicago/Turabian StyleWotherspoon, Amy, Robert L. Bradley, Daniel Houle, Stéphane Tremblay, Martin Barrette, and Krista Reicis. 2020. "Mechanisms by Which Pre-Commercial Thinning Increases Black Spruce Growth in Different Climates and Soil Types" Forests 11, no. 5: 599. https://doi.org/10.3390/f11050599