The Effects of Wind Exposure on Scots Pine Trees: Within-Stem Variability of Wood Density and Mechanical Properties

 ,
,  ,
, 
 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Stand Selection
2.2. Selection of Model Trees
2.3. Sampling Procedure
2.4. Measurement of Wood Properties
2.5. Statistical Analyses
3. Results
3.1. Characteristics of Model Trees
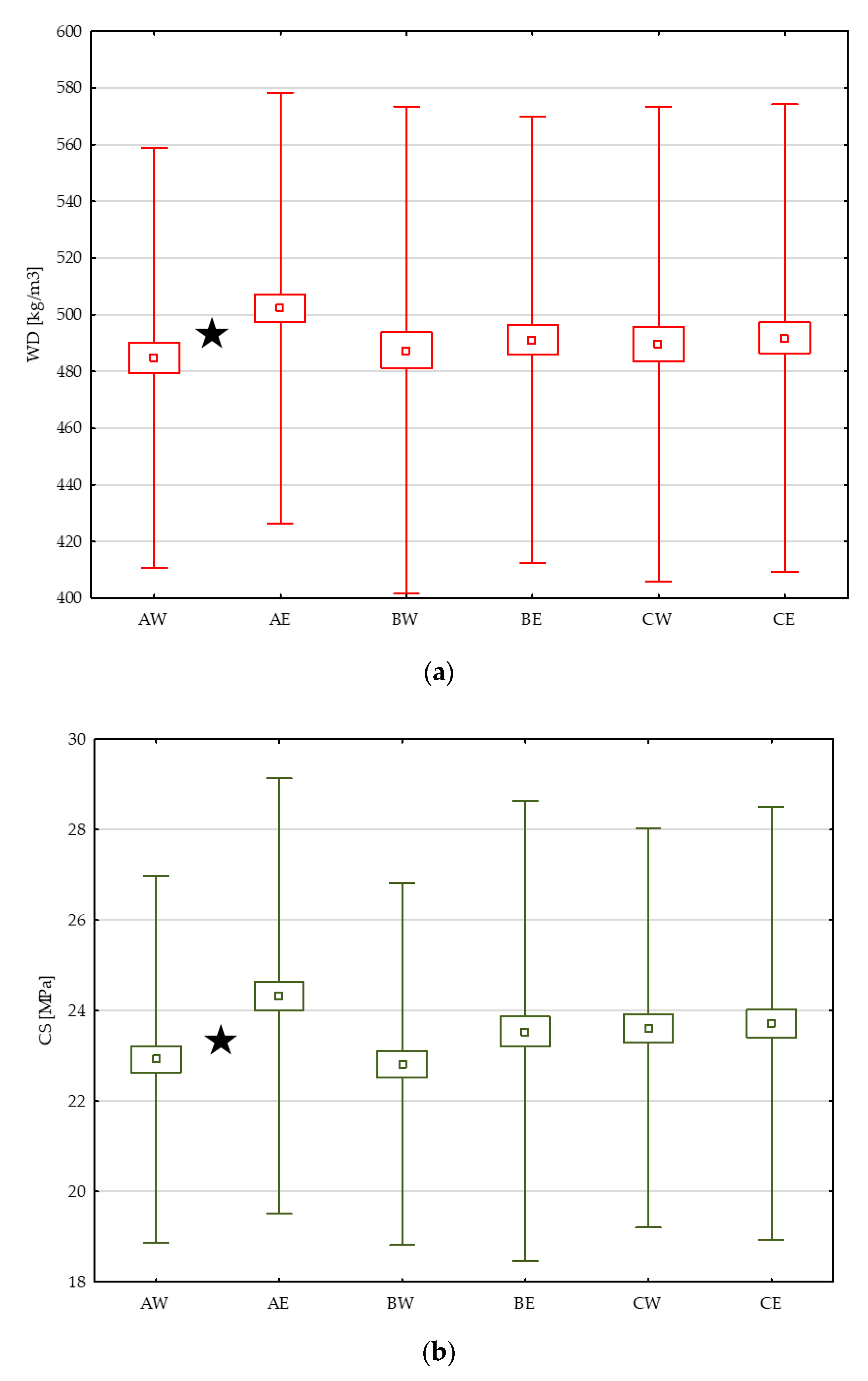
3.2. Differences of Wood Properties between Stand Edge and Forest Interior
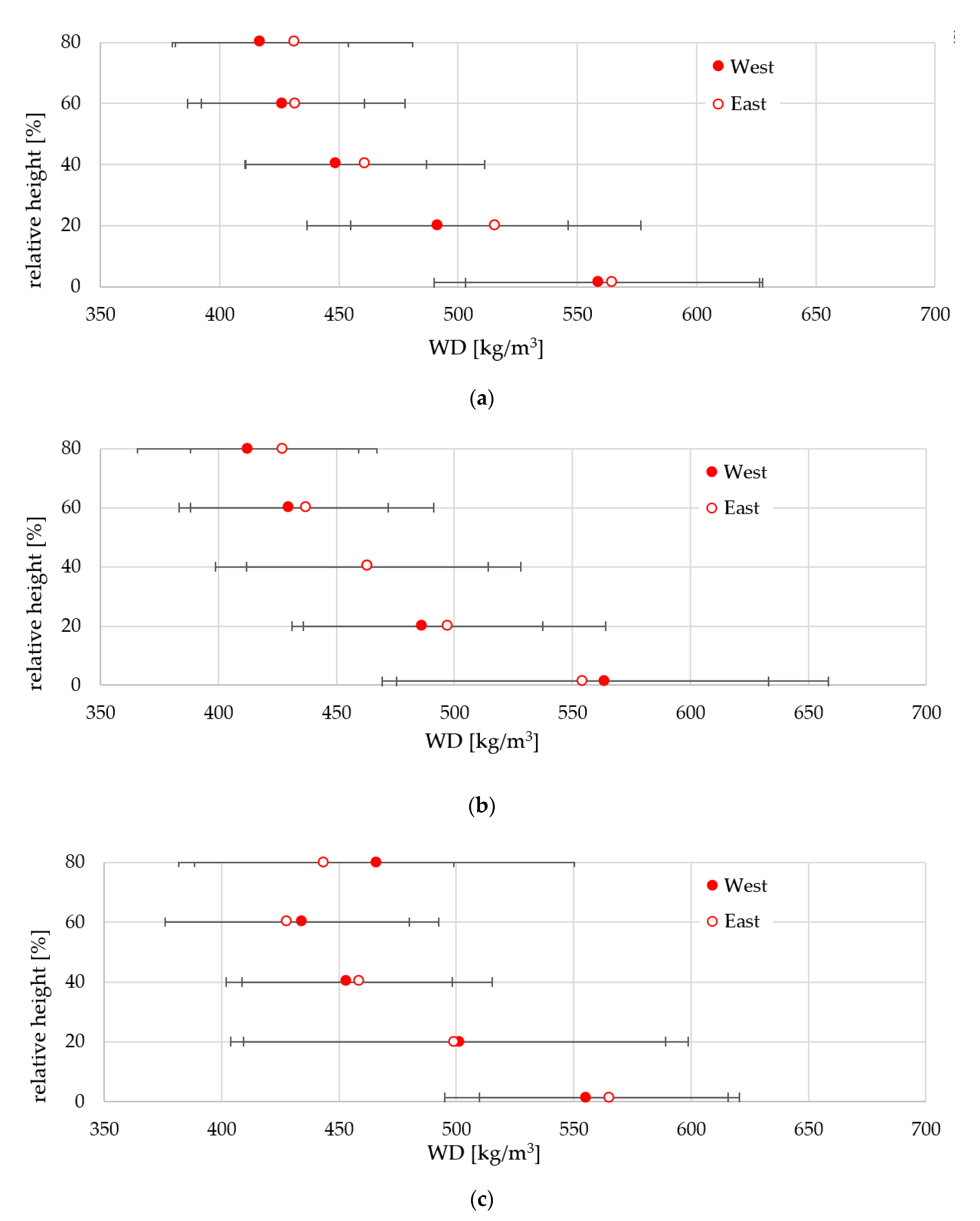
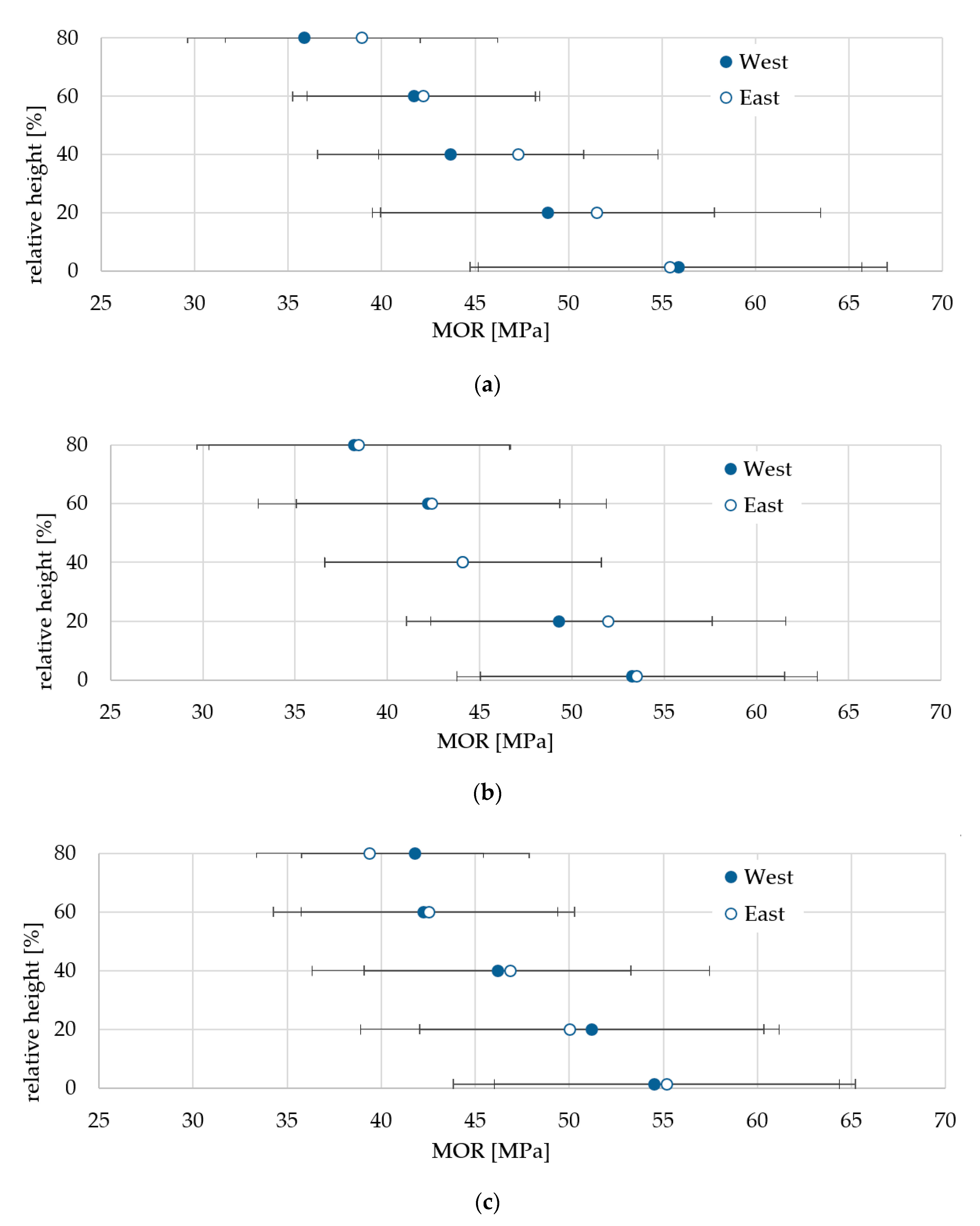
3.3. Variability of Wood Properties Across the Stem
3.4. Variability in Wood Properties Along the Stem
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ennos, A.R. Wind as an Ecological Factor. Trends Ecol. Evol. 1997, 12, 108–111. [Google Scholar] [CrossRef]
- Ulanova, N.G. The Effects of Windthrow on Forests at Different Spatial Scales: A Review. For. Ecol. Manag. 2000, 135, 155–167. [Google Scholar] [CrossRef]
- Tomczak, A.; Jelonek, T.; Jakubowski, M. Changes in the Structure and Properties of Wood as an Effect of the Impact of Wind on Trees. Sylwan 2012, 156, 776–783. [Google Scholar]
- Schelhaas, M.-J.; Nabuurs, G.-J.; Schuck, A. Natural Disturbances in the European Forests in the 19th and 20th Centuries. Glob. Chang. Biol. 2003, 9, 1620–1633. [Google Scholar] [CrossRef]
- Mitchell, S.J. Wind as a Natural Disturbance Agent in Forests: A Synthesis. Forestry 2013, 86, 147–157. [Google Scholar] [CrossRef] [Green Version]
- Gardiner, B.; Berry, P.; Moulia, B. Review: Wind Impacts on Plant Growth, Mechanics and Damage. Plant Sci. 2016, 245, 94–118. [Google Scholar] [CrossRef] [PubMed]
- Forzieri, G.; Pecchi, M.; Girardello, M.; Mauri, A.; Klaus, M.; Nikolov, C.; Rüetschi, M.; Gardiner, B.; Tomaštík, J.; Small, D.; et al. A Spatially Explicit Database of Wind Disturbances in European Forests Over the Period 2000–2018. Earth Syst. Sci. Data 2020, 12, 257–276. [Google Scholar] [CrossRef] [Green Version]
- Peltola, H.; Kellomäki, S.; Väisänen, H.; Ikonen, V.P. A Mechanistic Model for Assessing the Risk of Wind and Snow Damage to Single Trees and Stands of Scots Pine, Norway Spruce, and Birch. Can. J. For. Res. 1999, 22, 647–661. [Google Scholar] [CrossRef]
- Valinger, E.; Fridman, J. Models to Assess the Risk of Snow and Wind Damage in Pine, Spruce, and Birch Forests in Sweden. Environ. Manag. 1999, 24, 209–217. [Google Scholar] [CrossRef]
- Ancelin, P.; Courbaud, B.; Fourcaud, T. Development of an Individual Tree-Based Mechanical Model to Predict Wind Damage Within Forest Stands. For. Ecol. Manag. 2004, 203, 101–121. [Google Scholar] [CrossRef]
- Scott, R.E.; Mitchell, S.J. Empirical Modelling of Windthrow Risk in Partially Harvested Stands Using Tree, Neighbourhood, and Stand Attributes. For. Ecol. Manag. 2005, 218, 193–209. [Google Scholar] [CrossRef]
- Gardiner, B.; Byrne, K.; Hale, S.; Kamimura, K.; Mitchell, S.J.; Peltola, H.; Ruel, J.-C. A Review of Mechanistic Modelling of Wind Damage Risk to Forests. Forestry 2008, 81, 447–463. [Google Scholar] [CrossRef] [Green Version]
- Heinonen, T.; Pukkala, T.; Ikonen, V.-P.; Peltola, H.; Venäläinen, A.; Dupont, S. Integrating the Risk of Wind Damage into Forest Planning. For. Ecol. Manag. 2009, 258, 1567–1577. [Google Scholar] [CrossRef]
- Hanewinkel, M.; Peyron, J.L. The Economic Impact of Storms. In Living with Storm Damage to Forests: What Science Can Tell Us; Gardiner, B., Schuck, A., Schelhaas, M.-J., Orazio, C., Blennow, K., Nicoll, B., Eds.; European Forest Institute: Joensuu, Finland, 2013; pp. 55–63. [Google Scholar]
- Hale, S.E.; Gardiner, B.; Peace, A.; Nicoll, B.; Taylor, P.; Pizzirani, S. Comparison and Validation of Three Versions of a Forest Wind Risk Model. Environ. Model. Softw. 2015, 68, 27–41. [Google Scholar] [CrossRef] [Green Version]
- Anyomi, K.A.; Mitchell, S.J.; Ruel, J.-C. Windthrow Modelling in Old-Growth and Multi-Layered Boreal Forests. Ecol. Model. 2016, 327, 105–114. [Google Scholar] [CrossRef]
- Morimoto, J.; Nakagawa, K.; Takano, K.T.; Aiba, M.; Oguro, M.; Furukawa, Y.; Mishima, Y.; Ogawa, K.; Ito, R.; Takemi, T.; et al. Comparison of Vulnerability to Catastrophic Wind Between Abies Plantation Forests and Natural Mixed Forests in Northern Japan. For. Int. J. For. Res. 2019, 92, 436–443. [Google Scholar] [CrossRef]
- Dale, V.H.; Joyce, L.A.; McNulty, S.; Neilson, R.P. The Interplay Between Climate Change, Forests, and Disturbances. Sci. Total Environ. 2000, 262, 201–204. [Google Scholar] [CrossRef] [Green Version]
- Dale, V.H.; Joyce, L.A.; McNulty, S.; Neilson, R.P.; Ayres, M.P.; Flannigan, M.D.; Hanson, P.J.; Irland, L.C.; Lugo, A.E.; Peterson, C.J.; et al. Climate Change and Forest Disturbances. BioScience 2001, 51, 723. [Google Scholar] [CrossRef] [Green Version]
- Blennow, K.; Andersson, M.; Sallnäs, O.; Olofsson, E. Climate Change and the Probability of Wind Damage in Two Swedish Forests. For. Ecol. Manag. 2010, 259, 818–830. [Google Scholar] [CrossRef]
- Seidl, R.; Thom, D.; Kautz, M.; Martin-Benito, D.; Peltoniemi, M.; Vacchiano, G.; Wild, J.; Ascoli, D.; Petr, M.; Honkaniemi, J.; et al. Forest Disturbances Under Climate Change. Nat. Clim. Chang. 2017, 7, 395–402. [Google Scholar] [CrossRef] [Green Version]
- Stacey, G.R.; Belcher, R.E.; Wood, C.J.; Gardiner, B.A. Wind Flows and Forces in a Model Spruce Forest. Bound. Layer Meteorol. 1994, 69, 311–334. [Google Scholar] [CrossRef]
- Dupont, S.; Brunet, Y. Impact of Forest Edge Shape on Tree Stability: A Large-Eddy Simulation Study. Forestry 2008, 81, 299–315. [Google Scholar] [CrossRef] [Green Version]
- Irvine, M.R.; Gardiner, B.A.; Hill, M.K. The Evolution of Turbulence Across A Forest Edge. Bound. Layer Meteorol. 1997, 84, 467–496. [Google Scholar] [CrossRef]
- de Langre, E. Effects of Wind on Plants. Annu. Rev. Fluid Mech. 2008, 40, 141–168. [Google Scholar] [CrossRef] [Green Version]
- Marcolla, B.; Pitacco, A.; Cescatti, A. Canopy Architecture and Turbulence Structure in a Coniferous Forest. Bound. Layer Meteorol. 2003, 108, 39–59. [Google Scholar] [CrossRef]
- Cescatti, A.; Marcolla, B. Drag Coefficient and Turbulence Intensity in Conifer Canopies. Agric. For. Meteorol. 2004, 121, 197–206. [Google Scholar] [CrossRef]
- Dupont, S.; Brunet, Y. Edge Flow and Canopy Structure: A Large-Eddy Simulation Study. Bound. Layer Meteorol. 2008, 126, 51–71. [Google Scholar] [CrossRef]
- Queck, R.; Bernhofer, C. Constructing Wind Profiles in Forests from Limited Measurements of Wind and Vegetation Structure. Agric. For. Meteorol. 2010, 150, 724–735. [Google Scholar] [CrossRef]
- Ciftci, C.; Brena, S.F.; Kane, B.; Arwade, S.R. The Effect of Crown Architecture on Dynamic Amplification Factor of an Open-Grown Sugar Maple (Acer saccharum L.). Trees 2013, 27, 1175–1189. [Google Scholar] [CrossRef]
- Ver Planck, N.R.; MacFarlane, D.W. Branch Mass Allocation Increases Wind Throw Risk for Fagus Grandifolia. For. Int. J. For. Res. 2019, 92, 490–499. [Google Scholar] [CrossRef]
- Frederick, R.H. A Study of the Effect of Tree Leaves on Wind Movement. Mon. Wea. Rev. 1961, 89, 39–44. [Google Scholar] [CrossRef]
- Vogel, S. Drag and Reconfiguration of Broad Leaves in High Winds. J. Exp. Bot. 1989, 40, 941–948. [Google Scholar] [CrossRef]
- Dellwik, E.; Bingöl, F.; Mann, J. Flow Distortion at a Dense Forest Edge. Available online: https://rmets.onlinelibrary.wiley.com/doi/abs/10.1002/qj.2155 (accessed on 14 May 2020).
- Angelou, N.; Dellwik, E.; Mann, J. Wind Load Estimation on an Open-Grown European oak Tree. For. Int. J. For. Res. 2019, 92, 381–392. [Google Scholar] [CrossRef]
- Jaffe, M.J. Thigmomorphogenesis: The Response of Plant Growth and Development to Mechanical Stimulation: With Special Reference to Bryonia Dioica. Planta 1973, 114, 143–157. [Google Scholar] [CrossRef]
- Jaffe, M.J.; Forbes, S. Thigmomorphogenesis: The Effect of Mechanical Perturbation on Plants. Plant Growth Regul. 1993, 12, 313–324. [Google Scholar] [CrossRef]
- Coutand, C.; Martin, L.; Leblanc-Fournier, N.; Decourteix, M.; Julien, J.-L.; Moulia, B. Strain Mechanosensing Quantitatively Controls Diameter Growth and PtaZFP2 Gene Expression in Poplar. Plant Physiol. 2009, 151, 223–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonnesoeur, V.; Constant, T.; Moulia, B.; Fournier, M. Forest Trees Filter Chronic Wind-Signals to Acclimate to High Winds. New Phytol. 2016, 210, 850–860. [Google Scholar] [CrossRef] [Green Version]
- Lundström, T.; Jonas, T.; Volkwein, A. Analysing the Mechanical Performance and Growth Adaptation of Norway Spruce Using a Non-Linear Finite-Element Model and Experimental Data. J. Exp. Bot. 2008, 59, 2513–2528. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, M.; de Langre, E.; Moulia, B. A Scaling Law for the Effects of Architecture and Allometry on Tree Vibration Modes Suggests a Biological Tuning to Modal Compartmentalization. Available online: https://bsapubs.onlinelibrary.wiley.com/doi/abs/10.3732/ajb.0800161 (accessed on 13 May 2020).
- Valinger, E. Effects of Wind Sway on Stem Form and Crown Development of Scots Pine (Pinus sylvestris L.). Aust. For. 1992, 55, 15–21. [Google Scholar] [CrossRef]
- Telewski, F.W. Wind-Induced Physiological and Developmental Responses in Trees. In Wind and Trees; Coutts, M.P., Grace, J., Eds.; Cambridge University Press: Cambrige, UK, 1995; pp. 237–263. [Google Scholar]
- Telewski, F.W. Is Windswept Tree Growth Negative Thigmotropism? Plant Sci. 2012, 184, 20–28. [Google Scholar] [CrossRef]
- Telewski, F.W. Flexure Wood: Mechanical Stress Induced Secondary Xylem Formation. In Secondary Xylem Biology; Kim, Y.S., Funada, R., Singh, A.P., Eds.; Academic Press: Boston, MA, USA, 2016; pp. 73–91. ISBN 978-0-12-802185-9. [Google Scholar]
- Nicoll, B.C.; Ray, D. Adaptive Growth of Tree Root Systems in Response to Wind Action and Site Conditions. Tree Physiol. 1996, 16, 891–898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiao-jun, Z.; Zu-gen, L.; Xiu-fen, L.; Matsuzaki, T.; Gonda, Y. Review: Effects of Wind on Trees. J. For. Res. 2004, 15, 153–160. [Google Scholar] [CrossRef]
- Tamasi, E.; Stokes, A.; Lasserre, B.; Danjon, F.; Berthier, S.; Fourcaud, T.; Chiatante, D. Influence of Wind Loading on Root System Development and Architecture in Oak (Quercus robur L.) Seedlings. Trees 2005, 19, 374–384. [Google Scholar] [CrossRef]
- Schindler, D.; Bauhus, J.; Mayer, H. Wind Effects on Trees. Eur. J. For. Res. 2012, 131, 159–163. [Google Scholar] [CrossRef] [Green Version]
- Badel, E.; Ewers, F.W.; Cochard, H.; Telewski, F.W. Acclimation of Mechanical and Hydraulic Functions in Trees: Impact of the Thigmomorphogenetic Process. Front. Plant Sci. 2015, 6. [Google Scholar] [CrossRef] [Green Version]
- Kašpar, J.; Hosek, J.; Treml, V. How Wind Affects Growth in Treeline Picea Abies. Alp. Bot. 2017. [Google Scholar] [CrossRef]
- Tomczak, A.; Jelonek, T.; Pazdrowski, W. Characteristics of Selected Morphological Traits of Trees in Mature Scots Pine Stands Exposed to Wind. Sylwan 2014, 158, 183–191. [Google Scholar]
- Moulia, B.; Der Loughian, C.; Bastien, R.; Martin, O.; Rodríguez, M.; Gourcilleau, D.; Barbacci, A.; Badel, E.; Franchel, G.; Lenne, C.; et al. Integrative Mechanobiology of Growth and Architectural Development in Changing Mechanical Environments. In Mechanical Integration of Plant Cells and Plants; Wojtaszek, P., Ed.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 269–302. ISBN 978-3-642-19091-9. [Google Scholar]
- Moulia, B.; Coutand, C.; Julien, J.-L. Mechanosensitive Control of Plant Growth: Bearing the Load, Sensing, Transducing, and Responding. Front. Plant Sci. 2015, 6. [Google Scholar] [CrossRef]
- Jacobs, M. The Effect of Wind Sway on the Form and Development of Pinus Radiata D. Don. Aust. J. Bot. 1954, 2, 35–51. [Google Scholar] [CrossRef]
- Telewski, F.W.; Jaffe, M.J. Thigmomorphogenesis: Field and laboratory studies of Abies Fraseri in Response to Wind or Mechanical Perturbation. Physiol. Plant. 1986, 66, 211–218. [Google Scholar] [CrossRef]
- Telewski, F.W.; Jaffe, M.J. Thigmomorphogenesis: Anatomical, morphological and mechanical analysis of genetically different sibs of Pinus taeda in response to mechanical perturbation. Physiol. Plant. 1986, 66, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Robertson, A. Centroid of Wood Density, Bole Eccentricity, and Tree-Ring Width in Relation to Vector Winds in Wave Forests. Can. J. For. Res. 1991, 21, 73–82. [Google Scholar] [CrossRef] [Green Version]
- Pruyn, M.L.; Ewers, B.J., III; Telewski, F.W. Thigmomorphogenesis: Changes in the Morphology and Mechanical Properties of Two Populus Hybrids in Response to Mechanical Perturbation. Tree Physiol. 2000, 20, 535–540. [Google Scholar] [CrossRef] [PubMed]
- Alméras, T.; Fournier, M. Biomechanical Design and Long-Term Stability of Trees: Morphological and Wood Traits Involved in the Balance Between Weight Increase and the Gravitropic Reaction. J. Theor. Biol. 2009, 256, 370–381. [Google Scholar] [CrossRef] [Green Version]
- Koch, G.; Bauch, J.; Puls, J.; Schwab, E. Biological, Chemical and Mechanical Characteristics of “Wulstholz” as a Response to Mechanical Stress in Living Trees of Picea abies [L.] Karst. Holzforschung 2000, 54, 137–143. [Google Scholar] [CrossRef]
- Kern, K.A.; Ewers, F.W.; Telewski, F.W.; Koehler, L. Mechanical Perturbation Affects Conductivity, Mechanical Properties and Aboveground Biomass of Hybrid Poplars. Tree Physiol. 2005, 25, 1243–1251. [Google Scholar] [CrossRef]
- Fournier, M.; Alméras, T.; Clair, B.; Gril, J. Biomechanical Action and Biological Functions. In The Biology of Reaction Wood; Springer Series in Wood Science; Gardiner, B., Barnett, J., Saranpää, P., Gril, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 139–169. ISBN 978-3-642-10814-3. [Google Scholar]
- Bruchert, F.; Gardiner, B. The Effect of Wind Exposure on the Tree Aerial Architecture and Biomechanics of Sitka Spruce (Picea sitchensis, Pinaceae). Am. J. Bot. 2006, 93, 1512–1521. [Google Scholar] [CrossRef]
- Brüchert, F. The Influence of the Site Factor Wind Exposure on Wood Quality; Forestry Commission, Northern Research Station: Roslin, Scotland, 2000; p. 119.
- Zipse, A.; Mattheck, C.; Gräbe, D.; Gardiner, B. The Effect of Wind on the Mechanical Properties of the Wood of Beech (Fagus sylvatica L.) Growing in the Borders of Scotland. Arboric. J. 1998, 22, 247–257. [Google Scholar] [CrossRef]
- Bascuñán, A.; Moore, J.R.; Walker, J.C.F. Variations in the Dynamic Modulus of Elasticity with Proximity to the Stand Edge in Radiata Pine Stands on the Canterbury Plains, New Zealand. N. Z. J. For. 2006, 51, 4–8. [Google Scholar]
- Moore, J.; Gardiner, B.; Sellier, D. Tree Mechanics and Wind Loading. In Plant Biomechanics: From Structure to Function at Multiple Scales; Geitmann, A., Gril, J., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 79–106. ISBN 978-3-319-79099-2. [Google Scholar]
- Fournier, M.; Dlouhá, J.; Jaouen, G.; Almeras, T. Integrative Biomechanics for Tree Ecology: Beyond Wood Density and Strength. J. Exp. Bot. 2013, 64, 4793–4815. [Google Scholar] [CrossRef] [Green Version]
- Jelonek, T.; Tomczak, A.; Karaszewski, Z.; Jakubowski, M.; Arasimowicz-Jelonek, M.; Grzywiński, W.; Kopaczyk, J.; Klimek, K. The Biomechanical Formation of Trees. Drewno Prace Naukowe Doniesienia Komunikaty 2019, 62, 5–22. [Google Scholar] [CrossRef]
- Petty, J.A.; Swain, C. Factors Influencing Stem Breakage of Conifers in High Winds. For. Int. J. For. Res. 1985, 58, 75–84. [Google Scholar] [CrossRef]
- Nishimura, T.B. Tree Characteristics Related to Stem Breakage of Picea Glehnii and Abies Sachalinensis. For. Ecol. Manag. 2005, 215, 295–306. [Google Scholar] [CrossRef]
- Peltola, H.; Kellomäki, S.; Hassinen, A.; Granander, M. Mechanical Stability of Scots Pine, Norway Spruce and Birch: An Analysis of Tree-Pulling Experiments in Finland. For. Ecol. Manag. 2000, 135, 143–153. [Google Scholar] [CrossRef]
- Peltola, H.M. Mechanical Stability of Trees Under Static Loads. Am. J. Bot. 2006, 93, 1501–1511. [Google Scholar] [CrossRef]
- Cameron, A.D.; Dunham, R.A. Strength Properties of Wind- and Snow-Damaged Stems of Picea Sitchensis and Pinus Sylvestris in Comparison with Undamaged Trees. Can. J. For. Res. 1999, 29, 595–599. [Google Scholar] [CrossRef]
- Dunham, R.A.; Cameron, A.D. Crown, Stem and Wood Properties of Wind-Damaged and Undamaged Sitka Spruce. For. Ecol. Manag. 2000, 135, 73–81. [Google Scholar] [CrossRef]
- Meyer, F.D.; Paulsen, J.; Körner, C. Windthrow Damage in Picea Abies is Associated with Physical and Chemical Stem Wood Properties. Trees 2008, 22, 463–473. [Google Scholar] [CrossRef]
- Tomczak, A.; Jelonek, T.; Jakubowski, M. Density of Scots pine (Pinus sylvestris L.) Wood as an Indicator of Tree Resistance to Strong Winds. Sylwan 2013, 157, 539–545. [Google Scholar]
- Jakubowski, M.; Jelonek, T.; Tomczak, A. Compressive Strength Parallel to Grain of Scots Pine Wood of Wind−Damaged and Undamaged Trees. Sylwan 2014, 158, 787–794. [Google Scholar]
- Larjavaara, M.; Muller-Landau, H.C. Rethinking the Value of High Wood Density. Funct. Ecol. 2010, 24, 701–705. [Google Scholar] [CrossRef]
- Larjavaara, M.; Muller-Landau, H.C. Still Rethinking the Value of High Wood Density. Am. J. Bot. 2012, 99, 165–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dlouhá, J.; Alméras, T.; Beauchêne, J.; Clair, B.; Fournier, M. Biophysical Dependences among Functional Wood Traits. Funct. Ecol. 2018, 32, 2652–2665. [Google Scholar] [CrossRef] [Green Version]
- Loehle, C. Biomechanical Constraints on Tree Architecture. Trees 2016, 30, 2061–2070. [Google Scholar] [CrossRef]
- ISO 13061-2:2014. Physical and Mechanical Properties of Wood—Test Methods for Small Clear Wood Specimens—Part 2: Determination of Density for Physical and Mechanical Tests; International Organization for Standardization: Geneva, Switzerland, 2014. [Google Scholar]
- ISO 13061-17:2017. Physical and Mechanical Properties of Wood—Test Methods for Small Clear Wood Specimens—Part 17: Determination of Ultimate Stress in Compression Parallel to Grain; International Organization for Standardization: Geneva, Switzerland, 2017. [Google Scholar]
- PN-77/D-04103 Wood. Determination of Static Bending Strength; Polish Committee for Standardization: Warsaw, Poland, 1977. [Google Scholar]
- Kamimura, K.; Gardiner, B.A.; Koga, S. Observations and Predictions of Wind Damage to Larix Kaempferi Trees Following Thinning at an Early Growth Stage. For. Int. J. For. Res. 2017, 90, 530–540. [Google Scholar] [CrossRef]
- Tomczak, A.; Jelonek, T.; Zoń, L. Comparison of Selected Physical Properties of the Juvenile and Mature Wood of Scots Pine (Pinus sylvestris L.) from Mature Stands. Sylwan 2010, 154, 809–817. [Google Scholar]
- Auty, D.; Achim, A.; Macdonald, E.; Cameron, A.D.; Gardiner, B.A. Models for Predicting Wood Density Variation in Scots Pine. Forestry 2014, 87, 449–458. [Google Scholar] [CrossRef] [Green Version]
- Horáček, P.; Fajstavr, M.; Stojanović, M. The Variability of Wood Density and Compression Strength of Norway Spruce (Picea abies/L./Karst.) Within the Stem. Beskydy 2017, 10, 17–26. [Google Scholar] [CrossRef] [Green Version]
- Tomczak, A.; Jelonek, T. Technical Parameters of Juvenile and Mature Wood in Scots Pine (Pinus sylvestris L.). Sylwan 2012, 159, 695–702. [Google Scholar]
- Alméras, T.; Thibaut, A.; Gril, J. Effect of Circumferential Heterogeneity of Wood Maturation Strain, Modulus of Elasticity and Radial Growth on the Regulation of Stem Orientation in Trees. Trees 2005, 19, 457–467. [Google Scholar] [CrossRef]
- Coutand, C.; Pot, G.; Badel, E. Mechanosensing is Involved in the Regulation of Autostress Levels in Tension Wood. Trees 2014, 28, 687–697. [Google Scholar] [CrossRef]
- Pot, G.; Coutand, C.; Toussaint, E.; Le Cam, J.-B.; Saudreau, M. A Model to Simulate the Gravitropic Response and Internal Stresses in Trees, Considering the Progressive Maturation of Wood. Trees 2014, 28, 1235–1248. [Google Scholar] [CrossRef]
- Sellier, D.; Fourcaud, T. Crown Structure and Wood Properties: Influence on Tree Sway and Response to High Winds. Am. J. Bot. 2009, 96, 885–896. [Google Scholar] [CrossRef]
- King, D.A. Tree Form, Height Growth, and Susceptibility to Wind Damage in Acer Saccharum. Ecology 1986, 67, 980–990. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Forest District | Age | dbh | Height | Coordinates of Stands | Direction of the Stand Edge | |
|---|---|---|---|---|---|---|---|
| (cm) | (m) | N | E | ||||
| 1 | Miastko I | 84 | 34 | 26 | 53°58′28″ | 16°58′3″ | N-S |
| 2 | Miastko II | 89 | 36 | 26 | 54°0′56″ | 16°53′59″ | N-S |
| 3 | Warcino | 86 | 31 | 25 | 54°12′37″ | 16°52′28″ | NW-SE |
| 4 | Czaplinek | 87 | 35 | 27 | 53°37′35″ | 16°11′57″ | N-S |
| 5 | Łupawa | 83 | 37 | 24 | 54°23′11″ | 17°17′40″ | N-S |
| 6 | Świdwin | 83 | 32 | 26 | 53° 53′3″ | 15°44′18″ | N-S |
| 7 | Czarnobór | 82 | 31 | 24 | 53°36′59″ | 16°40′37″ | NE-SW |
| Plot | All | |||
|---|---|---|---|---|
| A (0–20 m) | B (30–50 m) | C (60–80 m) | ||
| model trees | 14 | 14 | 14 | 42 |
| logs for wood samples | 70 | 70 | 70 | 210 |
| samples for wood density (WD) | 421 | 403 | 421 | 1245 |
| samples for compressive strength (CS) | 421 | 409 | 434 | 1273 |
| samples for modulus of rupture (MOR) | 420 | 400 | 440 | 1260 |
| samples for modulus of elasticity (MOE) | 419 | 400 | 438 | 1257 |
| A (0–20 m) | B (30–50 m) | C (60–80 m) | |
|---|---|---|---|
| Mean ± SD | Mean ± SD | Mean ± SD | |
| tree height (th) (m) | 23.8 ± 1.2 | 25.9 ± 2.7 | 26.9 ± 3.3 |
| diameter at breast height (dbh) (cm) | 32 ± 6.9 | 34 ± 5.9 | 33 ± 7.5 |
| height-to-diameter-ratio (h:d) | 78 ± 16.5 | 81 ± 17.8 | 85 ± 15.1 |
| radius E d1.3—leeward (cm) | 17.8 ± 4.3 | 16.7 ± 4.5 | 15.7 ± 3.7 |
| radius W d1.3—windward (cm) | 13.2 ± 2.6 | 13.1 ± 2.7 | 12.1 ± 2.9 |
| crown length (cl) (m) | 8.4 ± 2.0 | 7.1 ± 2.2 | 6.3 ± 2.2 |
| crown ratio (cl:th) (crown length/tree height) | 0.35 ± 0.08 C | 0.27 ± 0.07 | 0.23 ± 0.06 A |
| crown diameter (cd) (m) | 4.3 ± 1.3 B | 3.3 ± 1.4 A | 4.0 ± 1.8 |
| A (0–20 m) | B (30–50 m) | C (60–80 m) | |
|---|---|---|---|
| Mean ± SD | Mean ± SD | Mean ± SD | |
| oven-dry density (WD) (kg/m3) | 495 ± 76 | 490 ± 82 | 491 ± 83 |
| compressive strength longitudinal to grain (CS) (MPa) | 23.7 ± 4.5 | 23.2 ± 4.4 | 23.6 ± 4.6 |
| modulus of rupture (MOR) (MPa) | 48.8 ± 10.5 | 48.3 ± 10.0 | 48.5 ± 10.5 |
| modulus of elasticity (MOE) (GPa) | 4.98± 1.23 C | 5.03 ± 1.29 | 5.23± 1.25 A |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tomczak, A.; Jelonek, T.; Pazdrowski, W.; Grzywiński, W.; Mania, P.; Tomczak, K. The Effects of Wind Exposure on Scots Pine Trees: Within-Stem Variability of Wood Density and Mechanical Properties. Forests 2020, 11, 1095. https://doi.org/10.3390/f11101095
Tomczak A, Jelonek T, Pazdrowski W, Grzywiński W, Mania P, Tomczak K. The Effects of Wind Exposure on Scots Pine Trees: Within-Stem Variability of Wood Density and Mechanical Properties. Forests. 2020; 11(10):1095. https://doi.org/10.3390/f11101095
Chicago/Turabian StyleTomczak, Arkadiusz, Tomasz Jelonek, Witold Pazdrowski, Witold Grzywiński, Przemysław Mania, and Karol Tomczak. 2020. "The Effects of Wind Exposure on Scots Pine Trees: Within-Stem Variability of Wood Density and Mechanical Properties" Forests 11, no. 10: 1095. https://doi.org/10.3390/f11101095