The Structure and Composition of Puerto Rico’s Urban Mangroves


Abstract
:1. Introduction
2. Materials and Methods
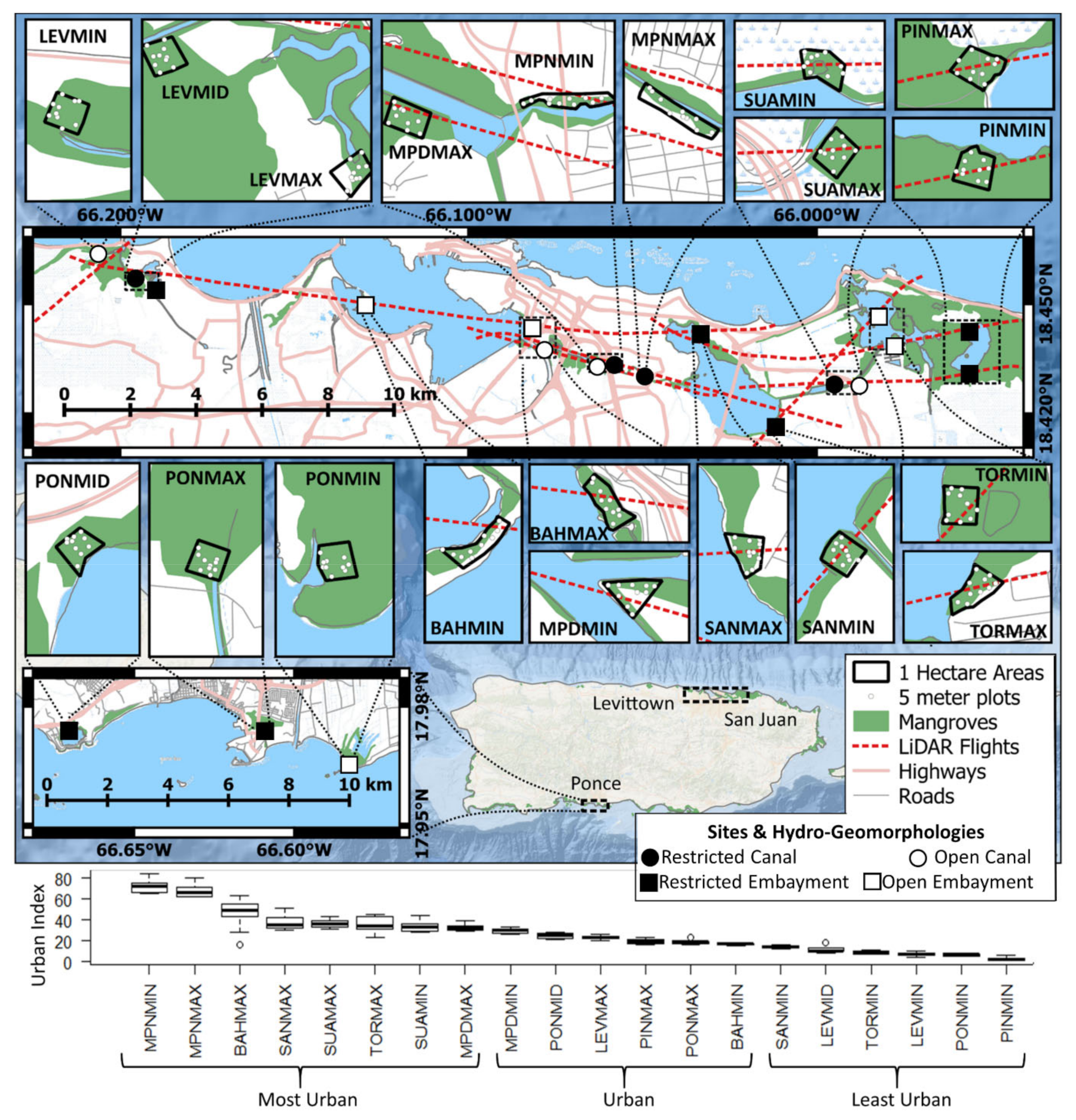
2.1. Study Location
2.2. Ground-Based Measurements
2.3. LiDAR
2.4. Land Use, Flooding, and Water Chemistry
2.5. Analyses and Statistical Testing
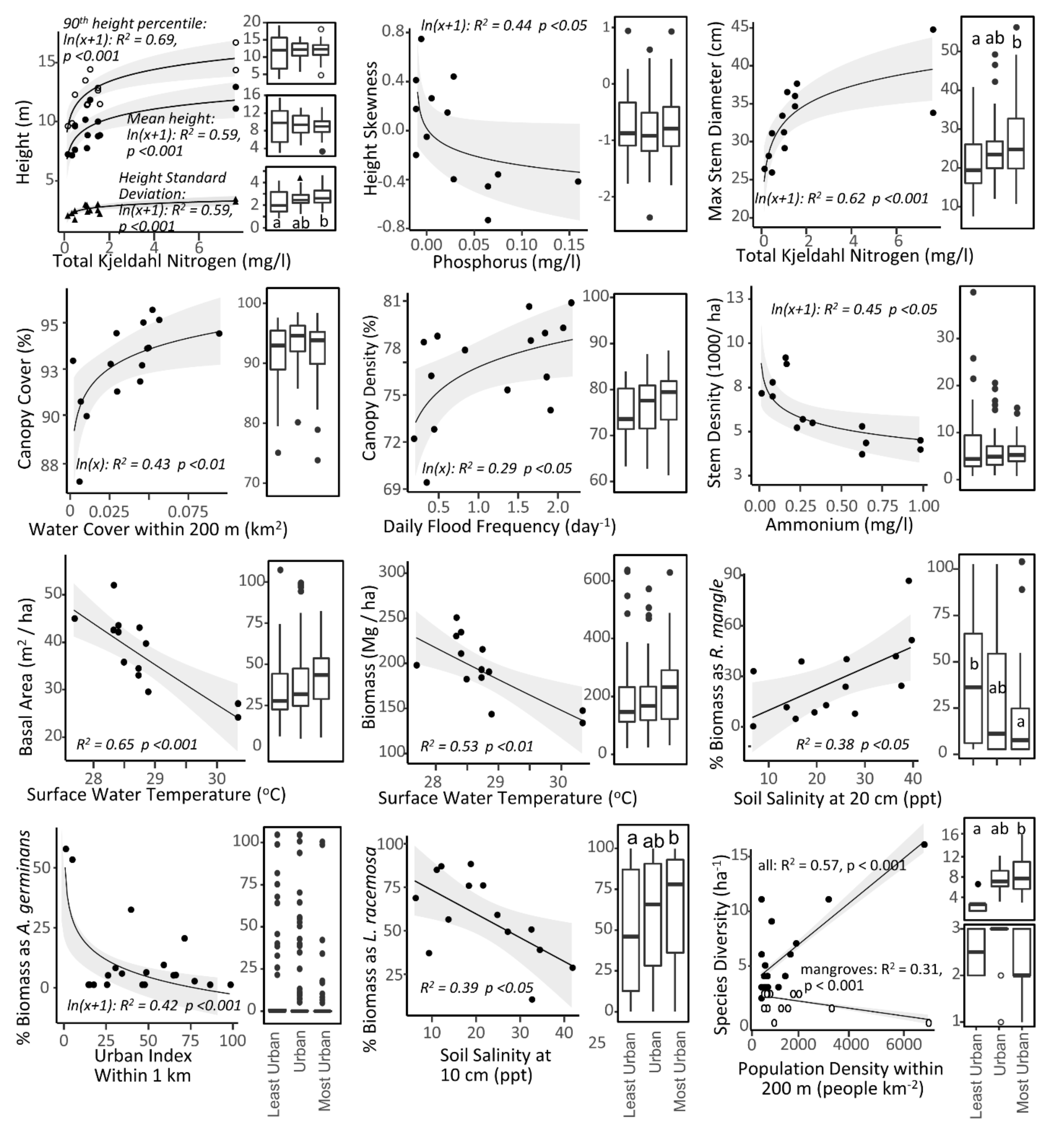
3. Results
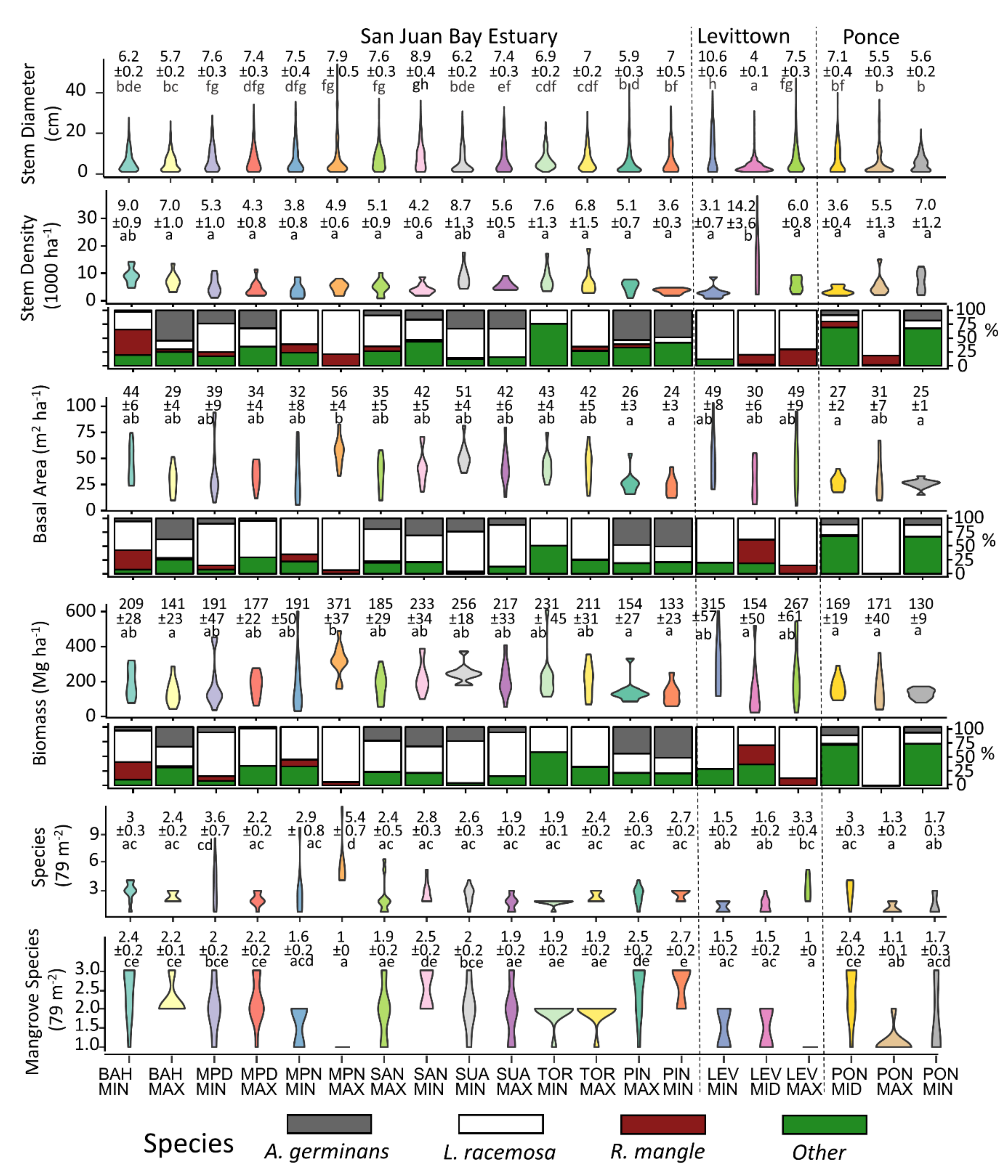
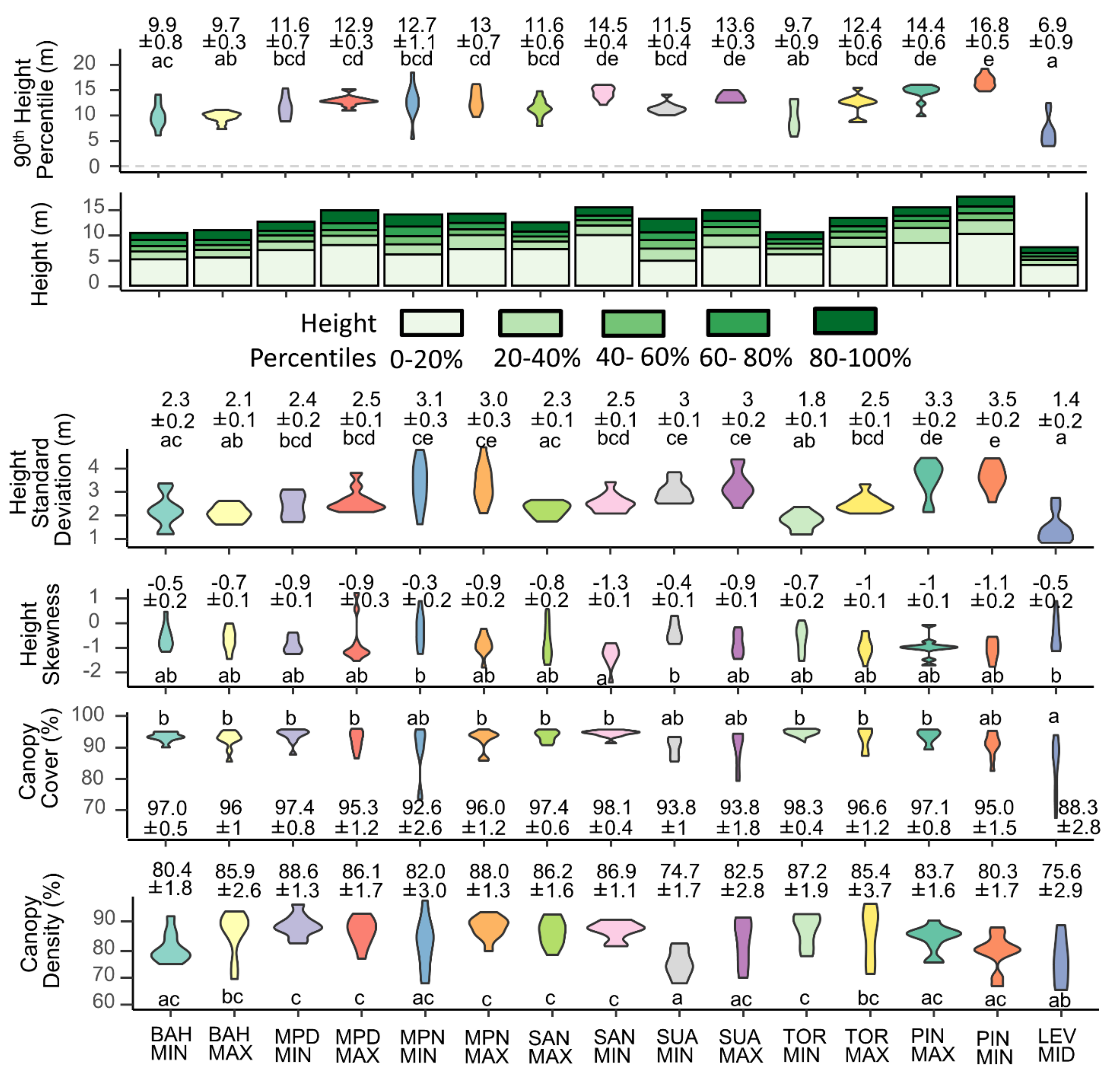
3.1. Structure and Composition of Mangroves
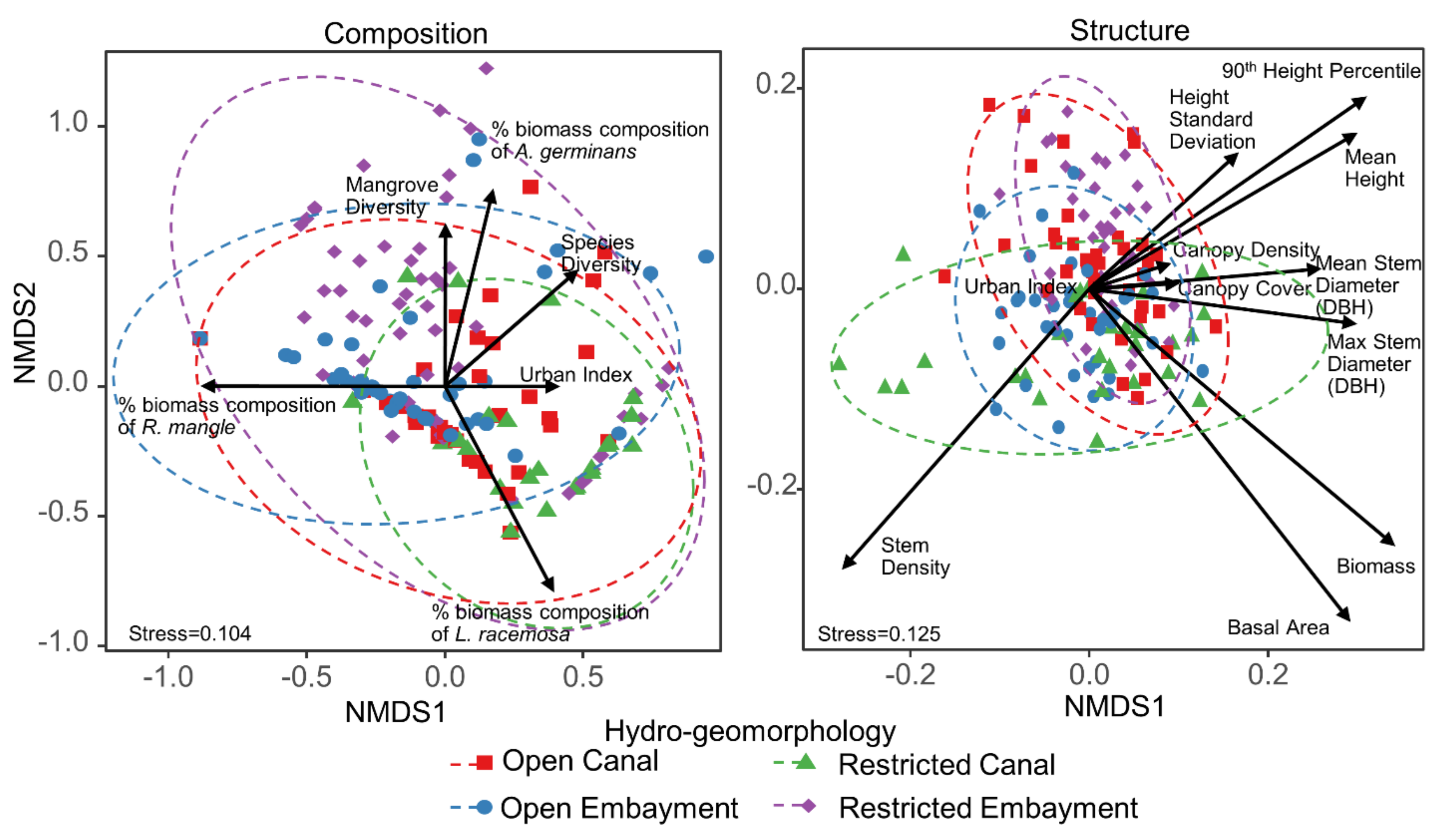
3.2. Non-Metric Multidimensional Scaling
3.3. Simple Regression
3.4. Multiple Regression
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Disclaimer:
References
- de Fries, R.S.; Rudel, T.; Uriarte, M.; Hansen, M. Deforestation driven by urban population growth and agricultural trade in the twenty-first century. Nat. Geosci. 2010, 3, 178–181. [Google Scholar] [CrossRef]
- Geist, H.J.; Lambin, E.F. Proximate Causes and Underlying Driving Forces of Tropical Deforestation: Tropical Forests are Disappearing as the Result of Many Pressures, Both Local and Regional, Acting in Various Combinations in Different Geographical Locations. BioScience 2002, 52, 143–150. [Google Scholar] [CrossRef]
- Jorgenson, A.K.; Burns, T.J. Effects of Rural and Urban Population Dynamics and National Development on Deforestation in Less-Developed Countries, 1990–2000. Sociol. Inq. 2007, 77, 460–482. [Google Scholar] [CrossRef]
- Seto, K.C.; Güneralp, B.; Hutyra, L.R. Global forecasts of urban expansion to 2030 and direct impacts on biodiversity and carbon pools. Proc. Natl. Acad. Sci. USA 2012, 109, 16083–16088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wright, S.J. Tropical forests in a changing environment. Trends Ecol. Evol. 2005, 20, 553–560. [Google Scholar] [CrossRef]
- Wright, S.J.; Muller-Landau, H.C. The Future of Tropical Forest Species. Biotropica 2006, 38, 287–301. [Google Scholar] [CrossRef]
- Grimm, N.B.; Faeth, S.H.; Golubiewski, N.E.; Redman, C.L.; Wu, J.; Bai, X.; Briggs, J.M. Global Change and the Ecology of Cities. Science 2008, 319, 756–760. [Google Scholar] [CrossRef] [Green Version]
- Pickett, S.T.A.; Cadenasso, M.L.; Grove, J.M.; Nilon, C.H.; Pouyat, R.; Zipperer, W.; Costanza, R. Urban Ecological Systems: Linking Terrestrial Ecological, Physical, and Socioeconomic Components of Metropolitan Areas. Annu. Rev. Ecol. Syst. 2001, 32, 127–157. [Google Scholar] [CrossRef] [Green Version]
- Rowntree, R.A. Ecology of the urban forest—Introduction to part I. Urban Ecol. 1984, 8, 1–11. [Google Scholar] [CrossRef]
- Alberti, M. The Effects of Urban Patterns on Ecosystem Function. Int. Reg. Sci. Rev. 2005, 28, 168–192. [Google Scholar] [CrossRef]
- McDonnell, M.J.; Pickett, S.T.A. Ecosystem Structure and Function along Urban-Rural Gradients: An Unexploited Opportunity for Ecology. Ecology 1990, 71, 1232–1237. [Google Scholar] [CrossRef]
- Dwyer, J.F.; McPherson, E.G.; Schroeder, H.W.; Rowntree, R.A. Assessing the Benefits and Costs of the Urban Forest. J. Arboric. 1992, 18, 227. [Google Scholar]
- McPherson, E.G.; Nowak, D.; Heisler, G.; Grimmond, S.; Souch, C.; Grant, R.; Rowntree, R. Quantifying urban forest structure, function, and value: The Chicago Urban Forest Climate Project. Urban Ecosyst. 1997, 1, 49–61. [Google Scholar] [CrossRef]
- Branoff, B.L. Quantifying the influence of urban land use on mangrove biology and ecology: A meta-analysis. Glob. Ecol. Biogeogr. 2017, 26, 1339–1356. [Google Scholar] [CrossRef]
- Benfield, S.L.; Guzmán, H.M.; Mair, J.M. Temporal mangrove dynamics in relation to coastal development in Pacific Panama. J. Environ. Manag. 2005, 76, 263–276. [Google Scholar] [CrossRef]
- Mohamed, M.O.S.; Neukermans, G.; Kairo, J.G.; Dahdouh-Guebas, F.; Koedam, N. Mangrove forests in a peri-urban setting: The case of Mombasa (Kenya). Wetl. Ecol. Manag. 2008, 17, 243–255. [Google Scholar] [CrossRef] [Green Version]
- Nortey, D.D.N.; Aheto, D.W.; Blay, J.; Jonah, F.; Asare, N.K. Comparative Assessment of Mangrove Biomass and Fish Assemblages in an Urban and Rural Mangrove Wetlands in Ghana. Wetlands 2016, 36, 717–730. [Google Scholar] [CrossRef]
- Zamprogno, G.C.; Tognella, M.M.P.; Quaresma, V.D.S.; da Costa, M.B.; Pascoalini, S.S.; Couto, G.F.D. The structural heterogeneity of an urbanised mangrove forest area in southeastern Brazil: Influence of environmental factors and anthropogenic stressors. Braz. J. Oceanogr. 2016, 64, 157–172. [Google Scholar] [CrossRef] [Green Version]
- DasGupta, R.; Shaw, R. Changing perspectives of mangrove management in India—An analytical overview. Ocean Coast. Manag. 2013, 80, 107–118. [Google Scholar] [CrossRef]
- Pham, T.D.; Yoshino, K. Impacts of mangrove management systems on mangrove changes in the Northern Coast of Vietnam. Tropics 2016, 24, 141–151. [Google Scholar] [CrossRef] [Green Version]
- Friess, D.A.; Richards, D.R.; Phang, V.X.H. Mangrove forests store high densities of carbon across the tropical urban landscape of Singapore. Urban Ecosyst. 2015, 19, 1–16. [Google Scholar] [CrossRef]
- Lugo, A.E.; Snedaker, S.C. The Ecology of Mangroves. Annu. Rev. Ecol. Syst. 1974, 5, 39–64. [Google Scholar] [CrossRef]
- Wolanski, E.; Mazda, Y.; Ridd, P. Mangrove Hydrodynamics. In Tropical Mangrove Ecosystems; Robertson, A.I., Alongi, D.M., Eds.; American Geophyscial Union: Washington, DC, USA, 1993; pp. 43–62. [Google Scholar]
- Krauss, K.W.; Doyle, T.W.; Twilley, R.R.; Rivera-Monroy, V.H.; Sullivan, J.K. Evaluating the relative contributions of hydroperiod and soil fertility on growth of south Florida mangroves. Hydrobiologia 2006, 569, 311–324. [Google Scholar] [CrossRef]
- McKee, K.L. Seedling recruitment patterns in a Belizean mangrove forest: Effects of establishment ability and physico-chemical factors. Oecologia 1995, 101, 448–460. [Google Scholar] [CrossRef]
- Cardona, P.; Botero, L. Soil Characteristics and Vegetation Structure in a Heavily Deteriorated Mangrove Forest in the Caribbean Coast of Colombia. Biotropica 1998, 30, 24–34. [Google Scholar] [CrossRef]
- Feller, I.C.; Whigham, D.F.; McKee, K.L.; Lovelock, C.E. Nitrogen Limitation of Growth and Nutrient Dynamics in a Disturbed Mangrove Forest, Indian River Lagoon, Florida. Oecologia 2003, 134, 405–414. [Google Scholar] [CrossRef]
- Joshi, H.; Ghose, M. Forest Structure and Species Distribution Along Soil Salinity and Ph Gradient in Mangrove Swamps of the Sundarbans. Trop. Ecol. 2003, 44, 195–204. [Google Scholar]
- McKee, K.L. Growth and Physiological Responses of Neotropical Mangrove Seedlings to Root Zone Hypoxia. Tree Physiol. 1996, 16, 883–889. [Google Scholar] [CrossRef] [Green Version]
- Reef, R.; Feller, I.C.; Lovelock, C.E. Nutrition of mangroves. Tree Physiol. 2010, 30, 1148–1160. [Google Scholar] [CrossRef] [Green Version]
- Marois, D.E.; Mitsch, W.J. A mangrove creek restoration plan utilizing hydraulic modeling. Ecol. Eng. 2017, 108, 537–546. [Google Scholar] [CrossRef]
- Seguinot-Barbosa, J. La Ecologia Urbana de San Juan. Una Interpretación Geográfica-Social. Anales Geografía Universidad Complutense 1996, 16, 161–184. [Google Scholar]
- Tian-Hong, L.; Peng, H.; Zhi-Jie, Z. Impact Analysis of Coastal Engineering Projects on Mangrove Wetland Area Change with Remote Sensing. China Ocean Eng 2008, 22, 347–358. [Google Scholar]
- Brandeis, T.J.; Escobedo, F.J.; Staudhammer, C.L.; Nowak, D.J.; Zipperer, W.C. San Juan Bay Estuary Watershed Urban Forest Inventory; Gen. Tech. Rep. SRS-GTR-190; USDA-Forest Service, Southern Research Station: Asheville, NC, USA, 2014; pp. 1–44. [Google Scholar]
- Martinuzzi, S.; Gould, W.A.; Lugo, A.E.; Medina, E. Conversion and recovery of Puerto Rican mangroves: 200 years of change. For. Ecol. Manag. 2009, 257, 75–84. [Google Scholar] [CrossRef]
- Branoff, B.L. Mangrove Disturbance and Response Following the 2017 Hurricane Season in Puerto Rico. Chesap. Sci. 2019, 43, 1248–1262. [Google Scholar] [CrossRef]
- McIntyre, N.E.; Knowles-Yánez, K.; Hope, D. Urban ecology as an interdisciplinary field: Differences in the use of “urban” between the social and natural sciences. Urban Ecosyst. 2000, 4, 5–24. [Google Scholar] [CrossRef]
- MacGregor-Fors, I. Misconceptions or misunderstandings? On the standardization of basic terms and definitions in urban ecology. Landsc. Urban Plan. 2011, 100, 347–349. [Google Scholar] [CrossRef]
- Alberti, M.; Botsford, E.; Cohen, A. Quantifying the Urban Gradient: Linking Urban Planning and Ecology. In Avian Ecology and Conservation in an Urbanizing World; Springer: Berlin/Heidelberg, Germany, 2001; pp. 89–115. [Google Scholar]
- McDonnell, M.; Hahs, A. The use of gradient analysis studies in advancing our understanding of the ecology of urbanizing landscapes: Current status and future directions. Landsc. Ecol. 2008, 23, 1143–1155. [Google Scholar] [CrossRef]
- Ellison, A.M.; Farnsworth, E.J. Anthropogenic Disturbance of Caribbean Mangrove Ecosystems: Past Impacts, Present Trends, and Future Predictions. Biotropica 1996, 28, 549–565. [Google Scholar] [CrossRef]
- Lugo, A.E.; Medina, E.; McGinley, K. Issues and Challenges of Mangrove conservation in the Anthropocene. Madera Bosques 2014, 20, 11–38. [Google Scholar] [CrossRef] [Green Version]
- Branoff, B.L. Urban Mangrove Biology and Ecology: Emergent Patterns and Management Implications. In Threats to Mangrove Forests; Springer: Berlin/Heidelberg, Germany, 2018; pp. 521–537. [Google Scholar]
- Rodríguez-Martínez, J.; Soler-López, L.R. Hydrogeology and Hydrology of the Punta Cabullones Wetland Area, Ponce, Southern Puerto Rico, 2007–08; United States Geological Survey: Reston, VA, USA, 2014. [Google Scholar]
- Branoff, B.L. The role of urbanization in the flooding and surface water chemistry of Puerto Rico’s mangroves. Hydrol. Sci. J. 2020, 65, 1326–1343. [Google Scholar] [CrossRef]
- US DOC/NOAA/NWS/NDBC > National Data Buoy Center. Meteorological and Oceanographic Data Collected from the National Data Buoy Center Coastal-Marine Automated Network (C-Man) and Moored (Weather) Buoys. Station Id: 9755371. 1971. Available online: https://tidesandcurrents.noaa.gov/stationhome.html?id=9755371#info (accessed on 7 July 2020).
- US DOC/NOAA/NWS/NDBC > National Data Buoy Center. Meteorological and Oceanographic Data Collected from the National Data Buoy Center Coastal-Marine Automated Network (C-Man) and Moored (Weather) Buoys. Station Id: 9759110. 1971. Available online: https://tidesandcurrents.noaa.gov/stationhome.html?id=9759110 (accessed on 7 July 2020).
- Sparks, J.C.; Masters, R.; Payton, M. Comparative Evaluation of Accuracy and Efficiency of Six Forest Sampling Methods. Proc. Okla. Acad. Sci. 2002, 82, 49–56. [Google Scholar]
- McKee, K.L.; Mendelssohn, I.A.; Hester, M.W. Reexamination of Pore Water Sulfide Concentrations and Redox Potentials Near the Aerial Roots of Rhizophora mangle and Avicennia germinans. Am. J. Bot. 1988, 75, 1352–1359. [Google Scholar] [CrossRef]
- Santos, J.; Al-Azzawi, M.; Aronson, J.; Flowers, T.J. Ehaloph a Database of Salt-Tolerant Plants: Helping put Halophytes to Work. Plant Cell Physiol. 2015, 57, e10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fromard, F.; Puig, H.; Mougin, E.; Marty, G.; Betoulle, J.L.; Cadamuro, L. Structure, above-ground biomass and dynamics of mangrove ecosystems: New data from French Guiana. Oecologia 1998, 115, 39–53. [Google Scholar] [CrossRef] [PubMed]
- Imbert, D.; Rollet, B. Phytomasse Aérienne Et Produciton Primaire Dans La Mangrove Du Grand Cul-De-Sac Marin (Guadeloupe, Antilles Françaises). Bull. Ecol. 1989, 20, 27–39. [Google Scholar]
- Smith, T.J.; Whelan, K.R.T. Development of allometric relations for three mangrove species in South Florida for use in the Greater Everglades Ecosystem restoration. Wetl. Ecol. Manag. 2006, 14, 409–419. [Google Scholar] [CrossRef]
- Chave, J.; Andalo, C.; Brown, S.; Cairns, M.A.; Chambers, J.Q.; Eamus, D.; Fölster, H.; Fromard, F.; Higuchi, N.; Kira, T. Tree allometry and improved estimation of carbon stocks and balance in tropical forests. Oecologia 2005, 145, 87–99. [Google Scholar] [CrossRef]
- Reyes, G.; Brown, S.; Chapman, J.; Lugo, A.E. Wood Densities of Tropical Tree Species; US Department of Agriculture, Forest Service, Southern Forest Experiment Station: New Orleans, LA, USA, 1992; p. 88.
- Cook, B.D.; Corp, L.W.; Nelson, R.F.; Middleton, E.M.; Morton, D.C.; McCorkel, J.T.; Masek, J.G.; Ranson, K.J.; Ly, V.; Montesano, P.M. Nasa Goddard’s Lidar, Hyperspectral and Thermal (G-Liht) Airborne Imager. Remote Sens. 2013, 5, 4045–4066. [Google Scholar] [CrossRef] [Green Version]
- Alonzo, M.; Bookhagen, B.; McFadden, J.P.; Sun, A.; Roberts, D.A. Mapping urban forest leaf area index with airborne lidar using penetration metrics and allometry. Remote. Sens. Environ. 2015, 162, 141–153. [Google Scholar] [CrossRef]
- Giannico, V.; Lafortezza, R.; John, R.; Sanesi, G.; Pesola, L.; Chen, J. Estimating Stand Volume and Above-Ground Biomass of Urban Forests Using LiDAR. Remote. Sens. 2016, 8, 339. [Google Scholar] [CrossRef] [Green Version]
- Yan, J.; Valdez, E.A.; Trivedi, P.K.; Zimmer, D.M. A Language and Environment for Statistical Computing. R Found. Stat. Comput. 2011, 1, 409. [Google Scholar]
- Lidr: Airborne Lidar Data Manipulation and Visualization for Forestry Applications R Package Version 3.0.2. Available online: https://CRAN.R-project.org/package=lidR (accessed on 9 October 2020).
- Rlidar: Lidar Data Processing and Visualization R Package Version 0.1.1. Available online: https://CRAN.R-project.org/package=rLiDAR (accessed on 9 October 2020).
- Andersen, H.E.; Mcgaughey, R.J.; Reutebuch, S.E. Assessing the influence of flight parameters, interferometric processing, slope and canopy density on the accuracy of X-band IFSAR-derived forest canopy height models. Int. J. Remote. Sens. 2008, 29, 1495–1510. [Google Scholar] [CrossRef] [Green Version]
- Pennington, D.N.; Hansel, J.; Blair, R.B. The conservation value of urban riparian areas for landbirds during spring migration: Land cover, scale, and vegetation effects. Biol. Conserv. 2008, 141, 1235–1248. [Google Scholar] [CrossRef]
- Frimpong, E.A.; Sutton, T.M.; Lim, K.J.; Hrodey, P.J.; Engel, B.A.; Simon, T.P.; Lee, J.G.; Le Master, D.C. Determination of optimal riparian forest buffer dimensions for stream biota–landscape association models using multimetric and multivariate responses. Can. J. Fish. Aquat. Sci. 2005, 62, 1–6. [Google Scholar] [CrossRef]
- Pebesma, E. Simple Features for R: Standardized Support for Spatial Vector Data. R J. 2018, 10, 439–446. [Google Scholar] [CrossRef] [Green Version]
- Raster: Geographic Data Analysis and Modeling R Package Version 2.5-8. Available online: https://CRAN.R-project.org/package=raster (accessed on 9 October 2020).
- Office for Coastal Management. C-Cap Land Cover, Puerto Rico, 2010 from 15 June 2010 to 15 August 2018. Available online: https://inport.nmfs.noaa.gov/inport/item/48301 (accessed on 9 October 2020).
- Tiger/Line Shapefile. Available online: https://catalog.data.gov/dataset/tiger-line-shapefile-2015-nation-u-s-primary-roads-national-shapefile (accessed on 9 October 2020).
- American Community Survey Census Tract Estimates for Puerto Rico. Available online: https://www.census.gov/geographies/mapping-files/time-series/geo/tiger-data.html (accessed on 9 October 2020).
- Vegan: Community Ecology Package R Package Version 2.5-1. Available online: https://CRAN.R-project.org/package=vegan (accessed on 9 October 2020).
- Glmulti: Model Selection and Multimodel Inference Made Easy. R Package Version 1.0.8. Available online: https://CRAN.R-project.org/package=glmulti (accessed on 9 October 2020).
- Olsrr: Tools for Building Ols Regression Models R Package Version 0.5.3. Available online: https://CRAN.R-project.org/package=olsrr (accessed on 9 October 2020).
- Grömping, U. Relative Importance for Linear Regression in R: The Package Relaimpo. J. Stat. Software 2006, 17, 1–27. [Google Scholar] [CrossRef] [Green Version]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2009. [Google Scholar]
- Clarke, K.R.; Ainsworth, M.A. Method of linking multivariate community structure to environmental variables. Mar. Ecol. Prog. Ser. 1993, 92, 205–219. [Google Scholar] [CrossRef]
- Bettez, N.D.; Marino, R.; Howarth, R.W.; Davidson, E.A. Roads as nitrogen deposition hot spots. Biogeochemistry 2013, 114, 149–163. [Google Scholar] [CrossRef]
- Brush, G.S. Historical Land Use, Nitrogen, and Coastal Eutrophication: A Paleoecological Perspective. Chesap. Sci. 2008, 32, 18–28. [Google Scholar] [CrossRef] [Green Version]
- Davidson, E.A.; Savage, K.E.; Bettez, N.D.; Marino, R.; Howarth, R.W. Nitrogen in Runoff from Residential Roads in a Coastal Area. Water Air Soil Pollut. 2009, 210, 3–13. [Google Scholar] [CrossRef]
- Agraz-Hernández, C.M.; del Río-Rodríguez, R.E.; Chan-Keb, C.A.; Osti-Saenz, J.; Muñiz-Salazar, R. Nutrient Removal Efficiency of Rhizophora mangle (L.) Seedlings Exposed to Experimental Dumping of Municipal Waters. Diversity 2018, 10, 16. [Google Scholar] [CrossRef] [Green Version]
- Naidoo, G. Differential effects of nitrogen and phosphorus enrichment on growth of dwarf Avicennia marina mangroves. Aquat. Bot. 2009, 90, 184–190. [Google Scholar] [CrossRef]
- Chen, R.; Twilley, R.R. A gap dynamic model of mangrove forest development along gradients of soil salinity and nutrient resources. J. Ecol. 1998, 86, 37–51. [Google Scholar] [CrossRef]
- Chen, R.; Twilley, R.R. Patterns of mangrove forest structure and soil nutrient dynamics along the Shark River Estuary, Florida. Estuaries 1999, 22, 955–970. [Google Scholar] [CrossRef]
- Ball, M.C. Patterns of secondary succession in a mangrove forest of Southern Florida. Oecologia 1980, 44, 226–235. [Google Scholar] [CrossRef]
- Ball, M.; Cowan, I.R.; Farquhar, G.D. Maintenance of Leaf Temperature and the Optimisation of Carbon Gain in Relation to Water Loss in a Tropical Mangrove Forest. Funct. Plant Biol. 1988, 15, 263–276. [Google Scholar] [CrossRef]
- Medina, E. Mangrove Physiology: The Challenge of Salt, Heat, and Light Stress under Recurrent Flooding. In Ecosistemas De Manglar En América Tropical; Yánez-Arancibia, A., Lara-Domínguez, A.L., Eds.; Instituto de Ecologia, AC; UICN/ORMA; NOAA/NMFS: Xalapa, Mexico, 1999; pp. 109–126. [Google Scholar]
- Miller, P.C. Bioclimate, Leaf Temperature, and Primary Production in Red Mangrove Canopies in South Florida. Ecology 1972, 53, 22–45. [Google Scholar] [CrossRef]
- Vila-Ruiz, C.P.; Meléndez-Ackerman, E.; Santiago-Bartolomei, R.; Garcia-Montiel, D.; Lastra, L.; Figuerola, C.E.; Fumero-Cabán, J. Plant species richness and abundance in residential yards across a tropical watershed: Implications for urban sustainability. Ecol. Soc. 2014, 19. [Google Scholar] [CrossRef] [Green Version]
- Berger, U.; Adams, M.; Grimm, V.; Hildenbrandt, H. Modelling secondary succession of neotropical mangroves: Causes and consequences of growth reduction in pioneer species. Perspect. Plant Ecol. Evol. Syst. 2006, 7, 243–252. [Google Scholar] [CrossRef]
- Krauss, K.W.; Lovelock, C.E.; McKee, K.L.; López-Hoffman, L.; Ewe, S.M.; Sousa, W.P. Environmental drivers in mangrove establishment and early development: A review. Aquat. Bot. 2008, 89, 105–127. [Google Scholar] [CrossRef]
- Odum, W.E.; McIvor, C.C. Mangroves. In Ecosystems of Florida; Myers, R.E., Ewel, J.J., Eds.; University Press of Florida: Gainesville, FL, USA, 1990; pp. 517–548. [Google Scholar]
- Nagelkerken, I.; Blaber, S.; Bouillon, S.; Green, P.; Haywood, M.; Kirton, L.; Meynecke, J.O.; Pawlik, J.; Penrose, H.; Sasekumar, A.; et al. The habitat function of mangroves for terrestrial and marine fauna: A review. Aquat. Bot. 2008, 89, 155–185. [Google Scholar] [CrossRef] [Green Version]
- Baird, R.C. Coastal urbanization: The challenge of management lag. Manag. Environ. Qual. Int. J. 2009, 20, 371–382. [Google Scholar]
- McGranahan, G.; Balk, D.; Anderson, B. The rising tide: Assessing the risks of climate change and human settlements in low elevation coastal zones. Environ. Urban. 2007, 19, 17–37. [Google Scholar] [CrossRef]
- Aburto-Oropeza, O.; Ezcurra, E.; Danemann, G.; Valdez, V.; Murray, J.; Sala, E. Mangroves in the Gulf of California Increase Fishery Yields. Proc. Natl. Acad. Sci. USA 2008, 105, 10456–10459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alongi, D.M. Mangrove forests: Resilience, protection from tsunamis, and responses to global climate change. Estuarine Coast. Shelf Sci. 2008, 76, 1–13. [Google Scholar] [CrossRef]
- Rönnbäck, P.; Troell, M.; Kautsky, N.; Primavera, J. Distribution Pattern of Shrimps and Fish AmongAvicenniaandRhizophoraMicrohabitats in the Pagbilao Mangroves, Philippines. Estuarine Coast. Shelf Sci. 1999, 48, 223–234. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Water-shed | Site | Lon. | Lat. | Urban Index | Urban Cover | Street Dens. | Pop. Dens. | Mangrove Cover | Urban: Mangrove |
|---|---|---|---|---|---|---|---|---|---|
| San Juan Bay Estuary Río Bayamón/ Río Hondo to The Río Puerto Nuevo/ Río Piedras | BAHMIN | −66.1302 | 18.44985 | 29 | 0.09 | 1.1 | 301.3 | 0.04 | 2.7 |
| BAHMAX | −66.0827 | 18.44436 | 63 | 0.45 | 6.4 | 2330.7 | 0.10 | 4.5 | |
| MPDMIN | −66.0788 | 18.43787 | 60 | 0.44 | 6.1 | 1338.2 | 0.20 | 2.1 | |
| MPDMAX | −66.0636 | 18.43329 | 73 | 0.59 | 11.3 | 3450.0 | 0.16 | 3.6 | |
| MPNMIN | −66.0591 | 18.43373 | 82 | 0.65 | 15.3 | 4317.9 | 0.11 | 6.1 | |
| MPNMAX | −66.0498 | 18.4307 | 98 | 0.75 | 19.4 | 7253.6 | 0.05 | 13.7 | |
| SANMAX | −66.0342 | 18.44253 | 46 | 0.30 | 5.3 | 2568.7 | 0.13 | 2.3 | |
| SANMIN | −66.0125 | 18.41686 | 35 | 0.21 | 4.3 | 1418.0 | 0.09 | 2.3 | |
| SUAMIN | −65.9957 | 18.42838 | 56 | 0.54 | 3.6 | 1460.4 | 0.10 | 5.4 | |
| SUAMAX | −65.9881 | 18.42849 | 64 | 0.60 | 6.8 | 2426.6 | 0.10 | 6.1 | |
| TORMIN | −65.9829 | 18.44735 | 14 | 0.10 | 1.3 | 376.2 | 0.32 | 0.3 | |
| TORMAX | −65.9782 | 18.43906 | 19 | 0.16 | 3.4 | 606.8 | 0.33 | 0.5 | |
| PINMAX | −65.9570 | 18.44315 | 5 | 0.06 | 1.8 | 115.2 | 0.55 | 0.1 | |
| PINMIN | −65.9572 | 18.43104 | 1 | 0.00 | 0.3 | 0.0 | 0.50 | 0.0 | |
| Levittown Río de la Plata | LEVMIN | −66.2068 | 18.46477 | 9 | 0.02 | 0.7 | 29.1 | 0.42 | 0.0 |
| LEVMID | −66.1961 | 18.45779 | 24 | 0.18 | 4.1 | 1942.2 | 0.41 | 0.4 | |
| LEVMAX | −66.1906 | 18.45468 | 48 | 0.40 | 9.3 | 2597.0 | 0.23 | 1.8 | |
| Ponce—Río Inabón to the Río Loco | PONMID | −66.6716 | 17.97408 | 31 | 0.13 | 1.4 | 321.2 | 0.12 | 1.0 |
| PONMAX | −66.6095 | 17.97253 | 42 | 0.37 | 3.1 | 598.8 | 0.15 | 2.4 | |
| PONMIN | −66.5833 | 17.96285 | 21 | 0.00 | 0.3 | 0.2 | 0.19 | 0.0 |
| Variable | Units | Description | Source | |
|---|---|---|---|---|
| Ground | dbh | cm | The diameter at breast height of each woody stem within the plot | Present study |
| Stem density | stems/ha | The number of stems over 1 cm in dbh within the plot over the area of the plot | Present study | |
| Basal area | m2/ha | The sum of cross-sectional areas: of all trees in the plot over the area of the plot | Present study | |
| Biomass | Mg/ha | The sum of biomass estimates from allometric equations for all trees in the plot over the area of the plot | Present study | |
| Percent stand biomass composition of spp. | % | The total biomass of a species at a plot over the total biomass of all species at that plot | Present study | |
| Diversity | n | The total number of all species encountered in a plot | Present study | |
| Mangrove Diversity | n | The total number of all true mangrove species encountered in a plot | Present study | |
| Soil Porewater Salinity | ppt | Porewater salinity measurements at soil depths of 0, 10, and 20 cm | Present study | |
| LiDAR | Mean height | m | Mean height of all returns above 1.4 m | NASA G-LiHT a |
| Height SD | m | Standard deviation of all return heights above 1.4 m | NASA G-LiHT a | |
| Height percentiles | m | Heights at which 10%, 20%...90%, 100% of returns are lower | NASA G-LiHT a | |
| Height skewness | m | Skewness of all return heights above 1.4 m, a measure of vertical height complexity | NASA G-LiHT a | |
| Canopy cover | % | Number of first returns above 2 m over the total number of first returns | NASA G-LiHT a | |
| Canopy density | % | Number of all returns above 2 m over the total number of all returns | NASA G-LiHT a | |
| Variable | Units | Description | Source | |
|---|---|---|---|---|
| Urbanness | Urban cover | km2/km2 | Aerial coverage of all urban and developed open space classes | LULC raster a |
| Green and blue cover | km2/km2 | Aerial coverage of all open water and vegetation classes | ||
| Mangrove cover | km2/km2 | Aerial coverage of all estuarine forested and estuarine scrub–shrub wetland classes | ||
| Road density | km/km2 | Length of roads over area of sampling circle | Road network vector b | |
| Population density | people/km2 | Number of people within the sampling circle over area of sampling circle | Census tract total population c | |
| Urban index | - | Normalized mean of the above urbanization variables such that 0 is the least urban and 100 is the most urban value | Above urbanness variables d | |
| Hydrology | Average depth | m | Mean depth of water at the site from 2012 to 2017 based on water level models | Water level models d |
| Proportion of time flooded | % | Proportion of time with positive depth at the site from 2012 to 2017 based on water level models | ||
| Mean daily flood frequency | floods/ day | Mean number of times per day a positive depth was estimated at the site from 2012 to 2017 based on water level models | ||
| Mean flood length | day | Mean length of positive depth estimated at the site from 2012 to 2017 based on water level models | ||
| Water chemistry | Dissolved oxygen | mg/L | Mean concentration of dissolved oxygen in surface waters measured monthly between 2012 and 2017 | San Juan Bay Estuary Program e |
| pH | - | Mean pH in surface waters measured monthly between 2012 and 2017 | ||
| Salinity | PSS | Mean salinity in surface waters measured monthly between 2012 and 2017 | ||
| Temperature | °C | Mean temperature in surface waters measured monthly between 2012 and 2017 | ||
| Ammonium | mg/L | Mean concentration of ammonium in surface waters measured bi-annually between 2012 and 2017 | ||
| Total Kjeldahl Nitrogen | mg/L | Mean concentration of total Kjeldahl nitrogen in surface waters measured bi-annually between 2012 and 2017 | ||
| Nitrate and nitrite | mg/L | Mean combined concentration of nitrate and nitrite in surface waters measured bi-annually between 2012 and 2017 | ||
| Total phosphorus | mg/L | Mean concentration of total phosphorus in surface waters measured bi-annually between 2012 and 2017 |
| San Juan Bay Estuary | Levittown | Ponce | All | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Species | Stems/ha | Basal Area m2/ha | Biomass Mg/ha | Stems/ha | Basal Area m2/ha | Biomass Mg/ha | Stems/ha | Basal Area m2/ha | Biomass Mg/ha | Stems/ha | Basal Area m2/ha | Biomass Mg/ha |
| All | 5758 ± 282.5 | 38.6 ± 1.6 | 207.9 ± 9.8 | 7724.3 ± 1242.8 | 42.6 ± 4.7 | 245.6 ± 33.5 | 5356.4 ± 632.1 | 27.7 ± 2.2 | 156 ± 14.2 | 5997.1 ± 291.2 | 37.6 ± 1.4 | 206 ± 8.9 |
| Laguncularia racemosa†22 | 2979.4 ± 275.8 | 28.1 ± 1.8 | 144.4 ± 10.1 | 6695.5 ± 1350.3 | 33.9 ± 4.4 | 186.4 ± 29.2 | 2569.6 ± 719.4 | 16.2 ± 3.8 | 86.1 ± 22.2 | 3556.9 ± 329.3 | 27.6 ± 1.6 | 144.1 ± 9.2 |
| Rhizophora mangle†SW | 2219.9 ± 189.3 | 9.2 ± 1 | 54.7 ± 7.8 | 463 ± 94.5 | 11.3 ± 3.8 | 101.5 ± 39.3 | 4165.3 ± 641.6 | 19.7 ± 1.7 | 115.4 ± 11.2 | 2366.3 ± 187.4 | 10.9 ± 0.9 | 66.9 ± 7.1 |
| Avicennia germinans†67 | 2062.6 ± 383.7 | 11.3 ± 1.5 | 59.7 ± 8.2 | - | - | - | 827.6 ± 355.3 | 3.5 ± 2 | 22.4 ± 16.1 | 1876.2 ± 334.8 | 10.1 ± 1.3 | 54.1 ± 7.5 |
| Thespesia populnea†SW | 2132.7 ± 524.7 | 7.3 ± 2.3 | 35 ± 11.7 | 3910.7 ± 687.2 | 19.5 ± 4 | 94.8 ± 22.4 | 1082.3 ± 191 | 0.2 ± 0 | 0.4 ± 0.1 | 2412.3 ± 432.8 | 9.3 ± 2.1 | 44.5 ± 10.6 |
| Calophyllum sp. | 1122.8 ± 323.8 | 1.9 ± 0.8 | 6.7 ± 3 | 286.5 ± 120.5 | 0.1 ± 0.0 | 0.2 ± 0.0 | - | - | - | 899.8 ± 256 | 1.4 ± 0.6 | 5 ± 2.3 |
| Dalbergia ecastaphyllum†22 | 615.4 ± 300.3 | 0.5 ± 0.3 | 1.4 ± 0.8 | 418.4 ± 110.1 | 0.6 ± 0.2 | 2.1 ± 0.6 | - | - | - | 509.3 ± 146.3 | 0.6 ± 0.2 | 1.8 ± 0.5 |
| Bucida buceras | 382 ± 76.8 | 0.5 ± 0.1 | 1.7 ± 0.5 | - | - | - | - | - | - | 382 ± 76.8 | 0.5 ± 0.1 | 1.7 ± 0.5 |
| unknown fabaceae | 848.8 ± 404.9 | 6 ± 3.1 | 30.4 ± 16 | - | - | - | - | - | - | 848.8 ± 404.9 | 6 ± 3.1 | 30.4 ± 16 |
| Jacquinia arborea | - | - | - | - | - | - | 483.8 ± 84.5 | 0.2 ± 0.1 | 0.5 ± 0.3 | 483.8 ± 84.5 | 0.2 ± 0.1 | 0.5 ± 0.3 |
| Roystonea borinquena | 291 ± 86.6 | 11.8 ± 6.4 | 88.6 ± 51.1 | 127.3 ± - | 2.9 ± - | 16.3 ± - | - | - | - | 270.6 ± 77.7 | 10.7 ± 5.7 | 79.6 ± 45.1 |
| Schefflera morototoni | 466.9 ± 278.3 | 1.1 ± 0.8 | 4.1 ± 2.8 | 127.3 ± - | 0.1 ± - | 0.2 ± - | - | - | - | 382 ± 214.3 | 0.9 ± 0.6 | 3.1 ± 2.2 |
| Terminalia catappa† | 350.1 ± 131.2 | 7.9 ± 3.5 | 52.5 ± 23.5 | 127.3 ± - | 2.7 ± - | 15.1 ± - | - | - | - | 305.6 ± 111 | 6.9 ± 2.9 | 45 ± 19.7 |
| Ardisia elliptica† | 466.9 ± 212.2 | 0.1 ± 0.1 | 0.4 ± 0.3 | - | - | - | - | - | - | 466.9 ± 212.2 | 0.1 ± 0.1 | 0.4 ± 0.3 |
| Cocos nuciferaδ6 | 169.8 ± 42.4 | 7.6 ± 1.7 | 54.7 ± 13.8 | 127.3 ± 0.0 | 7.6 ± 0.9 | 53.6 ± 7.3 | - | - | - | 159.2 ± 31.8 | 7.6 ± 1.3 | 54.4 ± 10.2 |
| Mammea americana | 159.2 ± 31.8 | 0.1 ± 0.1 | 0.3 ± 0.2 | - | - | - | - | - | - | 159.2 ± 31.8 | 0.1 ± 0.1 | 0.3 ± 0.2 |
| Andira inermis | 254.6 ± 127.3 | 0.1 ± 0.0 | 0.3 ± 0.2 | - | - | - | - | - | - | 254.6 ± 127.3 | 0.1 ± 0 | 0.3 ± 0.2 |
| Coccoloba uviferaδ32 | 509.3 ± - | 0.3 ± - | 0.8 ± - | - | - | - | - | - | - | 509.3 ± - | 0.3 ± - | 0.8 ± - |
| Tabebuia heterophylla | 509.3 ± - | 0.2 ± - | 0.5 ± - | - | - | - | - | - | - | 509.3 ± - | 0.2 ± - | 0.5 ± - |
| Annona glabra | - | - | - | 382 ± - | 5.4 ± - | 27.3 ± - | - | - | - | 382 ± - | 5.4 ± - | 27.3 ± - |
| Conocarpus erectus†SW | 254.6 ± - | 11 ± - | 76 ± - | - | - | - | 127.3 ± - | 3.8 ± - | 23.2 ± - | 191 ± 63.7 | 7.4 ± 3.6 | 49.6 ± 26.4 |
| Hippomane mancinellaδ | 382 ± - | 0.1 ± - | 0.2 ± - | - | - | - | - | - | - | 382 ± - | 0.1 ± - | 0.2 ± - |
| Pavonia fruticosa | 382 ± - | 0.1 ± - | 0.2 ± - | - | - | - | - | - | - | 382 ± - | 0.1 ± - | 0.2 ± - |
| Paullinia pinnata | 127.3 ± 0 | 0.9 ± 0.9 | 4.7 ± 4.5 | - | - | - | - | - | - | 127.3 ± 0 | 0.9 ± 0.9 | 4.7 ± 4.5 |
| Relative Importance | |||||||
|---|---|---|---|---|---|---|---|
| Response | Model: Response ~ Land Cover + Flooding + Water Chemistry | BIC | R2 | p | Land Cover | Water Chem. | Flood ing |
| Stem Density (ha−1) | ~ 7500 * ln(Mangrove Cover Within 500 m) − 3549 * Ammonium | 225 | 0.61 | 0.001 | 30 | 70 ** | - |
| Basal Area (m2 ha−1) | ~ −0.003 * Population Density Within 1 km − 8.8 * Temperature + 5 × 10−4 * Flood Length | 78 | 0.76 | 0.0004 | 24 ** | 58 *** | 18 * |
| Above ground Biomass (Mg ha−1) | ~ 107 * log(Vegetation Cover Within 1 km) − 66 * Temperature + 0.001 * Flood Length | 116 | 0.82 | 0.02 | 19 ** | 68 *** | 13 |
| Max DBH (cm) | ~ 2 * log(Population Density Within 1 km) + 2.4 * log(Total Kjeldahl Nitrogen) − 0.004 * Flood Days per Year | 62 | 0.84 | 0.0004 | 18 ** | 54 *** | 28 * |
| Mean DBH (cm) | ~ 0.23 * log(Population Density Within 100 m) + 5.32 * Nitrate & Nitrite − 0.23 * log(Dry Length) | 40 | 0.62 | 0.04 | 30 * | 38 ** | 32 * |
| 90th Height Percentile (m) | ~ 14.4 * Mangrove Cover Within 500 m − 0.08 * Dissolved Oxygen − 0.002 * Flood Days per Year | 39 | 0.84 | 0.0002 | 48 *** | 28 ** | 24 ** |
| Height SD (m) | ~ −1.5 * log(Open Water Within 500 m) + 0.13 * Total Kjeldahl Nitrogen − 3 × 10−4 * Flood Days per Year | 3 | 0.77 | 0.0002 | 33 * | 48 *** | 19 |
| Canopy Cover (%) | ~ 2.9 * log(Open Water Within 1 km) + 5.44 * Nitrate & Nitrite − 0.001 * Flood Length | 39 | 0.82 | 0.0005 | 53 ** | 20 ** | 28 * |
| Canopy Density (%) | ~ 1612* Urban Cover Within 50 m − 4.61 * log(Soil Salinity 0 cm) + 8.84 * Daily Flood Frequency | 66 | 0.61 | 0.01 | 24 ** | 20 * | 56 ** |
| Percent stand biomass composition of A. germinans | ~ −67.13 * log(Urban Cover Within 500 m) + 17.77 * Temperature − 1.5 * log(Flood Length) | 97 | 0.85 | 0.0001 | 28 ** | 66 *** | 6 * |
| Percent stand biomass composition of R. mangle | −11.5 * log(Urban Index Within 200 m) + 1 * Soil Salinity at 20 cm | 125 | 0.55 | 0.005 | 52 * | - | 48 * |
| Percent stand biomass composition of L. racemosa | ~ −583 * log(Open Water Within 200 m) − 1.52 * Salinity − 145.5 * Max Depth | 103 | 0.86 | 0.0002 | 15 *** | 16 *** | 68 *** |
| Percent stand biomass composition of non-mangrove | ~ 18150 * Open Water Within 50 m − 25.3 * log(Nitrate & Nitrite) + 15.04 * log(Mean Depth+1) | 82 | 0.76 | 0.001 | 88 *** | 8 | 4 |
| Tree Diversity | ~ 1136 * log(Open Water Within 50 m) + 0.23 * Temperature − 0.34 * Daily Flood Frequency | 7 | 0.74 | 0.002 | 78 *** | 7 * | 14 ** |
| Mangrove Diversity | ~ −1.31 * log(Urban Cover Within 500 m) + 3.93 * log(Phosphorus) + 0.06 * log(Dry Length) | −1 | 0.72 | 0.002 | 54 * | 29 ** | 17 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Branoff, B.L.; Martinuzzi, S. The Structure and Composition of Puerto Rico’s Urban Mangroves. Forests 2020, 11, 1119. https://doi.org/10.3390/f11101119
Branoff BL, Martinuzzi S. The Structure and Composition of Puerto Rico’s Urban Mangroves. Forests. 2020; 11(10):1119. https://doi.org/10.3390/f11101119
Chicago/Turabian StyleBranoff, Benjamin L., and Sebastián Martinuzzi. 2020. "The Structure and Composition of Puerto Rico’s Urban Mangroves" Forests 11, no. 10: 1119. https://doi.org/10.3390/f11101119