Post-Fire Recovery of Vegetation and Diversity Patterns in Semiarid Pinus halepensis Mill. Habitats after Salvage Logging

 ,
,  , ,
, ,  and
and
Abstract
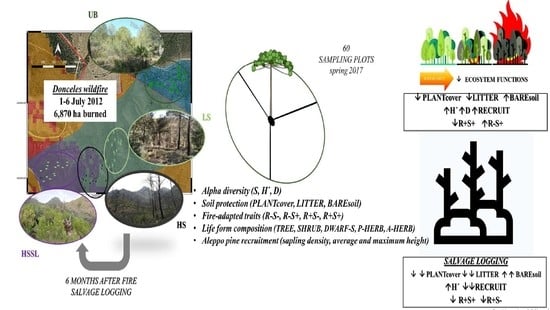
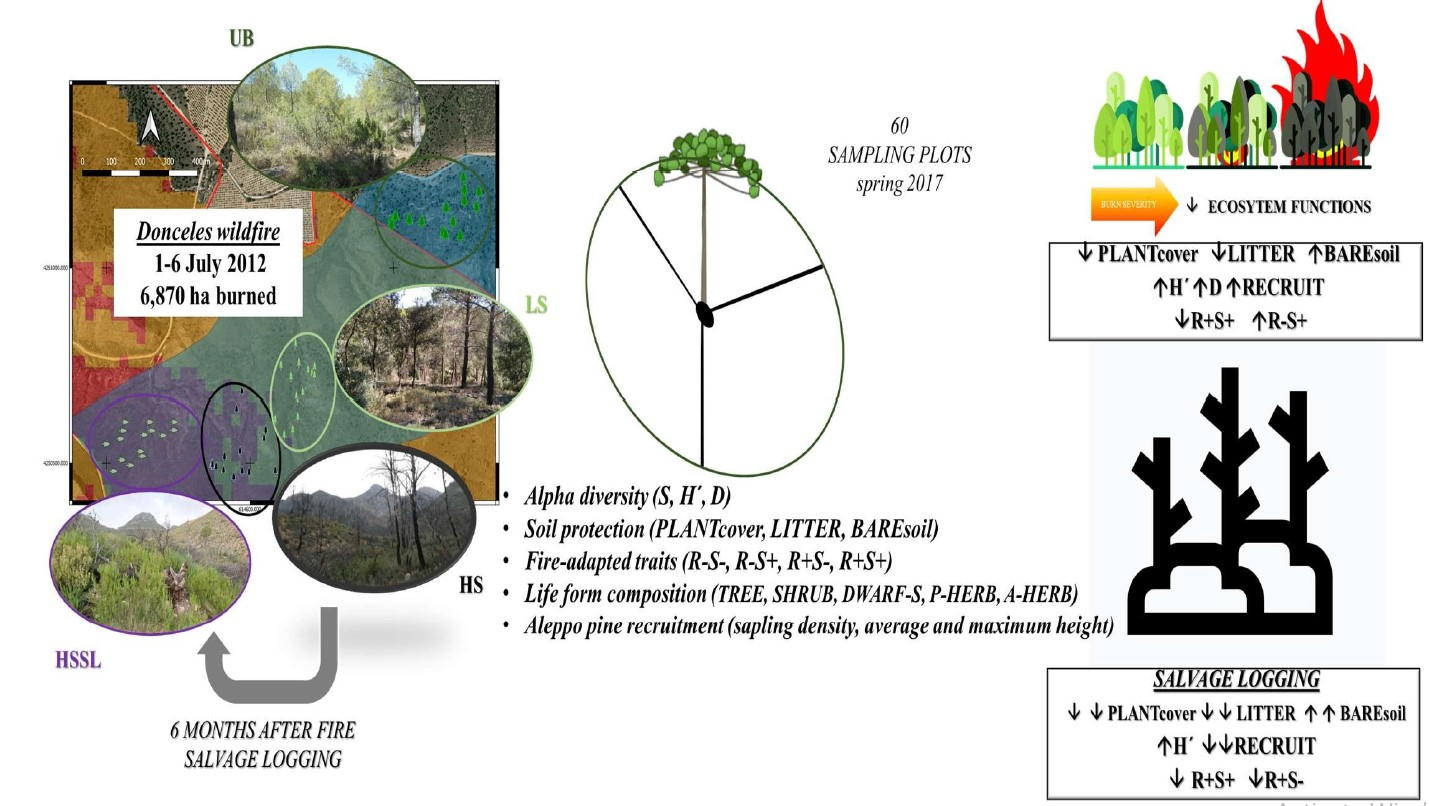
:
1. Introduction
2. Materials and Methods
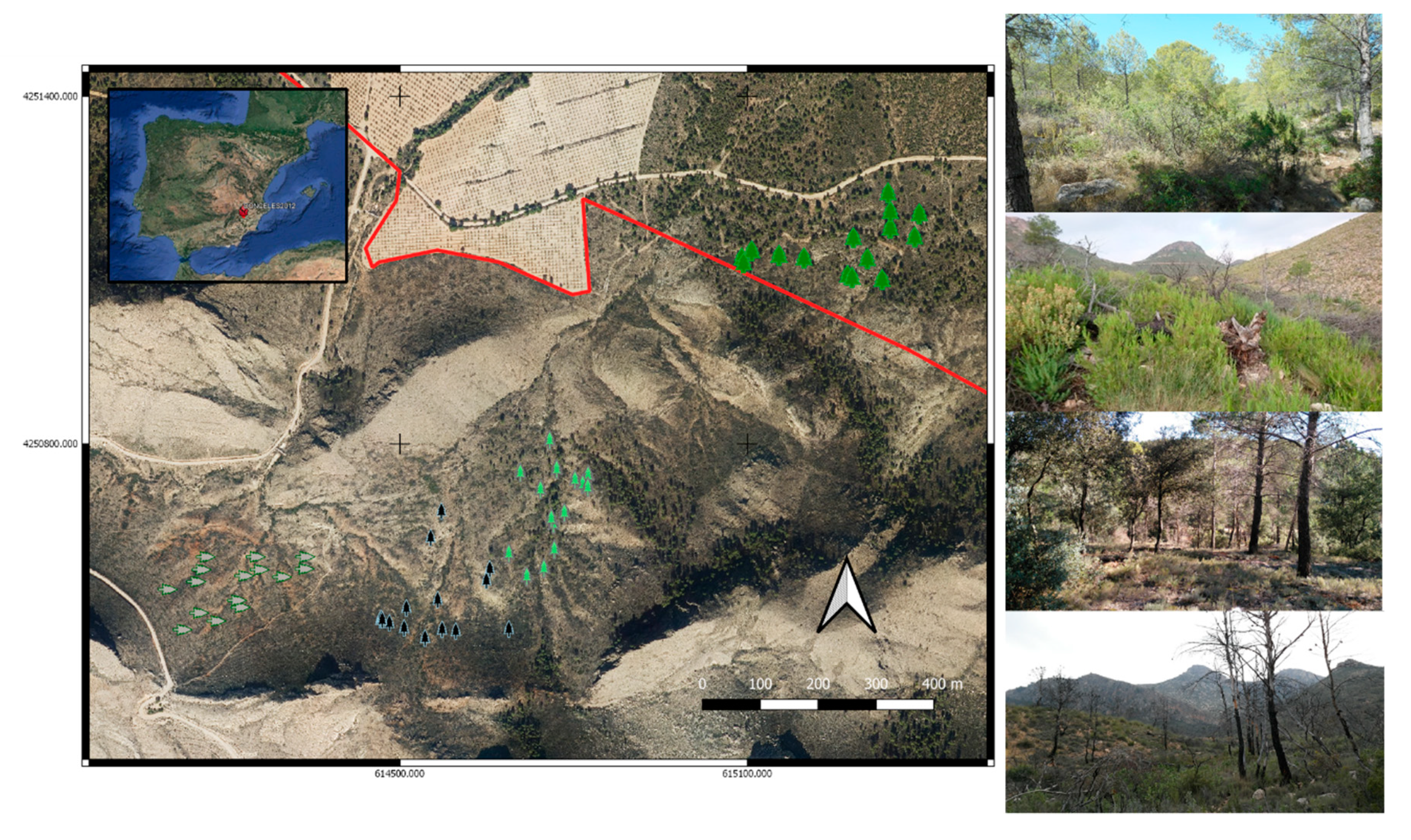
2.1. Study Area
2.2. Burn Severity and Post-Fire Management
2.3. Experimental Design and Field Sampling
2.4. Post-Fire Plant Community
2.4.1. Plant Diversity
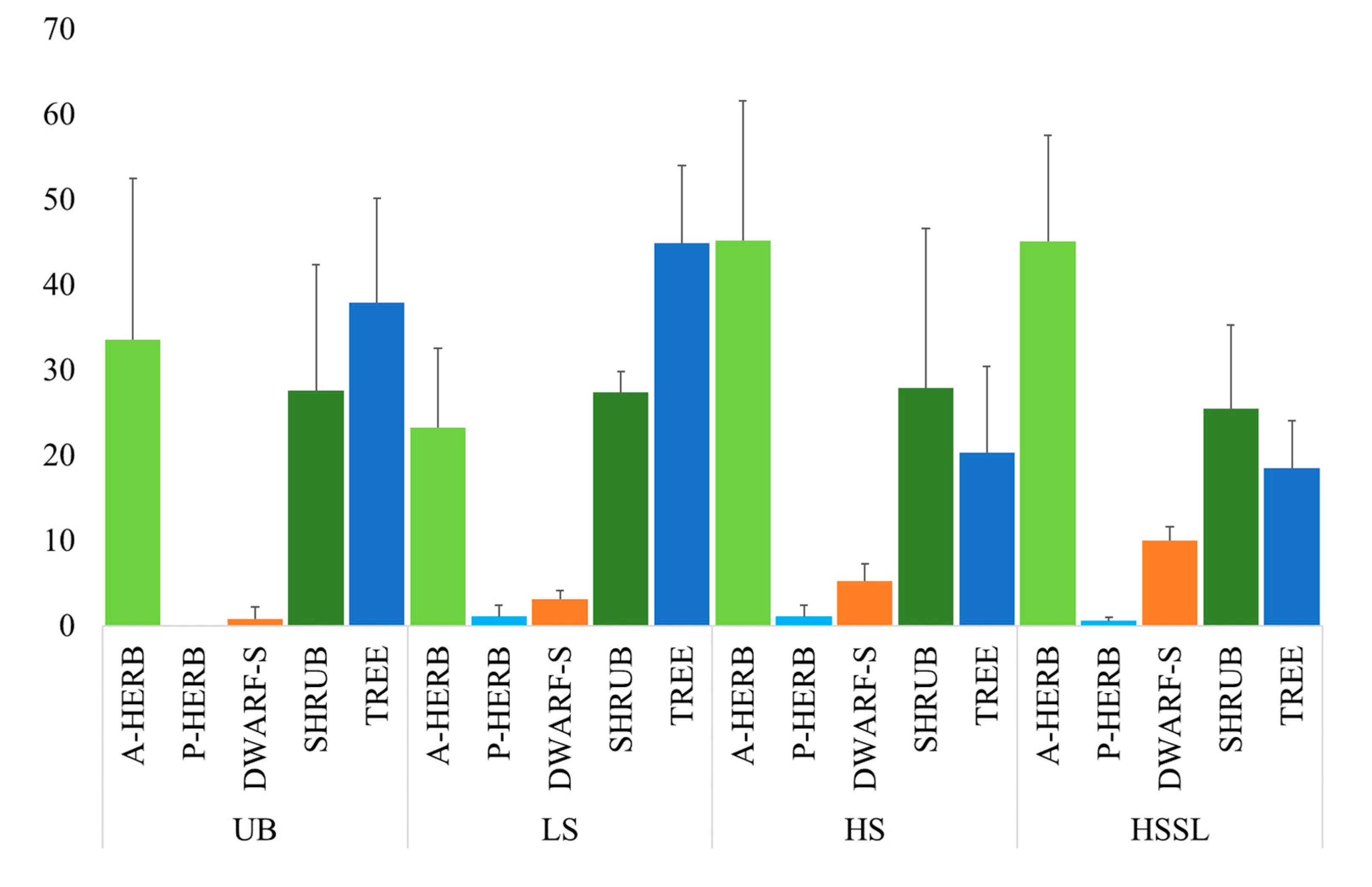
2.4.2. Fire-Adapted Traits and Life Forms
2.4.3. Pine Recruitment
2.5. Statistical Analyses
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pausas, J.G.; Keeley, J.E. Wildfires as an ecosystem service. Front. Ecol. Environ. 2019, 17, 289–295. [Google Scholar] [CrossRef] [Green Version]
- Pausas, J.G.; Fernández-Muñoz, S. Fire regime changes in the Western Mediterranean Basin: From fuel-limited to drought-driven fire regime. Clim. Chang. 2012, 110, 215–226. [Google Scholar] [CrossRef] [Green Version]
- Dale, V.H.; Joyce, L.A.; McNulty, S.; Neilson, R.P.; Ayres, M.P.; Flannigan, M.D.; Wotton, B.M. Climate Change and Forest Disturbances. Bioscience 2001, 51, 723–734. [Google Scholar] [CrossRef] [Green Version]
- Certini, G. Fire as a Soil-Forming Factor. AMBIO 2014, 43, 191–195. [Google Scholar] [CrossRef] [Green Version]
- Doblas-Miranda, E.; Alonso, R.; Arnan, X.; Bermejo, V.; Brotons, L.; de las Heras, J.; Estiarte, M.; Hódar, J.A.; Llorens, P.; Lloret, F.; et al. A review of the combination among global change factors in forests, shrublands and pastures of the Mediterranean. Glob. Planet. Chang. 2017, 148, 42–54. [Google Scholar] [CrossRef] [Green Version]
- Ibáñez, I.; Acharya, K.; Juno, E.; Karounos, C.; Lee, B.R.; McCollum, C.; Schaffer-Morrison, S.; Tourville, J. Forest resilience under global environmental change: Do we have the information we need? A systematic review. PLoS ONE 2019, 14, e0222207. [Google Scholar] [CrossRef] [PubMed]
- Newton, A.C.; Cantarello, E. Restoration of forest resilience: An achievable goal? New For. 2015, 46, 645–668. [Google Scholar] [CrossRef]
- Keeley, J.E. Fire intensity, fire severity and burn severity: A brief review and suggested usage. Int. J. Wildland Fire 2009, 18, 116–126. [Google Scholar] [CrossRef]
- López-Poma, R.; Bautista, S. Plant regeneration functional groups modulate the response to fire of soil enzyme activities in a Mediterranean shrubland. Soil Biol. Biochem. 2014, 79, 5–13. [Google Scholar] [CrossRef]
- González-De Vega, S.; de las Heras, J.; Moya, D. Post-Fire Regeneration and Diversity Response to Burn Severity in Pinus halepensis Mill. forests. Forests 2018, 9, 299. [Google Scholar] [CrossRef] [Green Version]
- Eugenio, M.; Verkaik, I.; Lloret, F.; Espelta, J. Recruitment and growth decline in Pinus halepensis populations after recurrent wildfires in Catalonia (NE Iberian Peninsula). For. Ecol. Manag. 2006, 231, 47–54. [Google Scholar]
- González-De Vega, S.; De las Heras, J.; Moya, D. Resilience of Mediterranean terrestrial ecosystems and fire severity in semiarid areas: Responses of Aleppo pine forests in the short, mid and long term. Sci. Total Environ. 2016, 573, 1171–1177. [Google Scholar] [CrossRef] [PubMed]
- Viana-Soto, A.; Aguado, I.; Martínez, S. Assessment of Post-Fire Vegetation Recovery Using Fire Severity and Geographical Data in the Mediterranean Region (Spain). Environments 2017, 4, 90. [Google Scholar] [CrossRef] [Green Version]
- Brown, J.K.; Smith, J.K. Wildland fire in ecosystems: Effects of fire on flora. In General Technical Reports Rocky Mountin Research Station; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Ogden, UT, USA, 2000; Volume 42, 257p. [Google Scholar]
- Santin, C.; Doerr, S.H. Fire effects on soils: The human dimension. Philos. Trans. R. Soc. Biol. Sci. 2016, 371, 20150171. [Google Scholar] [CrossRef] [Green Version]
- Smith, J.K. Wildland fire in ecosystems: Effects of fire on fauna. In General Technical Reports Rocky Mountin Research Station; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Ogden, UT, USA, 2000; Volume 42, 83p. [Google Scholar]
- Lee, C.; Schlemme, C.; Murray, J.; Unsworth, R. The cost of climate change: Ecosystem services and wildland fires. Ecol. Econ. 2015, 116, 261–269. [Google Scholar] [CrossRef] [Green Version]
- Kelly, L.T.; Brotons, L. Using fire to promote biodiversity. Science 2017, 355, 8–9. [Google Scholar] [CrossRef]
- Serrano-Ortiz, P.; Marañón-Jiménez, S.; Reverter, B.R.; Sánchez-Cañete, E.P.; Castro, J.; Zamora, R.; Kowalski, A.S. Post-fire salvage logging reduces carbon sequestration in Mediterranean coniferous forest. For. Ecol. Manag. 2011, 262, 2287–2296. [Google Scholar] [CrossRef]
- Lucas-Borja, M.E.; González-Romero, J.; Plaza-Álvarez, P.A.; Sagra, J.; Gómez, M.E.; Moya, D.; Cerdà, A.; de las Heras, J. The impact of straw mulching and salvage logging on post-fire runoff and soil erosion generation under Mediterranean climate conditions. Sci. Total Environ. 2019, 654, 441–451. [Google Scholar] [CrossRef]
- Tapias, R.; Gil, L.; Fuentes-Utrilla, P.; Pardos, J.A. Canopy seed banks in Mediterranean pines of southeastern Spain: A comparison between Pinus halepensis Mill.; P. pinaster Ait.; P. nigra Arn. and P. pinea L. J. Ecol. 2001, 89, 629–638. [Google Scholar] [CrossRef]
- Daskalakou, E.; Costas, A.T. Aleppo pine (Pinus halepensis) Postfire Regeneration: The Role of Canopy and Soil Seed Banks. Int. J. Wildland Fire 1996, 6, 59–66. [Google Scholar] [CrossRef]
- Goubitz, S.; Nathan, R.; Roitemberg, R.; Shmida, A.; Ne’eman, G. Canopy seed bank structure in relation to: Fire, tree size and density. Plant. Ecol. 2004, 173, 191–201. [Google Scholar] [CrossRef]
- Moya, D.; De las Heras, J.; López-Serrano, F.R.; Leone, V. Optimal intensity and age of management in young Aleppo pine stands for post-fire resilience. For. Ecol. Manag. 2008, 255, 3270–3280. [Google Scholar] [CrossRef]
- Saracino, A.; Bellino, A.; Allevato, E.; Mingo, A.; Conti, S.; Rossi, S.; Bonanomi, G.; Carputo, D.; Mazzoleni, S. Repeated Stand-Replacing Crown Fires Affect Seed Morphology and Germination in Aleppo pine. Front. Plant Sci. 2017, 8, 1160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarris, D.; Christodoulakis, D.; Körner, C. Impact of recent climatic change on growth of low elevation eastern Mediterranean forest trees. Clim. Chang. 2011, 106, 203–223. [Google Scholar] [CrossRef]
- Nathan, R.; Ne’eman, G. Serotiny seed dispersal and seed predation in Pinus halepensis. In Ecology, Biogeography and Management of Pinus halepensis and P. brutia Forest Ecosystems in the Mediterranean Basin; Ne’eman, G., Trabaud, L., Eds.; Backhuys Publishers: Leiden, The Netherlands, 2000; pp. 105–118. [Google Scholar]
- Hernandez-Serrano, A.; Verdu, M.; Gonzalez-Martinez, S.C.; Pausas, J.G. Fire structures pine Serotiny at different scales. Am. J. Bot. 2013, 100, 2349–2356. [Google Scholar] [CrossRef]
- Alfaro-Sanchez, R.; Camarero, J.J.; Lopez-Serrano, F.R.; Sanchez-Salguero, R.; Moya, D.; de las Heras, J. Positive coupling between growth and reproduction in young post-fire Aleppo pines depends on climate and site conditions. Int. J. Wildland Fire 2015, 24, 507–517. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Sánchez, J.J.; Ferrandis, P.; de las Heras, J.; Herranz, J.M. Effect of burnt wood removal on the natural regeneration of Pinus halepensis after fire in a pine forest in Tus valley (SE Spain). For. Ecol. Manag. 1999, 123, 1–10. [Google Scholar] [CrossRef]
- Moya, D.; de las Heras, J.; López-Serrano, F.R.; Ferrandis, P. Post-Fire Seedling Recruitment and Morpho-Ecophysiological Responses to Induced Drought and Salvage Logging in Pinus halepensis Mill. stands. Forests 2015, 6, 1858–1877. [Google Scholar] [CrossRef] [Green Version]
- Castro, J.; Allen, C.D.; Molina-Morales, M.; Marañón-Jiménez, S.; Sánchez-Miranda, A.; Zamora, R. Salvage Logging Versus the Use of Burnt Wood as a Nurse Object to Promote Post-Fire Tree Seedling Establishment. Restor. Ecol. 2011, 19, 537–544. [Google Scholar] [CrossRef]
- Marañón-Jiménez, S.; Castro, J.; Kowalski, A.S.; Serrano-Ortiz, P.; Reverter, B.R.; Sánchez-Cañete, E.P.; Zamora, R. Post-fire soil respiration in relation to burnt wood management in a Mediterranean mountain ecosystem. For. Ecol. Manag. 2011, 261, 1436–1447. [Google Scholar] [CrossRef]
- Puig-Gironès, R.; Imbeau, L.; Clavero, M.; Rost, J.; Pons, P. Does post-fire salvage logging affect foraging activity by rodents? Eur. J. For. Res. 2020, 139, 777–790. [Google Scholar] [CrossRef]
- Marcolin, E.; Marzano, R.; Vitali, A.; Garbarino, M.; Lingua, E. Post-Fire Management Impact on Natural Forest Regeneration through Altered Microsite Conditions. Forests 2019, 10, 1014. [Google Scholar] [CrossRef]
- Taboada, A.; Fernández-García, V.; Marcos, E.; Calvo, L. Interactions between large high-severity fires and salvage logging on a short return interval reduce the regrowth of fire-prone serotinous forests. For. Ecol. Manag. 2018, 414, 54–63. [Google Scholar] [CrossRef]
- DeRose, R.J.; Long, J.N. Resistance and Resilience: A Conceptual Framework for Silviculture. For. Sci. 2014, 60, 1205–1212. [Google Scholar] [CrossRef]
- Spanish Geographic Institute. Centro de Descargas del Centro Nacional de Información Geográfica, Spain. 2020. Available online: http://centrodedescargas.cnig.es/CentroDescargas/index.jsp (accessed on 16 December 2020).
- Kottek, M.; Grieser, J.; Beck, C.; Rudolf, B.; Rubel, F. World Map of the Köppen-Geiger climate classification updated. Meteorol. Z. 2006, 15, 259–263. [Google Scholar] [CrossRef]
- García-Morote, F.A.; Martínez-García, E.; Andrés-Abellán, M.; Caballero, E.R.; Miettinen, H.; López-Serrano, F.R. Direct seeding of Pinus halepensis Mill. for recovery of burned semi-arid forests: Implications for post-fire management for improving natural regeneration. Forests 2017, 8, 353. [Google Scholar] [CrossRef]
- Soil Survey Staff. Keys to Soil Taxonomy, 12th ed.; USDA-Natural Resources Conservation Service: Washington, DC, USA, 2014.
- Rivas-Martínez, S. Memoria del Mapa de Series de Vegetación de España 1: 400.000; ICONA, Ministerio de Agricultura, Pesca y Alimentación: Madrid, Spain, 1987; 268p.
- Miller, J.D.; Thode, A. Quantifying burn severity in a heterogeneous landscape with a relative version of the delta Normalized Burn Ratio (dNBR). Remote Sens. Environ. 2007, 109, 66–80. [Google Scholar] [CrossRef]
- Gómez-Sánchez, E.; De las Heras, J.; Lucas-Borja, M.; Moya, D. Ajuste de metodologías para evaluar severidad de quemado en zonas semiáridas (SE peninsular): Incendio Donceles 2012. Rev. Teledet. 2017, 49, 103–113. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Sánchez, E.; Lucas-Borja, M.E.; Plaza-Álvarez, P.A.; González-Romero, J.; Sagra, J.; Moya, D.; de las Heras, J. Effects of post-fire hillslope stabilisation techniques on chemical, physicochemical and microbiological soil properties in mediterranean forest ecosystems. J. Environ. Manag. 2019, 246, 229–238. [Google Scholar] [CrossRef]
- Chu, C.; Maestre, F.T.; Xiao, S.; Weiner, J.; Wang, Y.; Duan, Z.; Wang, G. Balance between facilitation and resource competition determines biomass–density relationships in plant populations. Ecol. Lett. 2008, 11, 1189–1197. [Google Scholar] [CrossRef]
- Stohlgren, T.J.; Falkner, M.; Schell, D.L. A Modified-Whittaker nested vegetation sampling method. Plant Ecol. 1995, 117, 113–121. [Google Scholar] [CrossRef]
- Buckland, S.T.; Borchers, D.L.; Johnston, A.; Henrys, P.A.; Marques, T.A. Line transect methods for plant surveys. Biometrics 2007, 63, 989–998. [Google Scholar] [CrossRef] [PubMed]
- Van der Maarel, E. Transformation of cover-abundance values for appropriate numerical treatment–Alternatives to the proposals by Podani. J. Veg. Sci. 2007, 18, 767–770. [Google Scholar]
- Tutin, T.G.; Heywood, V.H.; Burges, N.A.; Moore, D.M.; Valentine, D.H.; Walters, S.M.; Webb, D.A. Flora Europaea 1-5; Cambridge University Press: Cambridge, UK, 1964. [Google Scholar]
- Castroviejo, S. Flora Ibérica 1–8, 10–15, 17–18, 21; Real Jardín Botánico; CSIC: Madrid, Spain, 2012. [Google Scholar]
- Shannon, C. A mathematical theory of communication. Bell. Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef] [Green Version]
- Simpson, E.H. Measurement of diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Paula, S.; Arianoutsou, M.; Kazanis, D.; Tavsanoglu, Ç.; Lloret, F.; Buhk, C.; Ojeda, F.; Luna, B.; Moreno, J.M.; Rodrigo, A. Fire-related traits for plant species of the Mediterranean Basin. Ecology 2009, 90, 1420. [Google Scholar] [CrossRef] [Green Version]
- Pausas, J.G.; Bonet, A.; Maestre, F.T.; Climent, A. The role of the perch effect on the nucleation process in Mediterranean semi-arid oldfields. Acta Oecol. 2006, 29, 346–352. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2011; Available online: http://www.R-project.org (accessed on 16 December 2020).
- Addinsoft. XLSTAT Statistical and Data Analysis Solution; Addinsoft: New York, NY, USA, 2020; Available online: https://www.xlstat.com (accessed on 16 December 2020).
- Thorn, S.; Bässler, C.; Brandl, R.; Burton, P.J.; Cahall, R.; Campbell, J.L.; Castro, J.; Choi, C.Y.; Cobb, T.; Donato, D.C.; et al. Impacts of salvage logging on biodiversity: A meta-analysis. J. Appl. Ecol. 2018, 55, 279–289. [Google Scholar] [CrossRef]
- Leverkus, A.B.; Lindenmayer, D.B.; Thorn, S.; Gustafsson, L. Salvage logging in the world’s forests: Interactions between natural disturbance and logging need recognition. Glob. Ecol. Biogeogr. 2018, 27, 1140–1154. [Google Scholar] [CrossRef]
- Rost, J.; Clavero, M.; Brotons, L.; Pons, P. The effect of postfire salvage logging on bird communities in Mediterranean pine forests: The benefits for declining species. J. Appl. Ecol. 2012, 49, 644–651. [Google Scholar] [CrossRef]
- Zapata, V.M.; Robledano, F.; Ramos, V.; Martínez-López, V. Bird-mediated seed dispersal of fleshy fruits of mediterranean shrubs in semiarid forest patches: The role of Pinus halepensis Miller trees as seed receptors. Plant Ecol. 2014, 215, 1337–1350. [Google Scholar] [CrossRef]
- Castro, J.; Puerta-Piñero, C.; Leverkus, A.B.; Moreno-Rueda, G.; Sánchez-Miranda, A. Post-fire salvage logging alters a key plant-animal interaction for forest regeneration. Ecosphere 2012, 3, 1–12. [Google Scholar] [CrossRef]
- García Fayos, P.; Verdú, M. Soil seed bank, factors controlling germination and establishment of a Mediterranean shrub: Pistacia lentiscus L. Acta Oecol. 1998, 19, 357–366. [Google Scholar] [CrossRef]
- Bonet, A. Secondary succession of semi-arid Mediterranean old-fields in south-eastern Spain: Insights for conservation and restoration of degraded lands. J. Arid Environ. 2004, 56, 213–233. [Google Scholar] [CrossRef]
- Huston, M.A. Disturbance, productivity, and species diversity: Empiricism vs. logic in ecological theory. Ecology 2014, 95, 2382–2396. [Google Scholar] [CrossRef]
- Strand, E.K.; Satterberg, K.L.; Hudak, A.T. Does burn severity affect plant community diversity and composition in mixed conifer forests of the United States Intermountain West one decade post fire? Fire Ecol. 2019, 15, 25. [Google Scholar] [CrossRef] [Green Version]
- Schwilk, D.W.; Keeley, J.E.; Bond, W.J. The intermediate disturbance hypothesis does not explain fire and diversity pattern in fynbos. Plant Ecol. 1997, 132, 77–84. [Google Scholar] [CrossRef]
- Keeley, J.E.; Pausas, J.G.; Rundel, P.W.; Bond, W.J.; Bradstock, R.A. Fire as an evolutionary pressure shaping plant traits. Trends Plant. Sci. 2011, 16, 406–411. [Google Scholar] [CrossRef] [Green Version]
- Keeley, J.E. Fire in Mediterranean Ecosystems: Ecology, Evolution and Management; Cambridge University Press: Cambridge, UK, 2012. [Google Scholar]
- Buhk, C.; Götzenberger, L.; Wesche, K.; Sánchez-Gómez, P.; Hensen, I. Post-fire regeneration in a Mediterranean pine forest with historically low fire frequency. Acta Oecol. 2006, 30, 288–298. [Google Scholar] [CrossRef]
- Leverkus, A.B.; Gustafsson, L.; Lindenmayer, D.B.; Castro, J.; Rey-Benayas, J.M.; Ranius, T.; Thorn, S. Salvage logging effects on regulating ecosystem services and fuel loads. Front. Ecol. Environ. 2020, 18, 391–400. [Google Scholar] [CrossRef]
- Martínez-Murillo, J.F.; López-Vicente, M. Effect of Salvage Logging and Check Dams on Simulated Hydrological Connectivity in a Burned Area. Land Degrad. Dev. 2018, 29, 701–712. [Google Scholar] [CrossRef] [Green Version]
- Mauri, E.; Pons, P. Handbook of Good Practices in Post-Wildfire Management, 2nd ed.; Anifog Project I+D+i CGL2014-54094-R; Universitat de Girona: Girona, Spain, 2019. [Google Scholar]
- Moya, D.; de las Heras, J.; Ferrandis, P.; Herranz, J.M.; Martínez-Sánchez, J.J. Fire resilience and forest restoration in Mediterranean fire-prone areas. Tech. Know. Transf. E-Bull. 2011, 2. [Google Scholar]
- Clark, J.A.; Covey, K. Tree species richness and the logging of natural forests: A meta-analysis. For. Ecol. Manag. 2012, 276, 146–153. [Google Scholar] [CrossRef]
- Georgiadis, G.V.; Georgiadis, T. Post-Fire Vegetation Succession: The Case of Aleppo Pine (Pinus halepensis Miller) Forests of Northern Achaia (Greece). Fresenius Environ. Bull. 2002, 11, 186–193. [Google Scholar]
- Leverkus, A.B.; Lorite, J.; Navarro, F.B.; Sánchez-Cañete, E.P.; Castro, J. Post-fire salvage logging alters species composition and reduces cover, richness, and diversity in Mediterranean plant communities. J. Environ. Manag. 2014, 133, 323–331. [Google Scholar] [CrossRef]
- Martínez-Sánchez, J.J.; Ferrandis, P.; Trabaud, L.; Galindo, R.; Franco, J.A.; Herranz, J.M. Comparative root system structure of post-fire Pinus halepensis Mill. and Cistus monspeliensis L. saplings. Plant Ecol. 2003, 168, 309–320. [Google Scholar] [CrossRef]
- Vega, J.A.; Fernández-Filgueira, C.; Pérez-Gorostiaga, P.; Fonturbel, T. Response of maritime pine (Pinus pinaster Ait.) recruitment to fire severity and post-fire management in a coastal burned area in Galicia (NW Spain). Plant Ecol. 2010, 206, 297–308. [Google Scholar] [CrossRef]
- Lucas-Borja, M.E.; Ortega, R.; Miralles, I.; Plaza-Álvarez, P.A.; González-Romero, J.; Peña-Molina, E.; Moya, D.; Zema, A.; Wagenbrenner, J.W.; de las Heras, J. Effects of wildfire and logging on soil functionality in the short-term in Pinus halepensis M. forests. Eur. J. For. Res. 2020, 139, 935–945. [Google Scholar] [CrossRef]
- Lucas-Borja, M.E.; Plaza-Álvarez, P.A.; González-Romero, J.; Miralles, I.; Sagra, J.; Molina-Peña, E.; Moya, D.; de las Heras, J.; Fernández, C. Post-wildfire straw mulching and salvage logging affects initial pine seedling density and growth in two Mediterranean contrasting climatic areas in Spain. For. Ecol. Manag. 2020, 474, 118363. [Google Scholar] [CrossRef]
- García-Jiménez, R.; Palmero-Iniesta, M.; Espelta, J. Contrasting Effects of Fire Severity on the Regeneration of Pinus halepensis Mill. and Resprouter Species in Recently Thinned Thickets. Forests 2017, 8, 55. [Google Scholar] [CrossRef] [Green Version]
- Robichaud, P.R.; Waldrop, T.A. A comparison of surface runoff and sediment yields from low- and high-severity site preparation burns. Water Res. Bull. 1994, 30, 27–34. [Google Scholar] [CrossRef]
- Donato, D.C.; Fontaine, J.B.; Robinson, W.D.; Kauffman, J.B.; Law, B.E. Vegetation response to a short interval between high-severity wildfires in a mixed-evergreen forest. J. Ecol. 2009, 97, 142–154. [Google Scholar] [CrossRef] [Green Version]
- Van Wilgen, B.W.; Forsyth, G.G.; De Klerk, H.; Das, S.; Khuluse, S.; Schmitz, P. Fire management in Mediterranean-climate shrublands: A case study from the Cape fynbos, South Africa. J. Appl. Ecol. 2010, 47, 631–638. [Google Scholar] [CrossRef] [Green Version]
- Richter, C.; Rejmanek, M.; Miller, J.E.D.; Welch, K.R.; Weeks, J.; Safford, H. The species diversity x fire severity relationship is hump-shaped in semiarid yellow pine and mixed conifer forests. Ecosphere 2019, 10, e02882. [Google Scholar] [CrossRef]
- Doblas-Miranda, E.; Martínez-Vilalta, J.; Lloret, F.; Álvarez, A.; Ávila, A.; Bonet, F.J.; Brotons, L.; Castro, J.; Curiel Yuste, J.; Díaz, M.; et al. Reassessing global change research priorities in mediterranean terrestrial ecosystems: How far have we come and where do we go from here? Glob. Ecol. Biogeogr. 2015, 24, 25–43. [Google Scholar] [CrossRef] [Green Version]
- Johnstone, J.F.; Allen, C.D.; Franklin, J.F.; Frelich, L.E.; Harvey, B.J.; Higuera, P.E.; Mack, M.C.; Meentemeyer, R.K.; Metz, M.R.; Perry, G.L.W.; et al. Changing disturbance regimes, ecological memory, and forest resilience. Front. Ecol. Environ. 2016, 14, 369–378. [Google Scholar] [CrossRef]
- European Parliament. Directive 2009/147/EC on the Conservation of Wild Birds. 2009. Available online: https://eur-lex.europa.eu/eli/dir/2009/147/oj (accessed on 16 December 2020).





{kind=link}
{kind=link}
{kind=link}
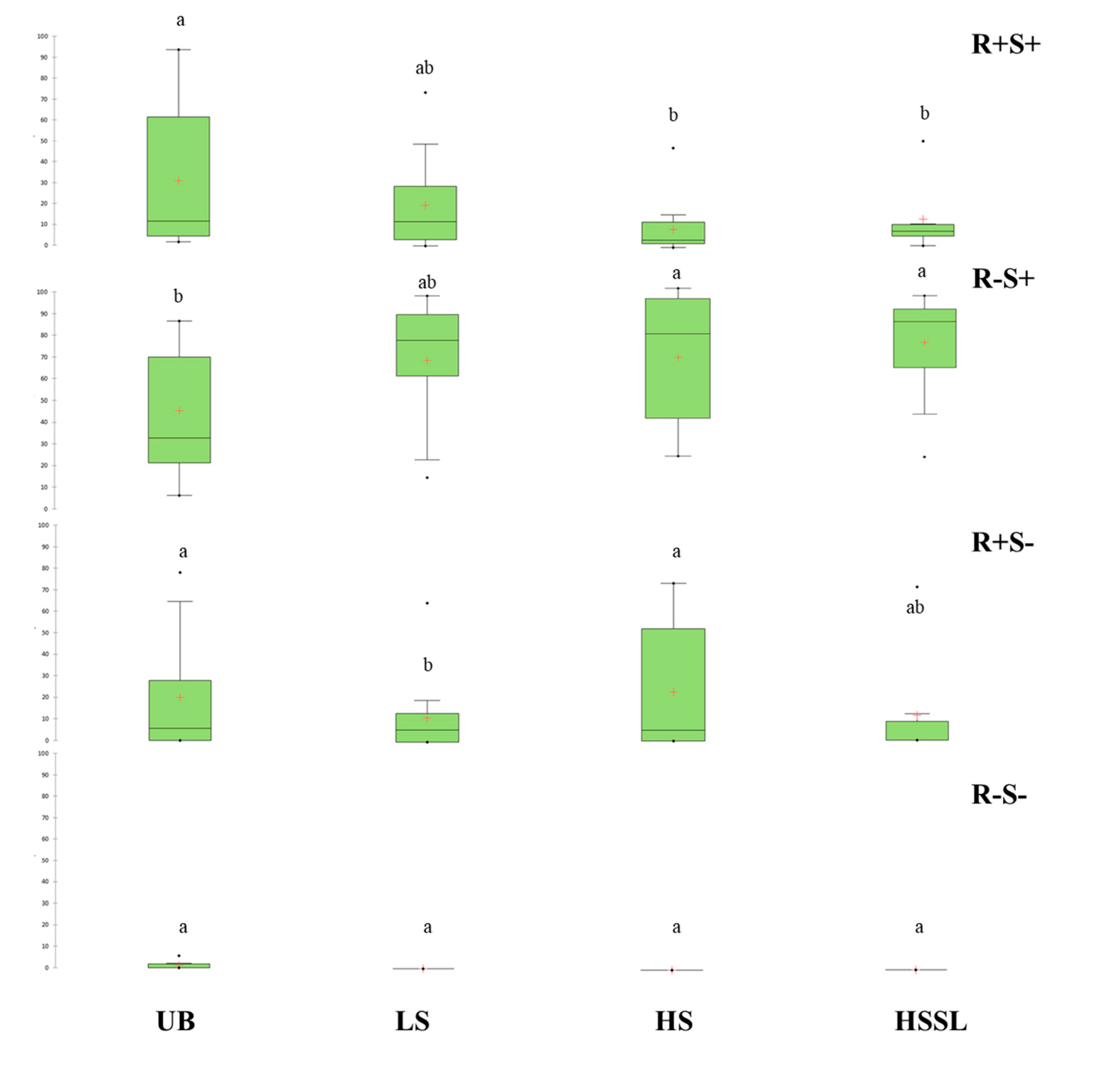{kind=link}
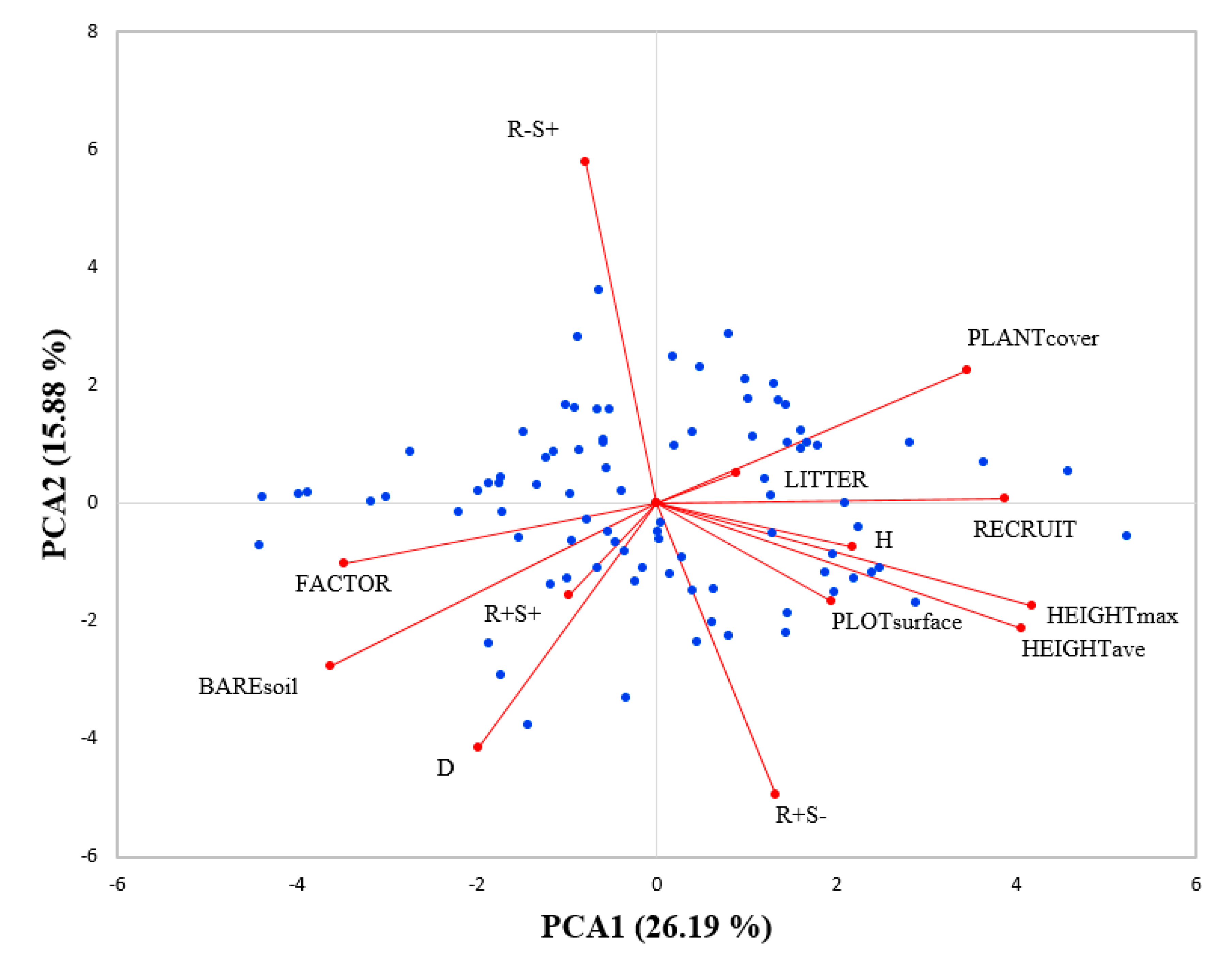{kind=link}
| S | H′ | D | PLANTcover | LITTER | BAREsoil | |
|---|---|---|---|---|---|---|
| UB | 20.3 ± 1.9 a | 0.952 ± 0.049 a | 0.738 ± 0.021 a | 133.86 ± 9.91 a | 31.10 ± 4.32 a | 16.02 ± 4.47 a |
| LS | 24.7 ± 2.3 a | 0.872 ± 0.046 a | 0.921 ± 0.011 c | 149.52 ± 8.54 a | 29.53 ± 7.29 a | 28.86 ± 3.08 ab |
| HS | 23.6 ± 2.0 a | 1.122 ± 0.041 b | 0.736 ± 0.021 a | 119.86 ± 6.67 b | 12.79 ± 1.83 b | 19.55 ± 3.65 ab |
| HSSL | 25.0 ± 2.3 a | 1.170 ± 0.048 b | 0.809 ± 0.010 b | 93.90 ± 6.89 c | 8.67 ± 0.98 c | 30.25 ± 2.64 b |
| p-value | 0.340 | 0.048 | 0.000 | 0.000 | 0.001 | 0.010 |
| F-value | 1.168 | 2.69 | 25.06 | 8.29 | 6.03 | 3.99 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moya, D.; Sagra, J.; Lucas-Borja, M.E.; Plaza-Álvarez, P.A.; González-Romero, J.; De Las Heras, J.; Ferrandis, P. Post-Fire Recovery of Vegetation and Diversity Patterns in Semiarid Pinus halepensis Mill. Habitats after Salvage Logging. Forests 2020, 11, 1345. https://doi.org/10.3390/f11121345
Moya D, Sagra J, Lucas-Borja ME, Plaza-Álvarez PA, González-Romero J, De Las Heras J, Ferrandis P. Post-Fire Recovery of Vegetation and Diversity Patterns in Semiarid Pinus halepensis Mill. Habitats after Salvage Logging. Forests. 2020; 11(12):1345. https://doi.org/10.3390/f11121345
Chicago/Turabian StyleMoya, Daniel, Javier Sagra, Manuel Esteban Lucas-Borja, Pedro Antonio Plaza-Álvarez, Javier González-Romero, Jorge De Las Heras, and Pablo Ferrandis. 2020. "Post-Fire Recovery of Vegetation and Diversity Patterns in Semiarid Pinus halepensis Mill. Habitats after Salvage Logging" Forests 11, no. 12: 1345. https://doi.org/10.3390/f11121345