

A Critical Review on the Status and Progress of Microalgae Cultivation in Outdoor Photobioreactors Conducted over 35 Years (1986–2021)
Abstract
:1. Introduction
2. Drivers of Algae Research
3. Trends and Concerns of Outdoor Algal Cultivation Methods
3.1. Ideal vs. Non-Ideal Conditions
3.2. Challenges and Limitations of Outdoor Experimentations
3.3. Batch Culture
3.4. Online Monitoring Tool
3.5. Virtual Laboratory
4. Design Question of Outdoor PBR Systems
5. Current Scenario of Outdoor Algal Cultivation
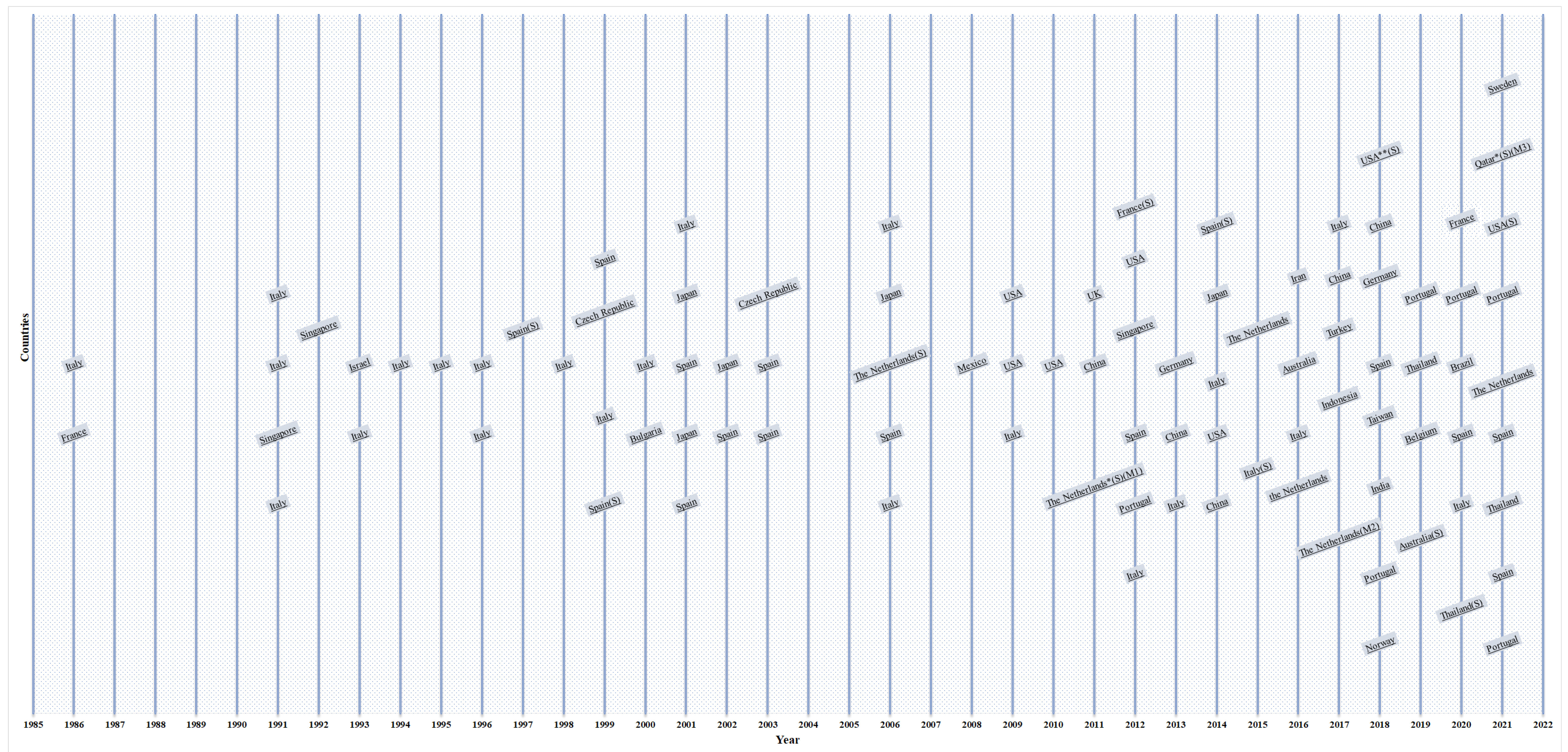
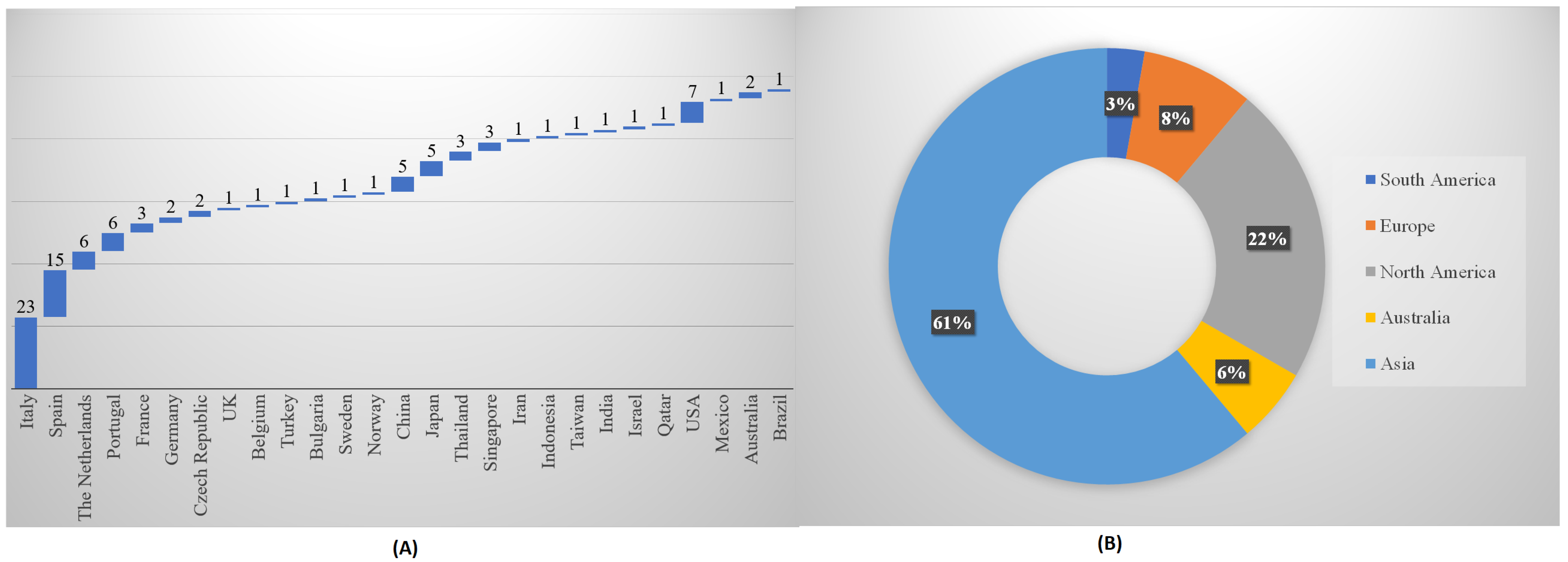
5.1. Chronological and Geographical Progress of Algal Research in Outdoor PBRs
5.2. Topics Covered by the Studies
5.3. Simulation Studies
5.4. Algae Species, Growth Media, and PBRs Used in the Studies

{kind=link}
{kind=link}
| Algae Species | Algae Growth Medium | PBRs |
|---|---|---|
| Alexandrium minutum [47], Anabaena azollae [48], Anabaena siamensis [49], Arthrospira platensis [22,50,51], Chaetoceros calcitrans [52], Chlamydomonas reinhardtii [25,53], Chlorella ellipsoidea [54], Chlorella pyrenoidosa [55,56,57], Chlorella sorokiniana [39,58,59], Chlorella sp. [60], Chlorella vulgaris [10,35,54,61,62,63], Chlorella zofingiensis [64,65], Chlorococcum littorale [52,66,67], Coccomyxa onubensis [68], Desmodesmus sp. [69], Fistulifera sp. (formerly Navicula sp.) [23], Galdieria sulphuraria [70], Haematococcus pluvialis [71], Heterosigma akashiwo [47], Karlodinium veneficum [47,72], Monoraphidium sp. [73], Monodus subterraneus [74], Muriellopsis sp. [75], Nannochloris atomus [76], Nannochloropsis oceanica [77,78,79,80], Nannochloropsis oculate [81,82], Nannochloropsis salina [81], Nannochloropsis granulata [83], Nannochloropsis sp. [84,85,86,87], Neochloris oleoabundans [67], Phaeodactylum tricornutum [34,88,89,90,91,92,93,94,95,96,97,98,99,100,101], Porphyridium cruentum [102], Pseudanabaena limnetica [37], Rhodobacter capsulatus [103], Rhodobacter sphearoides [53], Rhodopseudomonas palustris [104,105], Scenedesmus obliquus [62,106], Scenedesmus obtusus [69], Skeletonema marinoi [83], Spirulina maxima [107], Spirulina platensis [48,49,107,108,109,110,111,112,113,114,115], Synechocystis aquatilis [116], Tetradesmus obliquus [98], Tetraselmis suecica [117,118], Tetraselmis sp. [119,120], Thalassiosira pseudonana [34], Thalassiosira sp. [121], Tisochrysis lutea [93,94], Unknown wild species [24] | Lab-made chemical solutions: f/2 salts solution [101], Guillard’s f/2 medium [23,80], Jaworski formulation nutrients [106], Modified Zarouk’s medium [49], BG-11 medium [24,37,98,122], Mann and Myers’ medium [88,90,92], Modified SOT inorganic medium [123], Modified Ukeles medium [99], Modified van Niel medium (vN-B) [105], Norsker’s solution (new solution) [67], ½ SWES [98] Fertilizer solutions: Agricultural fertilizer-based media such as NPK-fertilizer solution, urea, and triple superphosphate [36,55,64,68,121], Flory Basic Fertilizer 1 (Euflor, Germany) with KNO3 and NH4NO3 [62] Others: Deep-sea water-based medium [77], Local mountain spring water [64], Enriched seawater [121], UV-treated natural seawater [85], Artificial seawater medium [66], Wastewater [10,35,54] Hybrid solution: Fresh f/2 (−Si) medium in seawater [63,120], Basal medium supplemented with f/2 salts [100], Groundwater supplemented with commercial NaCl [95], Modified seawater BG11 medium [37], Seawater supplemented with NutriBloom® plus [82,93] | Bubble column [23,47,71,72,74,99], Vertical column [23,68,117], Air lift [50,91,99,121], Flat panel [34,36,37,51,53,57,65,72,78,80,84,119], Vertical flat panel [38,70,82,116,123], Tubular (all the articles except where conical flasks were used and the articles cited in this column), Tubular loop [56], Vertical tubular [24,64,84], Horizontal tubular [36,66,84,86,87,106,119], Curved tubular [112], Helical tubular [90,124], Air-lift tubular [49,89], Air-lift bag column [69], Plastic bag type [77], Vertical cylindrical [63], Vertical alveolar panel [48], Dome-shaped reactor [52], Ring shaped [125], Floating horizontal [76], Underwater tubular [104,105], Penthouse-roof PBR (tubular) [108], Green Wall Panel (flat panel) [85,93,96,97,118,122], AGS 3 and AGS 4 (industrial outdoor PBRs) [81] |
| Parameters | Experimental | Simulation and Modelling | Total |
|---|---|---|---|
| Number of articles | 84 | 12 | 96 |
| Patent application | 4 | 4 | |
| PE | 18 | 1 | 19 |
| Multiple months | 17 | 17 | |
| Light intensity and dilution | 18 | 3 | 21 |
| Types of light and optical path | 4 | 1 | 5 |
| Temperature control | 12 | 3 | 15 |
| Outdoor adaptation | 9 | 9 | |
| Lipid | 14 | 14 | |
| Fatty acid | 9 | 9 | |
| EPA | 5 | 5 | |
| Carbohydrate | 4 | 4 | |
| Protein | 6 | 6 | |
| Lutein | 2 | 2 | |
| Fucoxanthin | 2 | 2 | |
| Astaxanthin and carotenoid | 2 | 2 | |
| Biochemical composition | 8 | 8 | |
| Novel and/or modified design | 18 | 3 | 21 |
| Economics | 15 | 3 | 18 |
| Comparison between different types of outdoor PBRs | 13 | 1 | 14 |
| Comparison of outdoor PBRs with indoor PBRs | 8 | 8 | |
| Comparison of outdoor PBRs with outdoor ponds | 8 | 8 | |
| Arrangement/orientation of outdoor PBRs | 6 | 2 | 8 |
| CO2 sequestration | 6 | 2 | 8 |
| Environmental impact | 4 | 1 | 5 |
| Bioenergy potential | 10 | 1 | 11 |
| Algae growth medium | 7 | 7 | |
| O2 accumulation | 5 | 1 | 6 |
| Fluid velocity | 8 | 8 | |
| Biomass loss at night/winter | 7 | 7 | |
| The initial concentration of pure strain | 6 | 6 | |
| Bacteria | 2 | 2 | |
| Pore size | 1 | 1 | |
| Algal species screening technology | 1 | 1 | |
| Comparison of multiple strains | 2 | 1 | 3 |
| Comparison of different operational modes | 1 | 1 | 2 |
| Comparison between multiple cities in one country | 1 | 1 | |
| Comparison between multiple countries | 1 | 2 | 3 |
6. A Review of the Findings on Upstream Processes
6.1. Temperature Control
6.2. Outdoor Adaption
6.3. Light Intensity and Dilution
6.4. Types of Light and Optical Path
6.5. Effect of Varying Outdoor Conditions
6.6. Photosynthetic Efficiency (PE)
6.7. Novel and Modified Design
6.8. Arrangement/Orientation of Outdoor PBRs
6.9. Fluid Velocity
6.10. O2 Accumulation
6.11. Bacteria
6.12. Biomass Loss at Night/Winter
6.13. Initial Concentration of Pure Strain and Harvest Rate
7. Downstream Processes
8. Comparisons
8.1. Comparison of Outdoor PBRs with Indoor PBRs
8.2. Comparison of Outdoor PBRs with Outdoor Ponds
8.3. Comparison between Different Types of Outdoor PBRs
9. Impacts
9.1. Economics
9.2. Environmental Impact
9.3. Bioenergy Potential
10. Conclusions and Outlook
Supplementary Materials
Funding
Data Availability Statement
Conflicts of Interest
References
- Singh, J.; Dhar, D.W. Overview of carbon capture technology: Microalgal biorefinery concept and state-of-the-art. Front Mar. Sci. 2019, 6, 29. [Google Scholar] [CrossRef] [Green Version]
- Bhadra, S.; Salam, P.A.; Sarker, N.K. Microalgae-based biodiesel production in open raceway ponds using coal thermal flue gas: A case of West Bengal, India. Environ. Qual. Manag. 2020, 29, 27–36. [Google Scholar] [CrossRef]
- Sarker, N.K. Theoretical effect of concentration, circulation rate, stages, pressure and temperature of single amine and amine mixture solvents on gas sweetening performance. Egypt. J. Pet. 2016, 25, 343–354. [Google Scholar] [CrossRef] [Green Version]
- Sarker, N.K. Effect of Concentration, Circulation Rate, Stages, Pressure and Temperature of Pure Glycols on Natural Gas Dehydration Performance and Sales Gas Characteristics. Pet. Coal 2021, 62, 477–502. [Google Scholar]
- Field, C.B.; Behrenfeld, M.J.; Randerson, J.T.; Falkowski, P. Primary production of the biosphere: Integrating terrestrial and oceanic components. Science 1998, 281, 237–240. [Google Scholar] [CrossRef] [Green Version]
- Gao, G.; Wu, M.; Fu, Q.; Li, X.; Xu, J. A two-stage model with nitrogen and silicon limitation enhances lipid productivity and biodiesel features of the marine bloom-forming diatom Skeletonema costatum. Bioresour. Technol. 2019, 289, 121717. [Google Scholar] [CrossRef]
- Jiang, X.; Han, Q.; Gao, X.; Gao, G. Conditions optimising on the yield of biomass, total lipid, and valuable fatty acids in two strains of Skeletonema menzelii. Food Chem. 2016, 194, 723–732. [Google Scholar] [CrossRef]
- Latsos, C.; van Houcke, J.; Timmermans, K.R. The Effect of Nitrogen Starvation on Biomass Yield and Biochemical Constituents of Rhodomonas sp. Front. Mar. Sci. 2020, 7, 900. [Google Scholar] [CrossRef]
- EnerGaia–EnerGaia–Producers of Fresh, Nutritional and Sustainable Spirulina. Available online: https://energaia.com/ (accessed on 26 March 2023).
- Sarker, N.K.; Salam, P.A. Indoor and outdoor cultivation of Chlorella vulgaris and its application in wastewater treatment in a tropical city—Bangkok, Thailand. SN Appl. Sci. 2019, 1, 1645. [Google Scholar] [CrossRef] [Green Version]
- Algalif ehf | Leading Supplier of High-Grade Natural Astaxanthin–Algalif, Icelandic Producer of Pure, High-Grade, Natural Astaxanthin from Microalgae. Available online: https://algalif.is/ (accessed on 26 March 2023).
- Griffiths, M.J.; Harrison, S.T.L. Lipid productivity as a key characteristic for choosing algal species for biodiesel production. J. Appl. Phycol. 2009, 21, 493–507. [Google Scholar] [CrossRef]
- Dahlin, L.R.; Van Wychen, S.; Gerken, H.G.; McGowen, J.; Pienkos, P.T.; Posewitz, M.C.; Guarnieri, M.T. Down-selection and outdoor evaluation of novel, halotolerant algal strains for winter cultivation. Front Plant. Sci. 2018, 871, 1513. [Google Scholar] [CrossRef] [Green Version]
- Leinemann, F.; Mabilia, V. European Union Legislation and Policies Relevant for Algae. In Grand Challenges in Biology and Biotechnology; Hallmann, A., Rampelotto, P.H., Eds.; Springer: Berlin/Heidelberg, Germany, 2019; pp. 577–591. [Google Scholar]
- Rumin, J.; de Oliveira Junior, R.G.; Bérard, J.B.; Picot, L. Improving microalgae research and marketing in the european atlantic area: Analysis of major gaps and barriers limiting sector development. Mar. Drugs 2021, 19, 319. [Google Scholar] [CrossRef]
- Onwezen, M.C.; Bouwman, E.P.; Reinders, M.J.; Dagevos, H. A systematic review on consumer acceptance of alternative proteins: Pulses, algae, insects, plant-based meat alternatives, and cultured meat. Appetite 2021, 159, 105058. [Google Scholar] [CrossRef]
- Birch, D.; Skallerud, K.; Paul, N.A. Who are the future seaweed consumers in a Western society? Insights from Australia. Br. Food J. 2019, 121, 603–615. [Google Scholar] [CrossRef] [Green Version]
- Moshood, T.D.; Nawanir, G.; Mahmud, F. Microalgae biofuels production: A systematic review on socioeconomic prospects of microalgae biofuels and policy implications. Environ. Chall. 2021, 5, 100207. [Google Scholar] [CrossRef]
- Loke Show, P. Global market and economic analysis of microalgae technology: Status and perspectives. Bioresour. Technol. 2022, 357, 127329. [Google Scholar] [CrossRef]
- Rao, N.R.; Tamburic, B.; Doan, Y.T.; Nguyen, B.D.; Henderson, R.K. Algal biotechnology in Australia and Vietnam: Opportunities and challenges. Algal. Res. 2021, 56, 102335. [Google Scholar] [CrossRef]
- Araújo, R.; Vázquez Calderón, F.; Sánchez López, J.; Azevedo, I.C.; Bruhn, A.; Fluch, S.; Garcia Tasende, M.; Ghaderiardakani, F.; Ilmjärv, T.; Laurans, M.; et al. Current Status of the Algae Production Industry in Europe: An Emerging Sector of the Blue Bioeconomy. Front Mar. Sci. 2021, 7, 626389. [Google Scholar] [CrossRef]
- Pruvost, J.; Cornet, J.F.; Goetz, V.; Legrand, J. Theoretical investigation of biomass productivities achievable in solar rectangular photobioreactors for the cyanobacterium Arthrospira platensis. Biotechnol. Prog. 2012, 28, 699–714. [Google Scholar] [CrossRef]
- Sato, R.; Maeda, Y.; Yoshino, T.; Tanaka, T.; Matsumoto, M. Seasonal variation of biomass and oil production of the oleaginous diatom Fistulifera sp. In outdoor vertical bubble column and raceway-type bioreactors. J. Biosci. Bioeng. 2014, 117, 720–724. [Google Scholar] [CrossRef] [Green Version]
- Huo, S.; Shang, C.; Wang, Z.; Zhou, W.; Cui, F.; Zhu, F.; Yuan, Z.; Dong, R. Outdoor Growth Characterization of an Unknown Microalga Screened from Contaminated Chlorella Culture. Biomed. Res. Int. 2017, 2017, 5681617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, E.; Pruvost, J.; He, X.; Munipalli, R.; Pilon, L. Design tool and guidelines for outdoor photobioreactors. Chem. Eng. Sci. 2014, 106, 18–29. [Google Scholar] [CrossRef] [Green Version]
- Norsker, N.H.; Cuaresma, M.; Uronen, P.; Barbosa, M.J.; Wijffels, R. Developing microalgal oil production for an outdoor photobioreactor. J. Appl. Phycol. 2021, 33, 1315–1325. [Google Scholar] [CrossRef]
- Kondo, T.; Chen, W.J.; Schlau-Cohen, G.S. Single-molecule fluorescence spectroscopy of photosynthetic systems. Chem. Rev. 2017, 117, 860–898. [Google Scholar] [CrossRef]
- Lavine, B.K.; Workman, J. Chemometrics: Past, Present, and Future; ACS Publication: Washington, DC, USA, 2005; pp. 1–13. [Google Scholar]
- Sá, M.; Bertinetto, C.G.; Ferrer-Ledo, N.; Jansen, J.J.; Wijffels, R.; Crespo, J.G.; Barbosa, M.; Galinha, C.F. Fluorescence spectroscopy and chemometrics for simultaneous monitoring of cell concentration, chlorophyll and fatty acids in Nannochloropsis oceanica. Sci. Rep. 2020, 10, 7688. [Google Scholar] [CrossRef]
- Sá, M.; Ramos, A.; Monte, J.; Brazinha, C.; Galinha, C.F.; Crespo, J.G. Development of a monitoring tool based on fluorescence and climatic data for pigments profile estimation in Dunaliella salina. J. Appl. Phycol. 2020, 32, 363–373. [Google Scholar] [CrossRef]
- Sá, M.; Monte, J.; Brazinha, C.; Galinha, C.F.; Crespo, J.G. Fluorescence coupled with chemometrics for simultaneous monitoring of cell concentration, cell viability and medium nitrate during production of carotenoid-rich Dunaliella salina. Algal. Res. 2019, 44, 101720. [Google Scholar] [CrossRef]
- Sá, M.; Monte, J.; Brazinha, C.; Galinha, C.F.; Crespo, J.G. 2D Fluorescence spectroscopy for monitoring Dunaliella salina concentration and integrity during membrane harvesting. Algal. Res. 2017, 24, 325–332. [Google Scholar] [CrossRef]
- Dormido, R.; Sánchez, J.; Duro, N.; Dormido-Canto, S.; Guinaldo, M.; Dormido, S. An interactive tool for outdoor computer controlled cultivation of microalgae in a tubular photobioreactor system. Sensors 2014, 14, 4466–4483. [Google Scholar] [CrossRef] [Green Version]
- Slegers, P.M.; Wijffels, R.H.; van Straten, G.; van Boxtel, A.J.B. Design scenarios for flat panel photobioreactors. Appl. Energy 2011, 88, 3342–3353. [Google Scholar] [CrossRef]
- Sarker, N.K.; Salam, P.A. Design of batch algal cultivation systems and ranking of the design parameters. Energy Ecol. Environ. 2020, 5, 196–210. [Google Scholar] [CrossRef]
- Schipper, K.; Al-Jabri, H.M.S.J.; Wijffels, R.H.; Barbosa, M.J. Techno-economics of algae production in the Arabian Peninsula. Bioresour. Technol. 2021, 331, 125043. [Google Scholar] [CrossRef]
- Sampat, M.C.; Arun, D.M. Operational Strategies for Cost Effective Mass Cultivation of Halophilic Microalgal Strain Pseudanabaena limnetica in 1000 L Flat Panel Photobioreactor. J. Pet Environ. Biotechnol. 2018, 9, 2. [Google Scholar] [CrossRef]
- Endres, C.H.; Roth, A.; Brück, T.B. Modeling Microalgae Productivity in Industrial-Scale Vertical Flat Panel Photobioreactors. Env. Sci. Technol. 2018, 52, 5490–5498. [Google Scholar] [CrossRef]
- Béchet, Q.; Muñoz, R.; Shilton, A.; Guieysse, B. Outdoor cultivation of temperature-tolerant Chlorella sorokiniana in a column photobioreactor under low power-input. Biotechnol. Bioeng. 2013, 110, 118–126. [Google Scholar] [CrossRef]
- Borowitzka, M.A.; Moheimani, N.R. Algae for biofuels and energy. In Developments in Applied Phycology; Springer: Berlin/Heidelberg, Germany, 2013; pp. 1–288. [Google Scholar]
- Belay, A. Mass Culture of Spirulina Outdoors—The Earthrise Farms Experience. In Spirulina Platensis Arthrospira, 1st ed.; Vonshak, A., Ed.; CRC Press: Boca Raton, FL, USA, 1997; pp. 149–176. [Google Scholar]
- Windschitl, M.; Andre, T. Using Computer Simulations to Enhance Conceptual Change: The Roles of Constructivist Instruction and Student Epistemological Beliefs. J. Res. Sci. Teach. 1998, 35, 145–160. [Google Scholar] [CrossRef]
- Perkins, K.; Moore, E.; Podolefsky, N.; Lancaster, K.; Denison, C. Towards research-based strategies for using PhET simulations in middle school physical science classes. In AIP Conference Proceedings 1413; American Institute of PhysicsAIP: Omaha, NE, USA, 2012; p. 295. [Google Scholar]
- Luo, W.; Pelletier, J.; Duffin, K.; Ormand, C.; Hung, W.C.; Shernoff, D.J.; Zhai, X.; Iverson, E.; Whalley, K.; Gallaher, C.; et al. Advantages of Computer Simulation in Enhancing Students’ Learning About Landform Evolution: A Case Study Using the Grand Canyon. J. Geosci. Educ. 2018, 64, 60–73. [Google Scholar] [CrossRef]
- Bilotta, F.F.; Werner, S.M.; Bergese, S.D.; Rosa, G. Impact and implementation of simulation-based training for safety. Sci. World J. 2013, 2013, 652956. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.K. Commercial production of microalgae in the Asia-Pacific rim. J. Appl. Phycol. 1997, 9, 403–411. [Google Scholar] [CrossRef]
- Fuentes-Grünewald, C.; Garcés, E.; Alacid, E.; Rossi, S.; Camp, J. Biomass and Lipid Production of Dinoflagellates and Raphidophytes in Indoor and Outdoor Photobioreactors. Mar. Biotechnol. 2013, 15, 37–47. [Google Scholar] [CrossRef]
- Tredici, M.R.; Carlozzi, P.; Chini Zittelli, G.; Materassi, R. A vertical alveolar panel (VAP) for outdoor mass cultivation of microalgae and cyanobacteria. Bioresour. Technol. 1991, 38, 153–159. [Google Scholar] [CrossRef]
- Richmond, A.; Boussiba, S.; Vonshak, A.; Kopel, R. A new tubular reactor for mass production of microalgae outdoors. J. Appl. Phycol. 1993, 5, 327–332. [Google Scholar] [CrossRef]
- Carlozzi, P. Hydrodynamic aspects and Arthrospira growth in two outdoor tubular undulating row photobioreactors. Appl. Microbiol. Biotechnol. 2000, 54, 14–22. [Google Scholar] [CrossRef] [PubMed]
- Tredici, M.R.; Zlttelli, G.C. Efficiency of sunlight utilization: Tubular versus flat photobioreactors. Biotechnol. Bioeng. 1998, 57, 187–197. [Google Scholar] [CrossRef]
- Sato, T.; Usui, S.; Tsuchiya, Y.; Kondo, Y. Invention of outdoor closed type photobioreactor for microalgae. Energy Convers. Manag. 2006, 47, 791–799. [Google Scholar] [CrossRef]
- Berberoǧlu, H.; Pilon, L. Maximizing the solar to H2 energy conversion efficiency of outdoor photobioreactors using mixed cultures. Int. J. Hydrog. Energy 2010, 35, 500–510. [Google Scholar] [CrossRef]
- Sarker, N.K. Exploring the potential of wastewater reclamation by means of outdoor cultivation of microalgae in photobioreactors. Energy Ecol. Env. 2022, 7, 473–488. [Google Scholar] [CrossRef]
- Lee, Y.-K.; Low, C.-S. Productivity of outdoor algal cultures in enclosed tubular photobioreactor. Biotechnol. Bioeng. 1992, 40, 1119–1122. [Google Scholar] [CrossRef]
- Lee, Y.-K.; Low, C.-S. Effect of photobioreactor inclination on the biomass productivity of an outdoor algal culture. Biotechnol. Bioeng. 1991, 38, 995–1000. [Google Scholar] [CrossRef]
- Ramos de Ortega, A.; Roux, J.C. Production of Chlorella biomass in different types of flat bioreactors in temperate zones. Biomass 1986, 10, 141–156. [Google Scholar] [CrossRef]
- Ugwu, C.U.; Ogbonna, J.C.; Tanaka, H. Improvement of mass transfer characteristics and productivities of inclined tubular photobioreactors by installation of internal static mixers. Appl. Microbiol. Biotechnol. 2002, 58, 600–607. [Google Scholar] [CrossRef]
- Menegazzo, M.L.; Nascimento, V.M.; Hestekin, C.N.; Hestekin, J.A.; Fonseca, G.G. Evaluation of Chlorella sorokiniana cultivated in outdoor photobioreactors for biodiesel production. Biofuels 2022, 13, 483–488. [Google Scholar] [CrossRef]
- Zhou, X.; Xia, L.; Ge, H.; Zhang, D.; Hu, C. Feasibility of biodiesel production by microalgae Chlorella sp. (FACHB-1748) under outdoor conditions. Bioresour. Technol. 2013, 138, 131–135. [Google Scholar] [CrossRef]
- Lopes, A.P.; Santos, F.M.; Silva, T.F.; Vilar, V.J.; Pires, J.C. Outdoor cultivation of the microalga Chlorella vulgaris in a new photobioreactor configuration: The effect of ultraviolet and visible radiation. Energy 2020, 13, 1962. [Google Scholar] [CrossRef]
- Hindersin, S.; Leupold, M.; Kerner, M.; Hanelt, D. Irradiance optimization of outdoor microalgal cultures using solar tracked photobioreactors. Bioprocess Biosyst. Eng. 2013, 36, 345–355. [Google Scholar] [CrossRef]
- Fatemeh, L.; Mohsen, D. Effects of Environmental Factors on the Growth, Optical Density and Biomass of the Green Algae Chlorella Vulgaris in Outdoor Conditions. J. Appl. Sci. Environ. Manag. 2016, 20, 133. [Google Scholar] [CrossRef] [Green Version]
- Huo, S.; Wang, Z.; Zhu, S.; Shu, Q.; Zhu, L.; Qin, L.; Zhou, W.; Feng, P.; Zhu, F.; Yuan, Z.; et al. Biomass accumulation of Chlorella zofingiensis G1 cultures grown outdoors in photobioreactors. Front Energy Res. 2018, 6, 49. [Google Scholar] [CrossRef] [Green Version]
- Feng, P.; Deng, Z.; Hu, Z.; Fan, L. Lipid accumulation and growth of Chlorella zofingiensis in flat plate photobioreactors outdoors. Bioresour. Technol. 2011, 102, 10577–10584. [Google Scholar] [CrossRef]
- Cabanelas, I.T.; Slegers, P.M.; Böpple, H.; Kleinegris, D.M.; Wijffels, R.H.; Barbosa, M.J. Outdoor performance of Chlorococcum littorale at different locations. Algal. Res. 2017, 27, 55–64. [Google Scholar] [CrossRef]
- Norsker, N.H.; Cuaresma, M.; de Vree, J.; Ruiz-Domínguez, M.C.; García, M.C.; Uronen, P.; Barbosa, M.J.; Wijffels, R. Neochloris oleoabundans oil production in an outdoor tubular photobioreactor at pilot scale. J. Appl. Phycol. 2021, 33, 1327–1339. [Google Scholar] [CrossRef]
- Fuentes, J.L.; Montero, Z.; Cuaresma, M.; Ruiz-Domínguez, M.C.; Mogedas, B.; Nores, I.G.; González del Valle, M.; Vílchez, C. Outdoor Large-Scale Cultivation of the Acidophilic Microalga Coccomyxa onubensis in a Vertical Close Photobioreactor for Lutein Production. Processes 2020, 8, 324. [Google Scholar] [CrossRef] [Green Version]
- Xia, L.; Song, S.; He, Q.; Yang, H.; Hu, C. Selection of microalgae for biodiesel production in a scalable outdoor photobioreactor in north China. Bioresour. Technol. 2014, 174, 274–280. [Google Scholar] [CrossRef] [PubMed]
- Greene, J.M.; Quiroz, D.; Compton, S.; Lammers, P.J.; Quinn, J.C. A validated thermal and biological model for predicting algal productivity in large scale outdoor cultivation systems. Algal Res. 2021, 54, 102224. [Google Scholar] [CrossRef]
- López, M.G.; Sánchez, E.D.; López, J.C.; Fernández, F.A.; Sevilla, J.F.; Rivas, J.; Guerrero, M.G.; Grima, E.M. Comparative analysis of the outdoor culture of Haematococcus pluvialis in tubular and bubble column photobioreactors. J. Biotechnol. 2006, 123, 329–342. [Google Scholar] [CrossRef] [PubMed]
- López-Rosales, L.; Sánchez-Mirón, A.; García-Camacho, F.; Place, A.R.; Chisti, Y.; Molina-Grima, E. Pilot-scale outdoor photobioreactor culture of the marine dinoflagellate Karlodinium veneficum: Production of a karlotoxins-rich extract. Bioresour. Technol. 2018, 253, 94–104. [Google Scholar] [CrossRef] [Green Version]
- Jimenez, R.; Markou, G.; Tayibi, S.; Barakat, A.; Chapsal, C.; Monlau, F. Production of Microalgal Slow-Release Fertilizer by Valorizing Liquid Agricultural Digestate: Growth Experiments with Tomatoes. Appl. Sci. 2020, 10, 3890. [Google Scholar] [CrossRef]
- Bosma, R.; van Zessen, E.; Reith, J.H.; Tramper, J.; Wijffels, R.H. Prediction of volumetric productivity of an outdoor photobioreactor. Biotechnol. Bioeng. 2007, 97, 1108–1120. [Google Scholar] [CrossRef]
- Del Campo, J.A.; Rodríguez, H.; Moreno, J.; Vargas, M.A.; Rivas, J.; Guerrero, M.G. Lutein production by Muriellopsis sp. in an outdoor tubular photobioreactor. J. Biotechnol. 2001, 85, 289–295. [Google Scholar] [CrossRef]
- Dogaris, I.; Welch, M.; Meiser, A.; Walmsley, L.; Philippidis, G. A novel horizontal photobioreactor for high-density cultivation of microalgae. Bioresour. Technol. 2015, 198, 316–324. [Google Scholar] [CrossRef]
- Chen, C.Y.; Nagarajan, D.; Cheah, W.Y. Eicosapentaenoic acid production from Nannochloropsis oceanica CY2 using deep sea water in outdoor plastic-bag type photobioreactors. Bioresour. Technol. 2018, 253, 1–7. [Google Scholar] [CrossRef]
- Norsker, N.H.; Michiels, M.; Slegers, P.M.; Swinkels, G.L.; Barbosa, M.J.; Wijffels, R.H. Productivity of Nannochloropsis oceanica in an industrial closely spaced flat panel photobioreactor. Algal. Res. 2019, 43, 101632. [Google Scholar] [CrossRef]
- Carneiro, M.; Cicchi, B.; Maia, I.B.; Pereira, H.; Zittelli, G.C.; Varela, J.; Malcata, F.X.; Torzillo, G. Effect of temperature on growth, photosynthesis and biochemical composition of Nannochloropsis oceanica, grown outdoors in tubular photobioreactors. Algal Res. 2020, 49, 101923. [Google Scholar] [CrossRef]
- Guerra, I.; Pereira, H.; Costa, M.; Silva, J.T.; Santos, T.; Varela, J.; Mateus, M.; Silva, J. Operation Regimes: A Comparison Based on Nannochloropsis oceanica Biomass and Lipid Productivity. Energy 2021, 14, 1542. [Google Scholar] [CrossRef]
- Quinn, J.C.; Yates, T.; Douglas, N.; Weyer, K.; Butler, J.; Bradley, T.H.; Lammers, P.J. Nannochloropsis production metrics in a scalable outdoor photobioreactor for commercial applications. Bioresour. Technol. 2012, 117, 164–171. [Google Scholar] [CrossRef]
- Olofsson, M.; Lamela, T.; Nilsson, E.; Bergé, J.P.; Del Pino, V.; Uronen, P.; Legrand, C. Seasonal variation of lipids and fatty acids of the microalgae Nannochloropsis oculata grown in outdoor large-scale photobioreactors. Energy 2012, 5, 1577–1592. [Google Scholar] [CrossRef] [Green Version]
- Cheregi, O.; Engelbrektsson, J.; Andersson, M.X.; Strömberg, N.; Ekendahl, S.; Godhe, A.; Spetea, C. Marine microalgae for outdoor biomass production—A laboratory study simulating seasonal light and temperature for the west coast of Sweden. Physiol. Plant 2021, 173, 543–554. [Google Scholar] [CrossRef]
- De Vree, J.H.; Bosma, R.; Janssen, M.; Barbosa, M.J.; Wijffels, R.H. Comparison of four outdoor pilot-scale photobioreactors. Biotechnol. Biofuels 2015, 8, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Rodolfi, L.; Chini Zittelli, G.; Bassi, N.; Padovani, G.; Biondi, N.; Bonini, G.; Tredici, M.R. Microalgae for oil: Strain selection, induction of lipid synthesis and outdoor mass cultivation in a low-cost photobioreactor. Biotechnol. Bioeng. 2009, 102, 100–112. [Google Scholar] [CrossRef]
- Zittelli, G.C.; Lavista, F.; Bastianini, A.; Rodolfi, L.; Vincenzini, M.; Tredici, M.R. Production of eicosapentaenoic acid by Nannochloropsis sp. cultures in outdoor tubular photobioreactors. J. Biotechnol. 1999, 70, 299–312. [Google Scholar] [CrossRef]
- de Vree, J.H.; Bosma, R.; Wieggers, R.; Gegic, S.; Janssen, M.; Barbosa, M.J.; Wijffels, R.H. Turbidostat operation of outdoor pilot-scale photobioreactors. Algal. Res. 2016, 18, 198–208. [Google Scholar] [CrossRef]
- Fernández, F.A.; Camacho, F.G.; Pérez, J.S.; Sevilla, J.F.; Grima, E.M. A model for light distribution and average solar irradiance inside outdoor tubular photobioreactors for the microalgal mass culture. Biotechnol. Bioeng. 1997, 55, 701–714. [Google Scholar] [CrossRef]
- Fernández, F.A.; Sevilla, J.F.; Pérez, J.S.; Grima, E.M.; Chisti, Y. Airlift-driven external-loop tubular photobioreactors for outdoor production of microalgae: Assessment of design and performance. Chem. Eng. Sci. 2001, 56, 2721–2732. [Google Scholar] [CrossRef] [Green Version]
- Hall, D.O.; Acién Fernández, F.G.; Guerrero, E.C.; Rao, K.K.; Grima, E.M. Outdoor helical tubular photobioreactors for microalgal production: Modeling of fluid-dynamics and mass transfer and assessment of biomass productivity. Biotechnol. Bioeng. 2003, 82, 62–73. [Google Scholar] [CrossRef] [PubMed]
- Camacho, F.G.; Gómez, A.C.; Fernández, F.A.; Sevilla, J.F.; Grima, E.M. Use of concentric-tube airlift photobioreactors for microalgal outdoor mass cultures. Enzym. Microb. Technol. 1999, 24, 164–172. [Google Scholar] [CrossRef]
- Fernández, F.A.; Hall, D.O.; Guerrero, E.C.; Rao, K.K.; Grima, E.M. Outdoor production of Phaeodactylum tricornutum biomass in a helical reactor. J. Biotechnol. 2003, 103, 137–152. [Google Scholar] [CrossRef]
- Gao, F.; Sá, M.; Teles, I.; Wijffels, R.H.; Barbosa, M.J. Production and monitoring of biomass and fucoxanthin with brown microalgae under outdoor conditions. Biotechnol. Bioeng. 2021, 118, 1355–1365. [Google Scholar] [CrossRef]
- Pereira, H.; Sá, M.; Maia, I.; Rodrigues, A.; Teles, I.; Wijffels, R.H.; Navalho, J.; Barbosa, M. Fucoxanthin production from Tisochrysis lutea and Phaeodactylum tricornutum at industrial scale. Algal. Res. 2021, 56, 102322. [Google Scholar] [CrossRef]
- Quelhas, P.M.; Trovão, M.; Silva, J.T.; Machado, A.; Santos, T.; Pereira, H.; Varela, J.; Simões, M.; Silva, J.L. Industrial production of Phaeodactylum tricornutum for CO2 mitigation: Biomass productivity and photosynthetic efficiency using photobioreactors of different volumes. J. Appl. Phycol. 2019, 31, 2187–2196. [Google Scholar] [CrossRef]
- Steinrücken, P.; Prestegard, S.K.; de Vree, J.H.; Storesund, J.E.; Pree, B.; Mjøs, S.A.; Erga, S.R. Comparing EPA production and fatty acid profiles of three Phaeodactylum tricornutum strains under western Norwegian climate conditions. Algal. Res. 2018, 30, 11–22. [Google Scholar] [CrossRef]
- Rodolfi, L.; Biondi, N.; Guccione, A.; Bassi, N.; D'Ottavio, M.; Arganaraz, G.; Tredici, M.R. Oil and eicosapentaenoic acid production by the diatom Phaeodactylum tricornutum cultivated outdoors in Green Wall Panel (GWP®) reactors. Biotechnol. Bioeng. 2017, 114, 2204–2210. [Google Scholar] [CrossRef] [Green Version]
- Buono, S.; Colucci, A.; Angelini, A.; Langellotti, A.L.; Massa, M.; Martello, A.; Fogliano, V.; Dibenedetto, A. Productivity and biochemical composition of Tetradesmus obliquus and Phaeodactylum tricornutum: Effects of different cultivation approaches. J. Appl. Phycol. 2016, 28, 3179–3192. [Google Scholar] [CrossRef]
- Mirón, A.S.; Garcıa, M.C.; Gómez, A.C.; Camacho, F.G.; Grima, E.M.; Chisti, Y. Shear stress tolerance and biochemical characterization of Phaeodactylum tricornutum in quasi steady-state continuous culture in outdoor photobioreactors. Biochem. Eng. J. 2003, 16, 287–297. [Google Scholar] [CrossRef]
- Silva Benavides, A.M.; Torzillo, G.; Kopecký, J.; Masojídek, J. Productivity and biochemical composition of Phaeodactylum tricornutum (Bacillariophyceae) cultures grown outdoors in tubular photobioreactors and open ponds. Biomass Bioenergy 2013, 54, 115–122. [Google Scholar] [CrossRef]
- Torzillo, G.; Faraloni, C.; Silva, A.M.; Kopecký, J.; Pilný, J.; Masojídek, J. Photoacclimation of Phaeodactylum tricornutum (Bacillariophyceae) cultures grown outdoors in photobioreactors and open ponds. Eur. J. Phycol. 2012, 47, 169–181. [Google Scholar] [CrossRef]
- Rubio, F.C.; Fernández, F.A.; Pérez, J.S.; Camacho, F.G.; Grima, E.M. Prediction of dissolved oxygen and carbon dioxide concentration profiles in tubular photobioreactors for microalgal culture. Biotechnol. Bioeng. 1999, 62, 71–86. [Google Scholar] [CrossRef]
- Androga, D.D.; Uyar, B.; Koku, H.; Eroglu, I. Dynamic modeling of temperature change in outdoor operated tubular photobioreactors. Bioprocess Biosyst. Eng. 2017, 40, 1017–1031. [Google Scholar] [CrossRef]
- Carlozzi, P.; Sacchi, A. Biomass production and studies on Rhodopseudomonas palustris grown in an outdoor, temperature controlled, underwater tubular photobioreactor. J. Biotechnol. 2001, 88, 239–249. [Google Scholar] [CrossRef]
- Carlozzi, P.; Pushparaj, B.; Degl’Innocenti, A.; Capperucci, A. Growth characteristics of Rhodopseudomonas palustris cultured outdoors, in an underwater tubular photobioreactor, and investigation on photosynthetic efficiency. Appl. Microbiol. Biotechnol. 2006, 73, 789–795. [Google Scholar] [CrossRef]
- Hulatt, C.J.; Thomas, D.N. Energy efficiency of an outdoor microalgal photobioreactor sited at mid-temperate latitude. Bioresour. Technol. 2011, 102, 6687–6695. [Google Scholar] [CrossRef]
- Torzillo, G.; Pushparaj, B.; Bocci, F.; Balloni, W.; Materassi, R.; Florenzano, G. Production of Spirulina biomass in closed photobioreactors. Biomass 1986, 11, 61–74. [Google Scholar] [CrossRef]
- Masojídek, J.; Papáček, Š.; Sergejevova, M.; Jirka, V.; Červený, J.; Kunc, J.; Korečko, J.; Verbovikova, O.; Kopecký, J.; Štys, D.; et al. A closed solar photobioreactor for cultivation of microalgae under supra-high irradiance: Basic design and performance. J. Appl. Phycol. 2003, 15, 239–248. [Google Scholar] [CrossRef]
- Torzillo, G.; Sacchi, A.; Materassi, R. Temperature as an important factor affecting productivity and night biomass loss in Spirulina platensis grown outdoors in tubular photobioreactors. Bioresour. Technol. 1991, 38, 95–100. [Google Scholar] [CrossRef]
- Torzillo, G.; Sacchi, A.; Materassi, R.; Richmond, A. Effect of temperature on yield and night biomass loss in Spirulina platensis grown outdoors in tubular photobioreactors. J. Appl. Phycol. 1991, 3, 103–109. [Google Scholar] [CrossRef]
- Vonshak, A.; Torzillo, G.; Tomaseli, L. Use of chlorophyll fluorescence to estimate the effect of photoinhibition in outdoor cultures of Spirulina platensis. J. Appl. Phycol. 1994, 6, 31–34. [Google Scholar] [CrossRef]
- Carlozzi, P.; Torzillo, G. Productivity of Spirulina in a strongly curved outdoor tubular photobioreactor. Appl. Microbiol. Biotechnol. 1996, 45, 18–23. [Google Scholar] [CrossRef]
- Prakash, J.; Torzillo, G.; Pushparaj, B.; Carlozzi, P.; Materassi, R. Transient analysis and performance studies of two tubular photobioreactors for outdoor culture of Spirulina. Int. J. Energy Res. 1995, 19, 479–492. [Google Scholar] [CrossRef]
- Vonshak, A.; Torzillo, G.; Accolla, P.; Tomaselli, L. Light and oxygen stress in Spirulina platensis (cyanobacteria) grown outdoors in tubular reactors. Physiol. Plant. 1996, 97, 175–179. [Google Scholar] [CrossRef]
- Torzillo, G.; Carlozzi, P.; Pushparaj, B.; Montaini, E.; Materassi, R. A two-plane tubular photobioreactor for outdoor culture of Spirulina. Biotechnol. Bioeng. 1993, 42, 891–898. [Google Scholar] [CrossRef]
- Zhang, K.; Miyachi, S.; Kurano, N. Evaluation of a vertical flat-plate photobioreactor for outdoor biomass production and carbon dioxide bio-fixation: Effects of reactor dimensions, irradiation and cell concentration on the biomass productivity and irradiation utilization efficiency. Appl. Microbiol. Biotechnol. 2001, 55, 428–433. [Google Scholar] [CrossRef]
- Chini Zittelli, G.; Rodolfi, L.; Biondi, N.; Tredici, M.R. Productivity and photosynthetic efficiency of outdoor cultures of Tetraselmis suecica in annular columns. Aquaculture 2006, 261, 932–943. [Google Scholar] [CrossRef]
- Tredici, M.R.; Bassi, N.; Prussi, M.; Biondi, N.; Rodolfi, L.; Zittelli, G.C.; Sampietro, G. Energy balance of algal biomass production in a 1-ha “Green Wall Panel” plant: How to produce algal biomass in a closed reactor achieving a high Net Energy Ratio. Appl. Energy 2015, 154, 1103–1111. [Google Scholar] [CrossRef] [Green Version]
- Pereira, H.; Páramo, J.; Silva, J.; Marques, A.; Barros, A.; Maurício, D.; Santos, T.; Schulze, P.; Barros, R.; Gouveia, L.; et al. Scale-up and large-scale production of Tetraselmis sp. CTP4 (Chlorophyta) for CO2 mitigation: From an agar plate to 100-m3 industrial photobioreactors. Sci. Rep. 2018, 8, 5112. [Google Scholar] [CrossRef]
- Narala, R.R.; Garg, S.; Sharma, K.K.; Thomas-Hall, S.R.; Deme, M.; Li, Y.; Schenk, P.M. Comparison of microalgae cultivation in photobioreactor, open raceway pond, and a two-stage hybrid system. Front. Energy Res. 2016, 4, 29. [Google Scholar] [CrossRef] [Green Version]
- Kusumaningtyas, P.; Nurbaiti, S.; Suantika, G.; Amran, M.B.; Nurachman, Z. Enhanced Oil Production by the Tropical Marine Diatom Thalassiosira Sp. Cultivated in Outdoor Photobioreactors. Appl. Biochem. Biotechnol. 2017, 182, 1605–1618. [Google Scholar] [CrossRef]
- Guccione, A.; Biondi, N.; Sampietro, G.; Rodolfi, L.; Bassi, N.; Tredici, M.R. Chlorella for protein and biofuels: From strain selection to outdoor cultivation in a Green Wall Panel photobioreactor. Biotechnol. Biofuels 2014, 7, 84. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.; Miyachi, S.; Kurano, N. Photosynthetic performance of a cyanobacterium in a vertical flat-plate photobioreactor for outdoor microalgal production and fixation of CO2. Biotechnol. Lett. 2001, 23, 21–26. [Google Scholar] [CrossRef]
- Gutiérrez, J.; Porta-Gándara, M.A.; Fernández, J.L. Passive temperature solar control of an outdoor photobioreactor. Renew. Energy 2008, 33, 1892–1903. [Google Scholar] [CrossRef]
- Sebök, S.; Herppich, W.B.; Hanelt, D. Outdoor cultivation of Ulva lactuca in a recently developed ring-shaped photobioreactor: Effects of elevated CO2 concentration on growth and photosynthetic performance. Bot. Mar. 2019, 62, 179–190. [Google Scholar] [CrossRef]
- Petkov, G.D. Absorber tower as a photobioreactor for microalgae. Russ. J. Plant Physiol. 2000, 47, 786–788. [Google Scholar] [CrossRef]
- Huang, J.; Hankamer, B.; Yarnold, J. Design scenarios of outdoor arrayed cylindrical photobioreactors for microalgae cultivation considering solar radiation and temperature. Algal. Res. 2019, 41, 101515. [Google Scholar] [CrossRef] [Green Version]
- Ong, S.C.; Kao, C.Y.; Chiu, S.Y.; Tsai, M.T.; Lin, C.S. Characterization of the thermal-tolerant mutants of Chlorella sp. with high growth rate and application in outdoor photobioreactor cultivation. Bioresour. Technol. 2010, 101, 2880–2883. [Google Scholar] [CrossRef]
- Hejazi, M.A.; Barzegari, A.; Gharajeh, N.H.; Hejazi, M.S. Introduction of a novel 18S rDNA gene arrangement along with distinct ITS region in the saline water microalga Dunaliella. Saline Syst. 2010, 6, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, S.; Xiong, J.; Yu, Y. Taxonomic Resolutions Based on 18S rRNA Genes: A Case Study of Subclass Copepoda. PLoS ONE 2015, 10, e0131498. [Google Scholar] [CrossRef] [PubMed]
- di Caprio, F.; Altimari, P.; Pagnanelli, F. New strategies enhancing feasibility of microalgal cultivations. In Studies in Surface Science and Catalysis; Elsevier: Amsterdam, The Netherlands, 2020; pp. 287–316. [Google Scholar]
- Fernández, F.A.; Camacho, F.G.; Pérez, J.S.; Sevilla, J.F.; Grima, E.M. Modeling of biomass productivity in tubular photobioreactors for microalgal cultures: Effects of dilution rate, tube diameter, and solar irradiance. Biotechnol. Bioeng. 2000, 58, 605–616. [Google Scholar] [CrossRef]
- Doucha, J.; Livansky, K. Process of Outdoor Thin-Layer Cultivation of Microalgae And Blue-Green Algae and Bioreactor for Performing the Process. U.S. Patent US5981271A, 11 September 1997. [Google Scholar]
- Eckerle, M.W.; Chalberg, T.W.; Hackworth, C.A.; Fertik, M.B. Photobioreactor Systems and Methods for Growing Organisms. U.S. Patent US20090203067A1, 18 September 2009. [Google Scholar]
- Laws, E.A.; Taguchi, S.; Hirata, J.; Pang, L. Optimization of microalgal production in a shallow outdoor flume. Biotechnol. Bioeng. 1988, 32, 140–147. [Google Scholar] [CrossRef]
- Terry, K.L. Photosynthesis in modulated light: Quantitative dependence of photosynthetic enhancement on flashing rate. Biotechnol. Bioeng. 1986, 28, 988–995. [Google Scholar] [CrossRef]
- Fuentes, J.L.; Garbayo, I.; Cuaresma, M.; Montero, Z.; González-del-Valle, M.; Vílchez, C. Impact of Microalgae-Bacteria Interactions on the Production of Algal Biomass and Associated Compounds. Mar. Drugs 2016, 14, 100. [Google Scholar] [CrossRef] [Green Version]
- Luo, H.; Moran, M.A. Evolutionary Ecology of the Marine Roseobacter Clade. Microbiol. Mol. Biol. Rev. 2014, 78, 573–587. [Google Scholar] [CrossRef] [Green Version]
- Afi, L.; Metzger, P.; Largeau, C.; Connan, J.; Berkaloff, C.; Rousseau, B. Bacterial degradation of green microalgae: Incubation of Chlorella emersonii and Chlorella vulgaris with Pseudomonas oleovorans and Flavobacterium aquatile. Org. Geochem. 1996, 25, 117–130. [Google Scholar] [CrossRef]
- Wang, X.; Li, Z.; Su, J.; Tian, Y.; Ning, X.; Hong, H.; Zheng, T. Lysis of a red-tide causing alga, Alexandrium tamarense, caused by bacteria from its phycosphere. Biol. Control. 2010, 52, 123–130. [Google Scholar] [CrossRef]
- Ramanan, R.; Kim, B.H.; Cho, D.H.; Oh, H.M.; Kim, H.S. Algae–bacteria interactions: Evolution, ecology and emerging applications. Biotechnol. Adv. 2016, 34, 14–29. [Google Scholar] [CrossRef] [Green Version]
- Hoekema, S.; Bijmans, M.; Janssen, M.; Tramper, J.; Wijffels, R.H. A pneumatically agitated flat-panel photobioreactor with gas re-circulation: Anaerobic photoheterotrophic cultivation of a purple non-sulfur bacterium. Int. J. Hydrogen Energy 2002, 27, 1331–1338. [Google Scholar] [CrossRef]
- Tredici, M.R. Photobiology of microalgae mass cultures: Understanding the tools for the next green revolution. Biofuels 2014, 1, 143–162. [Google Scholar] [CrossRef]
- Wang, B.; Lan, C.Q.; Horsman, M. Closed photobioreactors for production of microalgal biomasses. Biotechnol. Adv. 2012, 30, 904–912. [Google Scholar] [CrossRef]
- Carvalho, A.P.; Meireles, L.A.; Malcata, F.X. Microalgal Reactors: A Review of Enclosed System Designs and Performances. Biotechnol. Prog. 2006, 22, 1490–1506. [Google Scholar] [CrossRef] [PubMed]
- Endres, C.H.; Roth, A.; Brück, T.B. Thermal Reactor Model for Large-Scale Algae Cultivation in Vertical Flat Panel Photobioreactors. Env. Sci. Technol. 2016, 50, 3920–3927. [Google Scholar] [CrossRef] [PubMed]
- Nwoba, E.G.; Parlevliet, D.A.; Laird, D.W.; Alameh, K.; Moheimani, N.R. Light management technologies for increasing algal photobioreactor efficiency. Algal. Res. 2019, 39, 101433. [Google Scholar] [CrossRef]
- Sarker, N.K.; Sarkar, S. A comparative study on cost analysis, efficiency, and process mechanism of effluent treatment plants in Bangladesh. Environ. Qual. Manag. 2018, 27, 127–133. [Google Scholar] [CrossRef]
- Myint, M.T.; Ghassemi, A.; Nirmalakhandan, N. A generic stoichiometric equation for microalgae–microorganism nexus by using clarified domestic wastewater as growth medium. New Pub. Balaban 2013, 51, 6632–6640. [Google Scholar] [CrossRef]
- Liu, L.; Pohnert, G.; Wei, D. Extracellular Metabolites from Industrial Microalgae and Their Biotechnological Potential. Mar. Drugs 2016, 14, 191. [Google Scholar] [CrossRef]
- Eckerle, M.W. Patent Application Publication. U.S. Patent US 2009/0220638A1, 3 September 2009. [Google Scholar]
- Trosch, W.; Schmid-Staiger, U.; Zastrow, A.; Retze, A.; Brucker, F.; Fraunhofer Gesellschaft zur Forderung der Angewandten Forschung eV. Photobioreactor With Improved Supply Of Light By Surface Enlargement, Wavelength Shifter Bars Or Light Transport. U.S. Patent US6509188B1, 21 January 2003. [Google Scholar]
- Tredici, M.; Rodolfi, L. Reactor for Industrial Culture of Photosynthetic Micro-Organisms. U.S. Patent EP1599570A2, 24 February 2004. [Google Scholar]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sarker, N.K.; Kaparaju, P. A Critical Review on the Status and Progress of Microalgae Cultivation in Outdoor Photobioreactors Conducted over 35 Years (1986–2021). Energies 2023, 16, 3105. https://doi.org/10.3390/en16073105
Sarker NK, Kaparaju P. A Critical Review on the Status and Progress of Microalgae Cultivation in Outdoor Photobioreactors Conducted over 35 Years (1986–2021). Energies. 2023; 16(7):3105. https://doi.org/10.3390/en16073105
Chicago/Turabian StyleSarker, Nilay Kumar, and Prasad Kaparaju. 2023. "A Critical Review on the Status and Progress of Microalgae Cultivation in Outdoor Photobioreactors Conducted over 35 Years (1986–2021)" Energies 16, no. 7: 3105. https://doi.org/10.3390/en16073105