Size–Density Trajectory in Regenerated Maritime Pine Stands after Fire

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area Characteristics
2.2. Supporting Material
2.3. Selection of Maximum Values of Size–Density
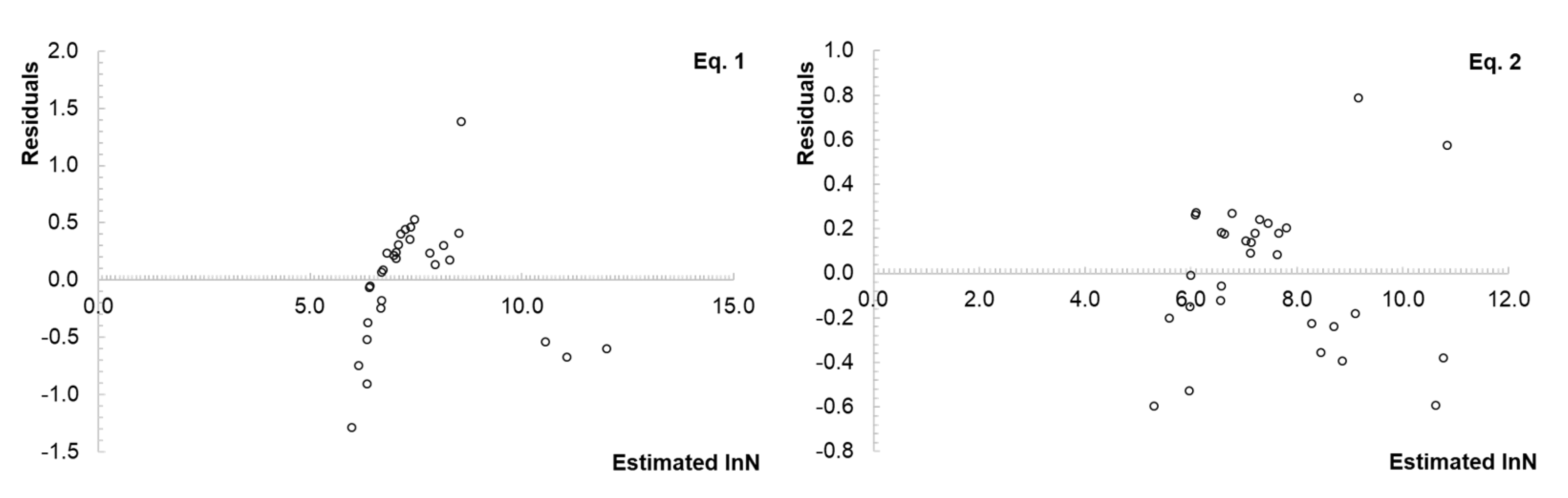
2.4. Size–Density Relationship Modeling and Statistical Analysis
3. Results
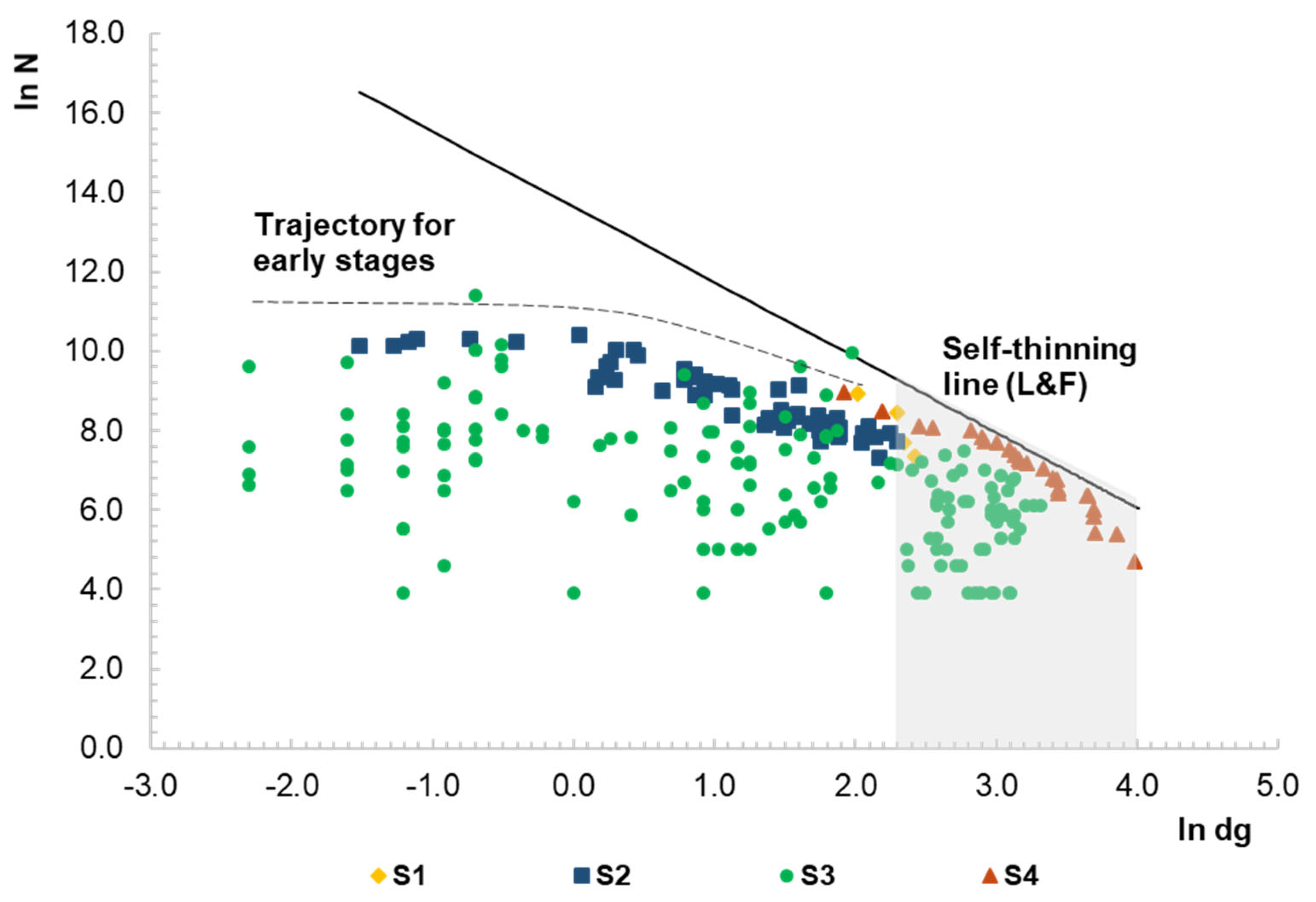
3.1. Representativeness of the Data Set and Pattern of the Density–Size Trajectory
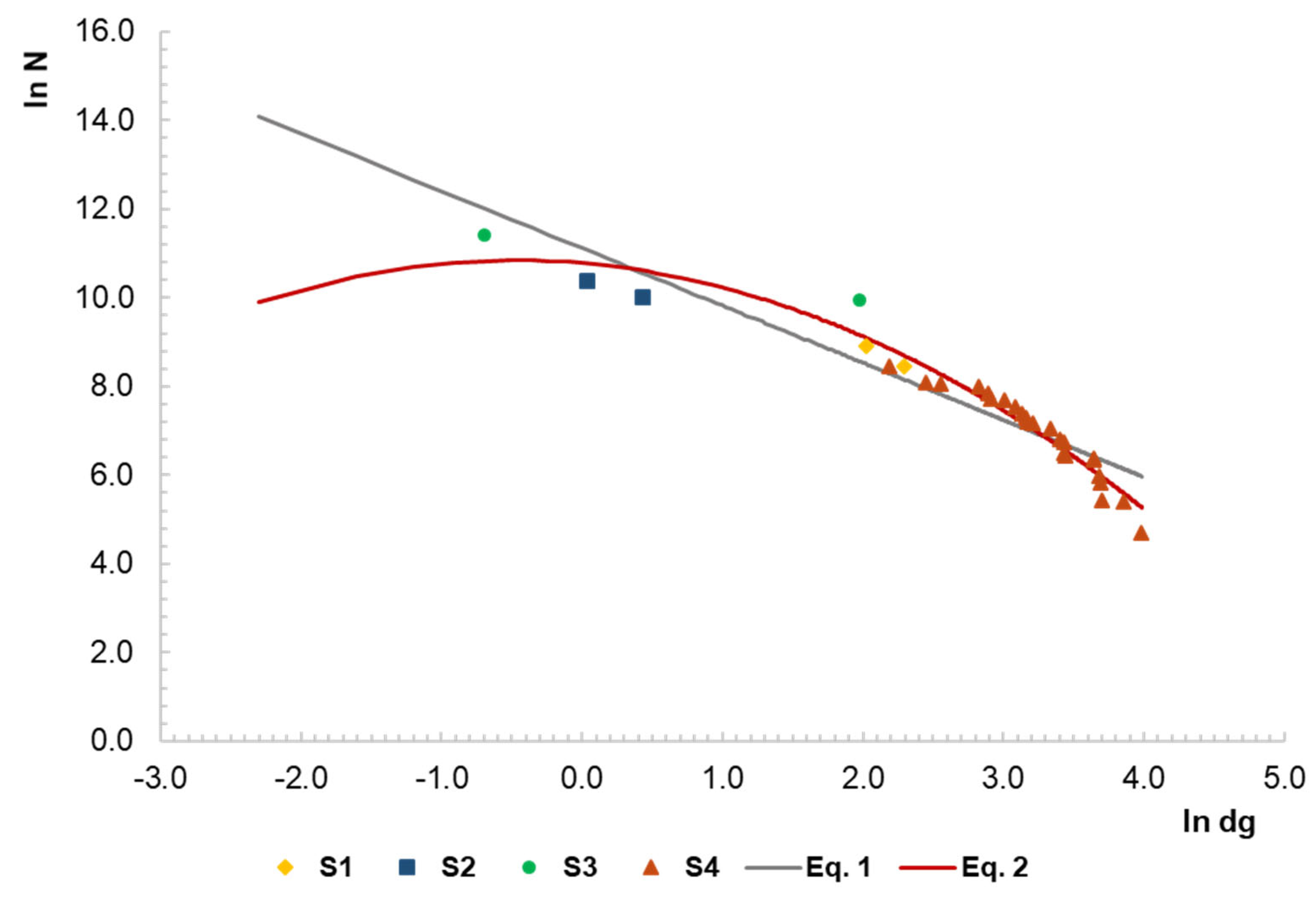
3.2. Maximum Size–Density Sample Data
3.3. Representation of the Size–Density Pattern for the Border Points and Fitted Lines
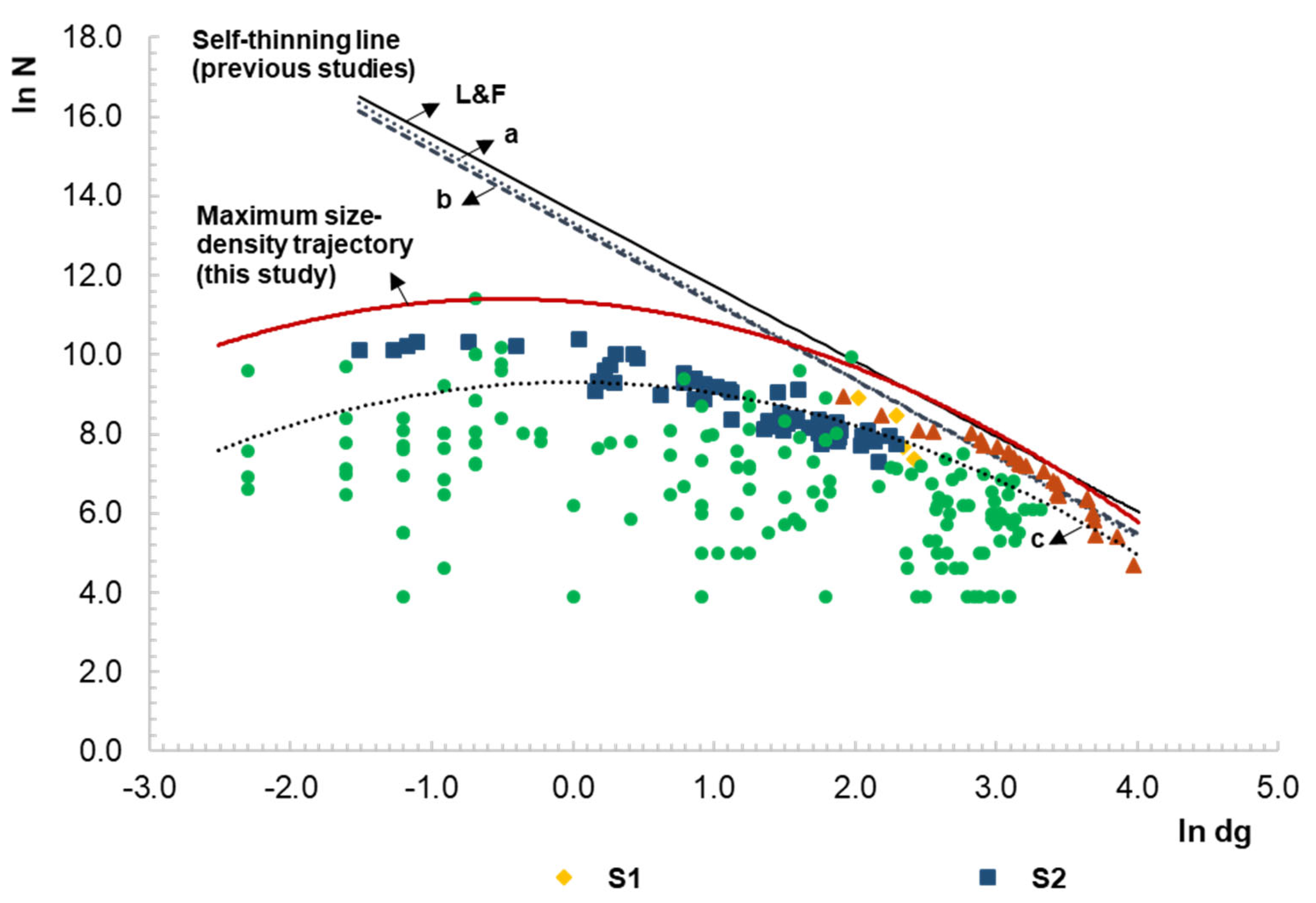
3.4. Maximum Size–Density Trajectory over All the Stages of Development
4. Discussion
4.1. Representativeness of the Database
4.2. Size–Density Trajectory and Self-Thinning Line
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Doerr, S.H.; Santin, C. The ‘wildfire problem’: Perceptions and realities in a changing world. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2016, 371, 1–10. [Google Scholar] [CrossRef] [PubMed]
- ICNF. I F N 6—Áreas dos Usos do Solo e das Espécies Florestais de Portugal Continental 1995–2005–2010. Resultados Preliminares. 2013. Available online: https://doi.org/10.1016/j.rasd.2006.07.002 (accessed on 11 January 2019).
- Verkerk, P.J.; Martinez de Arano, I.; Palahí, M. The bio-economy as an opportunity to tackle wildfires in Mediterranean forest ecosystems. For. Policy Econ. 2018, 86, 1–3. [Google Scholar] [CrossRef]
- AFN. FloreStat, 5th National Forest Inventory Information Retrieval Tool; Ministério da Agricultura, do Desenvolvimento Rural e das Pescas: Lisboa, Portugal, 2010; Available online: http://www2.icnf.pt/portal/florestas/ifn/ifn5/rel-fin (accessed on 26 January 2018).
- DR, Resolução do Conselho de Ministros n° 114/2006, Estratégia Nacional Para as Florestas. Diário da República, I Série—n° 179 de 15 de Setembro 2006. Available online: http://www.dre.pt/pdf1s/2006/07/13800/50145029.pdf (accessed on 20 April 2018).
- Fernandes, P.M.; Rigolot, E. The fire ecology and management of maritime pine (Pinus pinaster Ait.). For. Ecol. Manag. 2007, 241, 1–13. [Google Scholar] [CrossRef]
- Freitas, T.M.D. Avaliação da Biomassa em Regeneraçã de Pinheiro Bravo Pós Fogo; Final Project Report; UTAD: Vila Real, Portugal, 2013; p. 57. [Google Scholar]
- Rodrigues, M.R.C. Análise Multi-Temporal de Crescimento de Espécies Florestais. Criação de um SIG Para a Sua Gestão. Master Thesis, Universidade de Trás-os-Montes e Alto Douro, Vila Real, Portugal, 2008. [Google Scholar]
- Alegria, C.; Pedro, N.; do Carmo Horta, M.; Roque, N.; Fernandez, P. Ecological envelope maps and stand production of eucalyptus plantations and naturally regenerated maritime pine stands in the central inland of Portugal. For. Ecol. Manag. 2019, 432, 327–344. [Google Scholar] [CrossRef]
- Calvo, L.; Santalla, S.; Valbuena, L.; Marcos, E.; Tárrega, R.; Luis-Calabuig, E. Post-fire natural regeneration of a Pinus pinaster forest in NW Spain. Plant Ecol. 2008, 197, 81–90. [Google Scholar] [CrossRef]
- Tapias, R.; Climent, J.; Pardos, J.A.; Gil, L. Life histories of Mediterranean pines. Plant. Ecol. 2004, 171, 53–68. [Google Scholar] [CrossRef]
- Martínez-Sánchez, J.J.; Marin, A.; Herranz, J.P.; Ferrandis, P.; De las Heras, J. Effects of high temperatures on germination of Pinus halepensis Mill. and P. pinaster Aiton subsp, pinaster seeds in southeast Spain. J. Vegetatio 1995, 116, 69–72. [Google Scholar]
- Moya, D.; Espelta, J.M.; López-Serrano, F.R.; Eugenio, M.; Heras, J.D. Natural post-fire dynamics and serotiny in 10-year-old Pinus halepensis Mill. stands along a geographic gradient. Int. J. Wildl. Fire 2008, 17, 287–292. [Google Scholar] [CrossRef]
- Maia, P.; Keizer, J.; Vasques, A.; Abrantes, N.; Roxo, L.; Fernandes, P.; Ferreira, A.; Moreira, F. Post-fire plant diversity and abundance in pine and eucalypt stands in Portugal: Effects of biogeography, topography, forest type and post-fire management. For. Ecol. Manag. 2014, 334, 154–162. [Google Scholar] [CrossRef]
- Zeide, B. Analysis of the 3/2 Power Law of Self-Thinning. For. Sci. 1987, 33, 517–537. [Google Scholar]
- Reineke, L.H. Perfecting a stand-density index for even-aged forests. J. Agric. Res. 1933, 46, 627–638. [Google Scholar]
- Yoda, K. Self-thinning in overcrowded pure stands under cultivated and natural conditions (Intraspecific competition among higher plants. XI). J. Inst. Polytech. Osaka City Univ. Ser. D 1963, 14, 107–129. [Google Scholar]
- Harper, J.L. Population Biology of Plants; Academic Press: Cambridge, MA, USA, 1977; ISBN 9780123258526. [Google Scholar]
- Brunet-Navarro, P.; Sterck, F.J.; Vayreda, J.; Martinez-Vilalta, J.; Mohren, G.M.J. Self-thinning in four pine species: An evaluation of potential climate impacts. Ann. For. Sci. 2016, 73, 1025–1034. [Google Scholar] [CrossRef]
- Panayotov, M.; Kulakowski, D.; Tsvetanov, N.; Krumm, F.; Berbeito, I.; Bebi, P. Climate extremes during high competition contribute to mortality in unmanaged self-thinning Norway spruce stands in Bulgaria. For. Ecol. Manag. 2016, 369, 74–88. [Google Scholar] [CrossRef]
- Aguirre, A.; del Río, M.; Condés, S. Intra- and inter-specific variation of the maximum size-density relationship along an aridity gradient in Iberian pinewoods. For. Ecol. Manag. 2018, 411, 90–100. [Google Scholar] [CrossRef]
- Andrews, C.; Weiskittel, A.; D’Amato, A.W.; Simons-Legaard, E. Variation in the maximum stand density index and its linkage to climate in mixed species forests of the North American Acadian Region. For. Ecol. Manag. 2018, 417, 90–102. [Google Scholar] [CrossRef]
- Zeide, B. A relationship between size of trees and their number. For. Ecol. Manag. 1995, 72, 265–272. [Google Scholar] [CrossRef]
- Cao, Q.V.; Dean, T.J.; Baldwin, V.C. Modeling the size-density relationship in direct-seeded Slash pine stands. For. Sci. 2000, 46, 317–321. [Google Scholar]
- Monserud, R.A.; Ledermann, T.; Sterba, H. Are self-thinning constraints needed in a tree-specific mortality model? For. Sci. 2005, 50, 848–858. [Google Scholar]
- Charru, M.; Seynave, I.; Morneau, F. Significant differences and curvilinearity in the self-thinning relationships of 11 temperate tree species assessed from forest inventory data. Ann. For. Sci. 2012, 69, 195–205. [Google Scholar] [CrossRef]
- Fonseca, T.; Monteiro, L.; Enes, T.; Cerveira, A. Self-thinning dynamics in cork oak woodlands: Providing a baseline for managing density. For. Syst. 2017, 26, 1–10. [Google Scholar] [CrossRef]
- Luis, J.S.; Fonseca, T.F. The allometric model in the stand density management of Pinus pinaster Ait. in Portugal. Ann. For. Sci. 2004, 61, 807–814. [Google Scholar] [CrossRef]
- Fonseca, T.; Parresol, B.; Marques, C.; de Coligny, F. Models to Implement a Sustainable Forest Management—An Overview of the ModisPinaster Model. In Sustainable Forest Management/Book 1; Martín García, J., Diez Casero, J.J., Eds.; InTech—Open Access Publisher: Rijeka, Croatia, 2012; pp. 321–338. ISBN 978-953-51-0621-0. [Google Scholar]
- Riofrío, J.; Del Río, M.; Bravo, F. Mixing effects on growth efficiency in mixed pine forests. Forestry 2017, 90, 381–392. [Google Scholar] [CrossRef]
- Rivas-Martinez, S.; Fernández-González, F.; Loidi, J.; Lousa, M.; Penas, A. Vascular plant communities of Spain and Portugal. Addenda to the syntaxonomical checklist of 2001. Part II. In Itinera Geobotanica; Leon, Spain, 2002; Volume 15, ISSN 0213-8530. Available online: https://webs.ucm.es/info/cif/book/addenda/addenda1_00.htm (accessed on 13 November 2018).
- SNIRH. Sistema Nacional de Informação de Recursos Hídricos. Available online: http://www.snirh.pt (accessed on 13 November 2018).
- Marques, C.P. Evaluating site quality of even-aged maritime pine stands in northern Portugal using direct and indirect methods. For. Ecol. Manag. 1991, 41, 193–204. [Google Scholar] [CrossRef]
- Fonseca, T.J.F. Modelação do Crescimento, Mortalidade e Distribuição Diamétrica, do Pinhal Bravo no Vale do Tâmega. Ph.D. Thesis, Universidade de Trás-os-Montes e Alto Douro, Vila Real, Portugal, 2004. [Google Scholar]
- Keeney, R.; Raiffa, H.; Rajala, D. Decisions with Multiple Objectives: Preferences and Value Trade-Offs. Syst. Man Cybern. IEEE Trans. 1979, 9, 403. [Google Scholar] [CrossRef]
- Neter, J.; Kutner, M.; Nachtsheim, C.; Wassermaniption, W. Applied Linear Statistical Models, 3rd ed.; Irwin: Chicago, IL, USA, 1996. [Google Scholar]
- Myers, R.H. Classical and Modern Regression with Applications; Duxbury advanced series in statistics and decision sciences; PWS-KENT: Belmont, CA, USA, 1990; ISBN 9780534922412. [Google Scholar]
- Ningre, F.; Ottorini, J.M.; Le Goff, N. Modeling size-density trajectories for even-aged beech (Fagus silvatica L.) stands in France. Ann. For. Sci. 2016, 73, 765–776. [Google Scholar] [CrossRef]
- Zhang, X.; Cao, Q.V.; Duan, A.; Zhang, J. Self-thinning trajectories of Chinese fir plantations in Southern China. For. Sci. 2016, 62, 594–599. [Google Scholar] [CrossRef]
- Ningre, F.; Ottorini, J.-M.; Le Goff, N. Size-density trajectories for even-aged sessile oak (Quercus petraea (Matt.) Liebl.) and common beech (Fagus sylvatica L.) stands revealing similarities and differences in the mortality process. Ann. For. Sci. 2019, 76, 73. [Google Scholar] [CrossRef]
- Lauer, C.J.; Montgomery, C.A.; Dietterich, T.G. Spatial interactions and optimal forest management on a fire-threatened landscape. For. Policy Econ. 2017, 83, 107–120. [Google Scholar] [CrossRef]
- Fonseca, T.F.; Duarte, J.C. A silvicultural stand density model to control understory in maritime pine stands. iForest 2017, 10, 829–836. [Google Scholar] [CrossRef]
- Fernandes, P.M. Fire-smart management of forest landscapes in the Mediterranean basin under global change. Landsc. Urban. Plan. 2013, 110, 175–182. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | n | N (trees.ha−1) | dg (cm) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Min | Mean | Max | sd | Min | Mean | Max | sd | ||
| S1 | 4 | 1560 | 3985 | 7500 | 2708 | 7.5 | 9.8 | 11.2 | 1.6 |
| S2 | 59 | 1500 | 9536 | 32,500 | 8549 | 0.2 | 4.0 | 9.9 | 2.6 |
| S3 | 153 | 50 | 3091 | 90,000 | 8460 | 0.1 | 8.0 | 27.5 | 8.0 |
| S4 | 25 | 110 | 1724 | 7680 | 1705 | 6.8 | 27.2 | 53.3 | 12.0 |
| Source | n | N (trees.ha−1) | dg (cm) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Min | Mean | Max | sd | Min | Mean | Max | sd | ||
| Border points | 30 | 110 | 7121 | 90,000 | 17,366 | 0.5 | 23.3 | 53.3 | 14.1 |
| Model (Equation) | Estimates (Stand Error) | Fit Statistics | ||||
|---|---|---|---|---|---|---|
| Intercept | r | s | R2adj | RMSE | ||
| Equation (1) | 11.115 (0.270) | −1.290 (0.090) | - | - | 0.876 | 0.541 |
| Equation (2) | 12.969 (0.328) | −1.832 (0.100) | −0.280 (0.042) | 2.796 | 0.954 | 0.341 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Enes, T.; Lousada, J.; Aranha, J.; Cerveira, A.; Alegria, C.; Fonseca, T. Size–Density Trajectory in Regenerated Maritime Pine Stands after Fire. Forests 2019, 10, 1057. https://doi.org/10.3390/f10121057
Enes T, Lousada J, Aranha J, Cerveira A, Alegria C, Fonseca T. Size–Density Trajectory in Regenerated Maritime Pine Stands after Fire. Forests. 2019; 10(12):1057. https://doi.org/10.3390/f10121057
Chicago/Turabian StyleEnes, Teresa, José Lousada, José Aranha, Adelaide Cerveira, Cristina Alegria, and Teresa Fonseca. 2019. "Size–Density Trajectory in Regenerated Maritime Pine Stands after Fire" Forests 10, no. 12: 1057. https://doi.org/10.3390/f10121057