Assessing Habitat Suitability of Parasitic Plant Cistanche deserticola in Northwest China under Future Climate Scenarios


Abstract
:1. Introduction
2. Methods
2.1. Sampling and Area
2.2. Environmental Variables
2.3. RF Model Setting
2.4. Comprehensive Habitat Suitability (CHS) Model
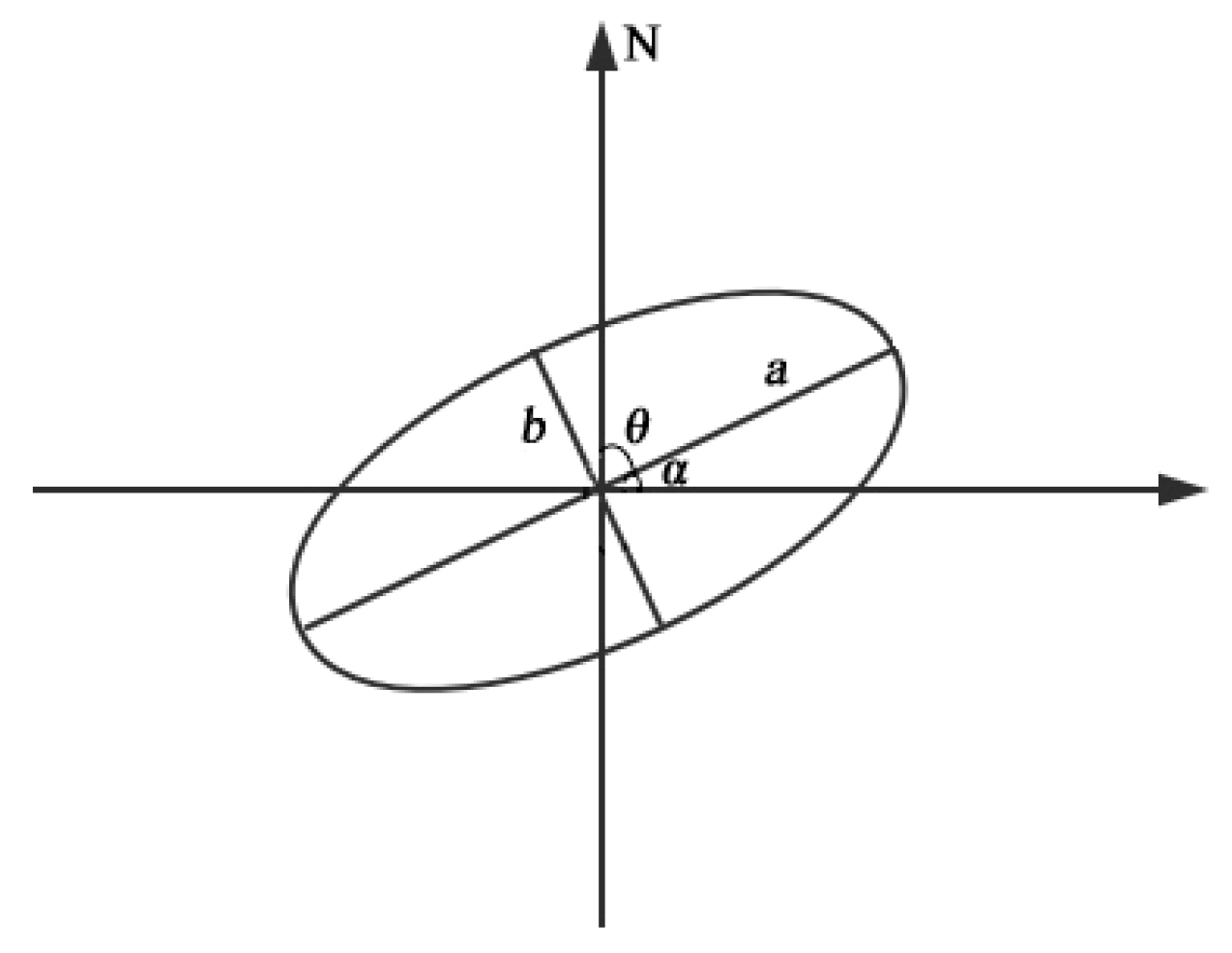
2.5. Calculation of Spatial Pattern
3. Results
3.1. Model Evaluation Analysis
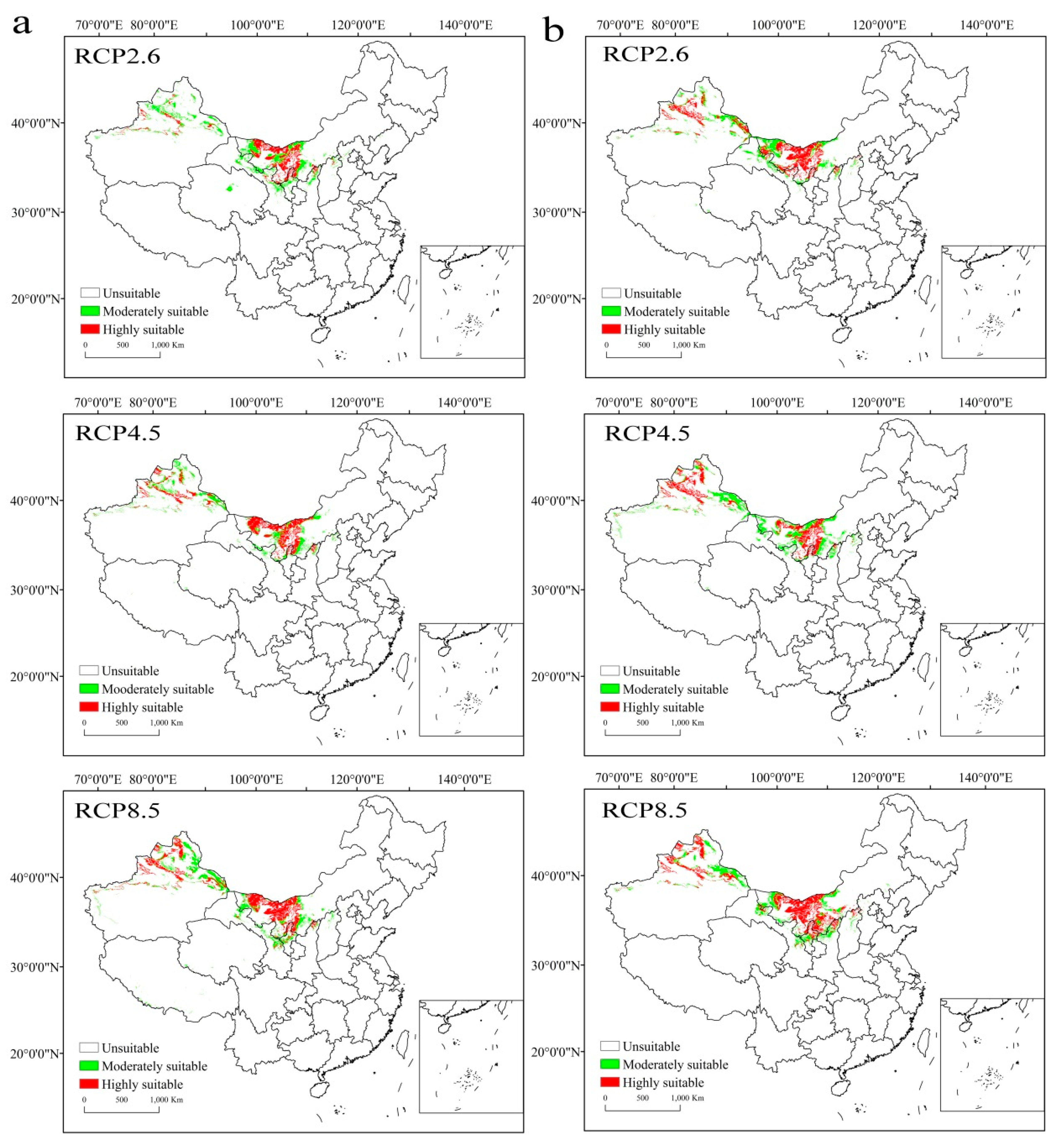
3.2. Potential Geographical Distribution of Suitable Habitats
3.3. Analysis of Factor Affecting the Distribution of C. deserticola
3.4. Identification of the Core Potential Distribution Area
3.5. Directional Distribution Pattern and Migration Trend
4. Discussion
4.1. RF Model
4.2. Evaluation Indexes
4.3. Environment Variable and Scenarios
4.4. Parasitic and Host Plants
4.5. Protection of Desert Forest Ecosystem
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- IPCC Climate Change. The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Stocker, T.F., Qin, D., Plattner, G.K., Tignor, M., Allen, S.K., Boschung, J., Nauels, A., Xia, Y., Bex, V., Midgley, P.M., Eds.; Cambridge University Press: Cambridge, UK, 2013; p. 1535. [Google Scholar]
- Ding, Y.; Ren, G.; Shi, G.; Gong, P.; Zheng, X.; Zhai, P.; Zhang, D.; Zhao, Z.; Wang, S.; Wang, H.; et al. China’s national assessment report on climate change (I): Climate change in china and the future trend. Adv. Clim. Chang. Res. 2007, 3, 1–5. [Google Scholar]
- Guo, Y.; Wei, H.; Lu, C.; Gao, B.; Gu, W. Predictions of potential geographical distribution and quality of Schisandra sphenanthera under climate change. PeerJ 2016, 4, e2554. [Google Scholar] [CrossRef] [PubMed]
- Meynard, C.N.; Gay, P.E.; Lecoq, M.; Foucart, A.; Piou, C.; Chapuis, M.P. Climate-driven geographic distribution of the desert locust during recession periods: Subspecies’ niche differentiation and relative risks under scenarios of climate change. Glob. Chang. Biol. 2017, 23, 4739–4749. [Google Scholar] [CrossRef]
- Shabani, F.; Kumar, L.; Esmaeili, A.; Saremi, H. Climate change will lead to larger areas of Spain being conducive to date palm cultivation. J. Food Agric. Environ. 2013, 11, 2441–2446. [Google Scholar]
- Easterling, D.R.; Meehl, G.A.; Parmesan, C.; Changnon, S.A.; Karl, T.R.; Mearns, L.O. Climate extremes: Observations, modeling, and impacts. Science 2000, 289, 2068–2074. [Google Scholar] [CrossRef] [PubMed]
- Mccarty, J.P. Ecological consequences of recent climate change. Conserv. Biol. 2001, 15, 320–331. [Google Scholar] [CrossRef]
- Chapman, J.W.; Bell, J.R.; Burgin, L.E.; Reynolds, D.R.; Pettersson, L.B.; Hill, J.K.; Bonsallg, M.B.; Thomasg, J.A. Seasonal migration to high latitudes results in major reproductive benefits in an insect. Proc. Natl. Acad. Sci. USA 2012, 109, 14924–14929. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Ruiz, E.P.; Lacher, T.E. Modelling the potential geographic distribution of an endangered pollination corridor in Mexico and the United States. Divers. Distrib. 2017, 23, 67–78. [Google Scholar] [CrossRef]
- Guo, Y.; Li, X.; Zhao, Z.; Wei, H.; Gao, B.; Wei, G. Prediction of the potential geographic distribution of the ectomycorrhizal mushroom Tricholoma matsutake under multiple climate change scenarios. Sci. Rep. 2017, 7, 46221. [Google Scholar] [CrossRef]
- Tang, C.; Dong, Y.; Herrandomoraira, S.; Matsui, T.; Ohashi, H.; He, L.; Nakao, K.; Tanaka, N.; Tomita, M.; Li, X.; et al. Potential effects of climate change on geographic distribution of the Tertiary relict tree species Davidia involucrata in China. Sci. Rep. 2017, 7, 43822. [Google Scholar] [CrossRef]
- Chen, S.; Cunningham, A.A.; Wei, G.; Yang, J.; Liang, Z.; Wang, J.; Wu, M.; Yan, F.; Xiao, H.; Harrison, X.A.; et al. Determining threatened species distributions in the face of limited data: Spatial conservation prioritization for the Chinese giant salamander (Andrias davidianus). Ecol. Evol. 2018, 8, 3098–3108. [Google Scholar] [CrossRef] [PubMed]
- Vencurik, J.; Bosela, M.; Sedmáková, D.; Pittner, J.; Kucbel, S.; Jaloviar, P.; Parobeková, Z.; Saniga, M. Tree species diversity facilitates conservation efforts of european yew. Biodivers. Conserv. 2019, 28, 791–810. [Google Scholar] [CrossRef]
- Shabani, F.; Kumar, L.; Taylor, S. Climate change impacts on the future distribution of date palms: A modeling exercise using climex. PLoS ONE 2012, 7, e48021. [Google Scholar] [CrossRef] [PubMed]
- Reside, A.E.; Vanderwal, J.; Garnett, S.T.; Utt, A.S. Vulnerability of Australian tropical savanna birds to climate change. Austral Ecol. 2016, 41, 106–116. [Google Scholar] [CrossRef]
- Chen, Y.; Vasseur, L.; You, M. Potential distribution of the invasive loblolly pine mealybug, Oracella acuta (Hemiptera: Pseudococcidae), in Asia under future climate change scenarios. Clim. Chang. 2017, 141, 719–732. [Google Scholar] [CrossRef]
- Wu, X.; Dong, S.; Liu, S.; Su, X.; Han, Y.; Shi, J.; Zhang, Y.; Zhao, Z.; Sha, W.; Zhang, X.; et al. Predicting the shift of threatened ungulates’ habitats with climate change in Altun Mountain National Nature Reserve of the Northwestern Qinghai-Tibetan Plateau. Clim. Chang. 2017, 142, 331–344. [Google Scholar] [CrossRef]
- Yang, L.; Zhang, C.; Chen, M.; Li, J.; Yang, L.; Huo, Z.; Ahmad, S.; Luan, X. Long-term ecological data for conservation: Range change in the black-billed capercaillie (Tetrao urogalloides) in northeast China (1970s–2070s). Ecol. Evol. 2018, 8, 3862–3870. [Google Scholar] [CrossRef]
- Koo, K.; Park, S.; Seo, C. Effects of climate change on the climatic niches of warm-adapted evergreen plants: Expansion or contraction? Forests 2017, 8, 500. [Google Scholar] [CrossRef]
- Wang, L.; Wang, W.; Wu, Z.; Du, H.; Zong, S.; Ma, S. Potential Distribution Shifts of Plant Species under Climate Change in Changbai Mountains, China. Forests 2019, 10, 498. [Google Scholar] [CrossRef]
- Keshtkar, H.; Voigt, W. Potential impacts of climate and landscape fragmentation changes on plant distributions: Coupling multi-temporal satellite imagery with GIS-based cellular automata model. Ecol. Inform. 2016, 32, 145–155. [Google Scholar] [CrossRef]
- Thodsen, H.; Baattrup-Pedersen, A.; Andersen, H.E.; Jensen, K.M.; Andersen, P.M.; Bolding, K.; Ovesen, N. Climate change effects on lowland stream flood regimes and riparian rich fen vegetation communities in Denmark. Int. Assoc. Sci. Hydrol. Bull. 2016, 61, 344–358. [Google Scholar] [CrossRef]
- Zhao, Z.; Guo, Y.; Wei, H.; Ran, Q.; Gu, W. Predictions of the potential geographical distribution and quality of a Gynostemma pentaphyllum base on the fuzzy matter element model in China. Sustainability 2017, 9, 1114. [Google Scholar] [CrossRef]
- Zhao, Z. Model construction and comparison of Species Distribution Models under climate change: A Case Study of Notopterygium incisum Ting Ex H. T. Chang. Master’s Thesis, Shaanxi Normal University, Xi’an, China, 2018. [Google Scholar]
- Aguilar-Soto, V.; Melgoza-Castillo, A.; Villarreal-Guerrero, F.; Wehenkel, C.; Pinedo-Alvares, C. Modeling the potential distribution of Picea chihuahuana Martínez, an endangered species at the Sierra Madre Occidental, Mexico. Forests 2015, 6, 692–707. [Google Scholar] [CrossRef]
- Alamgir, M.; Mukul, S.A.; Turton, S.M. Modelling spatial distribution of critically endangered Asian elephant and Hoolock gibbon in Bangladesh forest ecosystems under a changing climate. Appl. Geogr. 2015, 60, 10–19. [Google Scholar] [CrossRef]
- Brambilla, M.; Caprio, E.; Assandri, G.; Scridel, D.; Bassi, E.; Bionda, R.; Celada, C.; Falco, R.; Bogliani, G.; Pedrini, P.; et al. A spatially explicit definition of conservation priorities according to population resistance and resilience, species importance and level of threat in a changing climate. Divers. Distrib. 2017, 23, 1–12. [Google Scholar] [CrossRef]
- Ghaedi, M.; Ghaedi, A.M.; Negintaji, E.; Ansari, A.; Vafaei, A.; Rajabi, M. Random forest model for removal of bromophenol blue using activated carbon obtained from Astragalus bisulcatus tree. J. Ind. Eng. Chem. 2014, 20, 1793–1803. [Google Scholar] [CrossRef]
- Shruthi, R.B.; Kerle, N.; Jetten, V.; Stein, A. Object-based gully system prediction from medium resolution imagery using Random Forests. Geomorphology 2014, 216, 283–294. [Google Scholar] [CrossRef]
- Ma, Y.; Jiang, Q.; Meng, Z.; Li, Z.; Wang, D.; Liu, H. Classification of land use in farming area based on random forest algorithm. Trans. Chin. Soc. Agric. Mach. 2016, 47, 297–303. [Google Scholar]
- Peters, J.; Baets, B.D.; Verhoest, N.E.; Samson, R.; Degroeve, S.; Becker, P.D. Huybrechts W. Random forests as a tool for ecohydrological distribution modelling. Ecol. Model. 2007, 207, 304–318. [Google Scholar] [CrossRef]
- Evans, J.S.; Murphy, M.A.; Holden, Z.A.; Cushman, S.A. Modeling species distribution and change using random forest. In Predictive Species and Habitat Modeling in Landscape Ecology; Drew, C.A., Ed.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 139–159. [Google Scholar]
- Bradter, U.; Kunin, W.E.; Altringham, J.D.; Thom, T.J.; Benton, T.G. Identifying appropriate spatial scales of predictors in species distribution models with the random forest algorithm. Methods Ecol. Evol. 2013, 4, 167–174. [Google Scholar] [CrossRef]
- Gama, M.; Crespo, D.; Dolbeth, M.; Anastácio, P. Predicting global habitat suitability for Corbicula fluminea using species distribution models: The importance of different environmental datasets. Ecol. Model. 2016, 319, 163–169. [Google Scholar] [CrossRef]
- Pliscoff, P.; Luebert, F.; Hilger, H.H.; Guisan, A. Effects of alternative sets of climatic predictors on species distribution models and associated estimates of extinction risk: A test with plants in an arid environment. Ecol. Model. 2014, 288, 166–177. [Google Scholar] [CrossRef]
- Wang, T.; Zhang, X.; Xie, W. Cistanche deserticola Y. C. Ma, “Desert Ginseng”: A review. Am. J. Chin. Med. 2012, 40, 1123–1141. [Google Scholar] [CrossRef] [PubMed]
- Cai, R.L.; Yang, M.H.; Shi, Y.; Chen, J.; Li, Y.C.; Qi, Y. Antifatigue activity of phenylethanoid-rich extract from Cistanche deserticola. Phytother. Res. 2010, 24, 313–315. [Google Scholar] [CrossRef]
- Jin, G.C.; Moon, M.; Jeong, H.U.; Min, C.K.; Sun, Y.K.; Oh, M.S. Cistanches Herba enhances learning and memory by inducing nerve growth factor. Behav. Brain Res. 2011, 216, 652–658. [Google Scholar]
- Shamsutdinov, Z.S.; Ubaidullaev, S.R. Distribution of Poa bulbosa L. and Carex pachystylis Gay within the phytogenous field of black saxaul. Probl. Desert Dev. 1988, 1, 38–43. [Google Scholar]
- Tobe, K.; Li, X.; Omasa, K. Effects of sodium chloride on seed germination and growth of two Chinese desert shrubs, Haloxylon ammodendron and H. persicum (Chenopodiaceae). Aust. J. Bot. 2000, 48, 455–460. [Google Scholar] [CrossRef]
- Xu, G.Q.; Li, Y.; Zou, T. Hydraulic resistance partitioning between shoot and root system and plant water status of Haloxyolon ammodendron growing at sites of contrasting soil texture. J. Arid Land 2010, 2, 98–106. [Google Scholar] [CrossRef]
- Naran, R.; Ebringerova, A.; HromàdkovÁ, Z.; PatoprstÝ, V. Carbohydrate polymers from underground parts of Cistanche deserticola. Phytochemistry 1995, 40, 709–715. [Google Scholar] [CrossRef]
- Zheng, G.; Song, Y.; Guo, S.; Ma, H.; Niu, D. Soluble sugar accumulation and the activities of sugar metabolism related enzymes in Cistanche deserticola and its host Haloxylon ammodendron. Acta Bot. Boreal Occident. Sin. 2006, 26, 1175–1182. [Google Scholar]
- Tan, D.; Guo, Q.; Liu, Y.; Ma, C.; Wang, X. The physiological metabolism reaction of Haloxylon parasitized by Cistanche deserticola. For. Res. 2007, 20, 495–499. [Google Scholar]
- Tu, P.; Jiang, Y.; Guo, Y. Review on the research progress and industry development of cistanches herba. J. Chin. Pharm. Sci. 2011, 46, 882–887. [Google Scholar]
- Thuiller, W.; Lafourcade, B.; Engler, R.; Araujo, M.B. BIOMOD: A platform for ensemble forecasting of species distributions. Ecography 2009, 32, 369–373. [Google Scholar] [CrossRef]
- Booth, T.H.; Nix, H.A.; Busby, J.R.; Hutchinson, M.F. BIOCLIM: The first species distribution modelling package, its early applications and relevance to most current MAXENT studies. Divers. Distrib. 2014, 20, 1–9. [Google Scholar] [CrossRef]
- Breiman, L. Bagging predictors. Mach. Learn. 1996, 24, 123–140. [Google Scholar] [CrossRef] [Green Version]
- Guisan, A.; Thuiller, W.; Zimmermann, N.E. Habitat Suitability and Distribution Models with Applications in R; Cambridge University Press: Cambridge, UK, 2017; p. 203. [Google Scholar]
- Friedman, J.H. Greedy function approximation: A gradient boosting machine. Ann. Stat. 2001, 29, 1189–1232. [Google Scholar] [CrossRef]
- Wang, Y.; Xie, B.; Wan, F.; Xiao, Q.; Dai, L. Application of ROC curve analysis in evaluating the performance of alien species’ potential distribution models. Biodivers. Sci. 2007, 15, 365–372. [Google Scholar]
- Kwon, Y.S.; Bae, M.J.; Hwang, S.J.; Kim, S.H.; Park, Y.S. Predicting potential impacts of climate change on freshwater fish in Korea. Ecol. Inform. 2015, 29, 156–165. [Google Scholar] [CrossRef]
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the accuracy of species distribution models: Prevalence, kappa and the true skill statistic (TSS). J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar] [CrossRef]
- Guo, Y.; Li, X.; Zhao, Z.; Nawaz, Z. Predicting the impacts of climate change, soils and vegetation types on the geographic distribution of Polyporus umbellatus in China. Sci. Total Environ. 2019, 948, 1–11. [Google Scholar] [CrossRef]
- Lefever, D.W. Measuring geographic concentration by means of the standard deviational ellipse. Am. J. Sociol. 1926, 32, 88–94. [Google Scholar] [CrossRef]
- Gong, J.X. Clarifying the standard deviational ellipse. Geogr. Anal. 2002, 34, 155–167. [Google Scholar] [CrossRef]
- Rahmaniati, M.; Eryando, T.; Susanna, D.; Pratiwi, D.; Nugraha, F.; Ruliansah, A.; Riandi, M.U. The utilization of standard deviational ellipse (SDE) model for the analysis of dengue fever cases in Banjar city 2013. Aspirator 2014, 6, 21–28. [Google Scholar] [CrossRef]
- Tang, J.; Ma, M.; Liu, L. Study on temporal and spatial evolution of structure of lodging industry and Influencing Factors in Red Tourism City—A case on Xiangtan City. J. Hunan Univ. Financ. Econ. 2018, 34, 72–80. [Google Scholar]
- Mitchell, A. The ESRI Guide to GIS Analysis, Volume 2: Spatial Measurements and Statistics; Esri Guide to GIS Analysis; ESRI Press: Redlands, CA, USA, 2005. [Google Scholar]
- Fischer, M.M.; Getis, A. Handbook of Applied Spatial Analysis: Software Tools, Methods and Applications; Springer: Berlin/Heidelberg, Germany, 2011. [Google Scholar]
- Yang, H.; Yan, B.; Qi, C.; Chen, D. Evaluation and analysis of wind resources in Jin-Jing-Ji region of China. Procedia 2011, 11, 836–842. [Google Scholar]
- Pishgar-Komleh, S.H.; Keyhani, A.; Sefeedpari, P. Wind speed and power density analysis based on Weibull and Rayleigh distributions (a case study: Firouzkooh county of Iran). Renew. Sustain Energy Rev. 2015, 42, 313–322. [Google Scholar] [CrossRef]
- Chen, J.; Xie, C.; Chen, S.; Sun, C.; Zhao, R.; Xu, R. Suitability evaluation of Cistanche desertiola based on TCMGIS-I. China J. Chin. Mater. Med. 2007, 32, 1396–1401. [Google Scholar]
- Huang, X.; Xu, R.; Chen, J.; Yu, J.; Liu, S.; Liu, T. Status and prospect of studies on habitat characteristics, parasitic mechanism and nutrient transport of Cistanche deserticola. China J. Chin. Mater. Med. 2012, 37, 2831–2835. [Google Scholar]
- Ma, S.; Wei, B.; Li, X.; Luo, C.; Sun, F. The impacts of climate change on the potential distribution of Haloxylon ammodendron. Chin. J. Ecol. 2017, 36, 1243–1250. [Google Scholar]
- Cheng, A.; Feng, Q.; Zhang, J.; Li, Z.; Wang, G. A Review of Climate Change Scenario for Impacts Process Study. Sci. Geogr. Sin. 2015, 35, 84–90. [Google Scholar]
- Mullan, D.; Swindles, G.; Patterson, T.; Galloway, J.; Macumber, A.; Falck, H.; Crossley, L.; Chen, J.; Pisaric, M. Climate change and the long-term viability of the World’s busiest heavy haul ice road. Theor. Appl. Climatol. 2016, 129, 1089–1108. [Google Scholar] [CrossRef]
- Elith, J.; Kearney, M.; Phillips, S. The art of modelling range-shifting species. Methods Ecol. Evol. 2010, 1, 330–342. [Google Scholar] [CrossRef]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Breiman, L. Random forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Breiner, F.T.; Guisan, A.; Bergamini, A.; Nobis, M.P. Overcoming limitations of modelling rare species by using ensembles of small models. Methods Ecol. Evol. 2015, 6, 1210–1218. [Google Scholar] [CrossRef]
- Briscoe, N.J.; Elith, J.; Salguero-Gómez, R.; Lahoz-Monfort, J.J.; Camac, J.S.; Giljohann, K.M.; Holden, M.H.; Hradsky, B.A.; Kearney, M.R.; McMahon, S.M.; et al. Forecasting species range dynamics with process—Explicit models: Matching methods to applications. Ecol. Lett. 2019, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.; Chiu, C. The Relic Trochodendron aralioides Siebold & Zucc. (Trochodendraceae) in Taiwan: Ensemble distribution modeling and climate change impacts. Forests 2019, 10, 7. [Google Scholar]
- Lomba, A.; Pellissier, L.; Randin, C.F.; Vicente, J.; Moreira, F.; Honrado, J.; Guisan, A. Overcoming the rare species modelling paradox: A novel hierarchical framework applied to an Iberian endemic plant. Biol. Conserv. 2010, 143, 2647–2657. [Google Scholar] [CrossRef]
- VanDerWal, J.; Shoo, L.P.; Graham, C.; Williams, S.E. Selecting pseudo-absence data for presence-only distribution modeling: How far should you stray from what you know? Ecol. Model. 2009, 220, 589–594. [Google Scholar] [CrossRef]
- McPherson, J.; Jetz, W.; Rogers, D. The effects of species’ range sizes on the accuracy of distribution models: Ecological phenomenon or statistical artefact? J. Appl. Ecol. 2004, 41, 811–823. [Google Scholar] [CrossRef]
- Engler, R.; Guisan, A. MigClim: Predicting plant distribution and dispersal in a changing climate. Divers. Distrib. 2009, 15, 590–601. [Google Scholar] [CrossRef]
- Lobo, J.M.; Jimenez-Valverde, A.; Real, R. AUC: A misleading measure of the performance of predictive distribution models. Glob. Ecol. Biogeogr. 2008, 17, 145–151. [Google Scholar] [CrossRef]
- Swets, J.A. Measuring the accuracy of diagnostic systems. Science 1988, 240, 1285–1293. [Google Scholar] [CrossRef]
- Shabani, F.; Kumar, L.; Ahmadi, M. A comparison of absolute performance of different correlative and mechanistic species distribution models in an independent area. Ecol. Evol. 2016, 6, 5973–5986. [Google Scholar] [CrossRef] [Green Version]
- Pearson, G.A.; Mota, L.L. Frayed at the edges: Selective pressure and adaptive response to abiotic stressors are mismatched in low diversity edge populations. J. Ecol. 2009, 97, 450–462. [Google Scholar] [CrossRef]
- Crowther, M.S.; Lunney, D.; Lemon, J.; Stalenberg, E.; Ellis, M. Climate-mediated habitat selection in an arboreal folivore. Ecography 2013, 36, 1–8. [Google Scholar] [CrossRef]
- Lee, D.S.; Bae, Y.S.; Byun, B.K.; Lee, S.; Park, J.K.; Park, Y.S. Occurrence prediction of the citrus flatid planthopper (Metcalfa pruinosa (Say, 1830)) in South Korea using a random forest model. Forests 2019, 10, 583. [Google Scholar] [CrossRef]
- Shabani, F.; Ahmadi, M.; Peters, K.J.; Haberle, S.; Champreux, A.; Saltréand, F.; Bradshaw, C.J.A. Climate-driven shifts in the distribution of koala-browse species from the Last Interglacial to the near future. Ecography 2019, 42, 1–13. [Google Scholar] [CrossRef]
- Booth, T.H. Impacts of climate change on eucalypt distributions in Australia: An examination of a recent study. Aust. For. 2017, 80, 208–215. [Google Scholar] [CrossRef]
- Xu, X.; Zhang, H.; Xie, T.; Xu, Y.; Zhao, L.; Tian, W. Effects of Climate Change on the Potentially Suitable Climatic Geographical Range of Liriodendron chinense. Forests 2017, 8, 399. [Google Scholar] [CrossRef]
- Adhikari, P.; Shin, M.S.; Jeon, J.Y.; Kim, H.W.; Hong, S.; Seo, C. Potential impact of climate change on the species richness of subalpine plant species in the mountain national parks of south korea. J. Ecol. Environ. 2018, 42, 36. [Google Scholar] [CrossRef]
- Tan, D.; Guo, Q.; Wang, C.; Ma, C. Effects of the parasite plant (Cistanche deserticola) on growth and biomass of the host plant (Haloxylon ammodendron). For. Res. 2004, 17, 472–478. [Google Scholar]
- Li, L.I.; Xu, X.; Sun, Y.; Han, W. Effects of parasitic plant Cistanche deserticola on chlorophyll a fluorescence and nutrient accumulation of host plant Haloxylon ammodendron in the Taklimakan Desert. J. Arid Land 2012, 4, 342–348. [Google Scholar] [CrossRef]
- Zou, X.; Wang, S. Strengthening the protection of desert forest resources—Haloxylon ammodendron. J. Inn. Mong. For. 2004, 6, 19. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Environmental Variables | Variable | Label |
|---|---|---|
| Bioclimatic Variables | Annual Mean Temperature | Bio1 |
| Mean Diurnal Range (Mean of monthly (max temp − min temp)) | Bio2 | |
| Min Temperature of Coldest Month | Bio6 | |
| Temperature Annual Range (Bio5-Bio6) | Bio7 | |
| Mean Temperature of Driest Quarter | Bio9 | |
| Mean Temperature of Warmest Quarter | Bio10 | |
| Annual Precipitation | Bio12 | |
| Precipitation of Wettest Month | Bio13 | |
| Precipitation of Wettest Quarter | Bio16 | |
| Precipitation of Warmest Quarter | Bio18 | |
| Soil Variables | Topsoil USDA texture classification | T_USDA |
| Topsoil pH (H2O) | T_PH | |
| Topsoil Organic Carbon (% weight) | T_OC | |
| Topsoil Calcium Carbonate (% weight) | T_CACO3 | |
| Topsoil Gypsum (% weight) | T_CASO4 | |
| Subsoil pH (H2O) | S_PH | |
| Subsoil Organic Carbon (% weight) | S-OC | |
| Soil Type Variables | ST |
| Evaluation | Evaluation Grade | ||||
|---|---|---|---|---|---|
| Indexes | Best | Better | Moderate | General | Failed |
| AUC | 1.0–0.91 | 0.90–0.81 | 0.80–0.71 | 0.70–0.61 | <0.60 |
| KAPPA | 1.0–0.81 | 0.80–0.61 | 0.60–0.41 | 0.40–0.21 | <0.20 |
| TSS | 1.0–0.86 | 0.85–0.71 | 0.70–0.56 | 0.55–0.41 | <0.40 |
| Time | RCPs | Research | Mean Center (°) | SDE x(km) | SDE y(km) | Rotation θ (°) | Ellipticity | |
|---|---|---|---|---|---|---|---|---|
| Period | Area | |||||||
| 2050s | RCP2.6 | I11 | 84.193 | 42.927 | 528.040 | 185.032 | 78.821 | 0.937 |
| II11 | 105.908 | 39.740 | 483.449 | 212.168 | 91.725 | 0.899 | ||
| RCP4.5 | I21 | 85.771 | 43.874 | 662.348 | 222.273 | 80.391 | 0.942 | |
| II21 | 105.802 | 40.538 | 1707.532 | 430.200 | 96.585 | 0.968 | ||
| RCP8.5 | I31 | 86.332 | 43.613 | 730.405 | 246.197 | 81.775 | 0.941 | |
| II31 | 105.076 | 39.425 | 371.492 | 242.913 | 99.197 | 0.757 | ||
| 2070s | RCP2.6 | I12 | 87.005 | 43.929 | 726.342 | 207.777 | 85.109 | 0.958 |
| II12 | 105.032 | 40.311 | 565.755 | 144.648 | 95.894 | 0.967 | ||
| RCP4.5 | I22 | 85.474 | 43.911 | 613.714 | 262.113 | 79.400 | 0.904 | |
| II22 | 106.471 | 40.401 | 432.174 | 180.055 | 90.526 | 0.909 | ||
| RCP8.5 | I32 | 86.503 | 43.916 | 634.586 | 242.002 | 82.252 | 0.924 | |
| II32 | 106.845 | 39.712 | 522.806 | 225.796 | 92.148 | 0.902 | ||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, J.; Yang, Y.; Wei, H.; Zhang, Q.; Zhang, X.; Zhang, X.; Gu, W. Assessing Habitat Suitability of Parasitic Plant Cistanche deserticola in Northwest China under Future Climate Scenarios. Forests 2019, 10, 823. https://doi.org/10.3390/f10090823
Liu J, Yang Y, Wei H, Zhang Q, Zhang X, Zhang X, Gu W. Assessing Habitat Suitability of Parasitic Plant Cistanche deserticola in Northwest China under Future Climate Scenarios. Forests. 2019; 10(9):823. https://doi.org/10.3390/f10090823
Chicago/Turabian StyleLiu, Jing, Yang Yang, Haiyan Wei, Quanzhong Zhang, Xuhui Zhang, Xiaoyan Zhang, and Wei Gu. 2019. "Assessing Habitat Suitability of Parasitic Plant Cistanche deserticola in Northwest China under Future Climate Scenarios" Forests 10, no. 9: 823. https://doi.org/10.3390/f10090823