Understory Species Identity Rather than Species Richness Influences Fine Root Decomposition in a Temperate Plantation


Abstract
:1. Introduction
- (i)
- A faster fine root decomposition is associated with higher plant species diversity (SR);
- (ii)
- A faster fine root decomposition will be associated with higher SOC and N in the mineral soil;
- (iii)
- Plant species identity is a more important driver of fine root decomposition compared to plant SR per se.
2. Materials and Methods
2.1. Site Description
2.2. Experimental Design
2.3. Vegetation
2.4. Soil Sampling, Fine Root Decomposition and Root Chemistry
2.5. Soil Analyses
2.6. Statistical Analyses
3. Results
3.1. Effects of Silvicultural Treatments on Overstory and Understory Vegetation
3.2. Effects of Silvicultural Treatments on Fine Root Decomposition and Chemistry
3.3. Effects of Subsequent Vegetation on Fine Root Decomposition
3.4. Effects of Silvicultural Treatments on Surface Mineral Soil Properties
3.5. The Relationship between Fine Root Decomposition and Surface Mineral Soil Properties
4. Discussion
4.1. Silvicultural Treatment Effects on Overstory and Understory Vegetation
4.2. Silvicultural Treatment and Fine Root Decomposition Effects on Soil Properties
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Carle, J.; Holmgren, P. Wood from planted forests: A global outlook 2005–2030. For. Prod. J. 2008, 58, 6–18. [Google Scholar]
- Del Lungo, A.; Ball, J.; Carle, J. Global Planted Forests Thematic Study: Results and Analysis; Planted Forests and Trees Working Paper FP/38E; Food and Agricultural Organization: Rome, Italy, 2006. [Google Scholar]
- Stocker, T.F.; Qin, D.; Plattner, G.K.; Tignor, M.M.B.; Allen, S.K.; Boschung, J.; Nauels, A.; Xia, Y.; Bex, V.; Midgley, P.M. Climate Change 2013 the Physical Science Basis: Working Group I Contribution to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013; Volume 1535. [Google Scholar]
- Vesterdal, L.; Clarke, N.; Sigurdsson, B.D.; Gundersen, P. Do tree species influence soil carbon stocks in temperate and boreal forests? For. Ecol. Manag. 2013, 309, 4–18. [Google Scholar] [CrossRef]
- Poirier, V.; Coyea, M.R.; Angers, D.A.; Munson, A.D. Silvicultural treatments and subsequent vegetation impact long-term mineral. For. Ecol. Manag. 2016, 368, 140–150. [Google Scholar] [CrossRef]
- Díaz-Pinés, E.; Rubio, A.; Van Miegroet, H.; Montes, F.; Benito, M. Does tree species composition control soil organic carbon pools in Mediterranean mountain forests? For. Ecol. Manag. 2011, 262, 1895–1904. [Google Scholar] [CrossRef]
- Hansson, K.; Olsson, B.A.; Olsson, M.; Johansson, U.; Kleja, D.B. Differences in soil properties in adjacent stands of Scots pine, Norway spruce and silver birch in SW Sweden. For. Ecol. Manag. 2011, 262, 522–530. [Google Scholar] [CrossRef]
- Finér, L.; Helmisaari, H.-S.; Lõhmus, K.; Majdi, H.; Brunner, I.; Børja, I.; Eldhuset, T.; Godbold, D.; Grebenc, T.; Konôpka, B.; et al. Variation in fine root biomass of three European tree species: Beech (Fagus sylvatica L.), Norway spruce (Picea abies L. Karst.), and Scots pine (Pinus sylvestris L.). Plant Biosyst. Int. J. Deal. Asp. Plant Biol. 2007, 141, 394–405. [Google Scholar] [CrossRef]
- Meinen, C.; Hertel, D.; Leuschner, C. Root growth and recovery in temperate broad-leaved forest stands differing in tree species diversity. Ecosystems 2009, 12, 1103–1116. [Google Scholar] [CrossRef] [Green Version]
- Lei, P.; Scherer-Lorenzen, M.; Bauhus, J. The effect of tree species diversity on fine-root production in a young temperate forest. Oecologia 2012, 169, 1105–1115. [Google Scholar] [CrossRef]
- Rasse, D.P.; Rumpel, C.; Dignac, M.-F. Is soil carbon mostly root carbon? Mechanisms for a specific stabilisation. Plant Soil 2005, 269, 341–356. [Google Scholar] [CrossRef]
- Silver, W.L.; Miya, R.K. Global patterns in root decomposition: Comparisons of climate and litter quality effects. Oecologia 2001, 129, 407–419. [Google Scholar] [CrossRef]
- Chen, H.; Harmon, M.E.; Sexton, J.; Fasth, B. Fine-root decomposition and N dynamics in coniferous forests of the Pacific Northwest, USA. Can. J. For. Res. 2002, 32, 320–331. [Google Scholar] [CrossRef]
- Solly, E.F.; Schöning, I.; Boch, S.; Kandeler, E.; Marhan, S.; Michalzik, B.; Müller, J.; Zscheischler, J.; Trumbore, S.E.; Schrumpf, M. Factors controlling decomposition rates of fine root litter in temperate forests and grasslands. Plant Soil 2014, 382, 203–218. [Google Scholar] [CrossRef] [Green Version]
- Hobbie, S.E.; Oleksyn, J.; Eissenstat, D.M.; Reich, P.B. Fine root decomposition rates do not mirror those of leaf litter among temperate tree species. Oecologia 2010, 162, 505–513. [Google Scholar] [CrossRef]
- Freschet, G.T.; Aerts, R.; Cornelissen, J.H.C. Multiple mechanisms for trait effects on litter decomposition: Moving beyond home-field advantage with a new hypothesis: Substrate-matrix quality interactions in decay. J. Ecol. 2012, 100, 619–630. [Google Scholar] [CrossRef]
- Prieto, I.; Stokes, A.; Roumet, C. Root functional parameters predict fine root decomposability at the community level. J. Ecol. 2016, 104, 725–733. [Google Scholar] [CrossRef]
- Roumet, C.; Birouste, M.; Picon-Cochard, C.; Ghestem, M.; Osman, N.; Vrignon-Brenas, S.; Cao, K.; Stokes, A. Root structure-function relationships in 74 species: Evidence of a root economics spectrum related to carbon economy. New Phytol. 2016, 210, 815–826. [Google Scholar] [CrossRef]
- Hector, A.; Beale, A.J.; Minns, A.; Otway, S.J.; Lawton, J.H. Consequences of the reduction of plant diversity for litter decomposition: Effects through litter quality and microenvironment. Oikos 2000, 90, 357–371. [Google Scholar] [CrossRef]
- Scherer-Lorenzen, M. Functional diversity affects decomposition processes in experimental grasslands. Funct. Ecol. 2008, 22, 547–555. [Google Scholar] [CrossRef]
- Chen, H.; Oram, N.J.; Barry, K.E.; Mommer, L.; van Ruijven, J.; de Kroon, H.; Ebeling, A.; Eisenhauer, N.; Fischer, C.; Gleixner, G.; et al. Root chemistry and soil fauna, but not soil abiotic conditions explain the effects of plant diversity on root decomposition. Oecologia 2017, 185, 499–511. [Google Scholar] [CrossRef] [PubMed]
- Guerrero-Ramírez, N.R.; Craven, D.; Messier, C.; Potvin, C.; Turner, B.L.; Handa, I.T. Root quality and decomposition environment, but not tree species richness, drive root decomposition in tropical forests. Plant Soil 2016, 404, 125–139. [Google Scholar] [CrossRef]
- Prieto, I.; Birouste, M.; Zamora-Ledezma, E.; Gentit, A.; Goldin, J.; Volaire, F.; Roumet, C. Decomposition rates of fine roots from three herbaceous perennial species: Combined effect of root mixture composition and living plant community. Plant Soil 2017, 415, 359–372. [Google Scholar] [CrossRef]
- Munson, A.D.; Margolis, H.A.; Brand, D.G. Intensive silvicultural treatment: Impacts on soil fertility and planted conifer response. Soil Sci. Soc. Am. J. 1993, 57, 246–255. [Google Scholar] [CrossRef]
- Chapman, S.K.; Langley, J.A.; Hart, S.C.; Koch, G.W. Plants actively control nitrogen cycling: Uncorking the microbial bottleneck. New Phytol. 2006, 169, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Environment Canada. Canadian Climate Normal 1971–2000; Petawawa National Forestry Center: Petawawa, ON, Canada, 2016. Available online: http://climat.meteo.gc.ca/climate_normals/results_f.html?stnID=4353&lang=f&StationName=petawawa&SearchType=Contains&stnNameSubmit=go&dCode=0 (accessed on 22 June 2016).
- Soil Classification Working Group. The Canadian System of Soil Classification, 3rd ed.; Publication No. 1646; Agriculture & Agri-Food Canada; NRC Research Press: Ottawa, ON, Canada, 1998. [Google Scholar]
- Science & Education Administration. Soil Survey Staff Soil Taxonomy: A Basic System of Soil Classification for Making and Interpreting Soil Surveys, 2nd ed.; U.S. Government Printing Office: Washington, DC, USA, 1999.
- Bouyoucos, G.J. Hydrometer method improved for making particle size analyses of soils. Agron. J. 1962, 54, 464–465. [Google Scholar] [CrossRef]
- Amacher, M.C.; Henderson, R.E.; Breithaupt, M.D.; Seale, C.L.; La Bauve, J.M. Unbuffered and buffered salt methods for exchangeable cations and effective cation-exchange capacity. Soil Sci. Soc. Am. J. 1990, 54, 1036–1042. [Google Scholar] [CrossRef]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D. Linear and Nonlinear Mixed Effects Models; R Package Version; R Foundation for Statistical Computing: Vienna, Austria, 2007; Volume 3, p. 57. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Henry, M.; Stevens, H.; et al. Vegan: Community Ecology; R Package Version; R Foundation for Statistical Computing: Vienna, Austria, 2016. [Google Scholar]
- Harrell, F.E., Jr.; Dupont, C. With contributions from many others. In Hmisc: Harrell Miscellaneous; R Package Version; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Shan, J.; Morris, L.A.; Hendrick, R.L. The effects of management on soil and plant carbon sequestration in slash pine plantations. J. Appl. Ecol. 2001, 38, 932–941. [Google Scholar] [CrossRef]
- Tilman, D.; Isbell, F.; Cowles, J.M. Biodiversity and Ecosystem Functioning. Annu. Rev. Ecol. Evol. Syst. 2014, 45, 471–493. [Google Scholar] [CrossRef]
- Scherer-Lorenzen, M.; Luis Bonilla, J.; Potvin, C. Tree species richness affects litter production and decomposition rates in a tropical biodiversity experiment. Oikos 2007, 116, 2108–2124. [Google Scholar] [CrossRef]
- De Long, J.R.; Dorrepaal, E.; Kardol, P.; Nilsson, M.-C.; Teuber, L.M.; Wardle, D.A. Understory plant functional groups and litter species identity are stronger drivers of litter decomposition than warming along a boreal forest post-fire successional gradient. Soil Biol. Biochem. 2016, 98, 159–170. [Google Scholar] [CrossRef]
- Handa, I.T.; Aerts, R.; Berendse, F.; Berg, M.P.; Bruder, A.; Butenschoen, O.; Chauvet, E.; Gessner, M.O.; Jabiol, J.; Makkonen, M. Consequences of biodiversity loss for litter decomposition across biomes. Nature 2014, 509, 218–221. [Google Scholar] [CrossRef] [Green Version]
- Finér, L.; Jurgensen, M.; Palviainen, M.; Piirainen, S.; Page-Dumroese, D. Does clear-cut harvesting accelerate initial wood decomposition? A five-year study with standard wood material. For. Ecol. Manag. 2016, 372, 10–18. [Google Scholar] [CrossRef] [Green Version]
- Rousk, J.; Brookes, P.C.; Bååth, E. Contrasting soil pH effects on fungal and bacterial growth suggest functional redundancy in carbon mineralization. Appl. Environ. Microbiol. 2009, 75, 1589–1596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vitousek, P.M.; Matson, P.A. Disturbance, Nitrogen Availability, and Nitrogen Losses in an Intensively Managed Loblolly Pine Plantation. Ecology 1985, 66, 1360–1376. [Google Scholar] [CrossRef]
- Hobbie, S.E.; Ogdahl, M.; Chorover, J.; Chadwick, O.A.; Oleksyn, J.; Zytkowiak, R.; Reich, P.B. Tree species effects on soil organic matter dynamics: The role of soil cation composition. Ecosystems 2007, 10, 999–1018. [Google Scholar] [CrossRef]
- Binkley, D.; Giardina, C. Why do tree species affect soils? The Warp and Woof of tree-soil interactions. In Plant-Induced Soil Changes: Processes and Feedbacks; Van Breemen, N., Ed.; Springer: Dordrecht, The Netherlands, 1998; pp. 89–106. [Google Scholar]
- Côté, L.; Brown, S.; Paré, D.; Fyles, J.; Bauhus, J. Dynamics of carbon and nitrogen mineralization in relation to stand type, stand age and soil texture in the boreal mixedwood. Soil Biol. Biochem. 2000, 32, 1079–1090. [Google Scholar] [CrossRef]
- Augusto, L.; Ranger, J.; Binkley, D.; Rothe, A. Impact of several common tree species of European temperate forests on soil fertility. Ann. For. Sci. 2002, 59, 233–253. [Google Scholar] [CrossRef] [Green Version]
- Mueller, K.E.; Eissenstat, D.M.; Hobbie, S.E.; Oleksyn, J.; Jagodzinski, A.M.; Reich, P.B.; Chadwick, O.A.; Chorover, J. Tree species effects on coupled cycles of carbon, nitrogen, and acidity in mineral soils at a common garden experiment. Biogeochemistry 2012, 111, 601–614. [Google Scholar] [CrossRef]
- Laganiere, J.; Paré, D.; Bergeron, Y.; Chen, H.Y.H.; Brassard, B.W.; Cavard, X. Stability of soil carbon stocks varies with forest composition in the Canadian boreal biome. Ecosystems 2013, 16, 852–865. [Google Scholar] [CrossRef]
- Cotrufo, M.F.; Wallenstein, M.D.; Boot, C.M.; Denef, K.; Paul, E. The Microbial Efficiency-Matrix Stabilization (MEMS) framework integrates plant litter decomposition with soil organic matter stabilization: Do labile plant inputs form stable soil organic matter? Glob. Chang. Biol. 2013, 19, 988–995. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
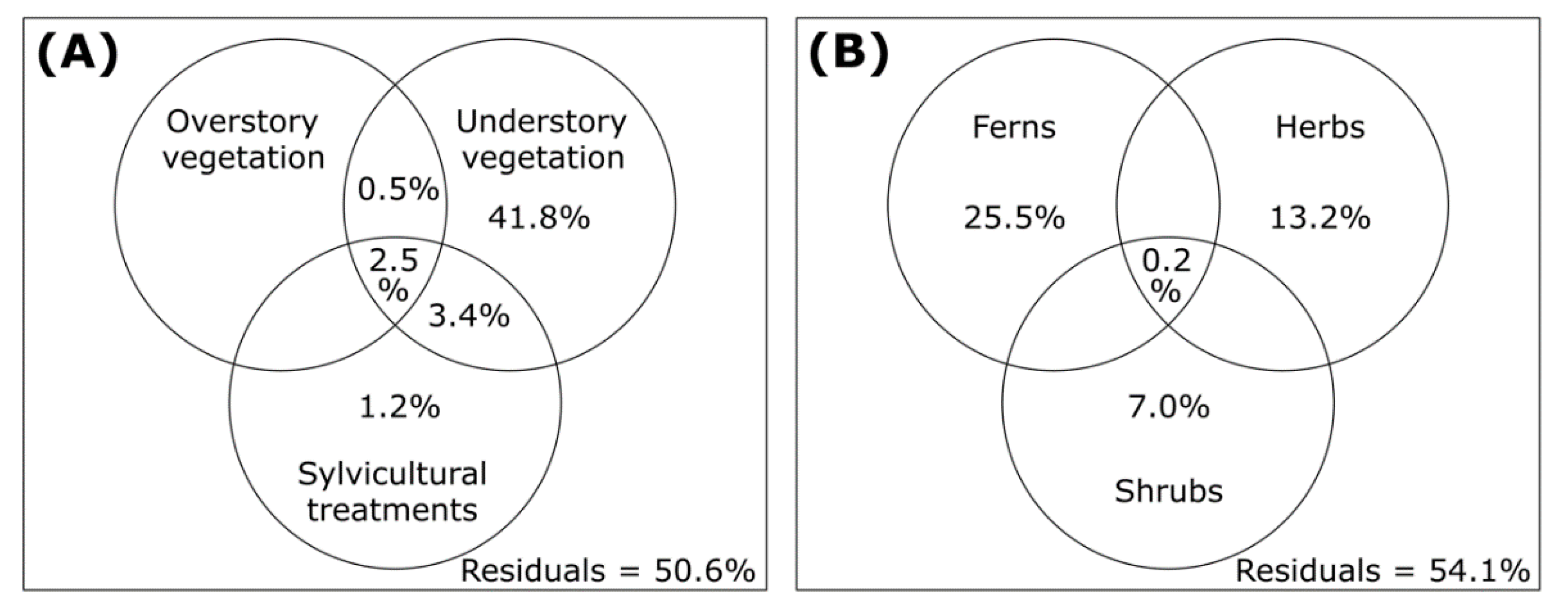{kind=link}
| Species Richness | Basal Area (m2 ha−1) | FRD (%) | C (g kg−1) | N (g kg−1) | Root C:N | P (g kg−1) | K (g kg−1) | Ca (g kg−1) | Mg (g kg−1) | |
|---|---|---|---|---|---|---|---|---|---|---|
| S0F0V0 | 4.2 (±0.3) | 28.0 (±1.9) | 20.6 (±0.01) | 411.7 (±11.8) | 7.4 (±0.5) | 58.6 (±4.2) | 1.27 (±0.15) | 3.35 (±0.35) | 11.1 (±0.96) | 0.93 (±0.12) |
| S1F0V0 | 5.7 (±0.6) | 23.0 (±1.8) | 19.0 (±0.01) | 392.6 (±15.2) | 7.9 (±0.5) | 51.0 (±3.4) | 1.23 (±0.16) | 4.55 (±0.71) | 9.17 (±0.79) | 0.86 (±0.10) |
| S0F1V0 | 3.8 (±0.5) | 31.5 (±2.1) | 18.1 (±0.01) | 414.5 (±13.8) | 6.7 (±0.5) | 65.3 (±4.5) | 1.27 (±0.12) | 4.2 (±0.58) | 12.67 (±0.7) | 0.85 (±0.05) |
| S1F1V0 | 5.6 (±0.3) | 26.0 (±1.5) | 26.1 (±0.01) | 425.6 (±7.8) | 9.0 (±0.7) | 49.7 (±3.5) | 1.6 (±0.19) | 4.55 (±0.58) | 8.75 (±0.83) | 1.05 (±0.20) |
| S0F0V1 | 1.7 (±0.5) | 46.5 (±3.1) | 13.6 (±0.00) | 429.4 (±10.2) | 7.5 (±0.6) | 61.4 (±4.9) | 1.23 (±0.12) | 2.47 (±0.17) | 4.96 (±0.36) | 0.76 (±0.07) |
| S1F0V1 | 2.2 (±0.5) | 41.5 (±1.7) | 18.2 (±0.01) | 436.1 (±8.9) | 7.2 (±0.4) | 62.9 (±3.9) | 1.31 (±0.11) | 2.78 (±0.22) | 4.94 (±0.81) | 0.63 (±0.03) |
| S0F1V1 | 4.0 (±0.8) | 27.0 (±1.1) | 16.0 (±0.01) | 431.4 (±11.0) | 7.4 (±0.4) | 60.2 (±3.5) | 1.27 (±0.09) | 3.31 (±0.2) | 8.28 (±1.01) | 0.83 (±0.06) |
| S1F1V1 | 2.8 (±0.7) | 37.0 (±2.4) | 18.5 (±0.01) | 417.4 (±12.6) | 7.9 (±0.5) | 55.0 (±3.5) | 1.35 (±0.14) | 3.54 (±0.44) | 5.7 (±0.86) | 0.75 (±0.04) |
| V * | V *** | V ** S × F × V ** | V ** | S ** | S ** | NS | V *** F ** | V *** S *** F ** | V ** |
| Correlation with FRD | Adj R2 | Adj R2 Cum | F | p-Values | |
|---|---|---|---|---|---|
| Onoclea sensibilis | (+) | 0.104 | 0.104 | 12.034 | 0.010 |
| Trientalis borealis | (+) | 0.106 | 0.210 | 13.553 | 0.006 |
| Polytrichum sp | (+) | 0.123 | 0.332 | 18.077 | 0.002 |
| Rubus idaeus | (+) | 0.052 | 0.385 | 8.841 | 0.015 |
| Maianthemum canadense | (+) | 0.031 | 0.415 | 5.771 | 0.016 |
| Lycopodium obscurum | (+) | 0.025 | 0.440 | 4.954 | 0.050 |
| Diervilla lonicera | (+) | 0.020 | 0.460 | 4.329 | 0.039 |
| Fern sp | (+) | 0.021 | 0.481 | 4.623 | 0.037 |
| pH | Exch.Ca (mmol (+) kg−1 soil) | Exch.Mg (mmol (+) kg−1 soil) | Exch.K (mmol (+) kg−1 soil) | C Concentration (g kg−1) | N Concentration (g kg−1) | Soil C: N | |
|---|---|---|---|---|---|---|---|
| S0F0V0 | 4.3 (±0.1) | 2.36 (±0.63) | 0.23 (±0.05) | 0.26 (±0.03) | 18.25 (±3.12) | 1.32 (±0.14) | 13.20 (±1.1) |
| S1F0V0 | 4.2 (±0.1) | 1.62 (±0.25) | 0.21 (±0.04) | 0.23 (±0.02) | 15.59 (±1.84) | 1.32 (±0.07) | 11.50 (±1.0) |
| S0F1V0 | 4.6 (±0.1) | 3.88 (±0.97) | 0.34 (±0.08) | 0.31 (±0.03) | 19.23 (±3.83) | 1.35 (±0.16) | 13.30 (±1.1) |
| S1F1V0 | 4.1 (±0.1) | 1.22 (±0.21) | 0.17 (±0.03) | 0.21 (±0.02) | 14.25 (±0.87) | 1.24 (±0.06) | 11.70 (±0.8) |
| S0F0V1 | 4.2 (±0.2) | 2.21 (±0.38) | 0.13 (±0.02) | 0.17 (±0.01) | 16.06 (±1.40) | 1.19 (±0.07) | 13.30 (±0.5) |
| S1F0V1 | 4.1 (±0.1) | 1.07 (±0.22) | 0.13 (±0.02) | 0.18 (±0.01) | 12.99 (±1.22) | 1.20 (±0.05) | 10.70 (±0.7) |
| S0F1V1 | 4.3 (±0.1) | 2.56 (±0.48) | 0.21 (±0.04) | 0.20 (±0.02) | 19.33 (±2.20) | 1.32 (±0.08) | 14.30 (±1.0) |
| S1F1V1 | 4.3 (±0.0) | 1.73 (±0.24) | 0.22 (±0.04) | 0.20 (±0.03) | 14.78 (±1.70) | 1.11 (±0.08) | 13.60 (±1.4) |
| S * | S *** | V ** | V *** | S * | NS | S * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khlifa, R.; Angers, D.A.; Munson, A.D. Understory Species Identity Rather than Species Richness Influences Fine Root Decomposition in a Temperate Plantation. Forests 2020, 11, 1091. https://doi.org/10.3390/f11101091
Khlifa R, Angers DA, Munson AD. Understory Species Identity Rather than Species Richness Influences Fine Root Decomposition in a Temperate Plantation. Forests. 2020; 11(10):1091. https://doi.org/10.3390/f11101091
Chicago/Turabian StyleKhlifa, Rim, Denis A. Angers, and Alison D. Munson. 2020. "Understory Species Identity Rather than Species Richness Influences Fine Root Decomposition in a Temperate Plantation" Forests 11, no. 10: 1091. https://doi.org/10.3390/f11101091