Quantitative Genetic Variation in Bark Stripping of Pinus radiata

 ,
,
Abstract
:1. Introduction
- (1)
- determine the extent to which bark stripping is under additive genetic control and if genetic differences are stable across sites and tree age;
- (2)
- determine the genetic correlation between the level of bark stripping, growth, stem and bark traits;
- (3)
- estimate possible genetic gains in reducing bark stripping damage from the field-based selection of the least damaged families.
2. Materials and Methods
2.1. Family Trials
2.1.1. Assessment of Bark Stripping Damage and Related Traits
2.1.2. Linear Models
2.2. Spatial Analyses
2.3. Estimation of Additive Genetic Variation and Heritability within Sites
2.4. Type B Genetic Correlations
2.5. Phenotypic and Type A Genetic Correlations
2.6. Estimation of Genetic Gain
3. Results
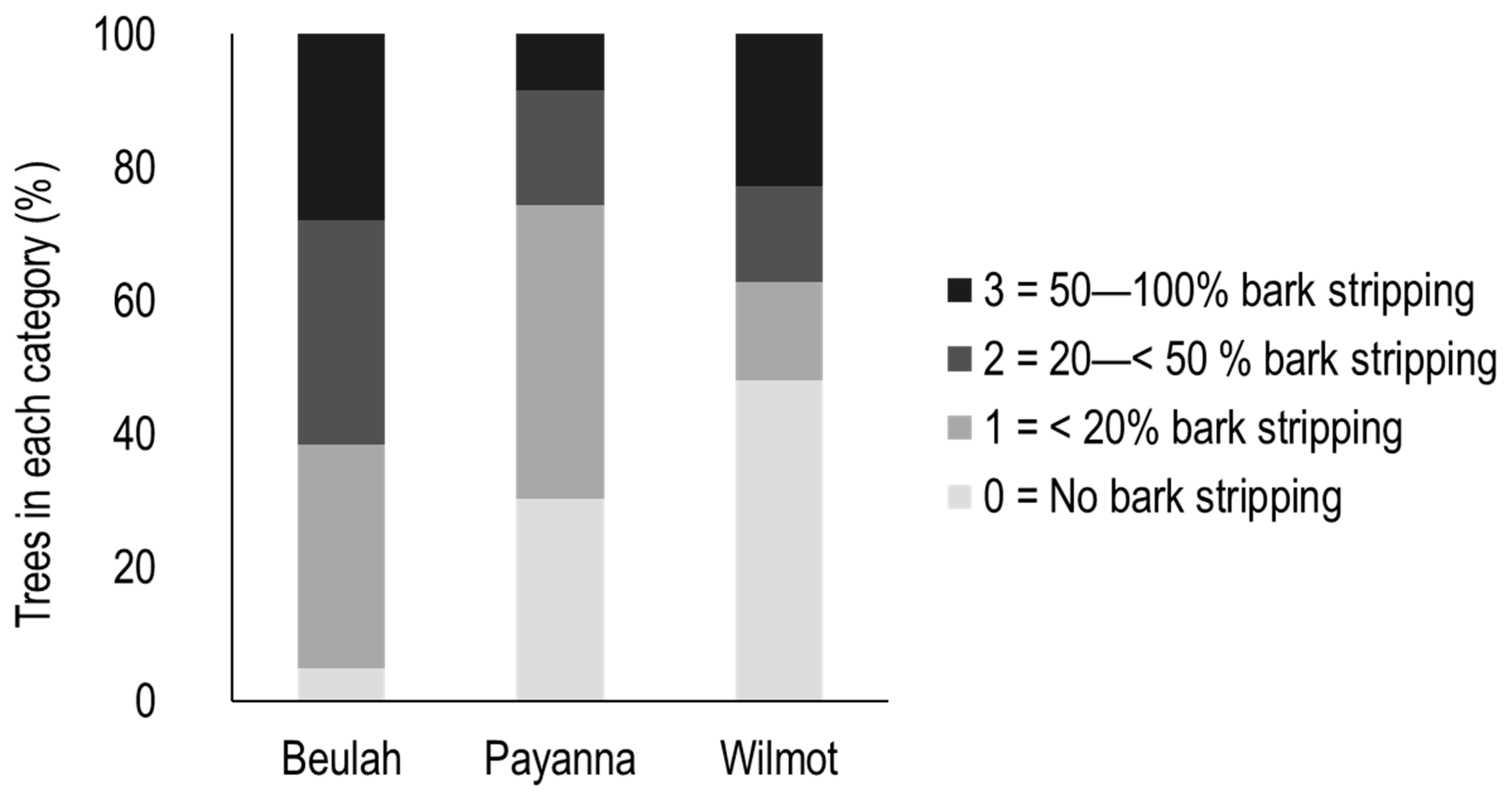
3.1. Differences between Sites in Bark Stripping and Associated Traits
3.2. Spatial Effects
3.3. Additive Genetic Variation for Bark Stripping
3.4. Genetic × Environment Interaction
3.5. Traits Associated with Bark Stripping
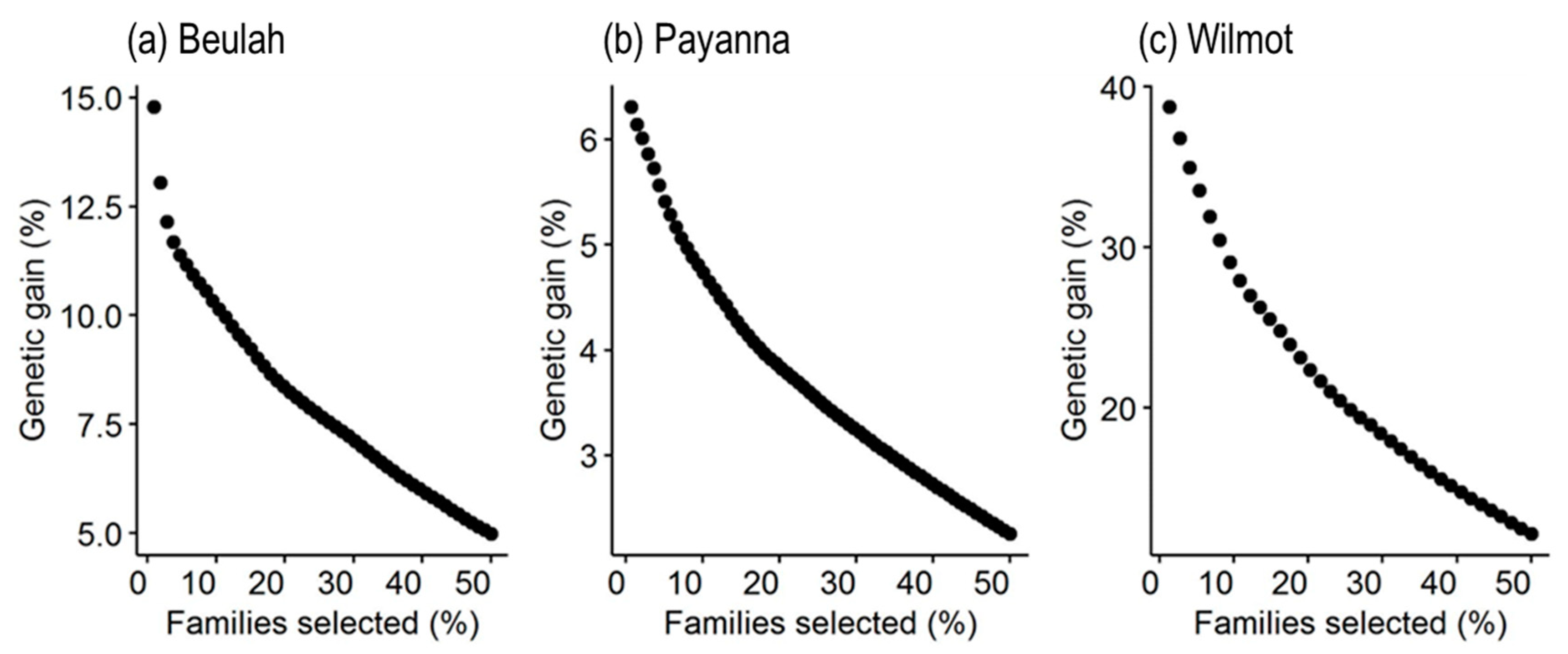
3.6. Estimation of Genetic Gain
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Schowalter, T.D. Insect Ecology: An Ecosystem Approach, 4th ed.; Academic Press: Cambridge, MA, USA, 2006; p. 762. [Google Scholar]
- Ward, A.I.; White, P.C.L.; Smith, A.; Critchley, C.H. Modelling the cost of roe deer browsing damage to forestry. For. Ecol. Manag. 2004, 191, 301–310. [Google Scholar] [CrossRef]
- Kimball, B.A.; Pfund, F.; Gourley, M.; Griffin, D.L.; Russell, J.H. Silvicultural Attempts to Induce Browse Resistance in Conifer Seedlings. Int. J. For. Res. 2011, 2011, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Nichols, C.P.; Drewe, J.A.; Gill, R.; Goode, N.; Gregory, N. A novel causal mechanism for grey squirrel bark stripping: The Calcium Hypothesis. For. Ecol. Manag. 2016, 367, 12–20. [Google Scholar] [CrossRef]
- Telford, A.; Cavers, S.; Ennos, R.A.; Cottrell, J.E. Can we protect forests by harnessing variation in resistance to pests and pathogens? Forestry 2014, 88, 3–12. [Google Scholar] [CrossRef] [Green Version]
- Miller, A.; O’Reilly-Wapstra, J.; Potts, B. Genetic Variation in Bark Stripping among Pinus radiata; National Centre for Future Forest Industries: Hobart, Australia, 2014; p. 30. [Google Scholar]
- Zas, R.; Björklund, N.; Nordlander, G.; Cendán, C.; Hellqvist, C.; Sampedro, L. Exploiting jasmonate-induced responses for field protection of conifer seedlings against a major forest pest, Hylobius abietis. For. Ecol. Manag. 2014, 313, 212–223. [Google Scholar] [CrossRef] [Green Version]
- Ehrlich, P.R.; Raven, P.H. butterflies and plants: A study in coevolution. Evolution 1964, 18, 586–608. [Google Scholar] [CrossRef]
- Maron, J.L.; Agrawal, A.A.; Schemske, D.W. Plant–herbivore coevolution and plant speciation. Ecology 2019, 100, e02704. [Google Scholar] [CrossRef]
- Falconer, D.S.; Mackay, T.F.C. Introduction to Quantitative Genetics, 4th ed.; Prentice Hall: Harlow, UK, 1996; p. 464. [Google Scholar]
- White, T.W.; Adams, W.T.; Neale, D.B. Forest Genetics; CABI Publishing: Cambridge, MA, USA, 2007; p. 704. [Google Scholar]
- Zas, R.; Björklund, N.; Sampedro, L.; Hellqvist, C.; Karlsson, B.; Jansson, S.; Nordlander, G. Genetic variation in resistance of Norway spruce seedlings to damage by the pine weevil Hylobius abietis. Tree Genet. Genomes 2017, 13, 111. [Google Scholar] [CrossRef] [Green Version]
- Zas, R.; Sampedro, L.; Prada, E.; Fernández-López, J. Genetic variation of Pinus pinaster Ait. seedlings in susceptibility to the pine weevil Hylobius abietis L. Ann. For. Sci. 2005, 62, 681–688. [Google Scholar] [CrossRef] [Green Version]
- O’Reilly-Wapstra, J.M.; Iason, G.R.; Fitzsimmons-Thoss, V. The role of genetic and chemical variation of Pinus sylvestris seedlings in influencing slug herbivory. Oecologia 2006, 152, 82–91. [Google Scholar] [CrossRef]
- Wise, M.J.; Rausher, M.D. Evolution of resistance to a multiple-herbivore community: Genetic correlations, diffuse coevolution, and constraints on the plant’s response to selection. Evolution 2013, 67, 1767–1779. [Google Scholar] [CrossRef] [PubMed]
- O’Reilly-Wapstra, J.M.; Potts, B.M.; McArthur, C.; Davies, N.W. Effects of nutrient variability on the genetic-based resistance of Eucalyptus globulus to a mammalian herbivore and on plant defensive chemistry. Oecologia 2004, 142, 597–605. [Google Scholar] [CrossRef] [PubMed]
- Klápště, J.; Meason, D.F.; Dungey, H.S.; Telfer, E.J.; Silcock, P.; Rapley, S. Genotype-by-environment interaction in coast redwood outside natural distribution—Search for environmental cues. BMC Genet. 2020, 21, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Dungey, H.S.; Carson, M.; Carson, S. Genotype by environment interaction for growth and Dothistroma resistance and clonal connectivity between environments in radiata pine in New Zealand and Australia. PLoS ONE 2018, 13, e0205402. [Google Scholar] [CrossRef]
- Sampedro, L.; Moreira, X.; Zas, R. Costs of constitutive and herbivore-induced chemical defences in pine trees emerge only under low nutrient availability. J. Ecol. 2011, 99, 818–827. [Google Scholar] [CrossRef] [Green Version]
- Barton, K.E.; Koricheva, J. The Ontogeny of Plant Defense and Herbivory: Characterizing General Patterns Using Meta-Analysis. Am. Nat. 2010, 175, 481–493. [Google Scholar] [CrossRef] [Green Version]
- Lawrence, R.; Potts, B.M.; Whitham, T.G. Relative importance of plant ontogeny, host genetic variation, and leaf age for a common herbivore. Ecology 2003, 84, 1171–1178. [Google Scholar] [CrossRef]
- Russell, J. Deployment of deer-resistant western redcedar (Thuja plicata). In National Proceedings: Forest and Conservation Nursery Associations—2007; Dumroese, R.K., Riley, L.E., Eds.; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2008; pp. 149–153. [Google Scholar]
- Moreira, X.; Zas, R.; Sampedro, L. Additive genetic variation in resistance traits of an exotic pine species: Little evidence for constraints on evolution of resistance against native herbivores. Heredity 2013, 110, 449–456. [Google Scholar] [CrossRef] [Green Version]
- Raffa, K.F.; Smalley, E.B. Interaction of pre-attack and induced monoterpene concentrations in host conifer defense against bark beetle-fungal complexes. Oecologia 1995, 102, 285–295. [Google Scholar] [CrossRef]
- Wainhouse, D.; Staley, J.T.; Jinks, R.; Morgan, G. Growth and defence in young pine and spruce and the expression of resistance to a stem-feeding weevil. Oecologia 2008, 158, 641–650. [Google Scholar] [CrossRef]
- Alfaro, R.I.; King, J.N.; Brown, R.; Buddingh, S.M. Screening of Sitka spruce genotypes for resistance to the white pine weevil using artificial infestations. For. Ecol. Manag. 2008, 255, 1749–1758. [Google Scholar] [CrossRef]
- Dimock, E.J., II; Silen, R.R.; Allen, V.E. Genetic resistance in Douglas-fir to damage by snowshoe hare and black-tailed deer. For. Sci. 1976, 22, 106–121. [Google Scholar]
- Duncan, A.J.; Hartley, S.E.; Thurlow, M.; Young, S.; Staines, B.W. Clonal variation in monoterpene concentrations in Sitka spruce (Picea sitchensis) saplings and its effect on their susceptibility to browsing damage by red deer (Cervus elaphus). For. Ecol. Manag. 2001, 148, 259–269. [Google Scholar] [CrossRef]
- Silen, R.R.; Randall, W.K.; Mandel, N.L. Estimates of genetic parameters for deer browsing of Douglas-fir. For. Sci. 1986, 32, 178–184. [Google Scholar]
- Cukor, J.; Vacek, Z.; Linda, R.; Vacek, S.; Marada, P.; Šimůnek, V.; Havránek, F. Effects of Bark Stripping on Timber Production and Structure of Norway Spruce Forests in Relation to Climatic Factors. Forests 2019, 10, 320. [Google Scholar] [CrossRef] [Green Version]
- Iijima, H.; Nagaike, T. Susceptible conditions for debarking by deer in subalpine coniferous forests in central Japan. For. Ecosyst. 2015, 2, 33. [Google Scholar] [CrossRef] [Green Version]
- Welch, D.; Scott, D. Bark-stripping damage by red deer in a Sitka spruce forest in western Scotland IV. Survival and performance of wounded trees. Forestry 1998, 71, 225–235. [Google Scholar] [CrossRef] [Green Version]
- Moreira, X.; Zas, R.; Sampedro, L. Differential Allocation of Constitutive and Induced Chemical Defenses in Pine Tree Juveniles: A Test of the Optimal Defense Theory. PLoS ONE 2012, 7, e34006. [Google Scholar] [CrossRef] [Green Version]
- Moreira, X.; Lundborg, L.; Zas, R.; Carrillo-Gavilán, A.; Borg-Karlson, A.-K.; Sampedro, L. Inducibility of chemical defences by two chewing insect herbivores in pine trees is specific to targeted plant tissue, particular herbivore and defensive trait. Phytochemistry 2013, 94, 113–122. [Google Scholar] [CrossRef] [Green Version]
- Iason, G.R.; O’Reilly-Wapstra, J.M.; Brewer, M.J.; Summers, R.W.; Moore, B.D. Do multiple herbivores maintain chemical diversity of Scots pine monoterpenes? Philos. Trans. R. Soc. B Biol. Sci. 2011, 366, 1337–1345. [Google Scholar] [CrossRef]
- Eldridge, K.G. Seed Collections of Pinus radiata and P. muricata in California. In Forest Genetic Resources Information; FAO: Canberra, Australia, 1979; pp. 44–45. [Google Scholar]
- Mead, D.J. Sustainable Management of Pinus radiata Plantations; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2013; p. 246. [Google Scholar]
- ABARES. Australia’s State of the Forests Report; Australian Government Department of Agriculture and Water Resources: Canberra, Australia, 2018; p. 56. [Google Scholar]
- Wu, H.X.; Eldridge, K.G.; Matheson, A.C.; Powell, M.P.; McRae, T.A. Successful introduction and breeding of radiata pine to Australia. In Growing Forest Values, Proceedings of ANZIF 2007 Conference, Coffs Harbour, NSW, Australia, 3–7 June 2007; Institute of Foresters of Australia and New Zealand Institute of Forestry: Canberra, Australia, 2007; pp. 506–517. [Google Scholar]
- Dungey, H.S.; Brawner, J.T.; Burger, F.; Carson, M.; Henson, M.; Jefferson, P.; Matheson, A.C. A New Breeding Strategy for Pinus radiata in New Zealand and New South Wales. Silvae Genet. 2009, 58, 28–38. [Google Scholar] [CrossRef] [Green Version]
- Sasse, J.; Elms, S.; Kube, P. Genetic resistance in Pinus radiata to defoliation by the pine aphid Essigella californica. Aust. For. 2009, 72, 25–31. [Google Scholar] [CrossRef]
- Ivković, M.; Baltunis, B.; Gapare, W.; Sasse, J.; Dutkowski, G.; Elms, S.; Wu, H. Breeding against dothistroma needle blight of radiata pine in Australia. Can. J. For. Res. 2010, 40, 1653–1660. [Google Scholar] [CrossRef]
- Graham, N.J.; Suontama, M.; Pleasants, T.; Li, Y.; Bader, M.K.-F.; Klápště, J.; Dungey, H.S.; Williams, N.M. Assessing the genetic variation of tolerance to red needle cast in a Pinus radiata breeding population. Tree Genet. Genomes 2018, 14, 55. [Google Scholar] [CrossRef] [Green Version]
- Suontama, M.; Li, Y.; Low, C.B.; Dungey, H.S. Genetic improvement of resistance to cyclaneusma needle cast in Pinus radiata. Can. J. For. Res. 2019, 49, 128–133. [Google Scholar] [CrossRef]
- Downham, R.; Gavran, M. Australian Plantation Statistics 2018 Update; Australian Government Department of Agriculture and Water Resources: Canberra, Australia, 2018. [Google Scholar]
- Page, D.E.; Close, D.C.; Beadle, C.L.; Wardlaw, T.J.; Mohammed, C. Seasonal dynamics in understorey abundance and carbohydrate concentration in relation to browsing and bark stripping of Tasmanian Pinus radiata plantations. For. Ecol. Manag. 2013, 296, 98–107. [Google Scholar] [CrossRef]
- Department of Primary Industries Parks Water and Environment. A Planning Toolkit for Managing Browsing and Grazing Losses from Wallabies and Brushtail Possums on Farms in Tasmania; Tasmanian Government: Launceston, Tasmania, 2018. [Google Scholar]
- Bannister, M.H. An early progeny trial in Pinus radiata 1. Juvenile characters. N. Z. J. Bot. 1969, 7, 316–335. [Google Scholar] [CrossRef]
- Smith, A.H.; Ratkowsky, D.A.; Wardlaw, T.J.; Mohammed, C. Ease of Access to An Alternative Food Source Enables Wallabies to Strip Bark in Tasmanian Pinus radiata Plantations. Forests 2020, 11, 387. [Google Scholar] [CrossRef] [Green Version]
- Department of Primary Industries Parks Water and Environment. Annual State-Wide Spotlight Surveys, Tasmania 2018/19: Regional Summary: Priority Harvested Species; Tasmanian Government: Habart, Australia, 2019. [Google Scholar]
- Butler, D.G.; Cullis, B.R.; Gilmour, A.R.; Gogel, B.J. ASReml-R Reference Manual; Queensland Government Department of Primary Industries and Fisheries: Brisbane, Australia, 2009; p. 149. [Google Scholar]
- Costa e Silva, J.; Dutkowski, G.W.; Gilmour, A.R. Analysis of Early Tree Height in Forest Genetic Trials Is Enhanced by Including a Spatially Correlated Residual. Can. J. For. Res. 2001, 31, 1887–1893. [Google Scholar] [CrossRef]
- Dungey, H.S.; Williams, N.M.; Low, C.B.; Stovold, G.T. First evidence of genetic-based tolerance to red needle cast caused by Phytophthora pluvialis in radiata pine. N. Z. J. For. Sci. 2014, 44, 31. [Google Scholar] [CrossRef] [Green Version]
- Isik, F.; Holland, J.; Maltecca, C. Genetic Data Analysis for Plant and Animal Breeding; Springer International: Cham, Switzerland, 2007; p. 400. [Google Scholar]
- Zas, R.; Solla, A.; Sampedro, L. Variography and kriging allow screening Pinus pinaster resistant to Armillaria ostoyae in field conditions. Forestry 2007, 80, 201–209. [Google Scholar] [CrossRef] [Green Version]
- Cappa, E.P.; Muñoz, F.; Sanchez, L. Performance of alternative spatial models in empirical Douglas-fir and simulated datasets. Ann. For. Sci. 2019, 76, 53. [Google Scholar] [CrossRef]
- Dutkowski, G.W.; Silva, J.C.E.; Gilmour, A.R.; Wellendorf, H.; Aguiar, A. Spatial analysis enhances modelling of a wide variety of traits in forest genetic trials. Can. J. For. Res. 2006, 36, 1851–1870. [Google Scholar] [CrossRef]
- Dungey, H.S.; Russell, J.H.; Silva, J.C.E.; Low, C.B.; Miller, M.A.; Fleet, K.R.; Stovold, G.T. The effectiveness of cloning for the genetic improvement of Mexican white cypress Cupressus lusitanica (Mill.). Tree Genet. Genomes 2012, 9, 443–453. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Haskard, K.A.; Cullis, B.R.; Verbyla, A.P. Anisotropic matern correlation and spatial prediction using REML. J. Agric. Biol. Environ. Stat. 2007, 2, 147–160. [Google Scholar] [CrossRef] [Green Version]
- Burnham, K.P.; Anderson, D.R. Multimodel inference: Understanding AIC and BIC in model selection. Sociol. Methods Res. 2004, 33, 261–304. [Google Scholar] [CrossRef]
- Burdon, R.D. Genetic correlation as a concept for studying genotype-environment interaction in forest tree breeding. Silvae Genet. 1977, 26, 168–175. [Google Scholar]
- Yamada, Y. Genotype by environment interaction and genetic correlation of the same trait under different environments. Jpn. J. Genet. 1962, 37, 498–509. [Google Scholar] [CrossRef] [Green Version]
- Olausson, A.; Rönningen, K. Estimation of Genetic Parameters for Threshold Characters. Acta Agric. Scand. 1975, 25, 201–208. [Google Scholar] [CrossRef]
- De la Mata, R.; Hood, S.M.; Sala, A. Insect outbreak shifts the direction of selection from fast to slow growth rates in the long-lived conifer Pinus ponderosa. Proc. Natl. Acad. Sci. USA 2017, 114, 7391–7396. [Google Scholar] [CrossRef] [Green Version]
- Yanchuk, A.; Murphy, J.C.; Wallin, K.F. Evaluation of genetic variation of attack and resistance in lodgepole pine in the early stages of a mountain pine beetle outbreak. Tree Genet. Genomes 2008, 4, 171–180. [Google Scholar] [CrossRef]
- King, J.N.; Yanchuk, A.D.; Kiss, G.K.; Alfaro, R.I. Genetic and phenotypic relationships between weevil (Pissodes strobi) resistance and height growth in spruce populations of British Columbia. Can. J. For. Res. 1997, 27, 732–739. [Google Scholar] [CrossRef]
- King, J.N.; Alfaro, R.I. Developing Sitka Spruce Populations for Resistance to the White Pine Weevil: Summary of Research and Breeding Program; Ministry of Forests and Range: Victoria, BC, Canada, 2009. [Google Scholar]
- Dieters, M.J.; Hodge, G.R.; White, T.L. Genetic parameter estimates for resistance to rust (Cronartium quercuum) infection from full-sib tests of slash pine (Pinus elliottii), modelled as functions of rust incidence. Silvae Genet. 1996, 45, 235–242. [Google Scholar]
- Mottet, M.-J.; Deblois, J.; Perron, M. High genetic variation and moderate to high values for genetic parameters of Picea abies resistance to Pissodes strobi. Tree Genet. Genomes 2015, 11, 58. [Google Scholar] [CrossRef]
- Murphy, G.E.; Cown, D. Within-tree, between-tree, and geospatial variation in estimated Pinus radiata bark volume and weight in New Zealand. N. Z. J. For. Sci. 2015, 45, 18. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Z.; Ueda, H.; Kitahara, M.; Imaki, H. Bark stripping by sika deer on veitch fir related to stand age, bark nutrition, and season in northern Mount Fuji district, central Japan. J. For. Res. 2005, 10, 359–365. [Google Scholar] [CrossRef]
- Nopp-Mayr, U.; Reimoser, F.; Voelk, F. Predisposition assessment of mountainous forests to bark peeling by red deer (Cervus elaphus L.) as a strategy in preventive forest habitat management. Wildl. Biol. Pract. 2011, 7, 66–89. [Google Scholar] [CrossRef]
- Månsson, J.; Jarnemo, A. Bark-stripping on Norway spruce by red deer in Sweden: Level of damage and relation to tree characteristics. Scand. J. For. Res. 2013, 28, 117–125. [Google Scholar] [CrossRef]
- Kuiters, A.T.; Van Der Sluijs, L.; Wytema, G. Selective bark-stripping of beech, Fagus sylvatica, by free-ranging horses. For. Ecol. Manag. 2006, 222, 1–8. [Google Scholar] [CrossRef]
- King, J.N.; Alfaro, R.I.; Lopez, M.G.; Van Akker, L. Resistance of Sitka spruce (Picea sitchensis (Bong.) Carr.) to white pine weevil (Pissodes strobi Peck): Characterizing the bark defence mechanisms of resistant populations. Forestry 2011, 84, 83–91. [Google Scholar] [CrossRef]
- Van Akker, L.; Alfaro, R.I.; Brockley, R. Effects of fertilization on resin canal defences and incidence of Pissodes strobi attack in interior spruce. Can. J. For. Res. 2004, 34, 855–862. [Google Scholar] [CrossRef] [Green Version]
- Ferrenberg, S.; Mitton, J.B. Smooth bark surfaces can defend trees against insect attack: Resurrecting a ‘slippery’ hypothesis. Funct. Ecol. 2014, 28, 837–845. [Google Scholar] [CrossRef]
- Zas, R.; Sampedro, L.; Moreira, X.; Martíns, P. Effect of fertilization and genetic variation on susceptibility of Pinus radiata seedlings to Hylobius abietis damage. Can. J. For. Res. 2008, 38, 63–72. [Google Scholar] [CrossRef] [Green Version]
- Mayle, B.A.; Proudfoot, J.; Poole, J. Influence of tree size and dominance on incidence of bark stripping by grey squirrels to oak and impact on tree growth. Forestry 2009, 82, 431–444. [Google Scholar] [CrossRef]
- Kenward, R.E.; Dutton, J.C.F.; Parish, T.; Doyle, F.I.B.; Walls, S.S.; Robertson, P.A. Damage by grey squirrels. I. Bark-stripping correlates and treatment. Q. J. For. 1996, 90, 135–142. [Google Scholar]
- Ferrenberg, S.; Kane, J.M.; Langenhan, J.M. To grow or defend? Pine seedlings grow less but induce more defences when a key resource is limited. Tree Physiol. 2015, 35, 107–111. [Google Scholar] [CrossRef] [Green Version]
- Akashi, N.; Terazawa, K. Bark stripping damage to conifer plantations in relation to the abundance of sika deer in Hokkaido, Japan. For. Ecol. Manag. 2005, 208, 77–83. [Google Scholar] [CrossRef]
- Gianoli, E.; Salgado-Luarte, C. Tolerance to herbivory and the resource availability hypothesis. Biol. Lett. 2017, 13, 20170120. [Google Scholar] [CrossRef] [Green Version]
- O’Reilly-Wapstra, J.M.; Moore, B.D.; Brewer, M.J.; Beaton, J.; Sim, D.; Wiggins, N.L.; Iason, G.R. Pinus sylvestris sapling growth and recovery from mammalian browsing. For. Ecol. Manag. 2014, 325, 18–25. [Google Scholar] [CrossRef]
- Zas, R.; Moreira, X.; Sampedro, L. Tolerance and induced resistance in a native and an exotic pine species: Relevant traits for invasion ecology. J. Ecol. 2011, 99, 1316–1326. [Google Scholar] [CrossRef] [Green Version]
- Lush, J.L. Family Merit and Individual Merit as Bases for Selection. Part I. Am. Nat. 1947, 81, 241–261. [Google Scholar] [CrossRef]
- Barbosa, P.; Hines, J.; Kaplan, I.; Martinson, H.; Szczepaniec, A.; Szendrei, Z. Associational Resistance and Associational Susceptibility: Having Right or Wrong Neighbors. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Watt, M.S.; Whitehead, D.; Mason, E.G.; Richardson, B.; Kimberley, M.O. The influence of weed competition for light and water on growth and dry matter partitioning of young Pinus radiata, at a dryland site. For. Ecol. Manag. 2003, 183, 363–376. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genetic Trial | Latitude (°) | Longitude (°) | Elevation (m) | Date Planted | Replicates | Incomplete Blocks | Families | Parents | Grand Parents | Number of Trees Assessed | Time of First Assessment (Age Years) | Time of Second Assessment (Age Years) | Time of Third Assessment (Age Years) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Beulah | 41°.29′50.22° S | 146°.25′12.43° E | 336 | 2011 | 25 | 75 | 101 (OP) | 101 | 194 | 2002 | 2015 (4 years) | 2016 (5 years) | |
| Payanna | 41°.06′47.29° S | 147°.43′12.19° E | 170 | 2011 | 20 | 80 | 138 (OP) | 138 | 195 | 2668 | 2015 (4 years) | 2016 (5 years) | |
| Wilmot | 41°.27′14.83° S | 146°.06′35.30° E | 580 | 2015 | 26 | 78 | 74 (CP) | 55 | 54 | 1372 | 2017 (2 years) | 2018 (3 years) | 2020 (5 years) |
| Trait | Year Assessed | Age of Trees (Years) | Number of Trees Assessed | Mean | Minimum | Maximum | SD | h2 (se) | Significance of Additive Genetic Variation | Significance of Spatial Model |
|---|---|---|---|---|---|---|---|---|---|---|
| Bark stripping (%) | 2015 | 4 | 1957 | 36.44 | 0.00 | 75.00 | 26.89 | 0.14 ± 0.05 | <0.001 | <0.001 |
| Bark stripping (%) | 2016 | 5 | 2002 | 40.08 | 0.00 | 75.00 | 26.55 | 0.13 ± 0.04 | <0.001 | <0.001 |
| Height (cm) | 2015 | 4 | 2032 | 391.18 | 40.00 | 707.00 | 89.38 | 0.18 ± 0.05 | <0.001 | <0.001 |
| Height (cm) | 2016 | 5 | 2002 | 473.92 | 1.00 | 807.00 | 117.29 | 0.13 ± 0.05 | <0.001 | <0.001 |
| DBH (cm) | 2016 | 5 | 1975 | 8.67 | 1.00 | 14.90 | 2.46 | 0.09 ± 0.04 | <0.05 | <0.001 |
| Bark thickness (mm) | 2016 | 5 | 1355 | 6.67 | 2.50 | 13.50 | 1.26 | 0.10 ± 0.03 | <0.01 | <0.001 |
| Rough bark * | 2016 | 5 | 2002 | 0.42 | 0.00 | 1.00 | 0.49 | 0.09 ± 0.03 | −46.82 + | −72.14 + |
| Rough bark height (cm) | 2016 | 5 | 847 | 47.88 | 2.00 | 420.00 | 63.21 | 1.00 ± 0.06 | <0.001 | 0.372 |
| Survival | 2016 | 5 | 2699 | 0.72 | 0.00 | 1.00 | 0.45 | NE | NE | NE |
| Trait | Year Assessed | Age (Years) | Sample Size | Mean | Minimum | Maximum | SD | h2 (se) | Significance of Additive Genetic Variation | Significance of Spatial Model |
|---|---|---|---|---|---|---|---|---|---|---|
| Bark stripping (%) | 2015 | 4 | 2647 | 8.21 | 0.00 | 75.00 | 15.99 | 0.07 ± 0.03 | <0.05 | 0.270 |
| Bark stripping (%) | 2016 | 5 | 2668 | 16.77 | 0.00 | 75.00 | 21.21 | 0.14 ± 0.04 | <0.001 | <0.001 |
| Height (cm) | 2015 | 4 | 2648 | 573.92 | 130.00 | 827.00 | 94.06 | 0.10 ± 0.04 | <0.001 | <0.001 |
| DBH (cm) | 2016 | 5 | 1727 | 7.90 | 3.00 | 19.50 | 6.48 | 0.03 ± 0.00 | 0.205 | <0.001 |
| Bark thickness (mm) | 2016 | 5 | 1727 | 6.01 | 0.00 | 16.00 | 1.78 | 0.11 ± 0.04 | <0.010 | <0.001 |
| Rough bark * | 2016 | 5 | 1694 | 0.77 | 0.00 | 1.00 | 0.42 | 0.07 ± 0.04 | −29.90 + | −35.12 + |
| Rough bark height (cm) | 2016 | 5 | 1327 | 93.54 | 5.00 | 350.00 | 60.26 | 0.53 ± 0.10 | <0.001 | <0.050 |
| Survival | 2016 | 5 | 2760 | 0.97 | 0.00 | 1.00 | 0.18 | NE | NE | NE |
| Trait | Year Assessed | Age (years) | Sample Size | Mean | Minimum | Maximum | SD | h2 (se) | Significance of Additive Genetic Variation | Significance of SCA Effect | Significance of Spatial Model |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Bark stripping (%) | 2017 | 2 | 1372 | 23.96 | 0.00 | 100.00 | 33.18 | 0.09 ± 0.03 | <0.001 | >0.05 | <0.001 |
| Bark stripping (%) | 2018 | 3 | 1269 | 22.50 | 0.00 | 100.00 | 25.10 | 0.06 ± 0.03 | <0.05 | >0.05 | <0.001 |
| Bark strip height (cm) | 2017 | 2 | 706 | 2.19 | 1.00 | 7.00 | 1.45 | 0.12 ± 0.04 | <0.001 | >0.05 | <0.001 |
| Height (cm) | 2017 | 2 | 1372 | 147.40 | 10.00 | 248.00 | 33.51 | 0.07 ± 0.04 | <0.001 | >0.05 | <0.001 |
| Height (cm) | 2018 | 3 | 1275 | 231.09 | 30.00 | 382.00 | 51.17 | 0.08 ± 0.04 | <0.001 | >0.05 | <0.001 |
| Height (cm) | 2020 | 5 | 1230 | 544.33 | 40.00 | 780.00 | 99.08 | 0.11 ± 0.04 | <0.001 | >0.05 | <0.001 |
| Basal diameter (cm) | 2017 | 2 | 140 | 2.99 | 1.00 | 5.30 | 0.78 | 0.04 ± 0.05 | >0.05 | >0.05 | <0.001 |
| DBH (cm) | 2020 | 5 | 1230 | 103.40 | 5.00 | 190.00 | 42.97 | 0.03 ± 0.02 | <0.01 | >0.05 | <0.001 |
| Stem access | 2017 | 2 | 1371 | 49.49 | 0.00 | 100.00 | 26.26 | 0.09 ± 0.03 | <0.05 | >0.05 | <0.001 |
| Survival | 2017 | 2 | 1372 | 0.86 | 0.00 | 1.00 | 0.40 | NE | NE | NE | NE |
| Trait | ra | se(ra) | χ2[ra = 0] | P[ra = 0] | P[ra = 1] |
|---|---|---|---|---|---|
| Bark stripping (year 4) | 0.23 | 0.39 | 0.44 | >0.05 | |
| Bark stripping (year 5) | 0.76 | 0.25 | 10.00 | <0.01 | <0.01 |
| Height (year 4) | 0.91 | 0.32 | 14.50 | <0.001 | <0.001 |
| Rough bark (year 5) | 0.74 | 0.42 | 23.80 | −10.72 + | −22.53 + |
| Rough bark height (year 5) | 0.25 | 0.21 | 1.50 | >0.05 | |
| Bark thickness (year 5) | 0.53 | 0.85 | 2.70 | >0.05 |
| Bark Stripping (Year 4) | Bark Stripping (Year 5) | Height (Year 4) | Height (Year 5) | Bark Thickness (Year 5) | DBH (Year 5) | Rough Bark (Year 5) | Rough Bark Height (Year 5) | |
|---|---|---|---|---|---|---|---|---|
| Bark stripping (year 4) | 0.42 (0.02) *** | −0.26 (0.02) *** | −0.29 (0.02) *** | −0.18 (0.02) *** | −0.26 (0.02) *** | −0.08 (0.02) *** | −0.17 (0.03) *** | |
| Bark stripping (year 5) | 0.78 (0.16) *** | −0.32 (0.02) *** | −0.40 (0.02) *** | −0.25 (0.02) *** | −0.42 (0.02) *** | −0.25 (0.02) *** | −0.25 (0.03) *** | |
| Height (year 4) | −0.19 (0.25) | −0.09 (0.02) | 0.86 (0.01) *** | 0.49 (0.02) *** | 0.79 (0.01) *** | 0.31 (0.02) *** | 0.13 (0.03) *** | |
| Height (year 5) | −0.09 (0.27) | 0.33 (0.34) | 0.98 (0.04) *** | 0.50 (0.02) *** | 0.80 (0.01) *** | 0.33 (0.02) *** | 0.10 (0.03) ** | |
| Bark thickness (year 5) | −0.24 (0.28) | −0.37 (0.29) | 0.24 (0.25) | 0.03 (0.33) | 0.57 (0.02) *** | 0.34 (0.02) *** | 0.34 (0.03) *** | |
| DBH (year 5) | −0.26 (0.02) | −0.04 (0.37) | 0.78 (0.12) * | 0.73 (0.15) | 0.40 (0.30) | 0.33 (0.02) *** | 0.17 (0.03) *** | |
| Rough bark (year 5) | −0.27 (0.22) | −0.25 (0.22) | 0.12 (0.21) | −0.05 (0.24) | 0.31 (0.23) | 0.21 (0.27) | NA | |
| Rough bark height (year 5) | −0.39 (0.27) | −0.52 (0.26) | 0.02 (0.01) | 0.02 (0.018) | 0.10 (0.02) | 0.37 (0.09) * | NA |
| Bark Stripping (Year 4) | Bark Stripping (Year 5) | Height (Year 4) | Bark Thickness (Year 5) | DBH (Year 5) | Rough Bark (Year 5) | Rough Bark Height (Year 5) | |
|---|---|---|---|---|---|---|---|
| Bark stripping (year 4) | 0.34 (0.02) *** | −0.30 (0.02) *** | −0.18 (0.02) *** | 0.01 (0.02) | −0.12 (0.02) *** | 0.00 (0.02) | |
| Bark stripping (year 5) | 0.91 (0.23) ** | −0.28 (0.02) *** | −0.21 (0.02) *** | −0.11 (0.02) *** | −0.29 (0.02) *** | −0.17 (0.03) *** | |
| Height (year 4) | −0.12 (0.34) | −0.20 (0.24) | 0.36 (0.02) *** | 0.22 (0.06) *** | 0.23 (0.02) *** | 0.19 (0.03) *** | |
| Bark thickness (year 5) | −0.34 (0.51) | −0.48 (0.22) * | 0.24 (0.27) | −0.02 (0.02) | 0.26 (0.02) *** | 0.29 (0.03) *** | |
| DBH (year 5) | −0.34 (0.33) | −0.40 (0.34) | 0.89 (0.27) ** | 0.56 (0.42) | 0.02 (0.02) | 0.32 (0.03) *** | |
| Rough bark (year 5) | 0.05 (0.31) | −0.47 (0.19) * | −0.41 (0.28) | 0.52 (0.25) | −0.58 (0.57) | NA | |
| Rough bark height (year 5) | −0.16 (0.25) | −0.37 (0.17) * | 0.05 (0.21) | 0.70 (0.17) ** | 0.11 (0.37) | NA |
| Bark Stripping (Year 2) | Bark Stripping (Year 3) | Height (Year 2) | Height (Year 3) | Height (Year 5) | Diameter at 10 cm (Year 2) | DBH (Year 5) | Stem Access (Year 2) | Bark Strip Height (Year 2) | ΔHt2-3 | ΔHt3-5 | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Bark stripping (year 2) | 0.71 (0.02) *** | 040 (0.03) *** | −0.24 (0.03) *** | 0.43 (0.04) *** | −0.23 (0.08) ** | −0.54 (0.02) *** | 0.41 (0.02) *** | 0.89(0.02) *** | −0.46 (0.02) *** | 0.45 (0.04) *** | |
| Bark stripping (year 3) | 1.00 (0.04) ** | −0.82 (0.31) * | −0.24 (0.03) *** | 0.50 (0.04) *** | −0.29 (0.55) | −0.29 (0.03) *** | 1.00 (0.09) *** | 0.62 (0.02) *** | −0.39 (0.03) *** | 0.52 (0.04) *** | |
| Height (year 2) | 0.56 (0.32) | 0.00 (0.03) | 0.80 (0.02) *** | 0.53 (0.02) *** | 0.70 (0.06) *** | 0.49 (0.02) *** | −0.11 (0.03) *** | 0.10 (0.03) *** | 0.59 (0.03) *** | 0.48 (0.03) *** | |
| Height (year 3) | 0.12 (0.) | 0.34 (0.41) | 0.79 (0.17) * | 0.69 (0.18) *** | 0.65 (0.07) *** | 0.81 (0.01) *** | 0.40 (0.38) | −0.17 (0.03) | 0.75 (0.01) *** | 0.63 (0.03) *** | |
| Height (year 5) | 0.42 (0.26) | 0.38 (0.32) | 0.45 (0.28) | 0.77 (0.16) ** | −0.82 (1.60) | 0.44 (0.30) | 0.49 (0.27) | 0.53 (0.23) | 0.49 (0.02) *** | 0.89 (0.01) *** | |
| Diameter at 10 cm (year 2) | −0.05 (0.57) | −0.27 (0.07) * | 0.22 (0.55) | −0.20 (0.62) | 0.47 (0.05) *** | 0.72 (0.02) *** | −0.20 (0.55) | 0.03 (0.56) | −0.12 (0.08) *** | 0.34 (0.06) *** | |
| DBH (year 5) | −0.84 (0.16) ** | −0.74 (0.24) * | −0.44 (0.42) | 0.49 (0.29) | 0.90 (0.01) *** | 0.75 (0.29) | −0.66 (0.24) | −0.88 (0.15) ** | 0.62 (0.02) *** | 0.72 (0.01) *** | |
| Stem access (year 2) | 0.95 (0.10) *** | 0.36 (0.03) *** | −0.76 (0.25) * | −0.28 (0.03) *** | 0.50 (0.04) *** | −0.24 (0.08) ** | −0.42 (0.03) *** | 0.42 (0.02) *** | −0.34 (0.03) *** | 0.50 (0.04) *** | |
| Bark strip height (year 2) | 0.97 (0.02) *** | 0.98 (0.02) *** | 0.66 (0.28) | 0.27 (0.33) | 0.34 (0.03) *** | −0.13 (0.08) | −0.47 (0.02) *** | 0.87 (0.13) *** | −0.44 (0.03) *** | 0.35 (0.04) *** | |
| ΔHt2−3 | −0.35 (0.27) | −0.20 (0.35) | 0.38 (0.33) | 0.89 (0.10) ** | 0.65 (0.22) * | −0.15 (0.59) | 0.68 (0.23) | −0.18 (0.33) | −0.25 (0.29) | 0.53 (0.03) *** | |
| ΔHt3−5 | 0.47 (0.27) | 0.38 (0.31) | 0.33 (0.34) | 0.53 (0.27) | 0.94 (0.04) *** | −0.99 (0.76) | 0.19 (0.37) | 0.47 (0.27) | 0.55 (0.22) | 0.44 (0.29) |
| Covariates Added to Model 1 | LRT χ2 [Va > 0] | p-Value [Va > 0] | h2 ± se |
|---|---|---|---|
| Payanna | |||
| y = Bark stripping (year 5), covariates =bark thickness (year 5), rough bark height (year 5) and rough bark | 101.8 | <0.001 | 0.12 ± 0.02 |
| Wilmot | |||
| y = Bark stripping (year 2), covariate = stem access (year 2) | 15.6 | <0.001 | 0.06 ± 0.02 |
| y = Bark stripping (year 3), covariates = height (year 2) and stem access (year 2) | 228.0 | <0.001 | 0.03 ± 0.02 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nantongo, J.S.; Potts, B.M.; Fitzgerald, H.; Newman, J.; Elms, S.; Aurik, D.; Dungey, H.; O’Reilly-Wapstra, J.M. Quantitative Genetic Variation in Bark Stripping of Pinus radiata. Forests 2020, 11, 1356. https://doi.org/10.3390/f11121356
Nantongo JS, Potts BM, Fitzgerald H, Newman J, Elms S, Aurik D, Dungey H, O’Reilly-Wapstra JM. Quantitative Genetic Variation in Bark Stripping of Pinus radiata. Forests. 2020; 11(12):1356. https://doi.org/10.3390/f11121356
Chicago/Turabian StyleNantongo, Judith S., Brad M. Potts, Hugh Fitzgerald, Jessica Newman, Stephen Elms, Don Aurik, Heidi Dungey, and Julianne M. O’Reilly-Wapstra. 2020. "Quantitative Genetic Variation in Bark Stripping of Pinus radiata" Forests 11, no. 12: 1356. https://doi.org/10.3390/f11121356