Development of Nutrient Uptake by Understory Plant Arrhenatherum elatius and Microbial Biomass during Primary Succession of Forest Soils in Post-Mining Land


Abstract
:1. Introduction
2. Material and Methods
2.1. Soil Sampling and Experiment Setup
2.2. Soil and Plant Analyses
2.3. Statistical Analyses
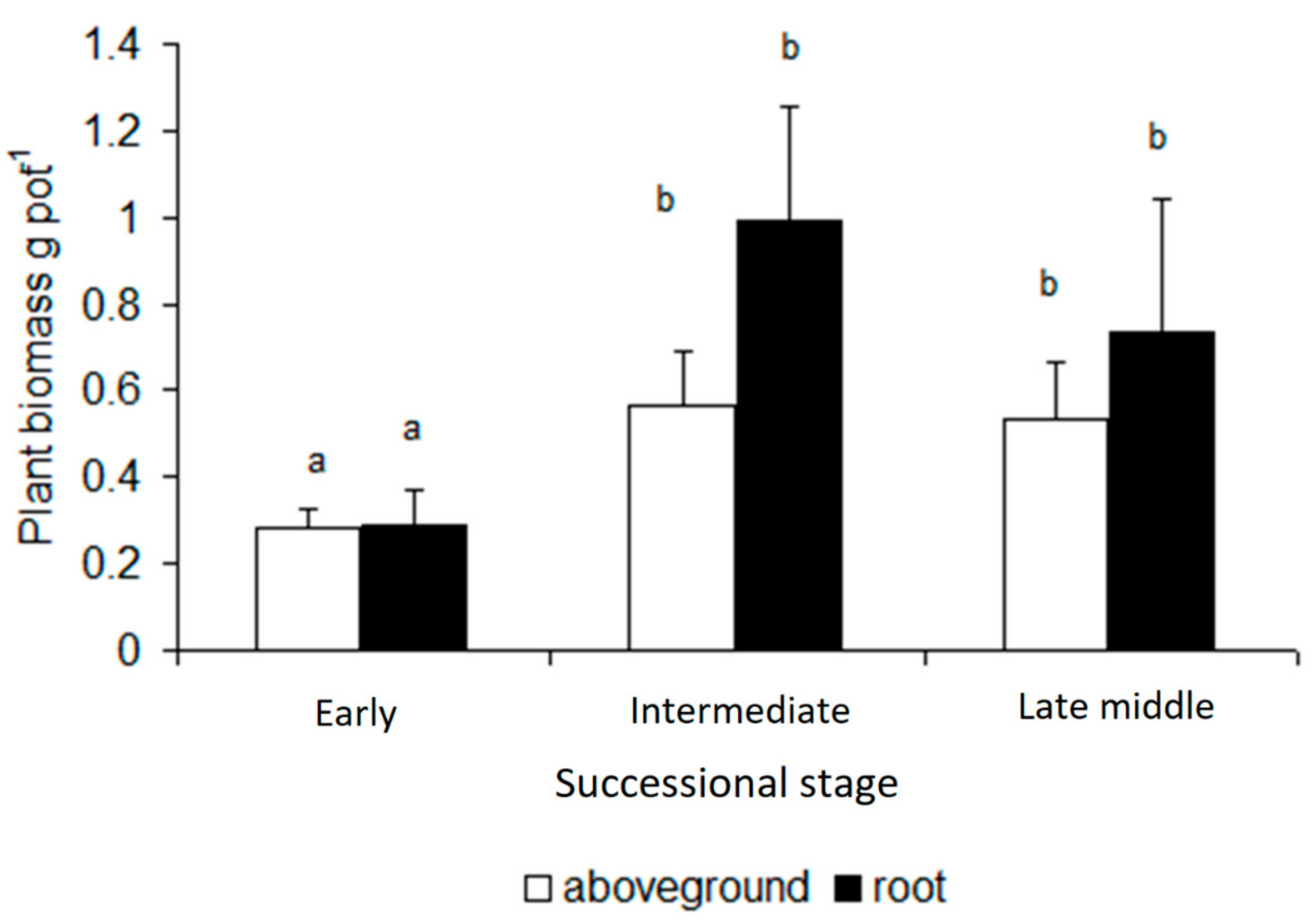
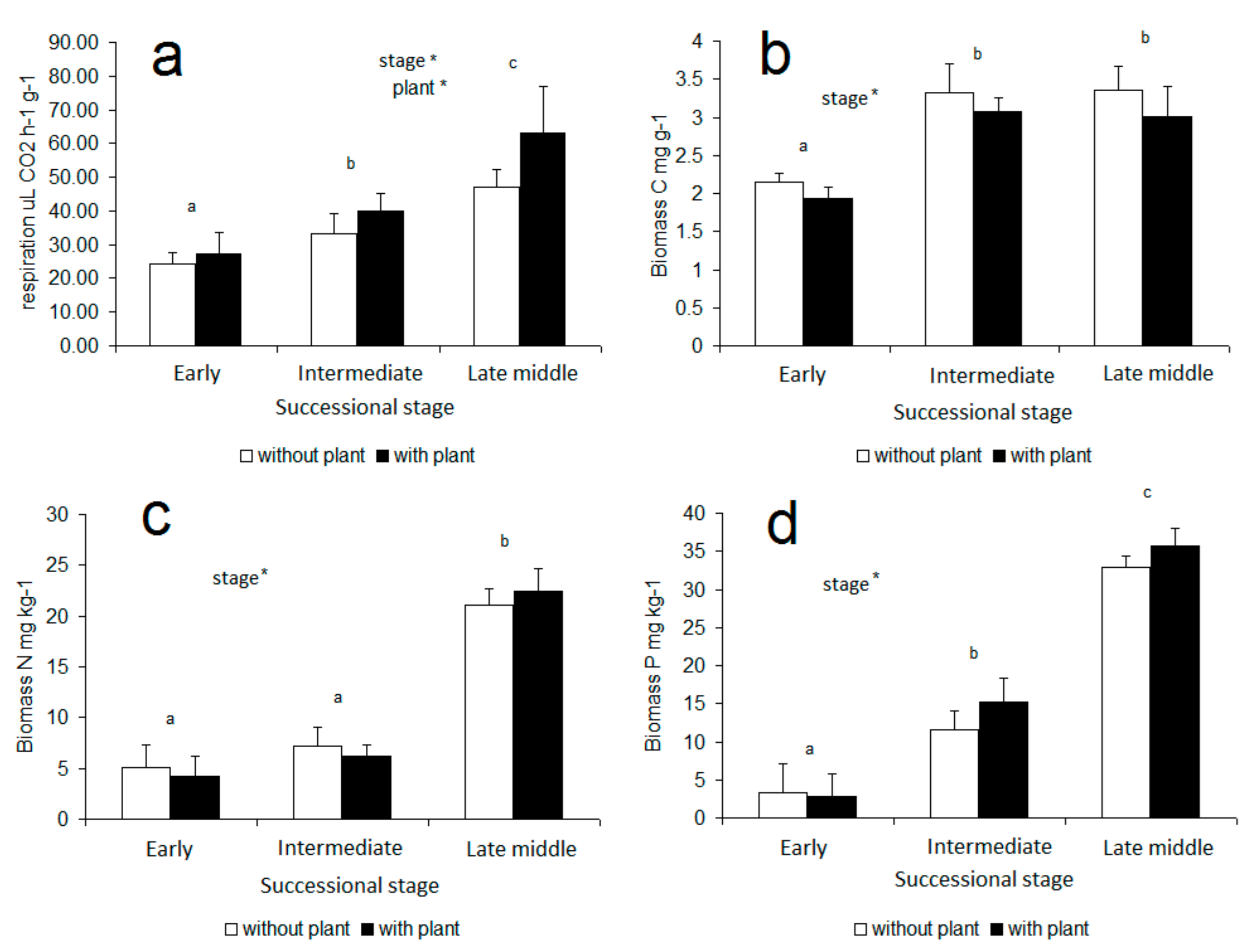
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Macdonald, S.E.; Landhäusser, S.M.; Skousen, J.; Franklin, J.; Frouz, J.; Hall, S.; Jacobs, D.F.; Quideau, S. Forest restoration following surface mining disturbance: Challenges and solutions. New For. 2015, 46, 703–732. [Google Scholar] [CrossRef] [Green Version]
- Frouz, J.; Prach, K.; Pižl, V.; Háněl, L.; Starý, J.; Tajovský, K.; Materna, J.; Balík, V.; Kalčík, J.; Řehounková, K. Interactions between soil development, vegetation and soil fauna during spontaneous succession in post mining sites. Eur. J. Soil Biol. 2008, 44, 109–121. [Google Scholar] [CrossRef]
- Frouz, J.; Dvorščík, P.; Vávrová, A.; Doušová, O.; Kadochová, Š.; Matějíček, L. Development of canopy cover and woody vegetation biomass on reclaimed and unreclaimed post-mining sites. Ecol. Eng. 2015, 84, 233–239. [Google Scholar] [CrossRef]
- Vitousek, P.M.; Matson, P.A.; Van Cleve, K. Nitrogen availability and nitrification during succession: Primary, secondary, and old-field seres. Plant Soil 1989, 115, 229–239. [Google Scholar] [CrossRef]
- Prescott, C.E.; Frouz, J.; Grayston, S.J.; Quideau, S.A.; Straker, J. Rehabilitating forest soils after disturbance. Dev. Soil Sci. 2019, 36, 309–343. [Google Scholar]
- Vitousek, P.M.; Porder, S.; Houlton, B.Z.; Oliver, A.; Vitousek, P.M.; Porder, S.; Houlton, B.Z.; Chadwick, O.A. Terrestrial phosphorus limitation: Mechanisms, implications, and nitrogen—Phosphorus interactions. Ecol. Appl. 2010, 20, 5–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vitousek, P.M.; Farrington, H. Nutrient limitation and soil development: Experimental test of a biogeochemical theory. Biogeochemistry 1997, 37, 63–75. [Google Scholar] [CrossRef]
- Van Der Heijden, M.G.A.; Bardgett, R.D.; Van Straalen, N.M. The unseen majority: Soil microbes as drivers of plant diversity and productivity in terrestrial ecosystems. Ecol. Lett. 2008, 11, 296–310. [Google Scholar] [CrossRef]
- Schimel, J.P.; Bennett, J. Nitrogen mineralization: Challenges of a changing paradigm. Ecology 2004, 85, 591–602. [Google Scholar] [CrossRef]
- Demoling, F.; Figueroa, D.; Bååth, E. Comparison of factors limiting bacterial growth in different soils. Soil Biol. Biochem. 2007, 39, 2485–2495. [Google Scholar] [CrossRef]
- Ekblad, A.; Nordgren, A. Is growth of soil microorganisms in boreal forests limited by carbon or nitrogen availability? Plant Soil 2002, 242, 115–122. [Google Scholar] [CrossRef]
- Spohn, M.; Kuzyakov, Y. Phosphorus mineralization can be driven by microbial need for carbon. Soil Biol. Biochem. 2013, 61, 69–75. [Google Scholar] [CrossRef]
- Čapek, P.; Kotas, P.; Manzoni, S.; Šantrůčková, H. Drivers of phosphorus limitation across soil microbial communities. Funct. Ecol. 2016, 30, 1705–1713. [Google Scholar] [CrossRef] [Green Version]
- Darcy, J.L.; Schmidt, S.K.; Knelman, J.E.; Cleveland, C.C.; Castle, S.C.; Nemergut, D.R. Phosphorus, not nitrogen, limits plants and microbial primary producers following glacial retreat. Sci. Adv. 2018, 4, eaaq0942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hart, S.C.; Stark, J.M. Nitrogen limitation of the microbial biomass in an old-growth forest soil. Écoscience 1997, 4, 91–98. [Google Scholar] [CrossRef]
- Marschner, B.; Kalbitz, K. Controls of bioavailability and biodegradability of dissolved organic matter in soils. Geoderma 2003, 113, 211–235. [Google Scholar] [CrossRef]
- Burger, M.; Jackson, L.E. Microbial immobilization of ammonium and nitrate in relation to ammonification and nitrification rates in organic and conventional cropping systems. Soil Biol. Biochem. 2003, 35, 29–36. [Google Scholar] [CrossRef]
- Richardson, A.E.; Barea, J.M.; McNeill, A.M.; Prigent-Combaret, C. Acquisition of phosphorus and nitrogen in the rhizosphere and plant growth promotion by microorganisms. Plant Soil 2009, 321, 305–339. [Google Scholar] [CrossRef]
- Bartuška, M.; Pawlett, M.; Frouz, J. Particulate organic carbon at reclaimed and unreclaimed post-mining soils and its microbial community composition. CATENA 2015, 131, 92–98. [Google Scholar] [CrossRef]
- Bartuška, M.; Frouz, J. Carbon accumulation and changes in soil chemistry in reclaimed open-cast coal mining heaps near Sokolov using repeated measurement of chronosequence sites. Eur. J. Soil Sci. 2015, 66, 104–111. [Google Scholar] [CrossRef]
- Harantová, L.; Mudrák, O.; Kohout, P.; Elhottová, D.; Frouz, J.; Baldrian, P. Development of microbial community during primary succession in areas degraded by mining activities. Land Degrad. Dev. 2017, 28, 2574–2584. [Google Scholar] [CrossRef]
- Zbíral, J.; Honsa, I.; Malý, S. Soil Analyses, Part III; Czech Central Institute for Supervising and Testing in Agriculture: Brno, Czech, 1997. [Google Scholar]
- Šnajdr, J.; Valášková, V.; Merhautová, V.; Cajthaml, T.; Baldrian, P. Activity and spatial distribution of lignocellulose-degrading enzymes during forest soil colonization by saprotrophic basidiomycetes. Enzyme Microb. Technol. 2008, 43, 186–192. [Google Scholar] [CrossRef]
- Joergensen, R.G. The fumigation-extraction method to estimate soil microbial biomass: Calibration of the kEC value. Soil Biol. Biochem. 1996, 28, 25–31. [Google Scholar] [CrossRef]
- Jonasson, S.; Michelsen, A.; Schmidt, I.K.; Nielsen, E.V.; Terry, V.; Michelsen, A.; Nielsen, E.V.; Callaghan, T.V. Microbial Biomass C, N and P in Two Arctic Soils and Responses to Addition of NPK Fertilizer and Sugar: Implications for Plant Nutrient Uptake. Oecologia 1996, 106, 507–515. [Google Scholar] [CrossRef]
- Brookes, P.C.; Powlson, D.S.; Jenkinson, D.S. Phosphorus in the soil microbial biomass. Soil Biol. Biochem. 1984, 16, 169–175. [Google Scholar] [CrossRef]
- Sommers, L.E.; Nelson, D.W. Determination of Total Phosphorus in Soils: A Rapid Perchloric Acid Digestion Procedure1. Soil Sci. Soc. Am. J. 1972, 36, 902–904. [Google Scholar] [CrossRef]
- Murphy, J.; Riley, J.P. A modified single solution method for the determination of phosphate in natural waters. Anal. Chim. Acta 1962, 27, 31–36. [Google Scholar] [CrossRef]
- Watanabe, F.S.; Olsen, S.R. Test of an Ascorbic Acid Method for Determining Phosphorus in Water and NaHCO3 Extracts from Soil1. Soil Sci. Soc. Am. J. 1965, 29, 677–678. [Google Scholar] [CrossRef]
- Gessner, M.O. Ergosterol as a measure of fungal biomass. In Methods to Study Litter Decomposition: A Practical Guide; Graça, M.A.S., Bärlocher, F., Eds.; Springer: Dordrech, The Netherlands, 2005; pp. 189–195. ISBN 9781402034664. [Google Scholar]
- Šourková, M.; Frouz, J.; Šantrůčková, H. Accumulation of carbon, nitrogen and phosphorus during soil formation on alder spoil heaps after brown-coal mining, near Sokolov (Czech Republic). Geoderma 2005, 124, 203–214. [Google Scholar] [CrossRef]
- Deubel, A.; Merbach, W. Influence of Microorganisms on Phosphorus Bioavailability in Soils. In Microorganisms in Soils: Roles in Genesis and Functions; Springer: Berlin/Heidelberg, Germany, 2005; pp. 177–191. [Google Scholar]
- Frouz, J.; Nováková, A. Development of soil microbial properties in topsoil layer during spontaneous succession in heaps after brown coal mining in relation to humus microstructure development. Geoderma 2005, 129, 54–64. [Google Scholar] [CrossRef]
- Bünemann, E.K.; Smernik, R.J.; Doolette, A.L.; Marschner, P.; Stonor, R.; Wakelin, S.A.; McNeill, A.M. Forms of phosphorus in bacteria and fungi isolated from two Australian soils. Soil Biol. Biochem. 2008, 40, 1908–1915. [Google Scholar] [CrossRef]
- Ezawa, T.; Cavagnaro, T.R.; Smith, S.E.; Smith, F.A.; Ohtomo, R. Rapid accumulation of polyphosphate in extraradical hyphae of an arbuscular mycorrhizal fungus as revealed by histochemistry and a polyphosphate kinase/luciferase system. New Phytol. 2004, 161, 387–392. [Google Scholar] [CrossRef] [Green Version]
- Bolan, N.S. A critical review on the role of mycorrhizal fungi in the uptake of phosphorus by plants. Plant Soil 1991, 134, 189–207. [Google Scholar] [CrossRef]
- Mudrák, O.; Hermová, M.; Tesnerová, C.; Rydlová, J.; Frouz, J. Above-ground and below-ground competition between the willow Salix caprea and its understorey. J. Veg. Sci. 2016, 27, 156–164. [Google Scholar] [CrossRef]
- Knoblochová, T.; Kohout, P.; Püschel, D.; Doubková, P.; Frouz, J.; Cajthaml, T.; Kukla, J.; Vosátka, M.; Rydlová, J. Asymmetric response of root-associated fungal communities of an arbuscular mycorrhizal grass and an ectomycorrhizal tree to their coexistence in primary succession. Mycorrhiza 2017, 27, 775–789. [Google Scholar] [CrossRef] [PubMed]
- Frouz, J.; Moradi, J.; Püschel, D.; Rydlová, J. Earthworms affect growth and competition between ectomycorrhizal and arbuscular mycorrhizal plants. Ecosphere 2019, 10, e02736. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Successional Stage | Early | Intermediate | Late Middle | |||
|---|---|---|---|---|---|---|
| C * (mg g−1) | 2.99 | 7.37 | 6.20 | |||
| N * (mg g−1) | 0.22 | 0.49 | 0.40 | |||
| C:N ratio * | 18.7 | 17.6 | 15.5 | |||
| No plant | Plant | No plant | Plant | No plant | Plant | |
| Ergosterol (ppm) | 0.49 ± 0.12a | 0.50 ± 0.05a | 1.98 ± 0.25b | 2.06 ± 0.22b | 4.11 ± 0.50c | 3.80 ± 0.39c |
| Successional Stage | Early | Intermediate | Late Middle | |||||
|---|---|---|---|---|---|---|---|---|
| Mass of Element in Microbes with Plants | ||||||||
| N | a 5.13 | ± 1.04 a | a 4.12 | ± 0.36 a | b 18.28 | ± 0.81 b | ||
| P | 3.46 | ± 1.75 a | b 10.09 | ± 1.0 9b | c 29.02 | ± 0.98 c | ||
| Mass of element in microbes without plants | ||||||||
| N | a 5.16 | ± 1.22 a | a 4.82 | ± 0.56 a | b 17.73 | ± 0.63 b | ||
| P | 3.99 | ± 2.33 a | a 7.75 | ± 0.81 b | b 26.73 | ± 0.59 c | ||
| Mass of element in plants | ||||||||
| N | b 7.38 | ± 0.53 a | b 12.72 | ± 1.21 b | a 13.30 | ± 1.59 b | ||
| P | 2.80 | ± 0.17 a | c 15.36 | ± 1.41 b | a 14.46 | ± 1.59 b | ||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaneda, S.; Angst, Š.; Frouz, J. Development of Nutrient Uptake by Understory Plant Arrhenatherum elatius and Microbial Biomass during Primary Succession of Forest Soils in Post-Mining Land. Forests 2020, 11, 247. https://doi.org/10.3390/f11020247
Kaneda S, Angst Š, Frouz J. Development of Nutrient Uptake by Understory Plant Arrhenatherum elatius and Microbial Biomass during Primary Succession of Forest Soils in Post-Mining Land. Forests. 2020; 11(2):247. https://doi.org/10.3390/f11020247
Chicago/Turabian StyleKaneda, Satoshi, Šárka Angst, and Jan Frouz. 2020. "Development of Nutrient Uptake by Understory Plant Arrhenatherum elatius and Microbial Biomass during Primary Succession of Forest Soils in Post-Mining Land" Forests 11, no. 2: 247. https://doi.org/10.3390/f11020247