Role of Plant Traits in Photosynthesis and Thermal Damage Avoidance under Warmer and Drier Climates in Boreal Forests


Abstract
:1. Introduction
2. Materials and Methods
2.1. APES-Atmosphere-Plant Exchange Simulator Model
2.2. Metrics Evaluating Thermal Risk and Assimilation Capacity
2.2.1. Maximum TL (TL,max)
2.2.2. Cumulated Anet (Anet,cum)
2.2.3. Maximum Anet (Anet,max)
2.2.4. Thermal Range of High Photosynthesis (∆T90)
2.3. Plant Traits Selection and Determination of Their Role
2.3.1. Selection of Plant Traits
2.3.2. General Sensitivity Analysis (GSA) and Determination of Dominant Traits
2.3.3. Selection of Focal Trait Combinations
2.4. Growing Conditions
2.4.1. Current Growing Conditions
2.4.2. Scenarios of Future Growing Conditions
3. Results
3.1. Dominant Traits Emerging from the General Sensitivity Analysis (GSA)
3.1.1. Maximum TL (TL,max)
3.1.2. Cumulated Anet (Anet,cum) and Maximum Anet (Anet,max)
3.1.3. Thermal Range of High Photosynthesis (∆T90)
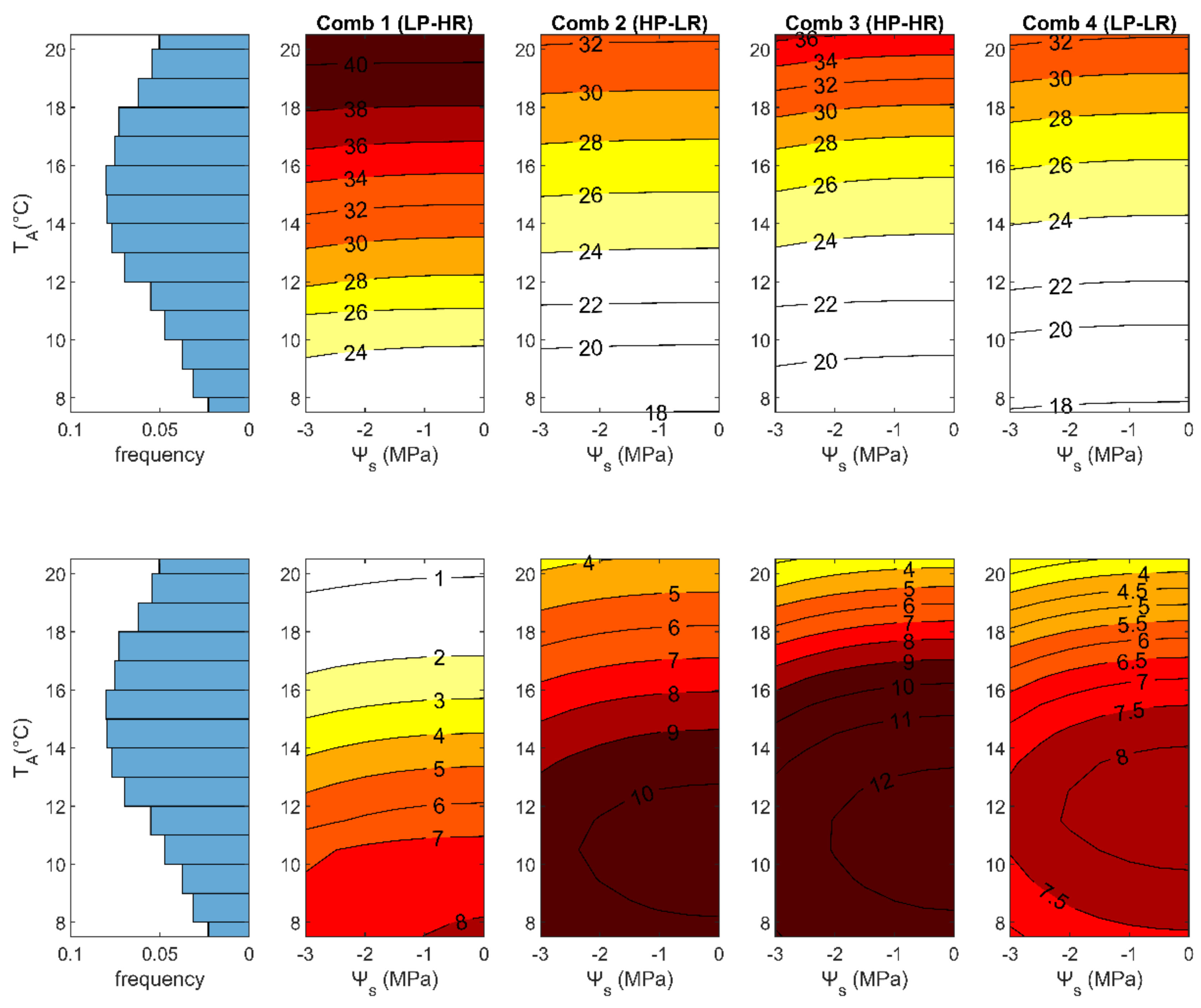
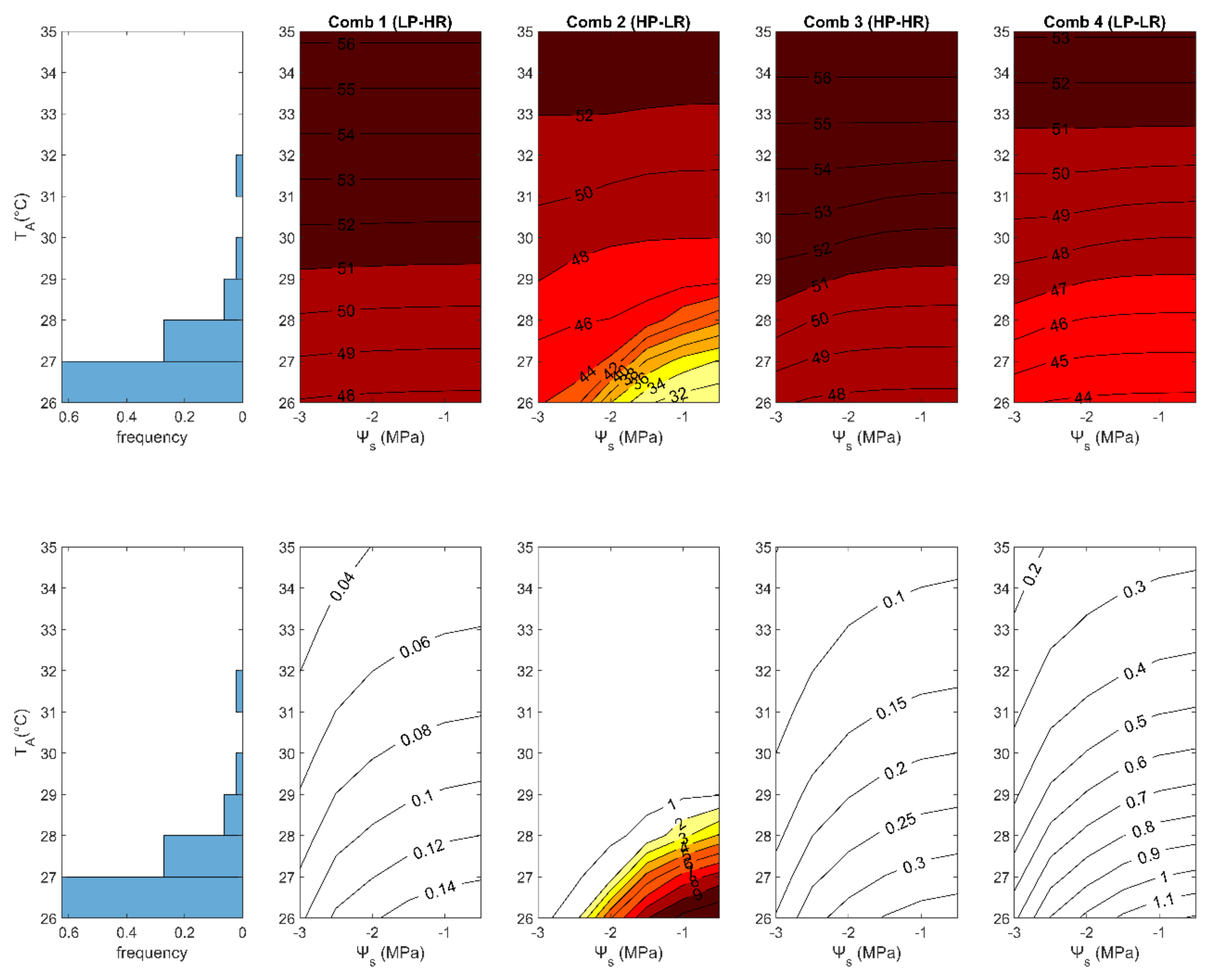
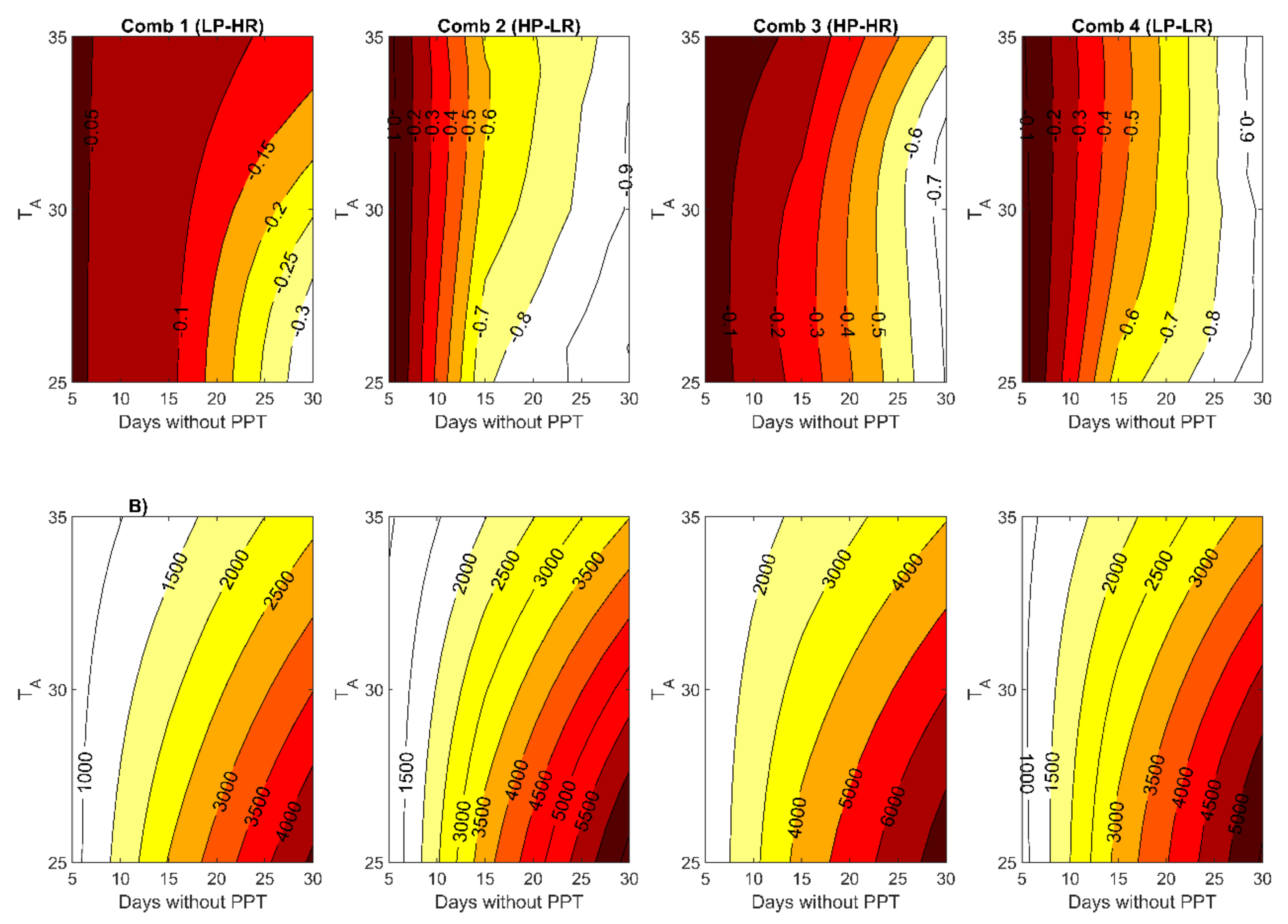
3.2. Role of the Growing Conditions
3.2.1. Differences between 2005 and 2006
3.2.2. Future Growing Conditions
4. Discussion
4.1. Role of Traits: Potential Mechanisms Explaining the Dominance of Plant Traits
4.2. Role of Growing Conditions: the Timing of Precipitatio Affects the Risk of Thermal Damage
4.3. Interactions of Traits and Growing Conditions: Most Suitable Traits for Enhanced CO2 Assimilation and Reduced Risk of Thermal Damage Under Current and Future Climates
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Yamori, W.; Hikosaka, K.; Way, D.A. Temperature response of photosynthesis in C-3, C-4, and CAM plants: Temperature acclimation and temperature adaptation. Photosynth. Res. 2014, 119, 101–117. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Alam, M.M.; Roychowdhury, R.; Fujita, M. Physiological, Biochemical, and Molecular Mechanisms of Heat Stress Tolerance in Plants. Int. J. Mol. Sci. 2013, 14, 9643–9684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Sullivan, O.S.; Heskel, M.A.; Reich, P.B.; Tjoelker, M.G.; Weerasinghe, L.K.; Penillard, A.; Zhu, L.L.; Egerton, J.J.G.; Bloomfield, K.J.; Creek, D.; et al. Thermal limits of leaf metabolism across biomes. Glob. Chang. Biol. 2017, 23, 209–223. [Google Scholar] [CrossRef] [PubMed]
- Schreiber, U.; Berry, J.A. Heat-induced changes of chlorophyll fluorescence in intact leaves correlated with damage of the photosynthetic apparatus. Planta 1977, 136, 233–238. [Google Scholar] [CrossRef]
- Huve, K.; Bichele, I.; Rasulov, B.; Niinemets, U. When it is too hot for photosynthesis: Heat-induced instability of photosynthesis in relation to respiratory burst, cell permeability changes and H2O2 formation. Plant Cell Environ. 2011, 34, 113–126. [Google Scholar] [CrossRef] [PubMed]
- Knight, C.A.; Ackerly, D.D. Evolution and plasticity of photosynthetic thermal tolerance, specific leaf area and leaf size: Congeneric species from desert and coastal environments. New Phytol. 2003, 160, 337–347. [Google Scholar] [CrossRef]
- O’Sullivan, O.S.; Weerasinghe, K.W.L.K.; Evans, J.R.; Egerton, J.J.G.; Tjoelker, M.G.; Atkin, O.K. High-resolution temperature responses of leaf respiration in snow gum (Eucalyptus pauciflora) reveal high-temperature limits to respiratory function. Plant, Cell 2013, 36, 1268–1284. [Google Scholar] [CrossRef]
- Chaves, M.; Costa, J.; Zarrouk, O.; Pinheiro, C.; Lopes, C.; Pereira, J.; Costa, J. Controlling stomatal aperture in semi-arid regions—The dilemma of saving water or being cool? Plant Sci. 2016, 251, 54–64. [Google Scholar] [CrossRef]
- Monteiro, M.V.; Blanuša, T.; Verhoef, A.; Hadley, P.; Cameron, R.W.F. Relative importance of transpiration rate and leaf morphological traits for the regulation of leaf temperature. Aust. J. Bot. 2016, 64, 32. [Google Scholar] [CrossRef] [Green Version]
- Radin, J.W.; Lu, Z.; Percy, R.G.; Zeiger, E. Genetic variability for stomatal conductance in Pima cotton and its relation to improvements of heat adaptation. Proc. Natl. Acad. Sci. USA 1994, 91, 7217–7221. [Google Scholar] [CrossRef]
- Okajima, Y.; Taneda, H.; Noguchi, K.; Terashima, I. Optimum leaf size predicted by a novel leaf energy balance model incorporating dependencies of photosynthesis on light and temperature. Ecol. Res. 2012, 27, 333–346. [Google Scholar] [CrossRef]
- Vogel, S. Leaves in the lowest and highest winds: temperature, force and shape. New Phytol. 2009, 183, 13–26. [Google Scholar] [CrossRef] [Green Version]
- Leigh, A.; Sevanto, S.; Ball, M.C.; Close, J.D.; Ellsworth, D.S.; Knight, C.A.; Nicotra, A.B.; Vogel, S. Do thick leaves avoid thermal damage in critically low wind speeds? New Phytol. 2012, 194, 477–487. [Google Scholar] [CrossRef] [Green Version]
- Luquet, D.; Bégué, A.; Vidal, A.; Clouvel, P.; Dauzat, J.; Olioso, A.; Gu, X.; Tao, Y. Using multidirectional thermography to characterize water status of cotton. Remote. Sens. Environ. 2003, 84, 411–421. [Google Scholar] [CrossRef]
- Intergovernmental Panel on Climate Change. Climate Change 2014—Impacts, Adaptation and Vulnerability: Part B: Regional Aspects; Cambridge University Press: Cambridge, UK, 2014. [Google Scholar]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E. (Ted); et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef]
- Boisvenue, C.; Running, S.W. Impacts of climate change on natural forest productivity—Evidence since the middle of the 20th century. Chang. Boil. 2006, 12, 862–882. [Google Scholar] [CrossRef]
- Sastry, A.; Barua, D. Leaf thermotolerance in tropical trees from a seasonally dry climate varies along the slow-fast resource acquisition spectrum. Sci. Rep. 2017, 7, 11246. [Google Scholar] [CrossRef] [Green Version]
- Bonan, G.B. Forests and Climate Change: Forcings, Feedbacks, and the Climate Benefits of Forests. Science 2008, 320, 1444–1449. [Google Scholar] [CrossRef] [Green Version]
- Pan, Y.; Birdsey, R.A.; Fang, J.; Houghton, R.; Kauppi, P.E.; Kurz, W.A.; Phillips, O.L.; Shvidenko, A.; Lewis, S.L.; Canadell, J.G.; et al. A Large and Persistent Carbon Sink in the World’s Forests. Science 2011, 333, 988–993. [Google Scholar] [CrossRef]
- Baul, T.K.; Alam, A.; Ikonen, A.; Strandman, H.; Asikainen, A.; Peltola, H.; Kilpeläinen, A. Climate Change Mitigation Potential in Boreal Forests: Impacts of Management, Harvest Intensity and Use of Forest Biomass to Substitute Fossil Resources. Forests 2017, 8, 455. [Google Scholar] [CrossRef]
- Grossiord, C.; Sevanto, S.; Adams, H.D.; Collins, A.D.; Dickman, L.T.; McBranch, N.; Michaletz, S.T.; Stockton, E.A.; Vigil, M.; McDowell, N.G. Precipitation, not air temperature, drives functional responses of trees in semi-arid ecosystems. J. Ecol. 2017, 105, 163–175. [Google Scholar] [CrossRef]
- Helman, D.; Osem, Y.; Yakir, D.; Lensky, I.M. Relationships between climate, topography, water use and productivity in two key Mediterranean forest types with different water-use strategies. Agric. Meteorol. 2017, 232, 319–330. [Google Scholar] [CrossRef]
- Grossiord, C.; Granier, A.; Gessler, A.; Jucker, T.; Bonal, D. Does Drought Influence the Relationship Between Biodiversity and Ecosystem Functioning in Boreal Forests? Ecosystems 2014, 17, 394–404. [Google Scholar] [CrossRef]
- Aubin, I.; Munson, A.; Cardou, F.; Burton, P.; Isabel, N.; Pedlar, J.; Paquette, A.; Taylor, A.; Delagrange, S.; Kebli, H.; et al. Traits to stay, traits to move: a review of functional traits to assess sensitivity and adaptive capacity of temperate and boreal trees to climate change. Environ. Rev. 2016, 24, 164–186. [Google Scholar] [CrossRef] [Green Version]
- Girardin, M.P.; Bouriaud, O.; Hogg, E.H.; Kurz, W.; Zimmermann, N.E.; Metsaranta, J.M.; de Jong, R.; Frank, D.C.; Esper, J.; Buntgen, U.; et al. No growth stimulation of Canada’s boreal forest under half-century of combined warming and CO2 fertilization. Proc. Natl. Acad. Sci. USA 2016, 113, E8406–E8414. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, M.J.; Engelbrecht, B.M.J.; Joswig, J.; Pereyra, G.; Schuldt, B.; Jansen, S.; Kattge, J.; Landhäusser, S.M.; Levick, S.R.; Preisler, Y.; et al. A synthesis of tree functional traits related to drought-induced mortality in forests across climatic zones. J. Appl. Ecol. 2017, 54, 1669–1686. [Google Scholar] [CrossRef]
- Launiainen, S.; Katul, G.G.; Kolari, P.; Lindroth, A.; Lohila, A.; Aurela, M.; Varlagin, A.; Grelle, A.; Vesala, T. Do the energy fluxes and surface conductance of boreal coniferous forests in Europe scale with leaf area? Chang. Boil. 2016, 22, 4096–4113. [Google Scholar] [CrossRef]
- Launiainen, S.; Katul, G.G.; Laurén, A.; Kolari, P. Coupling boreal forest CO2, H2O and energy flows by a vertically structured forest canopy – Soil model with separate bryophyte layer. Ecol. Model. 2015, 312, 385–405. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Von Caemmerer, S.; Berry, J.A. A biochemical model of photosynthetic CO2 assimilation in leaves of C3 species. Planta 1980, 149, 78–90. [Google Scholar] [CrossRef] [Green Version]
- Medlyn, B.E.; Dreyer, E.; Ellsworth, D.; Forstreuter, M.; Harley, P.C.; Kirschbaum, M.U.F.; Le Roux, X.; Montpied, P.; Strassemeyer, J.; Walcroft, A.; et al. Temperature response of parameters of a biochemically based model of photosynthesis. II. A review of experimental data. Plant Cell Environ. 2002, 25, 1167–1179. [Google Scholar] [CrossRef] [Green Version]
- Kellomäki, S.; Wang, K.-Y. Photosynthetic responses to needle water potentials in Scots pine after a four-year exposure to elevated CO2 and temperature. Tree Physiol. 1996, 16, 765–772. [Google Scholar] [CrossRef] [PubMed]
- Medlyn, B.E.; Duursma, R.A.; Eamus, D.; Ellsworth, D.; Prentice, I.C.; Barton, C.V.M.; Crous, K.Y.; De Angelis, P.; Freeman, M.; Wingate, L. Reconciling the optimal and empirical approaches to modelling stomatal conductance. Chang. Boil. 2011, 17, 2134–2144. [Google Scholar] [CrossRef] [Green Version]
- Zhao, W.; Qualls, R.J. A multiple-layer canopy scattering model to simulate shortwave radiation distribution within a homogeneous plant canopy. N.a. Resour. 2005, 41. [Google Scholar] [CrossRef] [Green Version]
- Zhao, W.; Qualls, R.J. Modeling of long-wave and net radiation energy distribution within a homogeneous plant canopy via multiple scattering processes. N.a. Resour. 2006, 42. [Google Scholar] [CrossRef] [Green Version]
- Schuepp, P.H. Tansley Review No. 59 Leaf Boundary-Layers. New Phytol. 1993, 125, 477–507. [Google Scholar] [CrossRef]
- Campbell, G.S.; Norman, J.M. An Introduction to Environmental Biophysics; Springer Nature: Basingstoke, UK, 1998. [Google Scholar]
- Medlyn, B.E.; Badeck, F.W.; De Pury, D.G.G.; Barton, C.V.M.; Broadmeadow, M.; Ceulemans, R.; De Angelis, P.; Forstreuter, M.; Jach, M.E.; Kellomaki, S.; et al. Effects of elevated [CO2] on photosynthesis in European forest species: a meta-analysis of model parameters. Plant Cell Eniviron. 1999, 22, 1475–1495. [Google Scholar] [CrossRef] [Green Version]
- Baldocchi, D.; Meyers, T. On using eco-physiological, micrometeorological and biogeochemical theory to evaluate carbon dioxide, water vapor and trace gas fluxes over vegetation: A perspective. Agric. Meteorol. 1998, 90, 1–25. [Google Scholar] [CrossRef]
- Kattge, J.; Knorr, W. Temperature acclimation in a biochemical model of photosynthesis: A reanalysis of data from 36 species. Plant Cell 2007, 30, 1176–1190. [Google Scholar] [CrossRef]
- Beerling, D.; Quick, W. A new technique for estimating rates of carboxylation and electron transport in leaves of C3 plants for use in dynamic global vegetation models. Chang. Boil. 1995, 1, 289–294. [Google Scholar] [CrossRef]
- Aerts, R.; Brovkin, V.; Cavender-Bares, J.; Cavender-Bares, J.; Verheijen, L.M.; Cornelissen, J.H.C.; Kattge, J.; Van Bodegom, P.M. Inclusion of ecologically based trait variation in plant functional types reduces the projected land carbon sink in an earth system model. Chang. Boil. 2015, 21, 3074–3086. [Google Scholar]
- A Arain, M.; A Black, T.; Barr, A.G.; Jarvis, P.G.; Massheder, J.M.; Verseghy, D.L.; Nesic, Z. Effects of seasonal and interannual climate variability on net ecosystem productivity of boreal deciduous and conifer forests. Can. J. 2002, 32, 878–891. [Google Scholar] [CrossRef]
- Medlyn, B.E.; Berbigier, P.; Clement, R.; Grelle, A.; Loustau, D.; Linder, S.; Wingate, L.; Jarvis, P.G.; Sigurdsson, B.D.; McMurtrie, R.E. Carbon balance of coniferous forests growing in contrasting climates: Model-based analysis. Agric. Meteorol. 2005, 131, 97–124. [Google Scholar] [CrossRef]
- Niinemets, Ülo Research review. Components of leaf dry mass per area - thickness and density - alter leaf photosynthetic capacity in reverse directions in woody plants. New Phytol. 1999, 144, 35–47. [CrossRef]
- Niinemets, Ülo GLOBAL-SCALE CLIMATIC CONTROLS OF LEAF DRY MASS PER AREA, DENSITY, AND THICKNESS IN TREES AND SHRUBS. Ecology 2001, 82, 453–469. [CrossRef]
- Lin, Y.-S.; Medlyn, B.E.; Duursma, R.A.; Prentice, I.C.; Wang, H.; Baig, S.; Eamus, D.; de Dios,, V.R.; Mitchell, P.; Ellsworth, D.S.; et al. Optimal stomatal behaviour around the world. Nat. Clim. Change 2015, 5, 459. [Google Scholar] [CrossRef]
- Breuer, L.; Eckhardt, K.; Frede, H.-G. Plant parameter values for models in temperate climates. Ecol. Model. 2003, 169, 237–293. [Google Scholar] [CrossRef]
- Bartlett, M.K.; Ollinger, S.V.; Wicklein, H.F.; Hollinger, D.Y.; Richardson, A.D. Canopy-scale relationships between foliar nitrogen and albedo are not observed in leaf reflectance and transmittance within temperate deciduous tree species. Botany 2011, 89, 491–497. [Google Scholar] [CrossRef]
- Dickinson, R.E. Land Surface Processes and Climate—Surface Albedos and Energy Balance. Adv. Geophys. 1983, 25, 305–353. [Google Scholar]
- Hornberger, G.M.; Spear, R.C. Eutrophication in Peel Inlet.1. Problem-Defining Behavior and a Mathematical-Model for the Phosphorus Scenario. Water Res. 1980, 14, 29–42. [Google Scholar] [CrossRef]
- McIntyre, N.; Jackson, B.; Wade, A.; Butterfield, D.; Wheater, H.; Wade, A. Sensitivity analysis of a catchment-scale nitrogen model. J. Hydrol. 2005, 315, 71–92. [Google Scholar] [CrossRef]
- Medici, C.; Wade, A.; Frances, F.; Wade, A. Does increased hydrochemical model complexity decrease robustness? J. Hydrol. 2012, 440, 1–13. [Google Scholar] [CrossRef]
- Ruiz-Pérez, G.; Medici, C.; Latron, J.; Llorens, P.; Gallart, F.; Francès, F.; Ruiz-Pérez, G. Investigating the behaviour of a small Mediterranean catchment using three different hydrological models as hypotheses. Hydrol. Process. 2016, 30, 2050–2062. [Google Scholar] [CrossRef]
- Kolari, P.; Kulmala, L.; Pumpanen, J.; Launiainen, S.; Ilvesniem, H.; Hari, P.; Nikinmaa, E. CO2 exchange and component CO2 fluxes of a boreal Scots pine forest. Boreal Environ. Res. 2009, 14, 761–783. [Google Scholar]
- Kulmala, L.; Launiainen, S.; Pumpanen, J.; Lankreijer, H.; Lindroth, A.; Hari, P.; Vesala, T. H2O and CO2fluxes at the floor of a boreal pine forest. N.a. B: Chem. Phys. Meteorol. 2008, 60, 167–178. [Google Scholar] [Green Version]
- Gao, Y.; Markkanen, T.; Aurela, M.; Mammarella, I.; Thum, T.; Tsuruta, A.; Yang, H.; Aalto, T. Response of water use efficiency to summer drought in a boreal Scots pine forest in Finland. Biogeosciences 2017, 14, 4409–4422. [Google Scholar] [CrossRef] [Green Version]
- Richardson, A.D.; Keenan, T.F.; Migliavacca, M.; Ryu, Y.; Sonnentag, O.; Toomey, M. Climate change, phenology, and phenological control of vegetation feedbacks to the climate system. Agric. Meteorol. 2013, 169, 156–173. [Google Scholar] [CrossRef]
- Gill, A.L.; Gallinat, A.S.; Sanders-DeMott, R.; Rigden, A.J.; Gianotti, D.J.S.; Mantooth, J.A.; Templer, P.H. Changes in autumn senescence in northern hemisphere deciduous trees: A meta-analysis of autumn phenology studies. Ann. Bot. 2015, 116, 875–888. [Google Scholar] [CrossRef]
- Michaletz, S.T.; Weiser, M.D.; McDowell, N.G.; Zhou, J.; Kaspari, M.; Helliker, B.R.; Enquist, B.J. Corrigendum: The energetic and carbon economic origins of leaf thermoregulation. Nat. Plants 2016, 2, 16147. [Google Scholar] [CrossRef]
- Davi, H.; Barbaroux, C.; Dufrene, E.; François, C.; Montpied, P.; Breda, N.; Badeck, F. Modelling leaf mass per area in forest canopy as affected by prevailing radiation conditions. Ecol. Model. 2008, 211, 339–349. [Google Scholar] [CrossRef]
- Xu, F.; Guo, W.; Xu, W.; Wei, Y.; Wang, R. Leaf morphology correlates with water and light availability: What consequences for simple and compound leaves? Prog. Sci. Mater. Int. 2009, 19, 1789–1798. [Google Scholar] [CrossRef]
- Vico, G.; Way, D.A.; Hurry, V.; Manzoni, S. Can leaf net photosynthesis acclimate to rising and more variable temperatures? Plant Cell Environ. 2019. [Google Scholar] [CrossRef]
- Way, D.A.; Yamori, W. Thermal acclimation of photosynthesis: On the importance of adjusting our definitions and accounting for thermal acclimation of respiration. Photosynth. Res. 2014, 119, 89–100. [Google Scholar] [CrossRef]
- Barnabas, B.; Jager, K.; Feher, A. The effect of drought and heat stress on reproductive processes in cereals. Plant Cell Environ. 2008, 31, 11–38. [Google Scholar] [CrossRef] [PubMed]
- De Boeck, H.J.; Bassin, S.; Verlinden, M.; Zeiter, M.; Hiltbrunner, E. Simulated heat waves affected alpine grassland only in combination with drought. New Phytol. 2016, 209, 531–541. [Google Scholar] [CrossRef]
- Suzuki, N.; Rivero, R.M.; Shulaev, V.; Blumwald, E.; Mittler, R. Abiotic and biotic stress combinations. New Phytol. 2014, 203, 32–43. [Google Scholar] [CrossRef] [Green Version]
- Zscheischler, J.; Westra, S.; van den Hurk, B.J.J.M.; Seneviratne, S.I.; Ward, P.J.; Pitman, A.; AghaKouchak, A.; Bresch, D.N.; Leonard, M.; Wahl, T.; et al. Future climate risk from compound events. Nat. Clim. Chang. 2018, 8, 469–477. [Google Scholar] [CrossRef]
- Belyazid, S.; Giuliana, Z. Water limitation can negate the effect of higher temperatures on forest carbon sequestration. Eur. J. 2019, 138, 287–297. [Google Scholar] [CrossRef]
- Smith, N.G.; Keenan, T.F.; Prentice, I.C.; Wang, H.; Wright, I.J.; Niinemets, Ü.; Crous, K.Y.; Domingues, T.F.; Guerrieri, R.; Ishida, F.Y.; et al. Global photosynthetic capacity is optimized to the environment. Ecol. Lett. 2019, 22, 506–517. [Google Scholar] [CrossRef] [PubMed]
- Keleş, Y.; Öncel, I. Response of antioxidative defence system to temperature and water stress combinations in wheat seedlings. N.a. Sci. 2002, 163, 783–790. [Google Scholar] [CrossRef]
- Rizhsky, L.; Liang, H.; Mittler, R. The Combined Effect of Drought Stress and Heat Shock on Gene Expression in Tobacco1. N.a. Physiol. 2002, 130, 1143–1151. [Google Scholar] [Green Version]
- Zscheischler, J.; Seneviratne, S.I. Dependence of drivers affects risks associated with compound events. Sci. Adv. 2017, 3, e1700263. [Google Scholar] [CrossRef]
- McVicar, T.R.; Roderick, M.L.; Donohue, R.J.; Van Niel, T.G. Less bluster ahead? Ecohydrological implications of global trends of terrestrial near-surface wind speeds. Ecohydrol. 2012, 5, 381–388. [Google Scholar] [CrossRef]
- Roderick, M.L.; Rotstayn, L.D.; Farquhar, G.D.; Hobbins, M.T. On the attribution of changing pan evaporation. Geophys. Lett. 2007, 34, L17401–L17403. [Google Scholar] [CrossRef]
- Wright, I.J.; Dong, N.; Maire, V.; Prentice, I.C.; Westoby, M.; Díaz, S.; Gallagher, R.V.; Jacobs, B.F.; Kooyman, R.; Law, E.A.; et al. Global climatic drivers of leaf size. Science 2017, 357, 917–921. [Google Scholar] [CrossRef]
- Niinemets, U. Stomatal conductance alone does not explain the decline in foliar photosynthetic rates with increasing tree age and size in Picea abies and Pinus sylvestris. Tree Physiol. 2002, 22, 515–535. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.W.; Wang, J.R.; Ji, M.F.; Milne, R.I.; Wang, M.H.; Liu, J.-Q.; Shi, S.; Yang, S.-L.; Zhao, C.-M. Higher Thermal Acclimation Potential of Respiration but Not Photosynthesis in Two Alpine Picea Taxa in Contrast to Two Lowland Congeners. PLoS One 2015, 10, e0123248. [Google Scholar] [CrossRef]
- Taeger, S.; Sparks, T.H.; Menzel, A. Effects of temperature and drought manipulations on seedlings of Scots pine provenances. Plant Biol. 2015, 17, 361–372. [Google Scholar] [CrossRef]
- Kurepin, L.V.; Stangl, Z.R.; Ivanov, A.G.; Bui, V.; Mema, M.; Hüner, N.P.; Öquist, G.; Way, D.; Hurry, V. Contrasting acclimation abilities of two dominant boreal conifers to elevated CO2 and temperature. Plant Cell 2018, 41, 1331–1345. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | Range | Range for Mid-to-high Latitude | References | References for Mid-to-high Latitude |
|---|---|---|---|---|
| VCMAX,25 (μmol m–2 s–1) | 20–120 | 35–70 | [39,40,41,42] | [29,39,40,43,44] |
| lt (m) | 0.005–0.15 | * | [11,45,46] | [29] |
| g1 (kPa0.5) | 1–15 | 1.5–7 | [33,38,47] | [29,33,38] |
| β (-) | 0.05–3 | ** | [29,33,38] | [29,33,38] |
| αPAR (-) | 0.02–0.3 | 0.04–0.12 | [48,49,50] | [29,48,49,50] |
| αNIR (-) | 0.1–0.7 | 0.2–0.55 |
| Trait Combination | VCMAX,25 μmol m–2 s–1 | lt m | g1 kPa0.5 | β (-) | αPAR (-) | αNIR (-) | Anet,cum μmol m–2 | TL,max °C | Anet,max μmol m–2s–1 | ∆T90 °C |
|---|---|---|---|---|---|---|---|---|---|---|
| Comb. 1 (LP-HR) | 36.16 | 0.14 | 6.35 | 2.41 | 0.11 | 0.39 | 25235 | 49.50 | 8.15 | 6.02 |
| Comb. 2 (HP-LR) | 57.36 | 0.02 | 2.19 | 0.70 | 0.27 | 0.60 | 32770 | 40.08 | 11.54 | 5.60 |
| Comb. 3 (HP-HR) | 56.93 | 0.10 | 6.53 | 2.34 | 0.19 | 0.49 | 36818 | 46.18 | 12.58 | 5.83 |
| Comb. 4 (LP-LR) | 36.37 | 0.03 | 6.18 | 1.92 | 0.22 | 0.52 | 25600 | 42.14 | 8.18 | 6.52 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruiz-Pérez, G.; Launiainen, S.; Vico, G. Role of Plant Traits in Photosynthesis and Thermal Damage Avoidance under Warmer and Drier Climates in Boreal Forests. Forests 2019, 10, 398. https://doi.org/10.3390/f10050398
Ruiz-Pérez G, Launiainen S, Vico G. Role of Plant Traits in Photosynthesis and Thermal Damage Avoidance under Warmer and Drier Climates in Boreal Forests. Forests. 2019; 10(5):398. https://doi.org/10.3390/f10050398
Chicago/Turabian StyleRuiz-Pérez, Guiomar, Samuli Launiainen, and Giulia Vico. 2019. "Role of Plant Traits in Photosynthesis and Thermal Damage Avoidance under Warmer and Drier Climates in Boreal Forests" Forests 10, no. 5: 398. https://doi.org/10.3390/f10050398