Difference in Regeneration Conditions in Pinus ponderosa Dominated Forests in Northern California, USA, over an 83 Year Period

Abstract
:1. Introduction
- What was the stocking of ponderosa pine, incense-cedar, and white fir for four different height classes in 1935 prior to active forest management across BMEF?
- Was historical stocking in BMEF associated with overstory stand structure, soil, and topography?
- Are there differences in stocking among contemporary (2018) forests that received several management treatments: control, prescribed fire, combination thinning, combination thinning with prescribed fire, thinning from below, and thinning from below with prescribed fire? If so, which treatments differ?
- How does historical stocking (1935) for the observed tree species differ from contemporary stocking (2018) for 12 compartments that received management treatments prior to 2018?
2. Materials and Methods
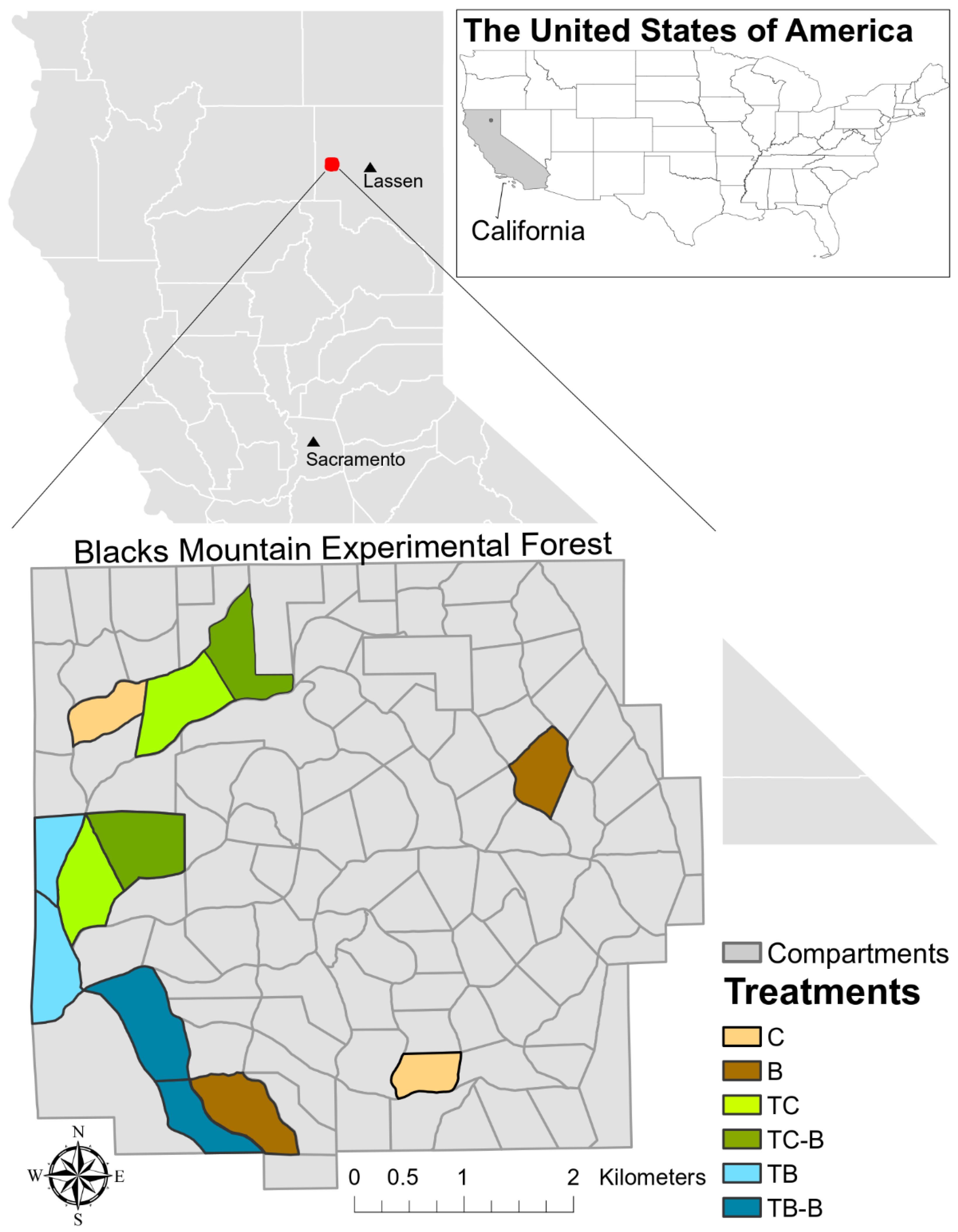
2.1. Study Area
2.2. Management History
- Control (C)
- Prescribed fire (B, burn only)—prescribed fire in 1999/1997 and 2011
- Combination thinning (TC)—thinning in 1996
- Combination thinning with prescribed fire (TC-B)—thinning in 1996 and prescribed fire in 1997
- Thinning from below only (TB)—thinning in 2012
- Thinning from below with prescribed fire (TB-B)—thinning in 2012 and prescribed fire in 2017
2.3. Regeneration Data
2.3.1. Historical Regeneration Data
2.3.2. Contemporary Regeneration Data
2.4. Data Compilation
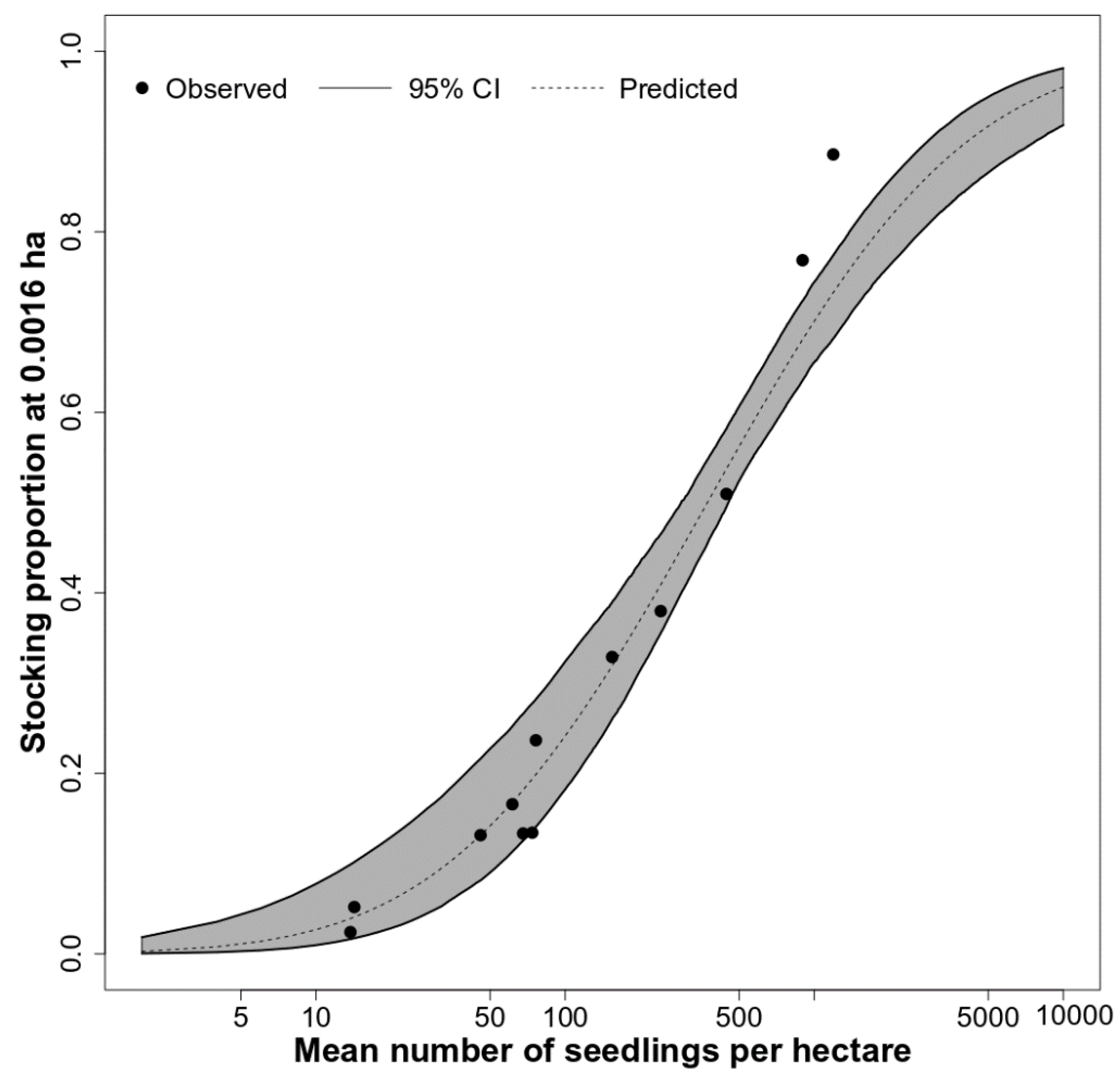
2.4.1. Stocking
2.4.2. Explanatory Variables for 1935 Conditions
2.5. Statistical Analysis
2.5.1. Historical Stocking
2.5.2. Effect of Overstory, Soil, and Topography on Stocking
2.5.3. Stocking in Contemporary Forests under Different Management Treatments
2.5.4. Stocking in Historical vs. Contemporary Forests
3. Results
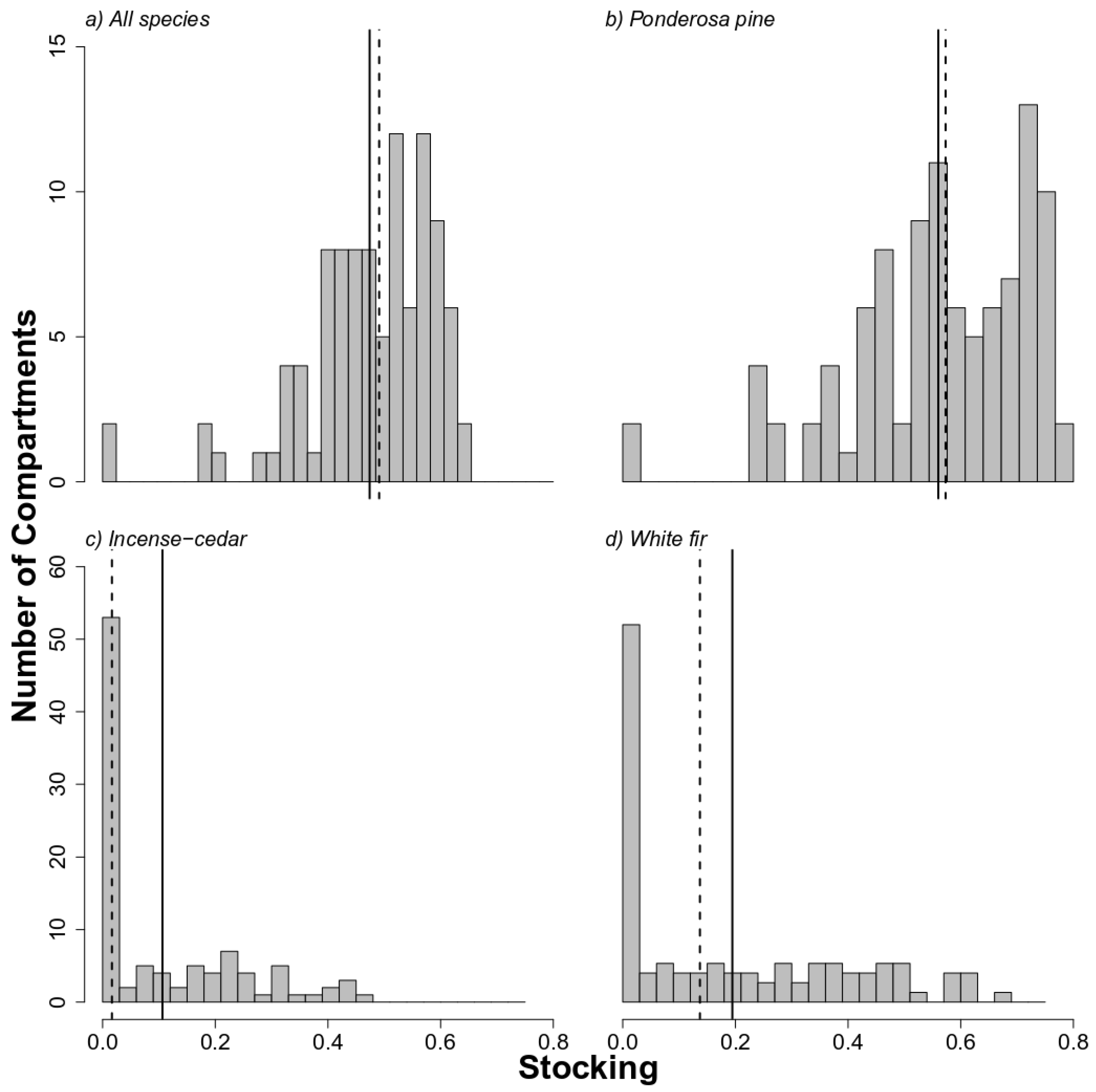
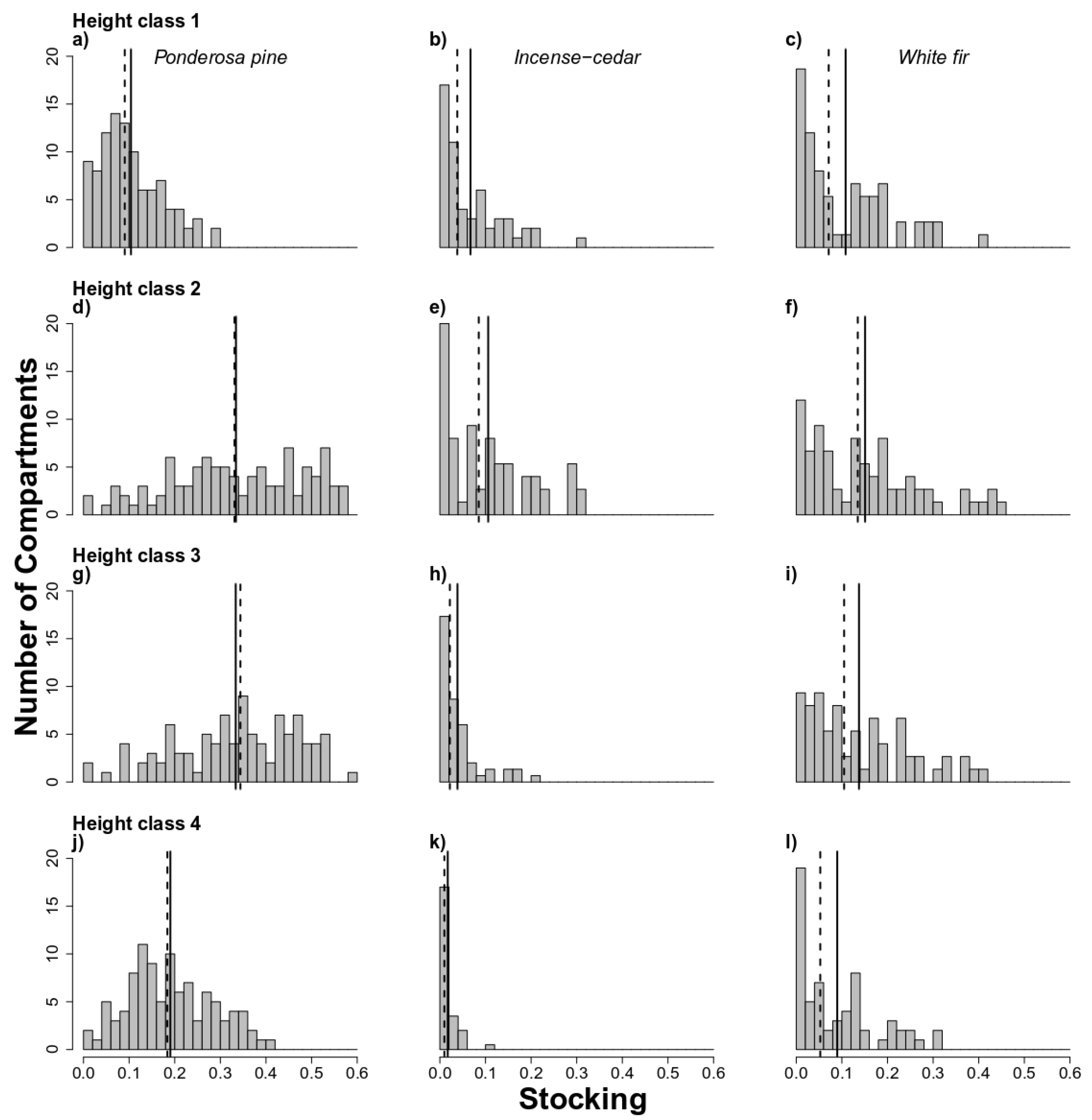
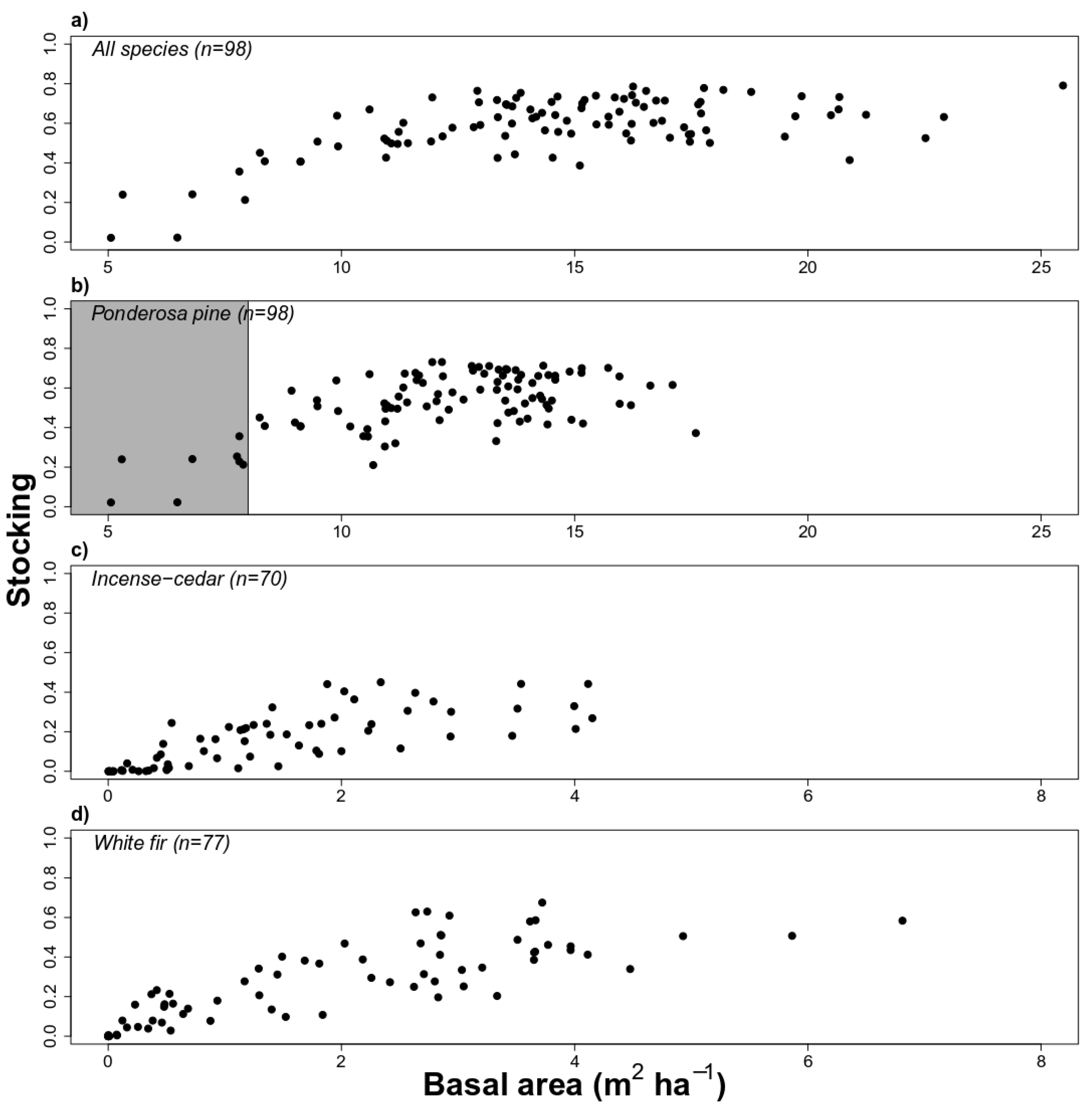
3.1. High Stocking for Ponderosa Pine Species in Historical BMEF
3.2. Overstory and Soil Variables Were Associated with Stocking
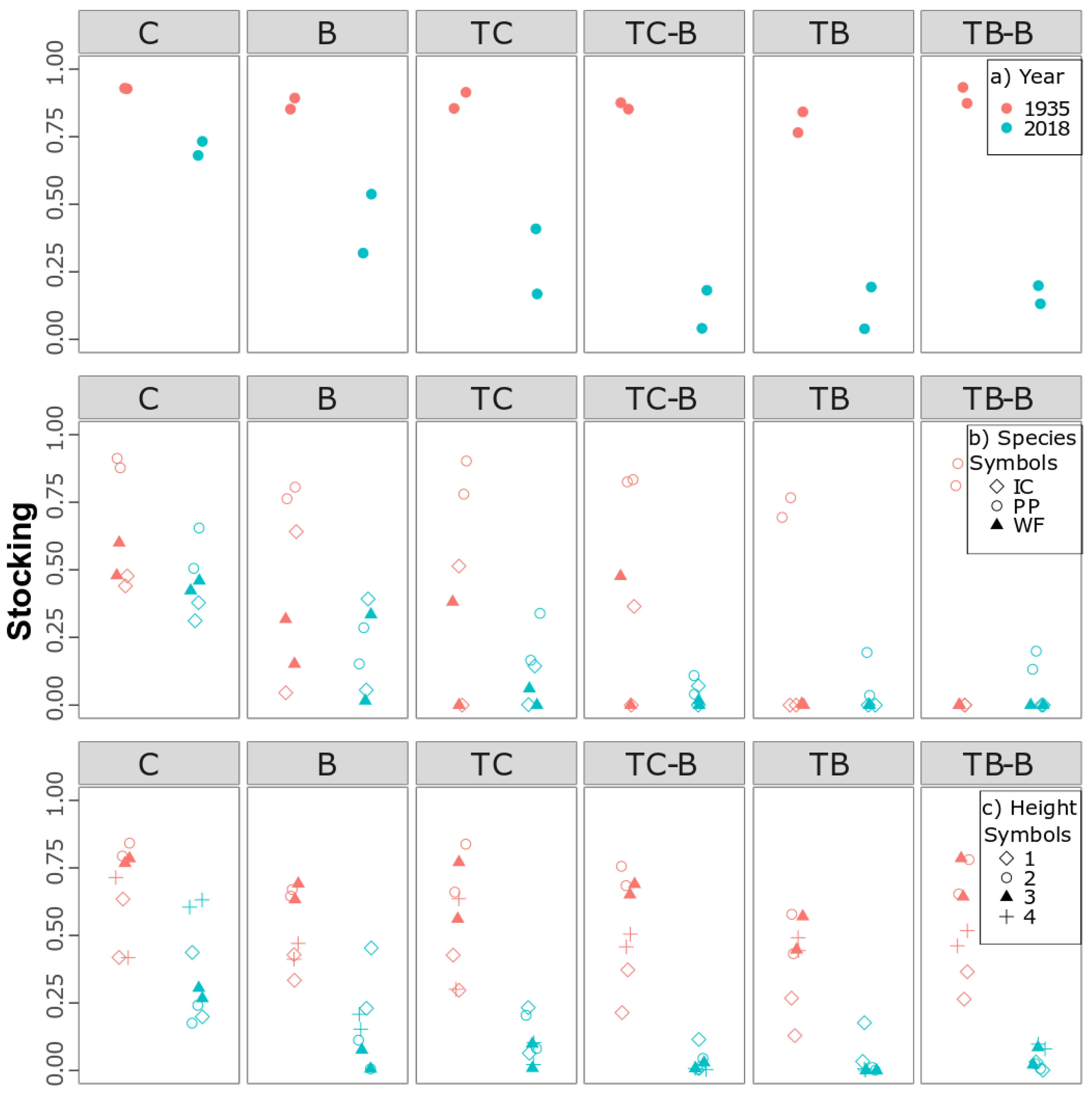
3.3. Differences in Stocking among Management Treatments in 2018
3.4. Historical Stocking Higher than Contemporary Stocking
4. Discussion
4.1. Historical Stocking Was Dominated by Ponderosa Pine at Low Elevations
4.2. Historical Stocking Was Dominated by Height Classes 2 and 3
4.3. Available Water Capacity at 150 cm and Overstory Basal Area Were Drivers of Historical Stocking
4.4. Contemporary Stocking Was Low in Thinned Stands and Differed across Treatments
4.5. Higher Historical Stocking than in Managed, Contemporary Stands
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- McKelvey, K.S.; Skinner, C.N.; Chang, C.; Erman, D.C.; Husari, S.J.; Parsons, D.J.; Van Wagtendonk, J.W.; Weatherspoon, C.P. An Overview of Fire in the Sierra Nevada; University of California, Davis, Centers for Water and Wildland Resources: Auckland, CA, USA, 1996; p. 8. [Google Scholar]
- Fitzgerald, S. Fire Ecology of Ponderosa Pine and the Rebuilding of Fire-Resilient Ponderosa Pine; Technical Report; Pacific Southwest Research Station, Forest Service, U.S. Department of Agriculture: Albany, CA, USA, 2005; pp. 197–225.
- Long, J.N.; Windmuller-Campione, M.; De Rose, R.J. Building resistance and resilience: Regeneration should not be left to chance. Forests 2018, 9, 270. [Google Scholar] [CrossRef] [Green Version]
- Puhlick, J.J.; Laughlin, D.C.; Moore, M.M. Factors influencing ponderosa pine regeneration in the southwestern USA. For. Ecol. Manag. 2012, 264, 10–19. [Google Scholar] [CrossRef]
- Brown, P.M.; Sieg, C.H. Fire history in interior ponderosa pine communities of the Black Hills, South Dakota, USA. Int. J. Wildl. Fire 1996, 6, 97–105. [Google Scholar] [CrossRef] [Green Version]
- Allen, C.D.; Savage, M.; Falk, D.A.; Suckling, K.F.; Swetnam, T.W.; Schulke, T.; Stacey, P.B.; Morgan, P.; Hoffman, M.; Klingel, J.T. Ecological restoration of southwestern ponderosa pine ecosystems: A broad perspective. Ecol. Appl. 2002, 12, 1418–1433. [Google Scholar] [CrossRef]
- Show, S.B.; Kotok, E.I. The Role of Fire in the California Pine Forests; Technical Bulletin; United States Department of Agriculture: Washington, DC, USA, 1924; p. 80.
- Taylor, A.H. Fire disturbance and forest structure in an old-growth Pinus ponderosa forest, southern Cascades, USA. J. Veg. Sci. 2010, 21, 561–572. [Google Scholar] [CrossRef]
- Weaver, H. Fire as an ecological and silvicultural factor in the ponderosa pine region of the Pacific slope. Fire Ecol. 1943, 41, 7–15. [Google Scholar] [CrossRef] [Green Version]
- Schubert, G.H. Silviculture of Southwestern Ponderosa Pine: The Status of Our Knowledge; United States Department of Agriculture Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 1974; p. 71.
- Cooper, C.F. Changes in vegetation, structure, and growth of southwestern pine forests since white settlement. Ecol. Monogr. 1960, 30, 129–164. [Google Scholar] [CrossRef]
- Dodson, E.K.; Root, H.T. Conifer regeneration following stand-replacing wildfire varies along an elevation gradient in a ponderosa pine forest, Oregon, USA. For. Ecol. Manage. 2013, 302, 163–170. [Google Scholar] [CrossRef]
- Van Pelt, R.; Franklin, J.F. Response of understory trees to experimental gaps in old-growth Douglas-fir forests. Ecol. Appl. 1999, 9, 504–512. [Google Scholar] [CrossRef]
- Angell, N.; Waring, K.M.; Graves, T.A. Predicting height growth of sugar pine regeneration using stand and individual tree characteristics. Forestry 2014, 87, 85–98. [Google Scholar] [CrossRef] [Green Version]
- Oliver, C.D.; Larson, B.C. Forest Stand Dynamics: Updated Edition; John Wiley & Sons: Hoboken, NJ, USA; C.D.Oliver & B.C. Larson: New York, NY, USA, 1996; ISBN 0471138339. [Google Scholar]
- Zald, H.S.J.; Gray, A.N.; North, M.; Kern, R.A. Initial tree regeneration responses to fire and thinning treatments in a Sierra Nevada mixed-conifer forest, USA. For. Ecol. Manag. 2008, 256, 168–179. [Google Scholar] [CrossRef]
- Bailey, J.D.; Covington, W.W. Evaluating ponderosa pine regeneration rates following ecological restoration treatments in northern Arizona, USA. For. Ecol. Manag. 2002, 155, 271–278. [Google Scholar] [CrossRef]
- McKelvey, K.S.; Johnston, J. Historical Perspectives on Forests of the Sierra Nevada and the Transverse Ranges of Southern California: Forest Conditions at the Turn of the Century; Technical Report; United States Forest Service, Pacific Southwest Research Station: Albany, CA, USA, 1992; p. 22. [Google Scholar]
- Van Wagtendonk, J.W.; Sugihara, N.G.; Stephens, S.L.; Thode, A.E.; Fites-Kaufman, J. Sierra Nevada bioregion. In Fire in California’s Ecosystems; Sugihara, N.G., Van Wagtendonk, J.W., Fites-Kaufman, J., Shaffer, K.E., Thode, A.E., Eds.; University of California Press: Auckland, CA, USA, 2006; pp. 264–294. ISBN 0520246055. [Google Scholar]
- Pearson, G.A. Management of Ponderosa Pine in the Southwest: As Developed by Research and Experimental Practice; United States Department of Agriculture: Washington, DC, USA, 1950; p. 218.
- Kilgore, B.M. The ecological role of fire in Sierran conifer forests. Its application to National Park management. Quat. Res. 1973, 3, 496–513. [Google Scholar] [CrossRef]
- Stephens, S.L.; Lydersen, J.M.; Collins, B.M.; Fry, D.L.; Meyer, M.D. Historical and current landscape-scale ponderosa pine and mixed conifer forest structure in the southern Sierra Nevada. Ecosphere 2015, 6, 1–63. [Google Scholar] [CrossRef]
- Farnden, C. Forest Regeneration in the ESSF Zone of North-Central British Columbia; Pacific and Yukon Region, Pacific Forestry Centre, Forestry Canada: Victoria, BC, Canada, 1994; p. 31. [Google Scholar]
- League, K.; Veblen, T. Climatic variability and episodic Pinus ponderosa establishment along the forest-grassland ecotones of Colorado. For. Ecol. Manag. 2006, 228, 98–107. [Google Scholar] [CrossRef]
- Savage, M.; Brown, P.M.; Feddema, J. The role of climate in a pine forest regeneration pulse in the southwestern United States. Ecoscience 1996, 3, 310–318. [Google Scholar] [CrossRef]
- Harley, G.; Baisan, C.; Brown, P.; Falk, D.; Flatley, W.; Grissino-Mayer, H.; Hessl, A.; Heyerdahl, E.; Kaye, M.; Lafon, C.; et al. Advancing dendrochronological studies of fire in the United States. Fire 2018, 1, 11. [Google Scholar] [CrossRef] [Green Version]
- Keyser, T.L.; Lentile, L.B.; Smith, F.W.; Shepperd, W.D. Changes in forest structure after a large, mixed-severity wildfire in ponderosa pine forests of the black hills, South Dakota, USA. For. Sci. 2008, 54, 328–338. [Google Scholar]
- Fulé, P.Z.; Crouse, J.E.; Roccaforte, J.P.; Kalies, E.L. Do thinning and/or burning treatments in western USA ponderosa or Jeffrey pine-dominated forests help restore natural fire behavior? For. Ecol. Manag. 2012, 269, 68–81. [Google Scholar] [CrossRef]
- Stein, W.I. Regeneration surveys and evaluation. In Reforestation Practices in Southwestern Oregon and Northern California; Hobbs, S.D., Ed.; Forest Research Laboratory, Oregon State University: Corvallis, OR, USA, 1992; pp. 346–382. [Google Scholar]
- Lowdermilk, W.C. A method for rapid surveys of vegetation. J. For. 1927, 25, 181–185. [Google Scholar]
- Lynch, D.W.; Schumacher, F.X. Concerning the dispersion of natural regeneration. J. For. 1941, 39, 49–51. [Google Scholar]
- Ritchie, M.W. Relating stocking and density for natural regeneration of conifers in northern California. For. Sci. 2020, fxaa005. [Google Scholar] [CrossRef]
- Skinner, C.N.; Taylor, A.H. Southern Cascades bioregion. In Fire in California’s Ecosystems; Sugihara, N.G., van Wagtendonk, J.W., Fites-Kaufmann, J., Shaffer, K.E., Thode, A.E., Eds.; University of California Press: Berkeley, CA, USA, 2006; pp. 195–224. [Google Scholar]
- Ritchie, M.W. Multi-scale reference conditions in an interior pine-dominated landscape in northeastern California. For. Ecol. Manag. 2016, 378, 233–243. [Google Scholar] [CrossRef]
- Wing, B.M.; Ritchie, M.W.; Boston, K.; Cohen, W.B.; Gitelman, A.; Olsen, M.J. Prediction of understory vegetation cover with airborne lidar in an interior ponderosa pine forest. Remote Sens. Environ. 2012, 124, 730–741. [Google Scholar] [CrossRef]
- Oliver, W.W. Ecological Research at the Blacks Mountain Experimental Forest in Northeastern California; Technical Report; United States Department of Agriculture Forest Service, Pacific Southwest Research Station: Albany, CA, USA, 2000; p. 66.
- Hallin, W.E. The Application of Unit Area Control in the Management of Ponderosa-Jeffrey Pine at Black Mountains Experimental Forest; Technical Bulletin; United States Department of Agriculture: Washington, DC, USA, 1959; p. 96.
- Eaton, C.B. Insect-Caused Maortality in Relation to Methods of Cutting in Ponderosa Pine on Blacks Mountain Experimental Forest; Technical Report; Pacific Southwest Forest and Range Experiment Station, Forest Service, USDA: Berkeley, CA, USA, 1959; p. 36. [Google Scholar]
- Salman, K.A.; Bongberg, J.W. Logging high risk trees to control insects in the pine stands of northern California. J. For. 1942, 40, 533–549. [Google Scholar]
- Camann, M.A.; Gillette, N.E.; Lamoncha, K.L.; Mori, S.R. Response of forest soil Acari to prescribed fire following stand structure manipulation in the southern Cascade Range. Can. J. For. Res. 2008, 38, 956–968. [Google Scholar] [CrossRef]
- Crotteau, J.S.; Ritchie, M.W. Long-term stand growth of interior ponderosa pine stands in response to structural modifications and burning treatments in northeastern California. J. For. 2014, 112, 412–423. [Google Scholar] [CrossRef] [Green Version]
- PSW Reserach Station Instruction for Type Map and Inventory of Experimental Forest; Technical Report; United States Dep. Agric. For. Serv.: Redding, CA, USA, 1935; p. 9.
- Ginrich, S. Measuring and evaluating stocking and stand density in upland hardwood forests in the central states. For. Sci. 1967, 13, 38–53. [Google Scholar]
- Gill, C. A study of the relationship between the number of trees per acre and dispersion. For. Chron. 1950, 186–196. [Google Scholar] [CrossRef] [Green Version]
- Ghent, A.W. Studies of regeneration in forest stands devastated by the spruce budworm. For. Sci. 1969, 15, 417–429. [Google Scholar]
- Alexander, E.B. Experimental Forests and Ranges of the Pacific Southwest Research Station; Unpubl. Rep. file Pacific Southwest Res. Station. United States Dep. Agric. For. Serv: Redding, CA, USA, 1994.
- Wilson, J.P. GIScience research at the 2017 Esri International User Conference. Trans. GIS 2017, 21, 431–433. [Google Scholar] [CrossRef]
- Stage, A. An expression for the effect of aspect, slope, and habitat type on tree growth. For. Sci. 1976, 22, 457–460. [Google Scholar]
- Reed, T.W.; Gulland, E.; West, G.; Mcmeekin, D.A.; Moncrieff, S. Geographic metadata searching with semantic and spatial filtering methods. In Proceedings of the GEOProcessing 2016: The Eighth International Conference on Advanced Geographic Information Systems, Applications, and Services, Venice, Italy, 24–28 April 2016. [Google Scholar]
- Beven, K.J.; Kirkby, M.J. A physically based, variable contributing area model of basin hydrology. Hydrol. Sci. Bull. 1979, 24, 43–69. [Google Scholar] [CrossRef] [Green Version]
- Dubayah, R.; Rich, P.M. Topographic solar radiation models for GIS. Int. J. Geogr. Inf. Syst. 1995, 9, 405–419. [Google Scholar] [CrossRef]
- Ferrari, S.L.P.; Cribari-Neto, F. Beta regression for modelling rates and proportions. J. Appl. Stat. 2004, 31, 799–815. [Google Scholar] [CrossRef]
- Core Team, R. R: A Language and Environment for Statistical Computing; R Foundation Statistical Computer: Vienna, Austria, 2018. [Google Scholar]
- Zuur, A.F.; Ieno, E.N.; Walker, N.J.; Saveliev, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology with R; Springer: Berlin/Heidelberg, Germany, 2009; p. 574. ISBN 978-0-387-87457-9. [Google Scholar]
- Akaike, H. Information theory and the maximum likelihood principle. In Proceedings of the 2nd International Symposium on Information Theory; Akademia Kiado: Budapest, Hungary, 1973. [Google Scholar]
- Moral, R.A.; Hinde, J.; Demétrio, C.G.B. Half-normal plots and overdispersed models in R: The hnp package. J. Stat. Softw. 2017, 81, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Kruskal, W.H.; Wallis, W.A. Use of ranks in one-criterion variance analysis. J. Am. Stat. Assoc. 1952, 47, 583–621. [Google Scholar] [CrossRef]
- Wilcoxon, F. Individual comparisons by ranking methods. Biometrics Bull. 1945, 1, 80–83. [Google Scholar] [CrossRef]
- Fowells, H.A.; Stark, N.B. Natural Regeneration in Relation to Environment in the Mixed Conifer Forest Type of California; Pacific Southwest Forest and Range Experiment Station, Forest Service, USDA: Berkeley, CA, USA, 1965; p. 14. [Google Scholar]
- Shive, K.L.; Preisler, H.K.; Welch, K.R.; Safford, H.D.; Butz, R.J.; O’Hara, K.L.; Stephens, S.L. From the stand scale to the landscape scale: Predicting the spatial patterns of forest regeneration after disturbance. Ecol. Appl. 2018, 28, 1626–1639. [Google Scholar] [CrossRef]
- Stein, S.J.; Kimberling, D.N. Germination, establishment, and mortality of naturally seeded southwestern ponderosa pine. West. J. Appl. For. 2003, 18, 109–114. [Google Scholar] [CrossRef] [Green Version]
- Flathers, K.N.; Kolb, T.E.; Bradford, J.B.; Waring, K.M.; Moser, W.K. Long-term thinning alters ponderosa pine reproduction in northern Arizona. For. Ecol. Manag. 2016, 374, 154–165. [Google Scholar] [CrossRef]
- Oliver, W.W.; Leroy Dolph, K. Mixed-conifer seedling growth varies in response to overstory release. For. Ecol. Manag. 1992, 48, 179–183. [Google Scholar] [CrossRef]
- Baker, W.L.; Williams, M.A. Bet-hedging dry-forest resilience to climate-change threats in the western USA based on historical forest structure. Front. Ecol. Evol. 2015, 2, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Oliver, W. Ponderosa pine can stagnate on a good site. J. For. 1967, 65, 814–816. [Google Scholar]
- Shepperd, W.D.; Edminster, C.B.; Mata, S.A. Long-term seedfall, establishment, survival, and growth of natural and planted ponderosa pine in the Colorado Front Range. West. J. Appl. For. 2006, 21, 19–26. [Google Scholar] [CrossRef] [Green Version]
- Kolb, T.E.; Agee, J.K.; Fulé, P.Z.; McDowell, N.G.; Pearson, K.; Sala, A.; Waring, R.H. Perpetuating old ponderosa pine. For. Ecol. Manag. 2007, 249, 141–157. [Google Scholar] [CrossRef]
- Airey Lauvaux, C.; Skinner, C.N.; Taylor, A.H. High severity fire and mixed conifer forest-chaparral dynamics in the southern Cascade Range, USA. For. Ecol. Manag. 2016, 363, 74–85. [Google Scholar] [CrossRef]
- Keyes, C.R.; Manso González, R. Climate-influenced ponderosa pine (Pinus ponderosa) seed masting trends in western Montana, USA. For. Syst. 2015, 24, 1–7. [Google Scholar] [CrossRef]
- Savage, M.; Mast, J.N. How resilient are southwestern ponderosa pine forests after crown fires? Can. J. For. Res. 2005, 35, 967–977. [Google Scholar] [CrossRef] [Green Version]
- Oliver, W.W.; Power, R.F. Early Height Growth of Ponderosa Pine: Forecast Dominance in Plantations; Pacific Southwest Forest and Range Experiment Station, Forest Service, USDA: Berkeley, CA, USA, 1971; p. 4. [Google Scholar]
- Lydersen, J.; North, M. Topographic variation in structure of mixed-conifer forests under an active-fire regime. Ecosystems 2012, 15, 1134–1146. [Google Scholar] [CrossRef]
- Haase, S.M. Effect of Prescribed Burning on Soil Moisture and Germination of Southwestern Ponderosa Pine Seed on Basaltic Soils; Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station: Fort Collins, CO, USA, 1986; p. 5. [Google Scholar]
- Harr, R.D. Water flux in soil and subsoil on a steep forested slope. J. Hydrol. 1977, 33, 37–58. [Google Scholar] [CrossRef]
- Daubenmire, R. Soil moisture in relation to vegetation distribution in the mountains of northern Idaho. Ecology 1968, 49, 431–438. [Google Scholar] [CrossRef]
- Zhang, J.; Oliver, W.W.; Ritchie, M.W.; Neal, D.L. Overstory and understory dynamics in a ponderosa pine plantation vary with stand density in the Sierra Nevada: 40-year results. For. Sci. 2013, 59, 670–680. [Google Scholar] [CrossRef]
- Clark, J.S.; Beckage, B.; Camill, P.; Cleveland, B.; HilleRisLambers, J.; Lichter, J.; McLachlan, J.; Mohan, J.; Wyckoff, P. Interpreting recruitment limitation in forests. Am. J. Bot. 1999, 86, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keeton, W.S.; Franklin, J.F. Do remnant old-growth trees accelerate rates of succession in mature Douglas-fir forests? Ecol. Monogr. 2005, 75, 103–118. [Google Scholar] [CrossRef]
- McDonald, P.M. Clearcutting and Natural Regeneration: Management Implications for the Northern Sierra Nevada; Techenical Report; Pacific Southwest Forest and Range Experiment Station, Forest Service, USDA: Berkeley, CA, USA, 1983; p. 11. [Google Scholar]
- Knapp, E.E.; Skinner, C.N.; North, M.P.; Estes, B.L. Long-term overstory and understory change following logging and fire exclusion in a Sierra Nevada mixed-conifer forest. For. Ecol. Manag. 2013, 310, 17. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Compartment | Management Description | Area (ha) | # of 0.016 ha Quadrats | |
|---|---|---|---|---|---|
| 1935 | 2018 | ||||
| RNA | |||||
| C | B24-4 (RNA A) | No treatment [36] | 35 | 296 | 140 |
| C | C4-21 (RNA D) | No treatment [36] | 38 | 210 | 108 |
| B | P21-28 (RNA B) | Prescribed fire in 1999 [36] | 39 | 288 | 157 |
| B | G2-12 (RNA C) | Prescribed fire in 1997 and 2011 [36] | 57 | 444 | 225 |
| LOD | |||||
| TC | G14-3 | Overstory above 50.8 cm removed and remaining stands thinned from below to a residual density of 250 tree per hectare in 1996 [36] | 45 | 350 | 175 |
| TC | B24-7 | Overstory above 50.8 cm removed and remaining stands thinned from below to a residual density of 250 tree per hectare in 1996 [36] | 41 | 322 | 158 |
| TC-B | G17-6 | Same as G14-3 followed by prescribed fire in 1997 [36] | 46 | 367 | 193 |
| TC-B | B27-11 | Same as B24-7 followed by prescribed fire in 1997 [36] | 77 | 642 | 210 |
| Bull thinning | |||||
| TB | G17-1 | Thinning from below up to 40.38 cm diameter at breast height (dbh) in 2012. For clumps of trees >40.38 cm dbh, half removed while largest and healthiest trees retained. Trees between 7.62–40.38 cm dbh and not in aggregates of large trees, a residual density stocking of 13.77 to 18.37 m2 ha−1 of basal area retained. Retain all snags >30.48 cm dbh except where they pose a hazard adjacent to roads or other improvements. | 32 | 230 | 131 |
| TB | G10-1 | Similar to G17-1 in 2012. But trees between 7.62–40.38 cm dbh and not in aggregates of large trees, a residual density stocking of 9.18 m2 ha−1 of basal area was retained. | 52 | 341 | 213 |
| TB-B | G7-6 | Thinning from below to a target of 18–23 m2 ha−1. Prescribed fire in fall of 2017. | 72 | 469 | 291 |
| TB-B | G2-8 | Thinning from below to a target of 18–23 m2 ha−1. Prescribed fire in fall of 2017. | 38 | 343 | 149 |
| Variables Group | Source |
|---|---|
| Response variable | |
| Stocking | Field data (1935) for 100 compartments Field data (2018) for 12 compartments |
| Explanatory variables | |
| Overstory | |
| Trees per hectare (TPH) | Field data (1934) |
| Basal area per hectare (BA, m2 ha−1) | Field data (1934) |
| Topographic variables | |
| Aspect (degrees) | DEM at 1 m resolution |
| Slope (%) | DEM at 1 m resolution |
| Elevation (m) | DEM at 1m resolution |
| Soil variables | |
| AWC150 (cm) | EUI Map [46] |
| AWC50 (cm) | EUI Map [46] |
| Sub-soil depth (cm) | EUI Map [46] |
| Surface soil depth (cm) | EUI Map [46] |
| Surface-stoniness (%) | EUI Map [46] |
| Topographic wetness index | DEM at 1 m resolution |
| Solar insolation | DEM at 1 m resolution |
| Variables | Min | First Quartile | Median | Mean | Third Quartile | Max |
|---|---|---|---|---|---|---|
| Stocking | 0.21 | 0.52 | 0.61 | 0.60 | 0.70 | 0.79 |
| Seedlings per hectare | 827 | 4097 | 6302 | 7070 | 10,051 | 16,772 |
| AWC150 (cm) | 4.5 | 12.5 | 15.0 | 14.7 | 17.7 | 21.6 |
| Overstory basal area (m2 ha−1) | 5.31 | 12.86 | 14.74 | 14.77 | 16.92 | 25.47 |
| Overstory trees per hectare (TPH) | 61 | 100 | 116 | 124 | 140 | 229 |
| Explanatory Variables | Pseudo R2 | AIC | Precision Parameter (phi) | Coefficients (p-Value) | |
|---|---|---|---|---|---|
| AWC150 (cm) | Overstory Basal Area (m2 ha−1) | ||||
| AWC150 + (Basal area)0.10 | 0.490 | −191.86 | 29.27 | 0.059 (<0.0001) | 11.05 (<0.0001) |
| AWC150 + (Basal area)0.20 | 0.483 | −190.46 | 28.85 | 0.059 (<0.0001) | 4.251 (<0.0001) |
| AWC150 + (Basal area)0.30 | 0.475 | −189.00 | 28.43 | 0.059 (<0.0001) | 2.178 (<0.0001) |
| AWC150 + (Basal area)0.40 | 0.467 | −187.51 | 28.00 | 0.059 (<0.0001) | 1.253 (<0.0001) |
| AWC150 + (Basal area)0.50 | 0.458 | −185.98 | 27.56 | 0.058 (<0.0001) | 0.768 (<0.0001) |
| AWC150+ (Basal area) | 0.413 | −178.13 | 25.45 | 0.057 (<0.0001) | 0.099 (<0.0001) |
| AWC150+ (Basal area)1.50 | 0.365 | −170.47 | 23.54 | 0.055 (<0.0001) | 0.016 (<0.0001) |
| AWC150+ (Basal area)2 | 0.319 | −163.51 | 21.92 | 0.053 (<0.0001) | 0.003 (<0.0001) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nepal, S.; Eskelson, B.N.I.; Ritchie, M.W. Difference in Regeneration Conditions in Pinus ponderosa Dominated Forests in Northern California, USA, over an 83 Year Period. Forests 2020, 11, 581. https://doi.org/10.3390/f11050581
Nepal S, Eskelson BNI, Ritchie MW. Difference in Regeneration Conditions in Pinus ponderosa Dominated Forests in Northern California, USA, over an 83 Year Period. Forests. 2020; 11(5):581. https://doi.org/10.3390/f11050581
Chicago/Turabian StyleNepal, Sushil, Bianca N.I. Eskelson, and Martin W. Ritchie. 2020. "Difference in Regeneration Conditions in Pinus ponderosa Dominated Forests in Northern California, USA, over an 83 Year Period" Forests 11, no. 5: 581. https://doi.org/10.3390/f11050581