
Simultaneous Hydrogen and Ethanol Production from Crude Glycerol by a Microbial Consortium Using Fed-Batch Fermentation


Abstract
:1. Introduction
2. Materials and Methods
2.1. Media Composition and Substrate
2.2. Inoculum Preparation and Growth Conditions
2.3. Fed-Batch Fermentations
2.4. Microbial Growth
2.5. Gas and Metabolite Analysis
2.6. Analysis of Microbial Community Structure
2.6.1. Isolation and Characterization of Main Microbial Communities of GCL
2.6.2. DNA Extraction and PCR Amplification
2.6.3. Denaturing Gradient Gel Electrophoresis (DGGE)
3. Results and Discussion
3.1. Fed-Batch Experiments for Hydrogen and Ethanol Production
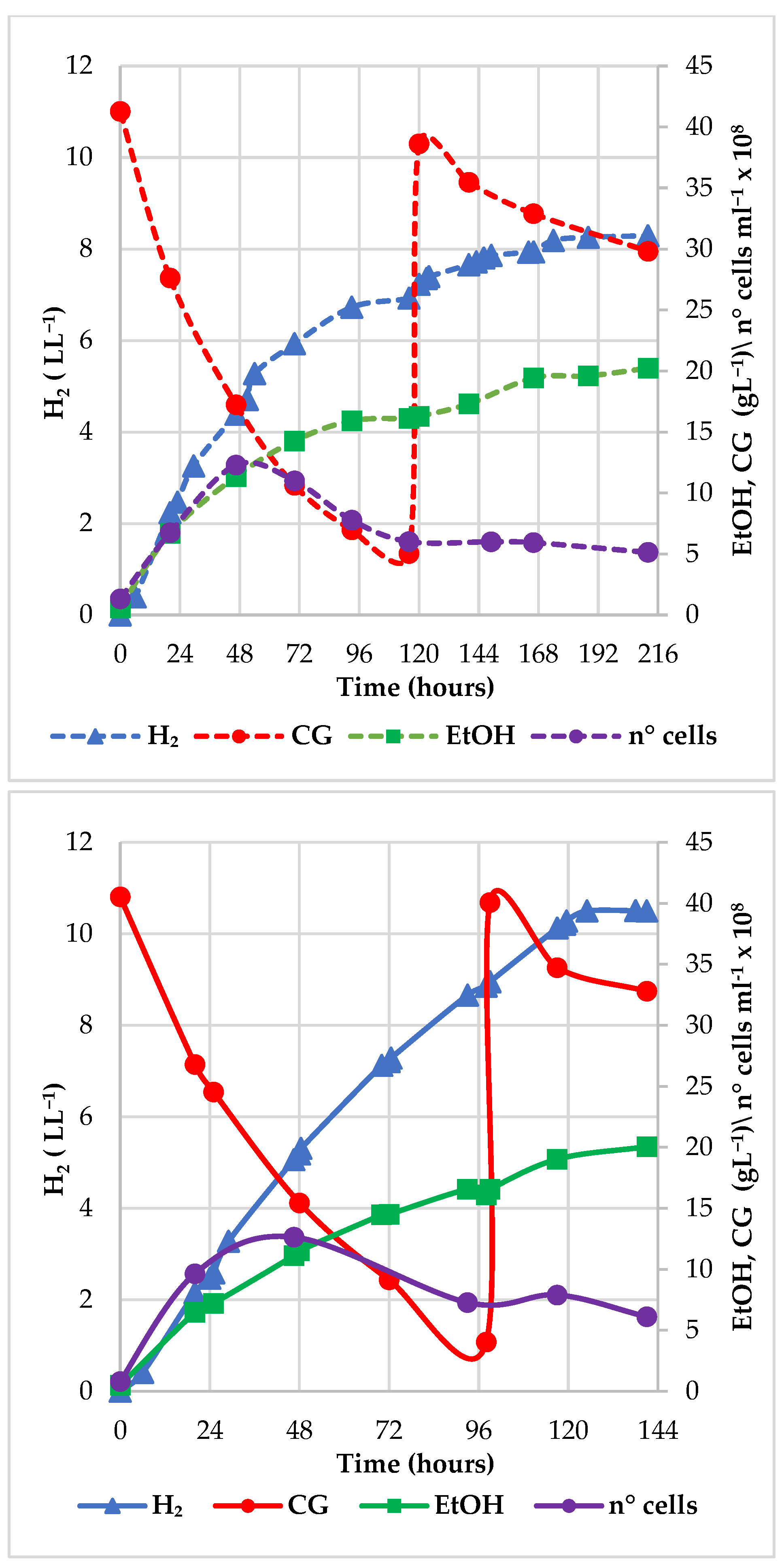
3.1.1. Increasing the CG Concentration: Performance of the Fed-Batch Processes
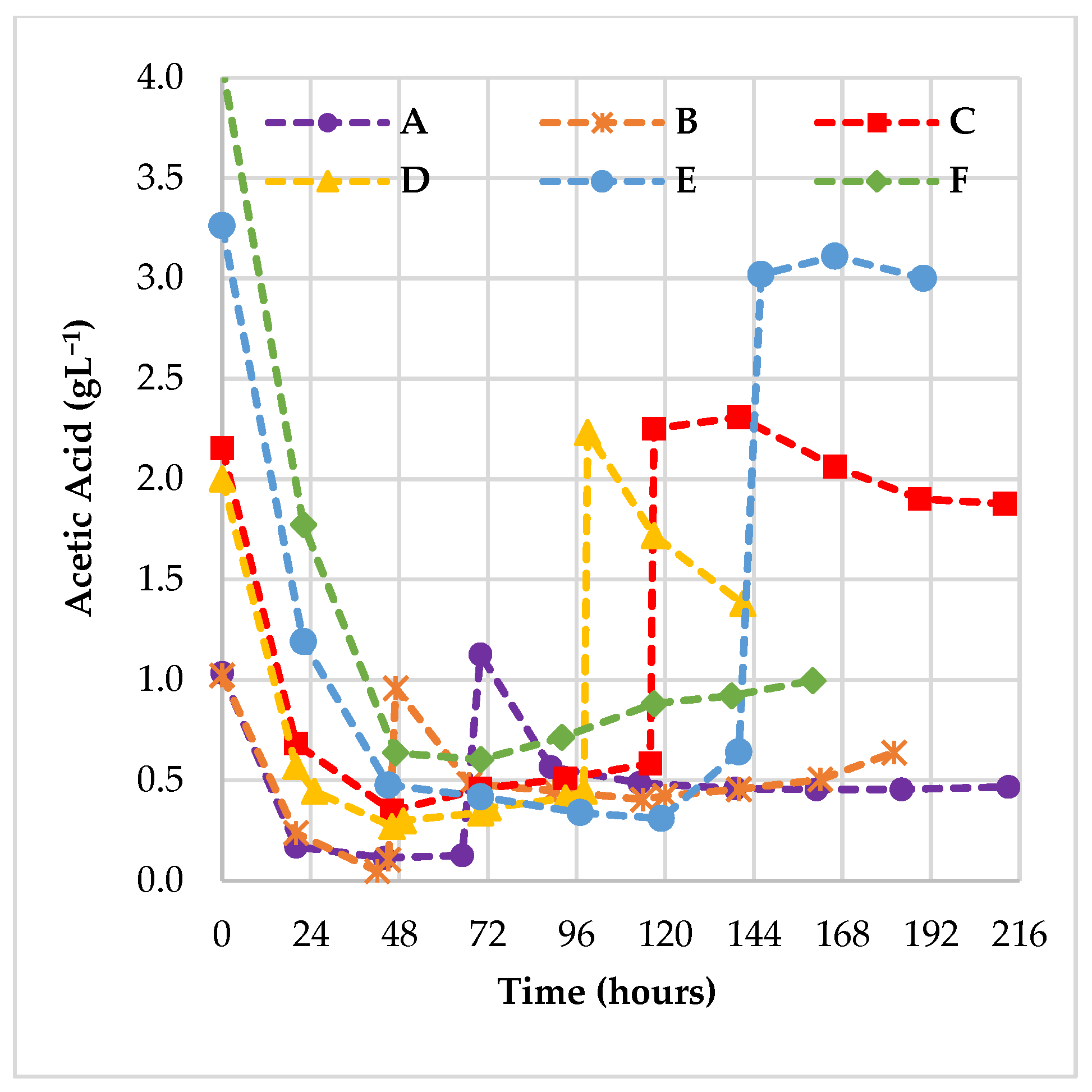
3.1.2. Role of Acetic Acid as the Main Impurities of CG during the Fed-Batch Fermentations
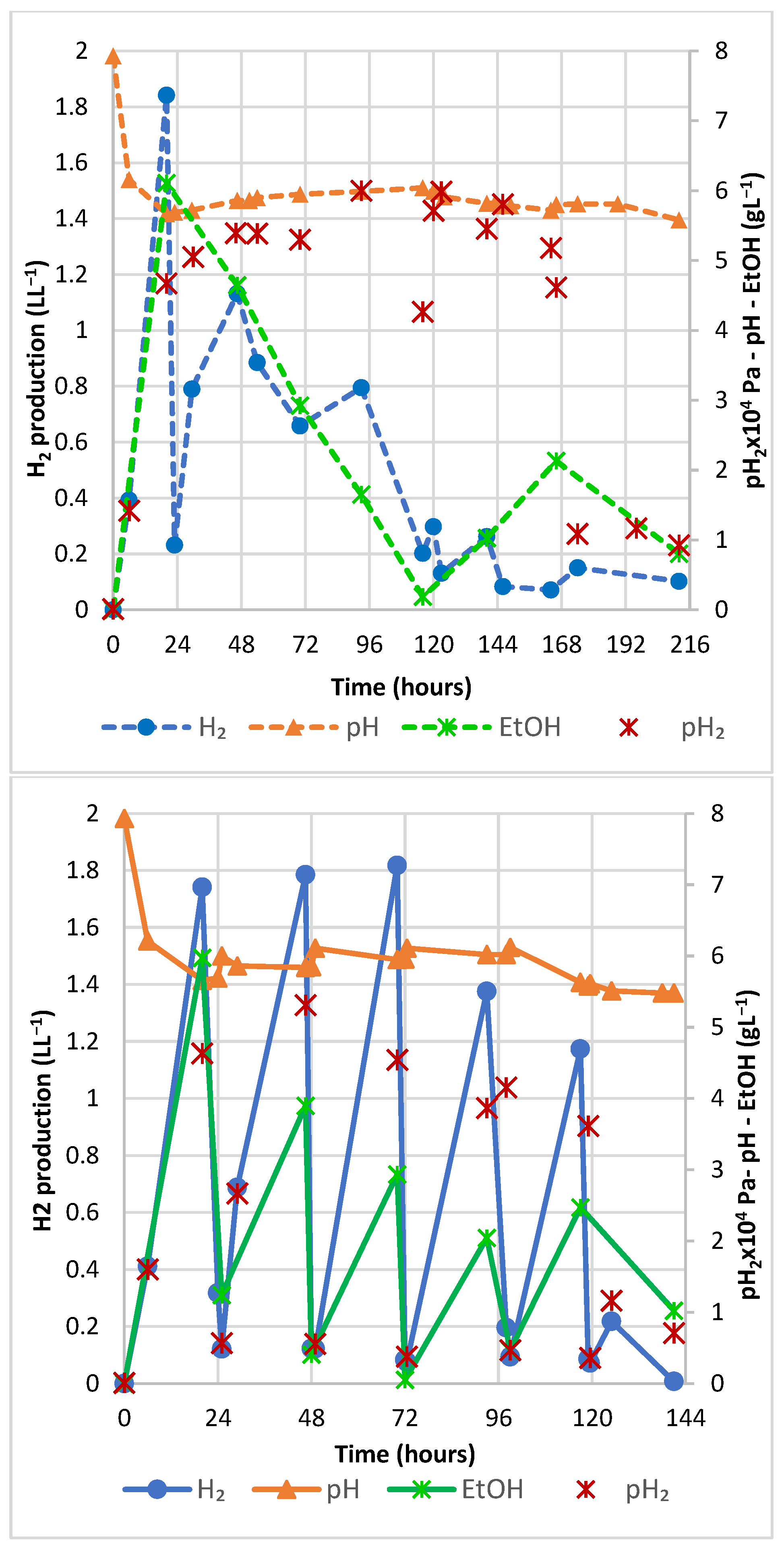
3.1.3. Effects of N2 Sparging on Instantaneous H2 and EtOH Production over Time
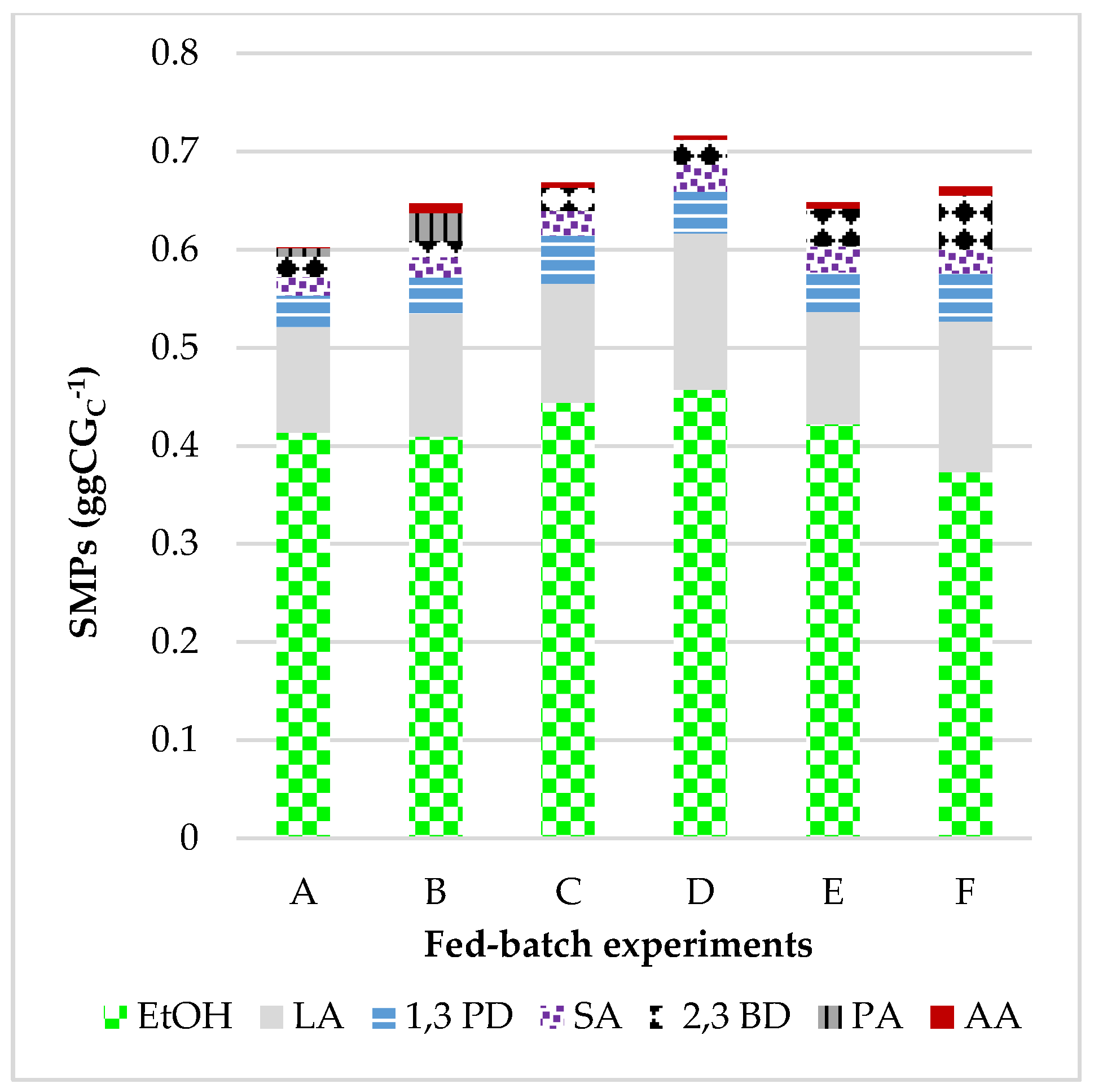
3.1.4. Increasing the CG Concentration: Metabolic Pathways and Products
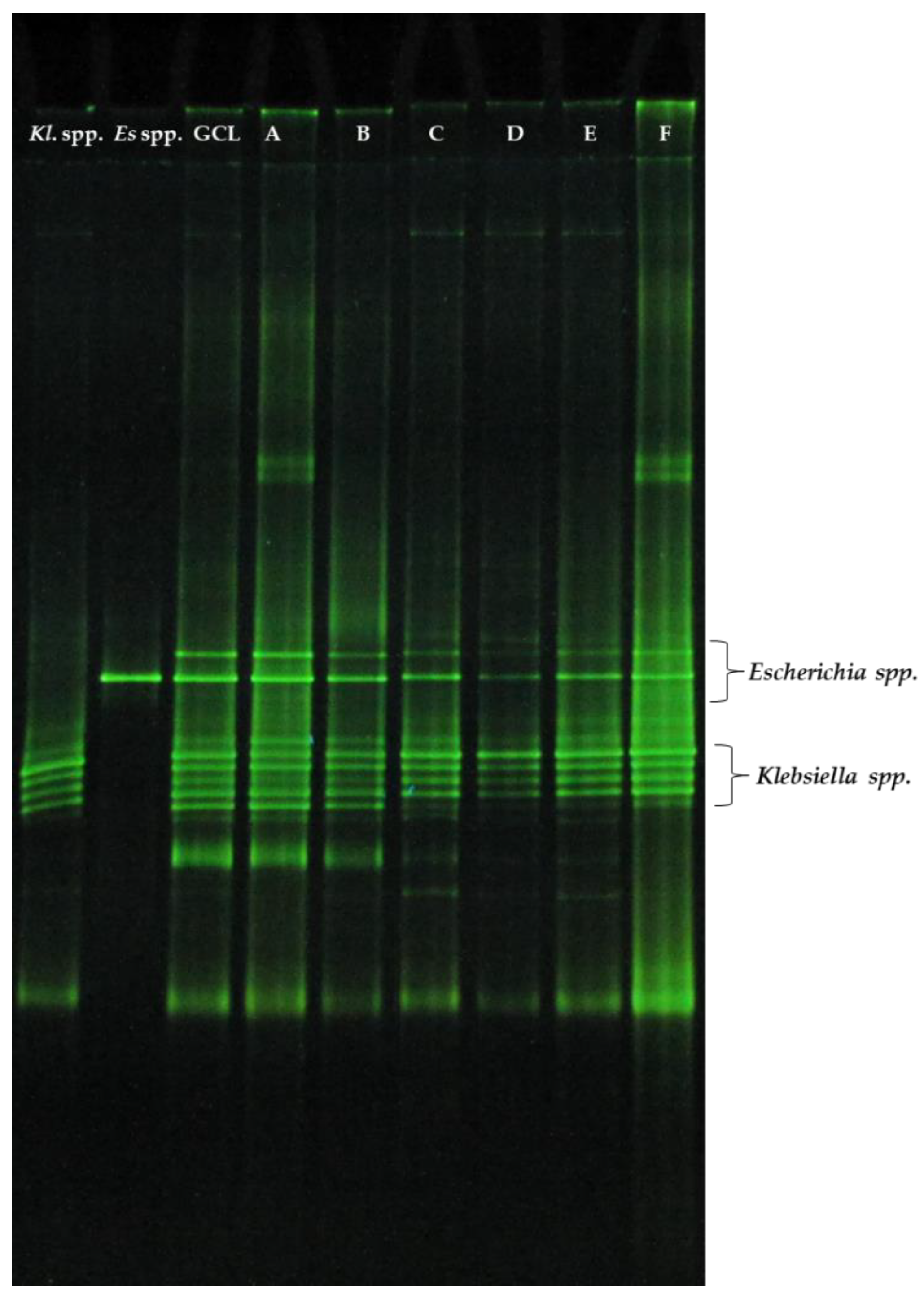
3.2. DGGE Profile of GCL Microbial Community at the End of Fed-Batch Fermentations
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- European Commission. Fit for 55: Delivering the EU’s 2030 Climate Target on the Way to Climate Neutrality; COM(2021) 550 Final; European Commission: Brussels, Belgium, 2021. [Google Scholar]
- Hosseini, S.E.; Wahid, M.A. Hydrogen production from renewable and sustainable energy resources: Promising green energy carrier for clean development. Renew. Sustain. Energy Rev. 2016, 57, 850–866. [Google Scholar] [CrossRef]
- Lepage, T.; Kammoun, M.; Schmetz, Q.; Richel, A. Biomass-to-hydrogen: A review of main routes production, processes evaluation and techno-economical assessment. Biomass Bioenergy 2021, 144, 105920. [Google Scholar] [CrossRef]
- Ma, Y.; Wang, X.R.; Li, T.; Zhang, J.; Gao, J.; Sun, Z.Y. Hydrogen and ethanol: Production, storage, and transportation. Int. J. Hydrog. Energy 2021, 46, 27330–27348. [Google Scholar] [CrossRef]
- Soo, C.S.; Yap, W.S.; Hon, W.M.; Phang, L.Y. Mini review: Hydrogen and ethanol co-production from waste materials via microbial fermentation. World J. Microbiol. Biotechnol. 2015, 31, 1475–1488. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhong, B.; Lawal, A. Recovery and utilization of crude glycerol, a biodiesel byproduct. RSC Adv. 2022, 12, 27997–28008. [Google Scholar] [CrossRef] [PubMed]
- Yazdani, S.S.; Gonzalez, R. Anaerobic fermentation of glycerol: A path to economic viability for the biofuels industry. Curr. Opin. Biotechnol. 2007, 18, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Murarka, A.; Dharmadi, Y.; Yazdani, S.S.; Gonzalez, R. Fermentative utilization of glycerol by Escherichia coli and its implications for the production of fuels and chemicals. Appl. Environ. Microbiol. 2008, 74, 1124–1135. [Google Scholar] [CrossRef] [Green Version]
- Clomburg, J.M.; Gonzalez, R. Anaerobic fermentation of glycerol: A platform for renewable fuels and chemicals. Trends Biotechnol. 2013, 31, 20–28. [Google Scholar] [CrossRef] [Green Version]
- Varrone, C.; Rosa, S.; Fiocchetti, F.; Giussani, B.; Izzo, G.; Massini, G.; Marone, A.; Signorini, A.; Wang, A. Enrichment of activated sludge for enhanced hydrogen production from crude glycerol. Int. J. Hydrog. Energy 2013, 38, 1319–1331. [Google Scholar] [CrossRef]
- Varrone, C.; Giussani, B.; Izzo, G.; Massini, G.; Marone, A.; Signorini, A.; Wang, A. Statistical optimization of biohydrogen and ethanol production from crude glycerol by microbial mixed culture. Int. J. Hydrog. Energy 2012, 37, 16479–16488. [Google Scholar] [CrossRef]
- Trchounian, K.; Trchounian, A. Hydrogen production from glycerol by Escherichia coli and other bacteria: An overview and perspectives. Appl. Energy 2015, 156, 174–184. [Google Scholar] [CrossRef]
- Kumar, V.; Park, S. Potential and limitations of Klebsiella pneumoniae as a microbial cell factory utilizing glycerol as the carbon source. Biotechnol. Adv. 2018, 36, 150–167. [Google Scholar] [CrossRef] [PubMed]
- Parate, R.; Mane, R.; Dharne, M.; Rode, C. Mixed bacterial culture mediated direct conversion of bio-glycerol to diols. Bioresour. Technol. 2018, 250, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Mangayil, R.; Karp, M.; Santala, V. Bioconversion of crude glycerol from biodiesel production to hydrogen. Int. J. Hydrog. Energy 2012, 37, 12198–12204. [Google Scholar] [CrossRef]
- de Oliveira Faber, M.; Ferreira-Leitão, V.S. Optimization of biohydrogen yield produced by bacterial consortia using residual glycerin from biodiesel production. Bioresour. Technol. 2016, 219, 365–370. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, C.V.; Nespeca, M.G.; Sakamoto, I.K.; de Oliveira, J.E.; Amâncio Varesche, M.B.; Maintinguer, S.I. Bioconversion of crude glycerol from waste cooking oils into hydrogen by sub-tropical mixed and pure cultures. Int. J. Hydrog. Energy 2019, 44, 144–154. [Google Scholar] [CrossRef] [Green Version]
- Wu, K.J.; Lin, Y.H.; Lo, Y.C.; Chen, C.Y.; Chen, W.M.; Chang, J.S. Converting glycerol into hydrogen, ethanol, and diols with a Klebsiella sp. HE1 strain via anaerobic fermentation. J. Taiwan Inst. Chem. Eng. 2011, 42, 20–25. [Google Scholar] [CrossRef]
- Cofré, O.; Ramírez, M.; Gómez, J.M.; Cantero, D. Pilot scale fed-batch fermentation in a closed loop mixed reactor for the biotransformation of crude glycerol into ethanol and hydrogen by Escherichia coli MG1655. Biomass Bioenergy 2016, 91, 37–47. [Google Scholar] [CrossRef]
- Shams Yazdani, S.; Gonzalez, R. Engineering Escherichia coli for the efficient conversion of glycerol to ethanol and co-products. Metab. Eng. 2008, 10, 340–351. [Google Scholar] [CrossRef]
- Soo, C.S.; Yap, W.S.; Hon, W.M.; Ramli, N.; Md Shah, U.K.; Phang, L.Y. Improvement of hydrogen yield of ethanol-producing Escherichia coli recombinants in acidic conditions. Electron. J. Biotechnol. 2017, 26, 27–32. [Google Scholar] [CrossRef]
- Paesi, S.; Schiavenin, A.; Almeida, L.G.; Andreis, D.; Magrini, F.E.; Marconatto, L.; dos Anjos Borges, L.G.; Giongo, A. Comparison of two different kinds of seed sludge and characterization of microorganisms producing hydrogen and soluble metabolites from raw glycerol. Braz. J. Chem. Eng. 2022, 39, 387–402. [Google Scholar] [CrossRef]
- Sittijunda, S.; Reungsang, A. Valorization of crude glycerol into hydrogen, 1,3-propanediol, and ethanol in an up-flow anaerobic sludge blanket (UASB) reactor under thermophilic conditions. Renew. Energy 2020, 161, 361–372. [Google Scholar] [CrossRef]
- Maru, B.T.; López, F.; Kengen, S.W.M.; Constantí, M.; Medina, F. Dark fermentative hydrogen and ethanol production from biodiesel waste glycerol using a co-culture of Escherichia coli and Enterobacter sp. Fuel 2016, 186, 375–384. [Google Scholar] [CrossRef]
- Marone, A.; Izzo, G.; Mentuccia, L.; Massini, G.; Paganin, P.; Rosa, S.; Varrone, C.; Signorini, A. Vegetable waste as substrate and source of suitable microflora for bio-hydrogen production. Renew. Energy 2014, 68, 6–13. [Google Scholar] [CrossRef]
- Yin, Y.; Song, W.; Wang, J. Inhibitory effect of acetic acid on dark-fermentative hydrogen production. Bioresour. Technol. 2022, 364, 128074. [Google Scholar] [CrossRef] [PubMed]
- Nath, K.; Das, D. Improvement of fermentative hydrogen production: Various approaches. Appl. Microbiol. Biotechnol. 2004, 65, 520–529. [Google Scholar] [CrossRef] [PubMed]
- Nunes Ferraz Júnior, A.D.; Pages, C.; Latrille, E.; Bernet, N.; Zaiat, M.; Trably, E. Biogas sequestration from the headspace of a fermentative system enhances hydrogen production rate and yield. Int. J. Hydrog. Energy 2020, 45, 11011–11023. [Google Scholar] [CrossRef]
- Ito, T.; Nakashimada, Y.; Senba, K.; Matsui, T.; Nishio, N. Hydrogen and ethanol production from glycerol-containing wastes discharged after biodiesel manufacturing process. J. Biosci. Bioeng. 2005, 100, 260–265. [Google Scholar] [CrossRef] [PubMed]
- Reungsang, A.; Sittijunda, S.; Angelidaki, I. Simultaneous production of hydrogen and ethanol from waste glycerol by Enterobacter aerogenes KKU-S1. Int. J. Hydrog. Energy 2013, 38, 1813–1825. [Google Scholar] [CrossRef]
- Magrini, F.E.; Castilhos, A.; Lora, L.B.; Paesi, S. Strategies of co-cultures and bioaugmentation by Bacillus amyloliquefaciens, Clostridium bifermentans, Enterobacter muelleri, and E. tabaci for increasing the production of hydrogen from raw glycerol. Biomass Bioenergy 2023, 168, 106672. [Google Scholar] [CrossRef]
- Rodrigues, C.V.; Rios Alcaraz, F.A.; Nespeca, M.G.; Rodrigues, A.V.; Motteran, F.; Tallarico Adorno, M.A.; Varesche, M.B.A.; Maintinguer, S.I. Biohydrogen production in an integrated biosystem using crude glycerol from waste cooking oils. Renew. Energy 2020, 162, 701–711. [Google Scholar] [CrossRef]
- Boecker, S.; Espinel-Ríos, S.; Bettenbrock, K.; Klamt, S. Enabling anaerobic growth of Escherichia coli on glycerol in defined minimal medium using acetate as redox sink. Metab. Eng. 2022, 73, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Murakami, N.; Oba, M.; Iwamoto, M.; Tashiro, Y.; Noguchi, T.; Bonkohara, K.; Abdel-Rahman, M.A.; Zendo, T.; Shimoda, M.; Sakai, K.; et al. L-Lactic acid production from glycerol coupled with acetic acid metabolism by Enterococcus faecalis without carbon loss. J. Biosci. Bioeng. 2016, 121, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Sawasdee, V.; Vikromvarasiri, N.; Pisutpaisal, N. Optimization of ethanol production from co-substrate of waste glycerol and acetic acid by Enterobacter aerogenes. Biomass Convers. Biorefinery 2021, 1–8. [Google Scholar] [CrossRef]
- Thauer, R.K.; Jungermann, K.; Decker, K. Energy conservation in chemotrophic anaerobic bacteria. Bacteriol. Rev. 1977, 41, 100–180. [Google Scholar] [CrossRef]
- Kyazze, G.; Martinez-Perez, N.; Dinsdale, R.; Premier, G.C.; Hawkes, F.R.; Guwy, A.J.; Hawkes, D.L. Influence of substrate concentration on the stability and yield of continuous biohydrogen production. Biotechnol. Bioeng. 2006, 93, 971–979. [Google Scholar] [CrossRef] [PubMed]
- Ngo, T.A.; Kim, M.S.; Sim, S.J. High-yield biohydrogen production from biodiesel manufacturing waste by Thermotoga neapolitana. Int. J. Hydrog. Energy 2011, 36, 5836–5842. [Google Scholar] [CrossRef]
- Marone, A.; Varrone, C.; Fiocchetti, F.; Giussani, B.; Izzo, G.; Mentuccia, L.; Rosa, S.; Signorini, A. Optimization of substrate composition for biohydrogen production from buffalo slurry co-fermented with cheese whey and crude glycerol, using microbial mixed culture. Int. J. Hydrog. Energy 2015, 40, 209–218. [Google Scholar] [CrossRef]
- Zeng, A.P.; Biebl, H.; Schlieker, H.; Deckwer, W.D. Pathway analysis of glycerol fermentation by Klebsiella pneumoniae: Regulation of reducing equivalent balance and product formation. Enzym. Microb. Technol. 1993, 15, 770–779. [Google Scholar] [CrossRef]
- Kongjan, P.; Jariyaboon, R.; Reungsang, A.; Sittijunda, S. Co-fermentation of 1,3-propanediol and 2,3-butanediol from crude glycerol derived from the biodiesel production process by newly isolated Enterobacter sp.: Optimization factors affecting. Bioresour. Technol. Rep. 2021, 13, 100616. [Google Scholar] [CrossRef]
- Biebl, H.; Zeng, A.P.; Menzel, K.; Deckwer, W.D. Fermentation of glycerol to 1,3-propanediol and 2,3-butanediol by Klebsiella pneumoniae. Appl. Microbiol. Biotechnol. 1998, 50, 24–29. [Google Scholar] [CrossRef] [PubMed]
- Sinha, P.; Roy, S.; Das, D. Role of formate hydrogen lyase complex in hydrogen production in facultative anaerobes. Int. J. Hydrog. Energy 2015, 40, 8806–8815. [Google Scholar] [CrossRef]
- Rosenzweig, R.F.; Sharp, R.R.; Treves, D.S.; Adams, J. Microbial evolution in a simple unstructured environment: Genetic differentiation in Escherichia coli. Genetics 1994, 137, 903–917. [Google Scholar] [CrossRef] [PubMed]
- Neilson, J.W.; Jordan, F.L.; Maier, R.M. Analysis of artifacts suggests DGGE should not be used for quantitative diversity analysis. J. Microbiol. Methods 2013, 92, 256–263. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Process Parameters and Strategies for Optimization | Fed-Batch Experiments | |||||
|---|---|---|---|---|---|---|
| A | B | C | D | E | F | |
| Fed-batch cycles | 1 | 1 | 1 | 1 | 1 | 0 |
| Feed (gCGL−1) | 20 | 20 | 40 | 40 | 60 | 80 |
| N2 sparging | - | - | - | + | + | + |
| CGc (g) | 31 | 29 | 45 | 44 | 50 | 43 |
| CG degradation (%) | 77 | 73 | 56 | 54 | 41 | 54 |
| Time (h) | 213 | 182 | 212 | 141 | 190 | 160 |
| µ (h−1) | 0.055 | 0.071 | 0.074 | 0.125 | 0.090 | 0.060 |
| EtOH (gL−1): | 14 | 12 | 20 | 20 | 21 | 16 |
| -yield (molEtOHmol−1CGC) | 0.84 | 0.82 | 0.87 | 0.92 | 0.83 | 0.78 |
| Total biogas (H2 + CO2, LL−1) | 13 | 13 | 18 | 17 | 20 | 17 |
| H2 (LL−1): | 6 | 6 | 8 | 10 | 11 | 8 |
| -yield (molH2mol−1CGC) | 0.63 | 0.74 | 0.67 | 0.87 | 0.78 | 0.75 |
| -Pmax (L) | 5.280 | 5.784 | 7.983 | 11.293 | 10.913 | 8.770 |
| -Rmax (Lh−1) | 0.054 | 0.096 | 0.096 | 0.112 | 0.100 | 0.113 |
| -λ (h) | 2.34 | 0.00 | 0.00 | 3.00 | 5.67 | 9.84 |
| -R2 | 0.992 | 0.988 | 0.994 | 0.998 | 0.998 | 1.000 |
| Inoculum | Fermentation | Glycerol | EtOH | H2 | Ref. | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Mode (st/ns) | N2 Source | T °C | pH Start/Co | Time h | C/P gL−1 | Degradation (%) | molmol−1/ gL−1 | molmol−1/ LL−1 | |||
| 1 | E. aerogenes HU-101 | Batch (st) | Ye/ tryp | 37 | 6.8 (Co) | 4 | C 1.7 | 100 | 0.96/nc | 1.12/nc | [29] |
| 4 | 3.3 | 0.83/nc | 0.90/nc | ||||||||
| 12 | 10 | 0.67/nc | 0.71/nc | ||||||||
| <48 | 25 | <100 | 0.56/nc | 0.71/nc | |||||||
| 2 | Klebsiella HE1 | Batch (-) | no | 35 | 6.0 | 35 | P 10 | 99.9 | 0.80/4.0 | 0.04/0.71 | [18] |
| 50 | 0.49/12.2 | 0.35/6.9 | |||||||||
| 3 | E. aerogenes KKU-S1 | Batch (ns) | Ye | 37 | 8.13 | nr | C 31 | nr | 0.85/5.5 | 0.12/nc | [30] |
| 4 | E. coli MG1655 | Exponential Fed-b/Continuous Ar sparging (st) | Pep | 37 | 6.30–6.35 (Co) | 88 | C 37.7 | 77 ± 7.6 | 0.66/6.3 | 0.56/2.2 * | [19] |
| 5 | E. coli SS1 wt | Batch | Ye/Pep | 37 | 7.5 5.8 | 72 | P10 | nr 56 | 0.57/2.8 0.70/4.8 | 0.57/1.7 0.46/1.0 | [21] |
| 6 | E. coli SS1 hydA | 7.5 5.8 | nr 49 | 0.76/3.3 0.68/2.4 | 0.28/0.8 0.58/1.0 | ||||||
| 7 | E. coli CECT432+ Enterobacter SPH1 | Batch (st) | Ye/ Me | 37 | 6.7 | 72 | C 26.7 * | 63 * | 1.21/10.0 * | 1.53/7.0 * | [24] |
| 8 | BA + CB + ET | Batch | Ye/ Pep | 37 | nr | 72 | C 30 | 59 | 0.14/1.3 * | 0.95/3.1 * | [31] |
| ET + CB | 50 * | 0.44 */3.3 | 0.5 */2.1 | ||||||||
| 9 | Thermophilic mixed culture | Continuous (UASB) (ns) | Urea | 55 | 5.5 (Co) | - | C 62.5 | 33 | 0.22/2.25 * | 2.05/12.3 * | [23] |
| 10 | Mixed culture (vegetable oil industry) | Batch (ns) | Ye/ Tryp | 37 | 6.0 | 72 | C 30 | 60.9 | 0.29 */1.6 | 1.12/2.7 * | [22] |
| 11 | Mixed culture (wine industry) | 63.5 | 0.27 */2.5 | 0.96/1.9 * | |||||||
| 12 | Thermophilic mixed Clostridiales | Batch | Ye/Me/ Pep | 37 | 5.5 | 85 | C 2.58 | 45.7 | 1.57 */0.86 * | 1.75/0.57 | [32] |
| 13 | Enriched consortium (anaerobic sludge) | Fed-b/ Intermittent N2 sparging (ns) | no | 37 | 7.9 | 141 | C 40 (batch + feed) | 54 | 0.92/20.0 | 0.87/10.0 | This study |
| Fed-Batch Experiments | Cycles | pH (Start/End) | TAA gL−1 | UAA mgL−1 |
|---|---|---|---|---|
| A | batch | 7.9/6.4 | 1.0 | 0.7 |
| B | 7.9/5.9 | 1.0 | 0.7 | |
| C | 7.9/6.0 | 2.1 | 1.4 | |
| D | 7.9/6.0 | 2.0 | 1.3 | |
| E | 8.0/6.0 | 3.3 | 1.8 | |
| F | 8.0/6.0 | 4.1 | 2.2 | |
| A | feed | 6.5/6.1 | 1.1 | 20.4 |
| B | 5.9/5.8 | 1.0 | 61.3 | |
| C | 5.9/5.6 | 2.2 | 121.0 | |
| D | 6.1/5.5 | 2.2 | 90.2 | |
| E | 6.0/5.8 | 3.0 | 159.0 | |
| F | - | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mehariya, S.; Signorini, A.; Marone, A.; Rosa, S. Simultaneous Hydrogen and Ethanol Production from Crude Glycerol by a Microbial Consortium Using Fed-Batch Fermentation. Energies 2023, 16, 4490. https://doi.org/10.3390/en16114490
Mehariya S, Signorini A, Marone A, Rosa S. Simultaneous Hydrogen and Ethanol Production from Crude Glycerol by a Microbial Consortium Using Fed-Batch Fermentation. Energies. 2023; 16(11):4490. https://doi.org/10.3390/en16114490
Chicago/Turabian StyleMehariya, Sanjeet, Antonella Signorini, Antonella Marone, and Silvia Rosa. 2023. "Simultaneous Hydrogen and Ethanol Production from Crude Glycerol by a Microbial Consortium Using Fed-Batch Fermentation" Energies 16, no. 11: 4490. https://doi.org/10.3390/en16114490