Mapping Tree Species Deciduousness of Tropical Dry Forests Combining Reflectance, Spectral Unmixing, and Texture Data from High-Resolution Imagery

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Field Data and Calculation of Leafless Trees Species
2.3. Remotely Sensed Data and Imagery Processing
2.4. Data Analysis
3. Results
3.1. Patterns of Tree Species Deciduousness
3.2. Modeling Tree Species Deciduousness and Model Validations
3.3. Relationships between Predictor Variables and Tree Species Deciduousness
3.4. Variance Partitioning of Tree Species Deciduousness
3.5. Mapping the Spatial Distribution of Tree Species Deciduousness and Its Uncertainty
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Miles, L.; Newton, A.C.; DeFries, R.S.; Ravilious, C.; May, I.; Blyth, S.; Kapos, V.; Gordon, J.E. A Global Overview of the Conservation Status of Tropical Dry Forests. J. Biogeogr. 2006, 33, 491–505. [Google Scholar] [CrossRef]
- Griscom, H.P.; Ashton, M.S. Restoration of Dry Tropical Forests in Central America: A Review of Pattern and Process. For. Ecol. Manag. 2011, 261, 1564–1579. [Google Scholar] [CrossRef]
- Murphy, P.G.; Lugo, A.E. Ecology of Tropical Dry Forest. Annu. Rev. Ecol. Syst. 1986, 17, 67–88. [Google Scholar] [CrossRef]
- Eamus, D.; Prior, L. Ecophysiology of Trees of Seasonally Dry Tropics: Comparisons among Phenologies. Adv. Ecol. Res. 2001, 32, 113–197. [Google Scholar] [CrossRef]
- Lebrija-Trejos, E.; Pérez-GarcíA, E.A.; Meave, J.A.; Bongers, F.; Poorter, L. Functional Traits and Environmental Filtering Drive Community Assembly in a Species-Rich Tropical System. Ecology 2010, 91, 386–398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lohbeck, M.; Lebrija-Trejos, E.; Martínez-Ramos, M.; Meave, J.A.; Poorter, L.; Bongers, F. Functional Trait Strategies of Trees in Dry and Wet Tropical Forests Are Similar but Differ in Their Consequences for Succession. PLoS ONE 2015, 10, e0123741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kikuzawa, K.; Lechowicz, M.J. Ecology of Leaf Longevity. Ecol. Res. Monogr. 2011. [Google Scholar] [CrossRef]
- Bohlman, S.A. Landscape Patterns and Environmental Controls of Deciduousness in Forests of Central Panama. Glob. Ecol. Biogeogr. 2010, 19, 376–385. [Google Scholar] [CrossRef]
- Singh, K.P.; Kushwaha, C.P. Deciduousness in Tropical Trees and Its Potential as Indicator of Climate Change: A Review. Ecol. Indic. 2016, 69, 699–706. [Google Scholar] [CrossRef]
- Zhou, G.; Houlton, B.Z.; Wang, W.; Huang, W.; Xiao, Y.; Zhang, Q.; Liu, S.; Cao, M.; Wang, X.; Wang, S.; et al. Substantial Reorganization of China’s Tropical and Subtropical Forests: Based on the Permanent Plots. Glob. Chang. Biol. 2014, 20, 240–250. [Google Scholar] [CrossRef]
- Condit, R.; Watts, K.; Bohlman, S.A.; Pérez, R.; Foster, R.B.; Hubbell, S.P. Quantifying the Deciduousness of Tropical Forest Canopies under Varying Climates. J. Veg. Sci. 2000, 11, 649–658. [Google Scholar] [CrossRef] [Green Version]
- Williams, L.J.; Bunyavejchewin, S.; Baker, P.J. Deciduousness in a Seasonal Tropical Forest in Western Thailand: Interannual and Intraspecific Variation in Timing, Duration and Environmental Cues. Oecologia 2008, 155, 571–582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kushwaha, C.P.; Tripathi, S.K.; Singh, G.S.; Singh, K.P. Diversity of Deciduousness and Phenological Traits of Key Indian Dry Tropical Forest Trees. Ann. For. Sci. 2010, 67, 310. [Google Scholar] [CrossRef] [Green Version]
- Gond, V.; Fayolle, A.; Pennec, A.; Cornu, G.; Mayaux, P.; Camberlin, P.; Doumenge, C.; Fauvet, N.; Gourlet-Fleury, S. Vegetation Structure and Greenness in Central Africa from Modis Multi-Temporal Data. Philos. Trans. R. Soc. B Biol. Sci. 2013, 368, 20120309. [Google Scholar] [CrossRef] [Green Version]
- Ouédraogo, D.Y.; Fayolle, A.; Gourlet-Fleury, S.; Mortier, F.; Freycon, V.; Fauvet, N.; Rabaud, S.; Cornu, G.; Bénédet, F.; Gillet, J.F.; et al. The Determinants of Tropical Forest Deciduousness: Disentangling the Effects of Rainfall and Geology in Central Africa. J. Ecol. 2016, 104, 924–935. [Google Scholar] [CrossRef]
- Valdez-Hernández, M.; González-Salvatierra, C.; Reyes-García, C.; Jackson, P.C.; Andrade, J.L. Physiological Ecology of Vascular Plants. In Biodiversity and Conservation of the Yucatan Peninsula; Springer: Cham, Switzerland, 2015. [Google Scholar] [CrossRef]
- Cuba, N.; Lawrence, D.; Rogan, J.; Williams, C.A. Local Variability in the Timing and Intensity of Tropical Dry Forest Deciduousness Is Explained by Differences in Forest Stand Age. GIScience Remote Sens. 2018, 55, 437–456. [Google Scholar] [CrossRef]
- Ganguly, S.; Friedl, M.A.; Tan, B.; Zhang, X.; Verma, M. Land Surface Phenology from MODIS: Characterization of the Collection 5 Global Land Cover Dynamics Product. Remote Sens. Environ. 2010, 114, 1805–1816. [Google Scholar] [CrossRef] [Green Version]
- Viennois, G.; Barbier, N.; Fabre, I.; Couteron, P. Multiresolution Quantification of Deciduousness in West-Central African Forests. Biogeosciences 2013, 10, 6957–6967. [Google Scholar] [CrossRef]
- Cuba, N.; Rogan, J.; Christman, Z.; Williams, C.A.; Schneider, L.C.; Lawrence, D.; Millones, M. Modelling Dry Season Deciduousness in Mexican Yucatán Forest Using MODIS EVI Data (2000-2011). GIScience Remote Sens. 2013, 50, 26–49. [Google Scholar] [CrossRef]
- Cuba, N.; Rogan, J.; Lawrence, D.; Williams, C. Cross-Scale Correlation between in Situ Measurements of Canopy Gap Fraction and Landsat-Derived Vegetation Indices with Implications for Monitoring the Seasonal Phenology in Tropical Forests Using MODIS Data. Remote Sens. 2018, 10, 979. [Google Scholar] [CrossRef] [Green Version]
- Pasher, J.; King, D.J. Multivariate Forest Structure Modelling and Mapping Using High Resolution Airborne Imagery and Topographic Information. Remote Sens. Environ. 2010, 114, 1718–1732. [Google Scholar] [CrossRef]
- Ploton, P.; Barbier, N.; Couteron, P.; Antin, C.M.; Ayyappan, N.; Balachandran, N.; Barathan, N.; Bastin, J.F.; Chuyong, G.; Dauby, G.; et al. Toward a General Tropical Forest Biomass Prediction Model from Very High Resolution Optical Satellite Images. Remote Sens. Environ. 2017, 200, 140–153. [Google Scholar] [CrossRef]
- Rzedowski, J. Vegetacion de Mexico; Comisión Nacional para el Conocimiento y Uso de la. Biodiversidad (CONABIO): Mexico City, Mexico, 2006. [Google Scholar]
- Islebe, G.A.; Schmook, B.; Calmé, S.; León-Cortés, J.L. Introduction: Biodiversity and Conservation of the Yucatán Peninsula, Mexico. In Biodiversity and Conservation of the Yucatan Peninsula; Springer: Cham, Switzerland, 2015. [Google Scholar] [CrossRef]
- Duch, G. La Conformación Territorial Del Estado de Yucatán, Los Componentes Del Medio Físico; Universidad Autonoma Chapingo. Centro Regional de la Península de Yucatán: Texcoco, México, 1988. [Google Scholar]
- Orellana, R.; Espadas, C.; Conde, C.; Gay, C. Atlas Escenarios de Cambio Climático En La Península de Yucatán; Centro de Investigación Científica de Yucatán (CICY): Mérida, México, 2009. [Google Scholar]
- Bautista-Zúñiga, F.; Batllori-Sampedro, E.; Ortiz-Pérez, M.A.; Palacio-Aponte, G.; Castillo-González, M. Geoformas, Agua y Suelo En La Península de Yucatán. In Naturaleza y Sociedad en el área Maya. Pasado, Presente y Futuro; Colunga, P., Larqué, A., Eds.; Centro de Investigación Científica de Yucatán (CICY): Mérida, México, 2003; pp. 21–36. [Google Scholar]
- Miranda, F.; Hernández-X, E. Los Tipos de Vegetación de México y Su Clasificación. Bot. Sci. 1963, 28, 29–179. [Google Scholar] [CrossRef]
- CONAFOR [Comisión Nacional Forestal]. Inventario Nacional y de Suelos. Manual y Procedimientos Para El Muestreo de Campo; CONAFOR: Zapopan, Jalisco, Mexico, 2013. [Google Scholar]
- Hernández-Stefanoni, J.L.; Reyes-Palomeque, G.; Castillo-Santiago, M.Á.; George-Chacón, S.P.; Huechacona-Ruiz, A.H.; Tun-Dzul, F.; Rondon-Rivera, D.; Dupuy, J.M. Effects of Sample Plot Size and GPS Location Errors on Aboveground Biomass Estimates from LiDAR in Tropical Dry Forests. Remote Sens. 2018, 10, 1586. [Google Scholar] [CrossRef] [Green Version]
- Gorelick, N.; Hancher, M.; Dixon, M.; Ilyushchenko, S.; Thau, D.; Moore, R. Google Earth Engine: Planetary-Scale Geospatial Analysis for Everyone. Remote Sens. Environ. 2017, 202, 18–27. [Google Scholar] [CrossRef]
- Sentinel, E.S.A. User Handbook; ESA Stand. Doc. 2AD; ESA: Paris, France, 2015; p. 64. [Google Scholar]
- Adams, J.B.; Gillespie, A.R. Remote Sensing of Landscapes with Spectral Images; Cambridge University Press: Cambridge, UK, 2006. [Google Scholar] [CrossRef]
- Boardman, J.W.; Kruse, F.A.; Green, R.O. Mapping Target Signatures via Partial Unmixing of AVIRIS Data. Summ. JPL Airborne Earth Sci. Work. 1995, 1, 23–26. [Google Scholar]
- Exelis Visual Information Solutions. Environment for Visualizing Images (ENVI); Exelis Visual Information Solutions: Boulder, CO, USA, 2013. [Google Scholar]
- Haralick, R.M.; Dinstein, I.; Shanmugam, K. Textural Features for Image Classification. IEEE Trans. Syst. Man Cybern. 1973, 610–621. [Google Scholar] [CrossRef] [Green Version]
- Zvoleff, A. Glcm: Calculate Textures from Grey-Level Co-Occurrence Matrices (GLCMs); R-CRAN Project; 2019; Available online: https://cran.r-project.org/web/packages/glcm/index.html (accessed on 26 March 2019).
- Liaw, A.; Wiener, M. Classification and Regression by RandomForest. R News 2002, 2, 18–22. [Google Scholar]
- Breiman, L. Random Forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef] [Green Version]
- Borcard, D.; Legendre, P.; Avois-Jacquet, C.; Tuomisto, H. Dissecting the Spatial Structure of Ecological Data at Multiple Scales. Ecology 2004, 85, 1826–1832. [Google Scholar] [CrossRef] [Green Version]
- Freeman, E.; Frescino, T.; Moisen, G. ModelMap: An R Package for Modeling and Map Production Using Random Forest and Stochastic Gradient Boosting; USDA Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2009. [Google Scholar]
- Zar, J. Biostatistical Analysis, 4nd ed.; Prentice Hall: Upper Saddle River, NJ, USA, 1999. [Google Scholar]
- Freeman, E.A.; Moisen, G.G.; Coulston, J.W.; Wilson, B.T. Random Forests and Stochastic Gradient Boosting for Predicting Tree Canopy Cover: Comparing Tuning Processes and Model Performance. Can. J. For. Res. 2015, 46, 323–339. [Google Scholar] [CrossRef] [Green Version]
- Adole, T.; Dash, J.; Atkinson, P.M. A Systematic Review of Vegetation Phenology in Africa. Ecol. Inform. 2016, 34, 117–128. [Google Scholar] [CrossRef]
- Feret, J.B.; Corbane, C.; Alleaume, S. Detecting the Phenology and Discriminating Mediterranean Natural Habitats with Multispectral Sensors-An Analysis Based on Multiseasonal Field Spectra. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2015, 8, 2294–2305. [Google Scholar] [CrossRef]
- Xu, X.; Medvigy, D.; Powers, J.S.; Becknell, J.M.; Guan, K. Diversity in Plant Hydraulic Traits Explains Seasonal and Inter-Annual Variations of Vegetation Dynamics in Seasonally Dry Tropical Forests. New Phytol. 2016, 212, 80–95. [Google Scholar] [CrossRef] [PubMed]
- Restrepo-Coupe, N.; Levine, N.M.; Christoffersen, B.O.; Albert, L.P.; Wu, J.; Costa, M.H.; Galbraith, D.; Imbuzeiro, H.; Martins, G.; da Araujo, A.C.; et al. Do Dynamic Global Vegetation Models Capture the Seasonality of Carbon Fluxes in the Amazon Basin? A Data-Model Intercomparison. Glob. Chang. Biol. 2017, 23, 191–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van der Sande, M.T.; Peña-Claros, M.; Ascarrunz, N.; Arets, E.J.M.M.; Licona, J.C.; Toledo, M.; Poorter, L. Abiotic and Biotic Drivers of Biomass Change in a Neotropical Forest. J. Ecol. 2017, 105, 1223–1234. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Stefanoni, J.L.; Castillo-Santiago, M.Á.; Mas, J.F.; Wheeler, C.E.; Andres-Mauricio, J.; Tun-Dzul, F.; George-Chacón, S.P.; Reyes-Palomeque, G.; Castellanos-Basto, B.; Vaca, R.; et al. Improving Aboveground Biomass Maps of Tropical Dry Forests by Integrating LiDAR, ALOS PALSAR, Climate and Field Data. Carbon Balance Manag. 2020, 15, 1–17. [Google Scholar] [CrossRef]
- Vieira, I.C.G.; De Almeida, A.S.; Davidson, E.A.; Stone, T.A.; Reis De Carvalho, C.J.; Guerrero, J.B. Classifying Successional Forests Using Landsat Spectral Properties and Ecological Characteristics in Eastern Amazônia. Remote Sens. Environ. 2003, 87, 470–481. [Google Scholar] [CrossRef]
- Thenkabail, P.S.; Enclona, E.A.; Ashton, M.S.; Legg, C.; De Dieu, M.J. Hyperion, IKONOS, ALI, and ETM+ Sensors in the Study of African Rainforests. Remote Sens. Environ. 2004, 90, 23–43. [Google Scholar] [CrossRef]
- Muldavin, E.H.; Neville, P.; Harper, G. Indices of Grassland Biodiversity in the Chihuahuan Desert Ecoregion Derived from Remote Sensing. Conserv. Biol. 2001, 15, 844–855. [Google Scholar] [CrossRef]
- Gallardo-Cruz, J.A.; Meave, J.A.; González, E.J.; Lebrija-Trejos, E.E.; Romero-Romero, M.A.; Pérez-García, E.A.; Gallardo-Cruz, R.; Hernández-Stefanoni, J.L.; Martorell, C. Predicting Tropical Dry Forest Successional Attributes from Space: Is the Key Hidden in Image Texture? PLoS ONE 2012, 7, e30506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viedma, O.; Torres, I.; Pérez, B.; Moreno, J.M. Modeling Plant Species Richness Using Reflectance and Texture Data Derived from QuickBird in a Recently Burned Area of Central Spain. Remote Sens. Environ. 2012, 119, 208–221. [Google Scholar] [CrossRef]
- George-Chacon, S.P.; Dupuy, J.M.; Peduzzi, A.; Hernandez-Stefanoni, J.L. Combining High Resolution Satellite Imagery and Lidar Data to Model Woody Species Diversity of Tropical Dry Forests. Ecol. Indic. 2019, 101, 975–984. [Google Scholar] [CrossRef]
- Zhou, J.; Yan Guo, R.; Sun, M.; Di, T.T.; Wang, S.; Zhai, J.; Zhao, Z. The Effects of GLCM Parameters on LAI Estimation Using Texture Values from Quickbird Satellite Imagery. Sci. Rep. 2017, 7, 7366. [Google Scholar] [CrossRef]
- Chen, L.; Wang, Y.; Ren, C.; Zhang, B.; Wang, Z. Optimal Combination of Predictors and Algorithms for Forest Above-Ground Biomass Mapping from Sentinel and SRTM Data. Remote Sens. 2019, 11, 414. [Google Scholar] [CrossRef] [Green Version]
- Reyes-Palomeque, G.; Dupuy, J.M.; Johnson, K.D.; Castillo-Santiago, M.A.; Hernández-Stefanoni, J.L. Combining LiDAR Data and Airborne Imagery of Very High Resolution to Improve Aboveground Biomass Estimates in Tropical Dry Forests. For. Int. J. For. Res. 2019, 92, 599–615. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Acquisition Time | Sensor Type | Tile Numbers |
|---|---|---|---|
| El Palmar | 24 March 2018 | S2A | T15QYC, T15QYD, T15QZC, T15QZD |
| 26 March 2018 | S2B | T16QBH, T16QBJ | |
| Kaxil Kiuic | 6 March 2018 | S2B | T15QZB, T15QZC |
| FCP | 16 March 2018 | S2B | T16QCF, T16QCG |
| Site | n | Mean | SD | Min | Max | Range |
|---|---|---|---|---|---|---|
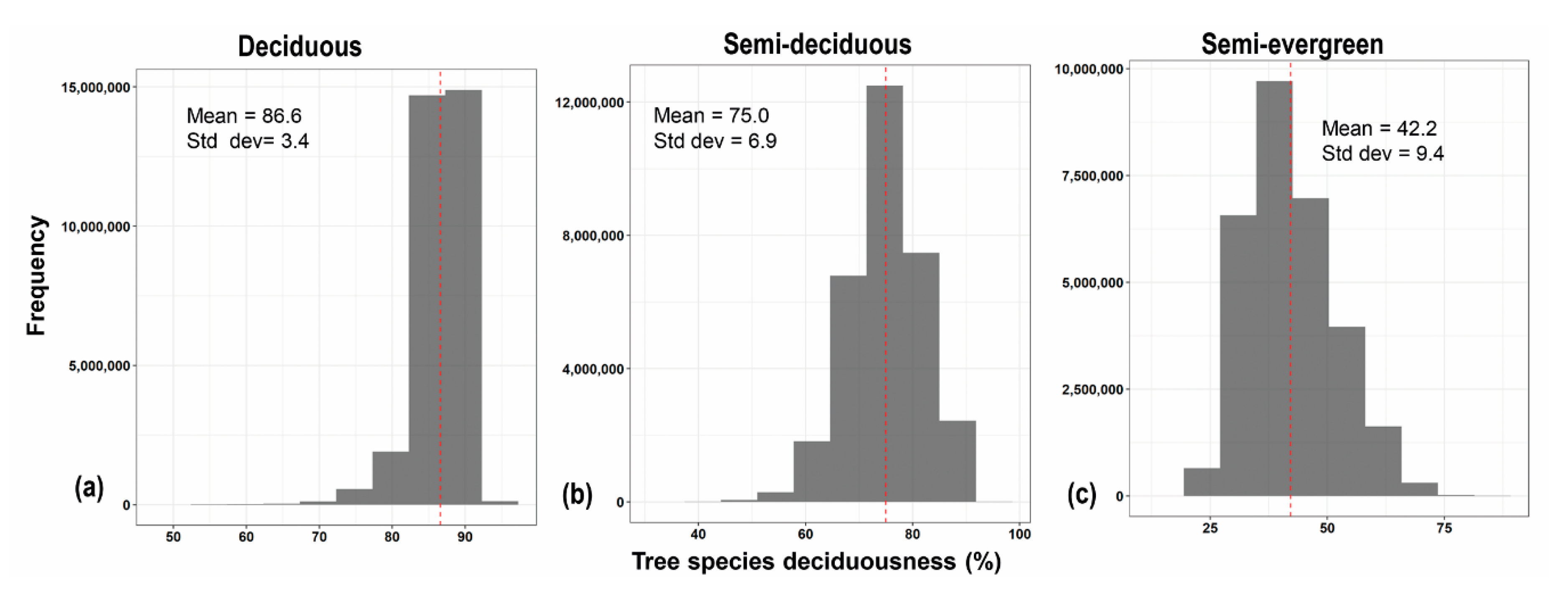
| Palmar | 33 | 91.5 | 7.9 | 65.9 | 100.0 | 34.1 |
| Kaxil Kiuic | 134 | 80.4 | 14.5 | 0.0 | 100.0 | 100.0 |
| FCP | 121 | 43.3 | 18.2 | 0.8 | 100.0 | 99.2 |
| Model | Explanatory Variables | R2 | RMSE (%) |
|---|---|---|---|
| Combining spectral and texture variables | Spectral values from blue, green red, and NIR bands + NDVI + SMA deciduous fraction + Texture metrics of spectral bands, NDVI and SMA | 0.60 | 16.2 |
| Spectral variables | Spectral values from blue, green red, and NIR bands + NDVI + SMA deciduous fraction | 0.56 | 16.9 |
| Texture variables | Texture metrics of spectral bands, NDVI, and SMA | 0.59 | 16.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huechacona-Ruiz, A.H.; Dupuy, J.M.; Schwartz, N.B.; Powers, J.S.; Reyes-García, C.; Tun-Dzul, F.; Hernández-Stefanoni, J.L. Mapping Tree Species Deciduousness of Tropical Dry Forests Combining Reflectance, Spectral Unmixing, and Texture Data from High-Resolution Imagery. Forests 2020, 11, 1234. https://doi.org/10.3390/f11111234
Huechacona-Ruiz AH, Dupuy JM, Schwartz NB, Powers JS, Reyes-García C, Tun-Dzul F, Hernández-Stefanoni JL. Mapping Tree Species Deciduousness of Tropical Dry Forests Combining Reflectance, Spectral Unmixing, and Texture Data from High-Resolution Imagery. Forests. 2020; 11(11):1234. https://doi.org/10.3390/f11111234
Chicago/Turabian StyleHuechacona-Ruiz, Astrid Helena, Juan Manuel Dupuy, Naomi B. Schwartz, Jennifer S. Powers, Casandra Reyes-García, Fernando Tun-Dzul, and José Luis Hernández-Stefanoni. 2020. "Mapping Tree Species Deciduousness of Tropical Dry Forests Combining Reflectance, Spectral Unmixing, and Texture Data from High-Resolution Imagery" Forests 11, no. 11: 1234. https://doi.org/10.3390/f11111234