
In Situ Biogas Upgrading in a Randomly Packed Gas-Stirred Tank Reactor (GSTR)

Abstract
:1. Introduction
2. Materials and Methods
2.1. Inoculum and Substrate
2.2. Reactor Set Up and Operation
2.3. Analytical Methods
2.4. Calculation
2.5. Illumina Sequencing
3. Results and Discussion
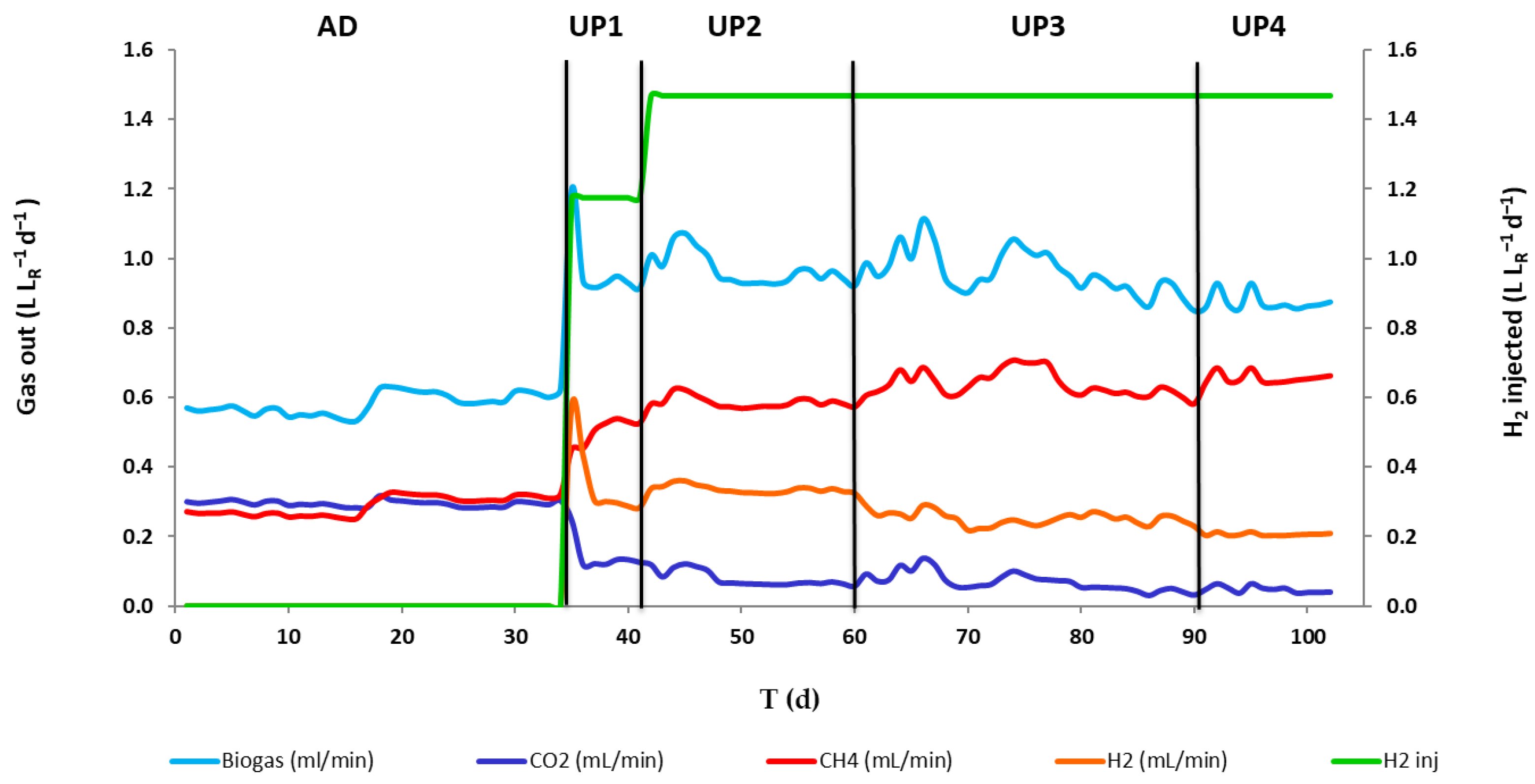
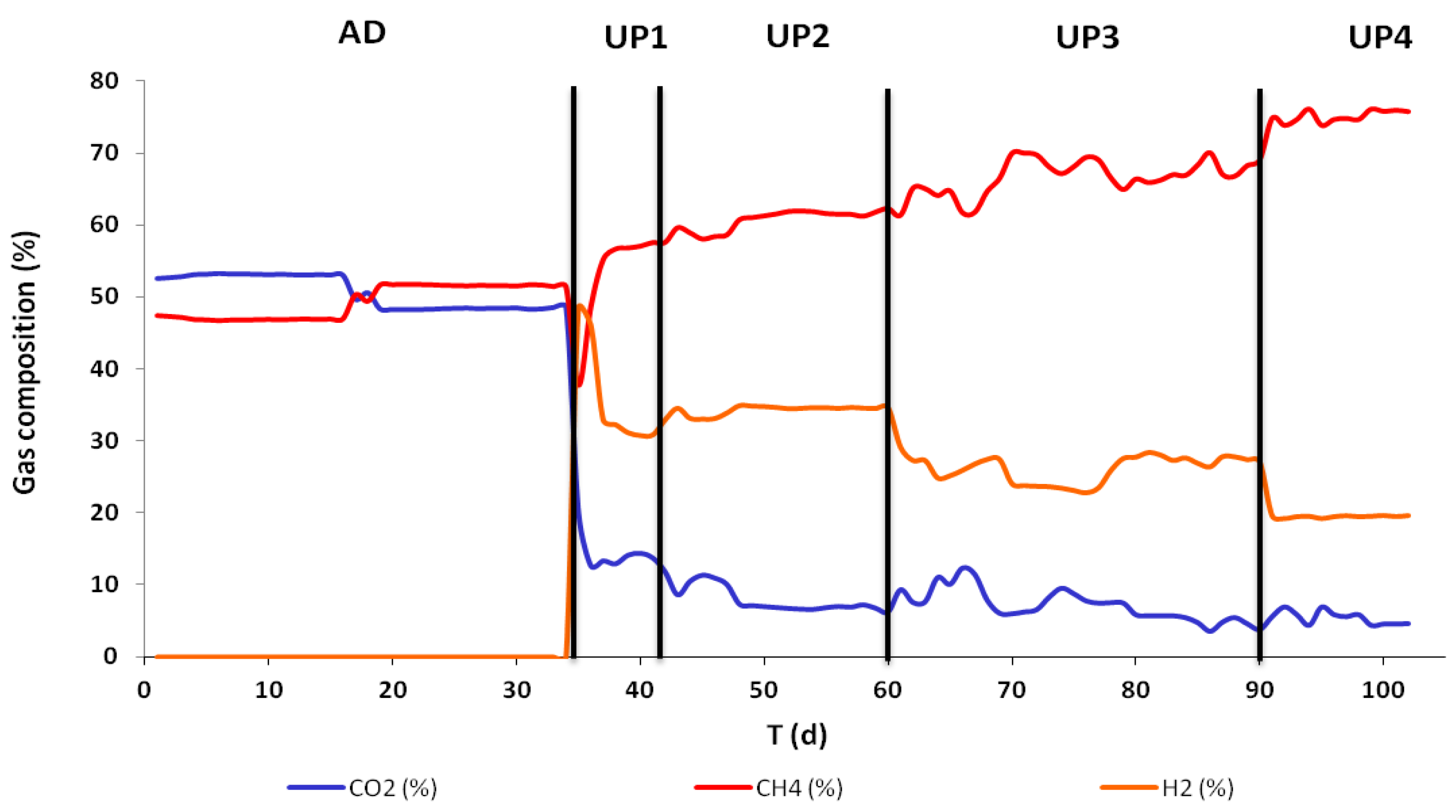
3.1. Reactor Performance
3.2. pH, VFAs Evolution and Organic Matter Removal
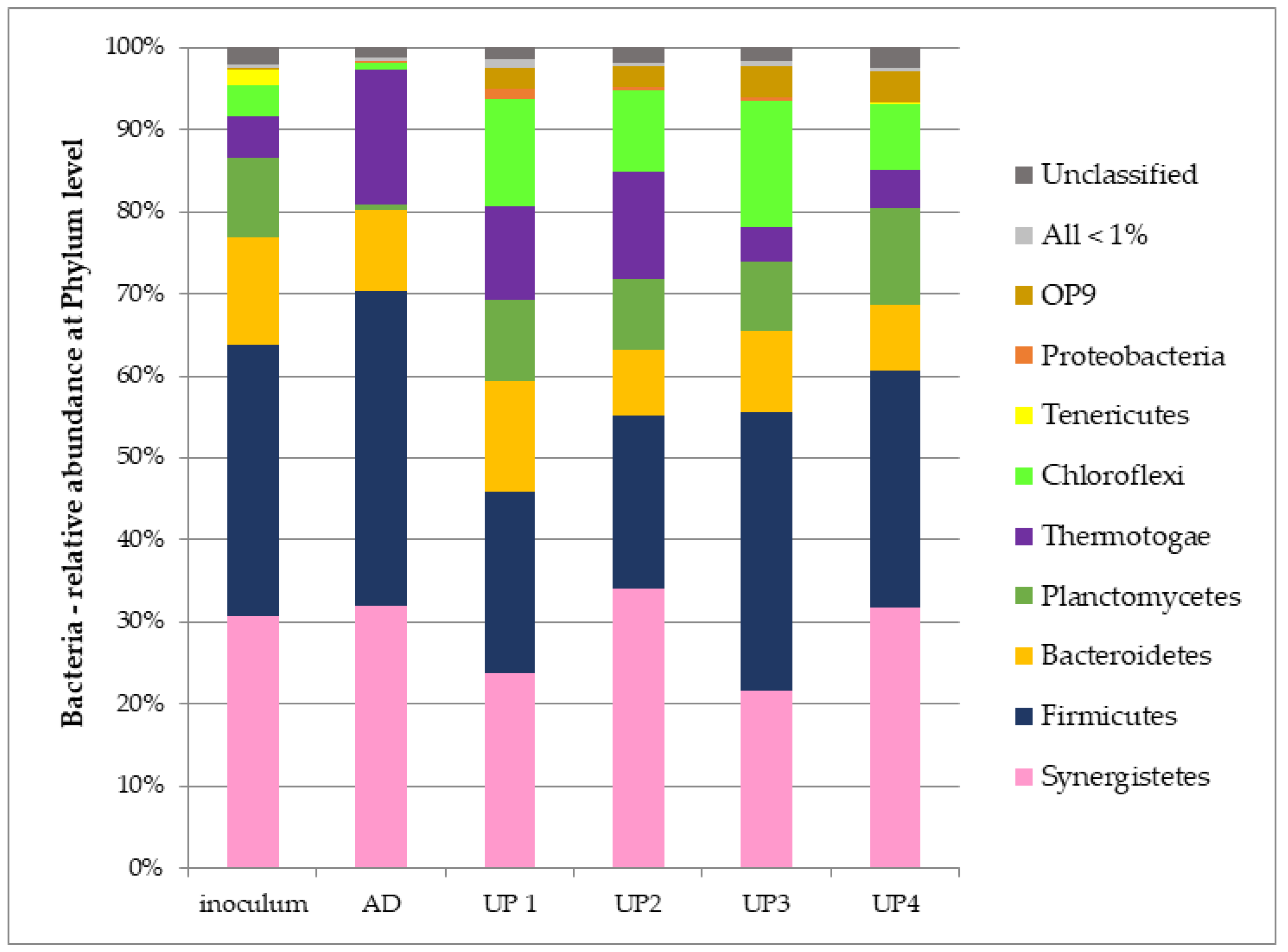
3.3. Microbial Communities
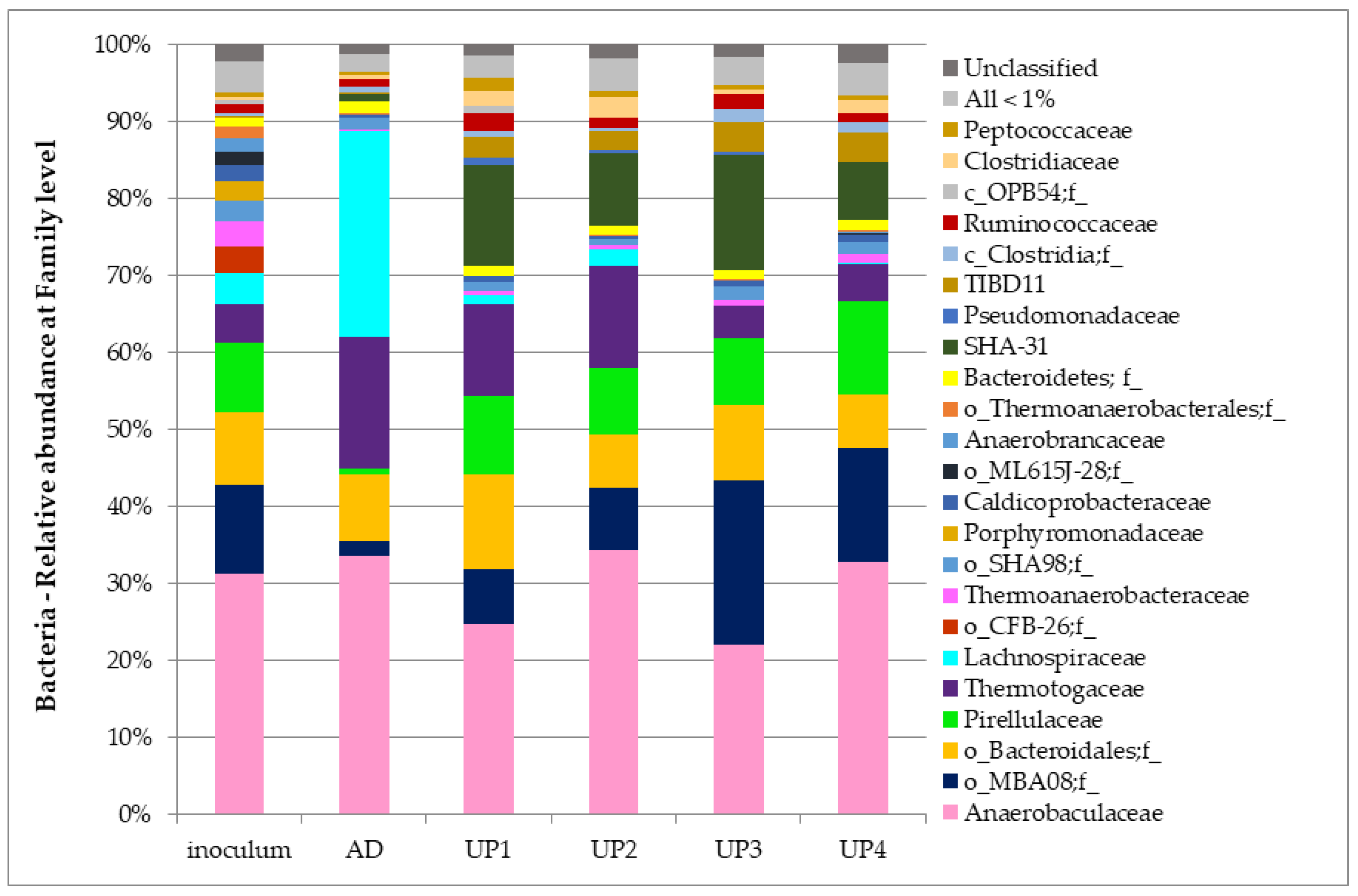
3.4. Bacteria Communities
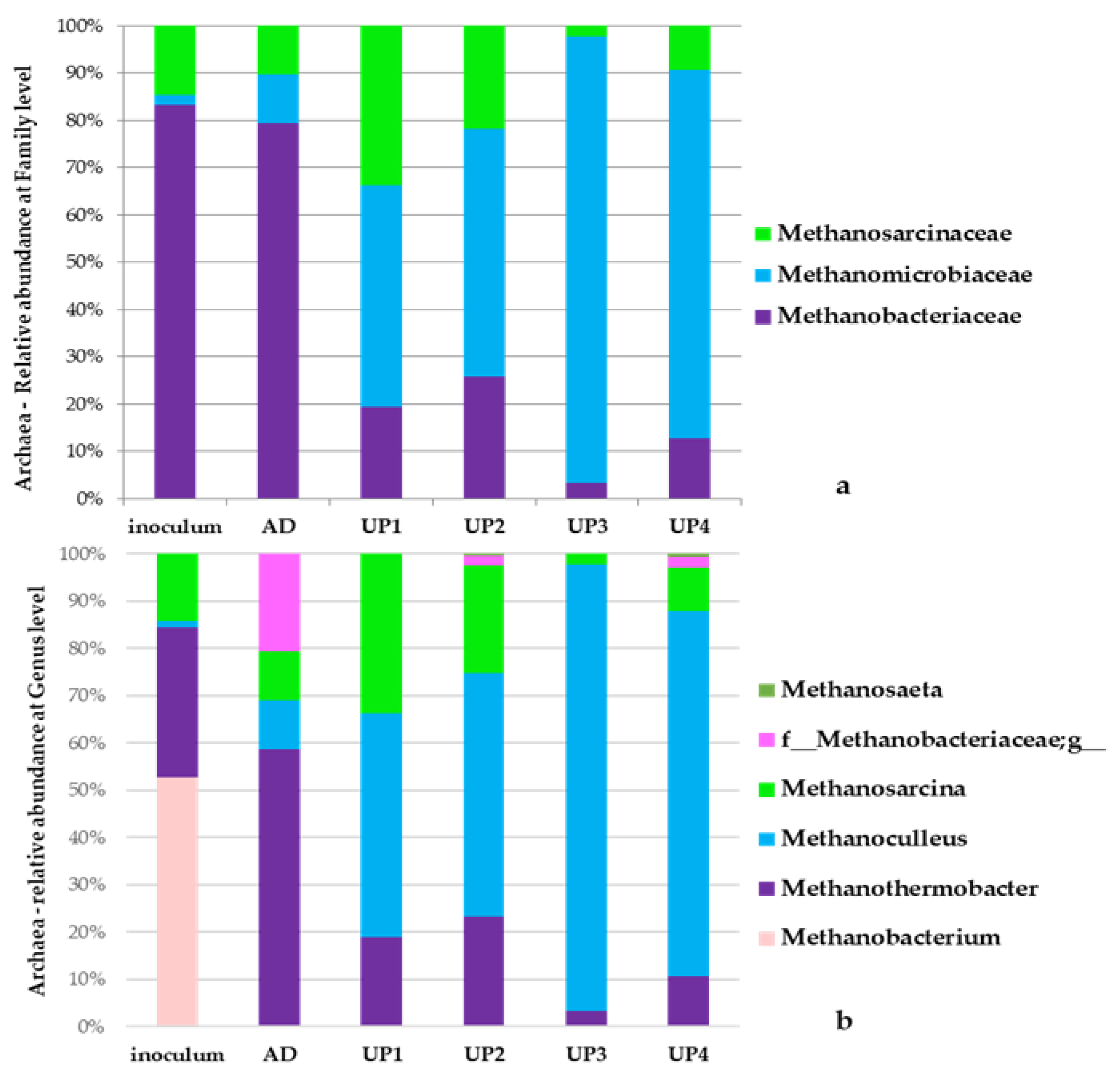
3.5. Archaea Communities
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| AD | Anaerobic digestion |
| CHP | Combined heat and power units |
| CSTR | Continuously stirred tank reactor |
| GSTR | Gas-stirred tank reactor |
| HDPE | High-density polietylene |
| HFM | Hollow fiber membrane |
| HPLC | High-pressure liquid chromatography |
| HRT | Hydraulic retention time |
| MER | Methane evolution rate |
| NaCl | Sodium chloride |
| OLR | Organic loading rate |
| SAO | Syntrophic acetate oxidation |
| SCW | Second cheese whey |
| UP | Upgrading phase |
| VFA | Volatile fatty acid |
| VS | Volatile solids |
| Symbols | |
| H2 | H2 conversion efficiency |
| rt | H2 gas-liquid mass transfer rate |
| LR | Reactor Working volume (L) |
| KH | Henry’s constant |
| LH2 | Volume of H2 |
| LCH4 | Volume of CH4 |
| RH2 | Volumetric hydrogen mass transfer rate |
| (KLa)H2 | Volumetric hydrogen mass coefficient |
| CH2,G | Gaseous hydrogen concentration |
| CH2,L | Dissolved hydrogen concentration |
References
- Osman, A.I.; Chen, L.; Yang, M.; Msigwa, G.; Farghali, M.; Fawzy, S.; Rooney, D.W.; Yap, P.S. Cost, environmental impact, and resilience of renewable energy under a changing climate: A review. Environ. Chem. Lett. 2022, 21, 741–764. [Google Scholar] [CrossRef]
- Chen, L.; Msigwa, G.; Yang, M.; Osman, A.I.; Fawzy, S.; Rooney, D.W.; Yap, P.S. Strategies to achieve a carbon neutral society: A review. Environ. Chem. Lett. 2022, 20, 2277–2310. [Google Scholar] [CrossRef] [PubMed]
- Farghali, M.; Osman, A.I.; Umetsu, K.; Rooney, D.W. Integration of biogas systems into a carbon zero and hydrogen economy: A review. Environ. Chem. Lett. 2022, 20, 2853–2927. [Google Scholar] [CrossRef]
- Kougias, P.G.; Angelidaki, I. Biogas and its opportunities—A review. Front. Environ. Sci. Eng. 2018, 12, 14. [Google Scholar] [CrossRef]
- Ghaib, K.; Ben-Fares, F.Z. Power-to-Methane: A state-of-the-art review. Renew. Sustain. Energy Rev. 2018, 81, 433–446. [Google Scholar] [CrossRef]
- Kapoor, R.; Ghosh, P.; Kumar, M.; Vijay, V.K. Evaluation of biogas upgrading technologies and future perspectives: A review. Environ. Sci. Pollut. Res. 2019, 26, 11631–11661. [Google Scholar] [CrossRef]
- Bassani, I.; Kougias, P.G.; Treu, L.; Porté, H.; Campanaro, S.; Angelidaki, I. Optimization of hydrogen dispersion in thermophilic up-flow reactors for ex situ biogas upgrading. Bioresour. Technol. 2017, 234, 310–319. [Google Scholar] [CrossRef]
- Vo, T.T.Q.; Wall, D.M.; Ring, D.; Rajendran, K.; Murphy, J.D. Techno-economic analysis of biogas upgrading via amine scrubber, carbon capture and ex-situ methanation. Appl. Energy 2018, 212, 1191–1202. [Google Scholar] [CrossRef]
- Luo, G.; Angelidaki, I. Co-digestion of manure and whey for in situ biogas upgrading by the addition of H2: Process performance and microbial insights. Appl. Microbiol. Biotechnol. 2013, 97, 1373–1381. [Google Scholar] [CrossRef]
- Thauer, R.K.; Jungermann, K.; Decker, K. Energy conservation in chemotrophic anaerobic bacteria. Bacteriol. Rev. 1977, 41, 100–180. [Google Scholar] [CrossRef]
- Zabranska, J.; Pokorna, D. Bioconversion of carbon dioxide to methane using hydrogen and hydrogenotrophic methanogens. Biotechnol. Adv. 2018, 36, 707–720. [Google Scholar] [CrossRef]
- Alfaro, N.; Fdz-Polanco, M.; Fdz-Polanco, F.; Díaz, I. H2 addition through a submerged membrane for in-situ biogas upgrading in the anaerobic digestion of sewage sludge. Bioresour. Technol. 2019, 280, 1–8. [Google Scholar] [CrossRef]
- Narayanan, C.M.; Narayan, V. Biological wastewater treatment and bioreactor design: A review. Sustain. Environ. Res. 2019, 1, 33. [Google Scholar] [CrossRef] [Green Version]
- Voelklein, M.A.; Rusmanis, D.; Murphy, J.D. Biological methanation: Strategies for in-situ and ex-situ upgrading in anaerobic digestion. Appl. Energy 2019, 235, 1061–1071. [Google Scholar] [CrossRef]
- Angelidaki, I.; Treu, L.; Tsapekos, P.; Luo, G.; Campanaro, S.; Wenzel, H.; Kougias, P.G. Biogas upgrading and utilization: Current status and perspectives. Biotechnol. Adv. 2018, 36, 452–466. [Google Scholar] [CrossRef] [Green Version]
- Ullrich, T.; Lindner, J.; Bär, K.; Mörs, F.; Graf, F.; Lemmer, A. Influence of operating pressure on the biological hydrogen methanation in trickle-bed reactors. Bioresour. Technol. 2018, 247, 7–13. [Google Scholar] [CrossRef]
- Luo, G.; Johansson, S.; Boe, K.; Xie, L.; Zhou, Q.; Angelidaki, I. Simultaneous hydrogen utilization and in situ biogas upgrading in an anaerobic reactor. Biotechnol. Bioeng. 2012, 109, 1088–1094. [Google Scholar] [CrossRef]
- Luo, G.; Angelidaki, I. Hollow fiber membrane based H2 diffusion for efficient in situ biogas upgrading in an anaerobic reactor. Appl. Microbiol. Biotechnol. 2013, 97, 3739–3744. [Google Scholar] [CrossRef]
- Agneessens, L.M.; Ottosen, L.D.M.; Voigt, N.V.; Nielsen, J.L.; de Jonge, N.; Fischer, C.H.; Kofoed, M.V.W. In-situ biogas upgrading with pulse H2 additions: The relevance of methanogen adaption and inorganic carbon level. Bioresour. Technol. 2017, 233, 256–263. [Google Scholar] [CrossRef]
- Zhao, J.; Li, Y.; Dong, R. Recent progress towards in-situ biogas upgrading technologies. Sci. Total Environ. 2021, 800, 149667. [Google Scholar] [CrossRef]
- Lecker, B.; Illi, L.; Lemmer, A.; Oechsner, H. Biological hydrogen methanation—A review. Bioresour. Technol. 2017, 245, 1220–1228. [Google Scholar] [CrossRef]
- Burkhardt, M.; Busch, G. Methanation of hydrogen and carbon dioxide. Appl. Energy 2013, 111, 74–79. [Google Scholar] [CrossRef]
- Rachbauer, L.; Voitl, G.; Bochmann, G.; Fuchs, W. Biological biogas upgrading capacity of a hydrogenotrophic community in a trickle-bed reactor. Appl. Energy 2016, 180, 483–490. [Google Scholar] [CrossRef]
- Kougias, P.G.; Tsapekos, P.; Treu, L.; Kostoula, M.; Campanaro, S.; Lyberatos, G.; Angelidaki, I. Biological CO2 fixation in up-flow reactors via exogenous H2 addition. J. Biotechnol. 2020, 319, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Lembo, G.; Rosa, S.; Miritana, V.M.; Marone, A.; Massini, G.; Fenice, M.; Signorini, A. Thermophilic anaerobic digestion of second cheese whey: Microbial community response to H2 addition in a partially immobilized anaerobic hybrid reactor. Processes 2021, 9, 43. [Google Scholar] [CrossRef]
- Schink, B. Energetics of syntrophic cooperation in methanogenic degradation. Microbiol. Mol. Biol. Rev. 1997, 61, 262–280. [Google Scholar]
- Labatut, R.A.; Angenent, L.T.; Scott, N.R. Conventional mesophilic vs. thermophilic anaerobic digestion: Atrade-off between performance and stability? Water Res. 2014, 53, 249–258. [Google Scholar] [CrossRef]
- Zhu, X.; Chen, L.; Chen, Y.; Cao, Q.; Liu, X.; Li, D. Differences of methanogenesis between mesophilic and thermophilic in situ biogas-upgrading systems by hydrogen addition. J. Ind. Macrobiol. Biotecnol. 2019, 46, 1569–1581. [Google Scholar] [CrossRef]
- Baird, B.; Eaton, D.A.; Rice, E.W. Standard Methods for the Examination of Water and Wastewater; APHA: Washington, DC, USA, 2017; Volume 23. [Google Scholar]
- Díaz, I.; Pérez, C.; Alfaro, N.; Fdz-Polanco, F. A feasibility study on the bioconversion of CO2 and H2 to biomethane by gas sparging through polymeric membranes. Bioresour. Technol. 2015, 185, 246–253. [Google Scholar] [CrossRef]
- Agneessens, L.M.; Ottosen, L.D.M.; Andersen, M.; Berg Olesen, C.; Feilberg, A.; Kofoed, M.V.W. Parameters affecting acetate concentrations during in-situ biological hydrogen methanation. Bioresour. Technol. 2018, 258, 33–40. [Google Scholar] [CrossRef]
- Mulat, D.G.; Mosbæk, F.; Ward, A.J.; Polag, D.; Greule, M.; Keppler, F.; Nielsen, J.L.; Feilberg, A. Exogenous addition of H2 for an in situ biogas upgrading through biological reduction of carbon dioxide into methane. Waste Manag. 2017, 68, 146–156. [Google Scholar] [CrossRef]
- Treu, L.; Tsapekos, P.; Peprah, M.; Campanaro, S.; Giacomini, A.; Corich, V.; Kougias, P.G.; Angelidaki, I. Microbial profiling during anaerobic digestion of cheese whey in reactors operated at different conditions. Bioresour. Technol. 2019, 275, 375–385. [Google Scholar] [CrossRef]
- Kougias, P.G.; Treu, L.; Benavente, D.P.; Boe, K.; Campanaro, S.; Angelidaki, I. Ex-situ biogas upgrading and enhancement in different reactor systems. Bioresour. Technol. 2017, 225, 429–437. [Google Scholar] [CrossRef]
- Jabari, L.; Gannoun, H.; Cayol, J.L.; Hamdi, M.; Fauque, G.; Ollivier, B.; Fardeau, M.L. Characterization of Defluviitalea saccharophila gen. nov., sp. nov., a thermophilic bacterium isolated from an upflow anaerobic filter treating abattoir wastewaters, and proposal of defluviitaleaceae fam. nov. Int. J. Syst. Evol. Microbiol. 2012, 62, 550–555. [Google Scholar] [CrossRef]
- Fontana, A.; Campanaro, S.; Treu, L.; Kougias, P.G.; Cappa, F.; Morelli, L.; Angelidaki, I. Performance and genome-centric metagenomics of thermophilic single and two-stage anaerobic digesters treating cheese wastes. Water Res. 2018, 134, 181–191. [Google Scholar] [CrossRef]
- Li, D.; Ran, Y.; Chen, L.; Cao, Q.; Li, Z.; Liu, X. Instability diagnosis and syntrophic acetate oxidation during thermophilic digestion of vegetable waste. Water Res. 2018, 139, 263–271. [Google Scholar] [CrossRef]
- Rivière, D.; Desvignes, V.; Pelletier, E.; Chaussonnerie, S.; Guermazi, S.; Weissenbach, J.; Li, T.; Camacho, P.; Sghir, A. Towards the definition of a core of microorganisms involved in anaerobic digestion of sludge. ISME J. 2009, 3, 700–714. [Google Scholar] [CrossRef] [Green Version]
- Nelson, M.C.; Morrison, M.; Yu, Z. A meta-analysis of the microbial diversity observed in anaerobic digesters. Bioresour. Technol. 2011, 102, 3730–3739. [Google Scholar] [CrossRef]
- Treu, L.; Campanaro, S.; Kougias, P.G.; Sartori, C.; Bassani, I.; Angelidaki, I. Hydrogen-fueled microbial pathways in biogas upgrading systems revealed by genome-centric metagenomics. Front. Microbiol. 2018, 9, 1079. [Google Scholar] [CrossRef] [Green Version]
- Braga Nan, L.; Trably, E.; Santa-Catalina, G.; Bernet, N.; Delgenès, J.P.; Escudié, R. Biomethanation processes: New insights on the effect of a high H2partial pressure on microbial communities. Biotechnol. Biofuels 2020, 13, 141. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Indicators | Range |
|---|---|
| pH | 5.9–6.2 |
| Lactose (gL−1) | 40–60 |
| Total Solids (TS) (gL−1) | 47–64 |
| Volatile Solids (VS) (gL−1) | 40–54 |
| COD (gL−1) | 45–70 |
| Proteins (gL−1) | 0.45–0.90 |
| NH4+ (gL−1) | 0.10–0.12 |
| Total Volatile fatty acids (TVFAs) (gL−1) | 1.5–2.5 |
| Parameters | AD | UP1 | UP2 | UP3 | UP4 |
|---|---|---|---|---|---|
| OLR (g VS LR−1 d−1) | 1.20 ± 0.02 | 1.16 ± 0.08 | 1.16 ± 0.05 | 1.26 ± 0.09 | 1.28 ± 0.08 |
| H2In Flow rate (LH2 LR−1 d−1) | / | 1.18 ± 0.01 | 1.47 ± 0.01 | 1.47 ± 0.01 | 1.47 ± 0.01 |
| Biogas recirculation rate (L LR−1 d−1) | 118 ± 5 | 118 ± 5 | 118 ± 5 | 176 ± 5 | 235 ± 5 |
| Parameters | AD | UP1 | UP2 | UP3 | UP4 |
|---|---|---|---|---|---|
| Biogasout production rate (L LR−1 d−1) | 0.58 ± 0.03 | 0.93 ± 0.01 | 0.97 ± 0.05 | 0.96 ± 0.06 | 0.87 ± 0.03 |
| H2out% | / | 31.64 ± 0.96 | 34.3 ± 0.67 | 26.2 ± 1.86 | 19.5 ± 0.14 |
| CH4out% | 49.3 ± 2.3 | 56.6 ± 0.89 | 60.6 ±1.52 | 66.6 ± 2.43 | 75.0 ± 0.83 |
| CO2out% | 50.7 ± 2.3 | 13.7 ± 0.59 | 8.04 ± 1.87 | 7.17 ± 2.23 | 5.44 ± 0.93 |
| H2out Flow rate (L LR−1 d−1) | / | 0.29 ± 0.01 | 0.33 ± 0.01 | 0.25 ± 0.02 | 0.17 ± 0.001 |
| CH4out Flow rate (L LR−1 d−1) | 0.29 ± 0.01 | 0.53 ± 0.01 | 0.59 ± 0.02 | 0.64 ± 0.02 | 0.66 ± 0.02 |
| CO2out Flow rate (L LR−1 d−1) | 0.3 ± 0.03 | 0.13 ± 0.01 | 0.08 ± 0.02 | 0.07 ± 0.03 | 0.05 ± 0.01 |
| rt (L LR−1 d−1) | / | 0.88 | 1.14 | 1.22 | 1.30 |
| H2 (%) | / | 75 | 77 | 83 | 88 |
| MER (LCH4 LR−1 d−1) | / | 0.24 | 0.30 | 0.35 | 0.37 |
| Lactose (mg/L) | 224 ± 13 | 168 ± 12 | 148 ± 13 | 159 ± 6.85 | 172 ± 10 |
| Acetic Acid (mg/L) | 33 ± 1.8 | 32 ± 1.5 | 41 ± 7 | 159 ± 15 | 358 ± 10 |
| pH | 7.1 ± 0.14 | 7.81 ± 0.20 | 8.12 ± 0.18 | 8.2 ± 0.15 | 8.24 ± 0.10 |
| VS Effluent (g/L) | 4.78 ± 0.15 | 4.76 ± 0.12 | 4.81 ± 0.14 | 4.96 ± 2.1 | 4.85 ± 0.45 |
| Samples | Diversity | Evenness | ||
|---|---|---|---|---|
| Simpson | Shannon | Simpson | Shannon | |
| (1-D) | (H) | (1/DS) | (H/ln(S)) | |
| Inoculum | 0.85 | 2.23 | 0.31 | 0.72 |
| AD | 0.77 | 1.67 | 0.21 | 0.55 |
| UP1 | 0.87 | 2.13 | 0.43 | 0.74 |
| UP2 | 0.82 | 1.97 | 0.31 | 0.68 |
| UP3 | 0.85 | 2.00 | 0.38 | 0.69 |
| UP4 | 0.83 | 1.98 | 0.26 | 0.64 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lembo, G.; Rosa, S.; Marone, A.; Signorini, A. In Situ Biogas Upgrading in a Randomly Packed Gas-Stirred Tank Reactor (GSTR). Energies 2023, 16, 3296. https://doi.org/10.3390/en16073296
Lembo G, Rosa S, Marone A, Signorini A. In Situ Biogas Upgrading in a Randomly Packed Gas-Stirred Tank Reactor (GSTR). Energies. 2023; 16(7):3296. https://doi.org/10.3390/en16073296
Chicago/Turabian StyleLembo, Giuseppe, Silvia Rosa, Antonella Marone, and Antonella Signorini. 2023. "In Situ Biogas Upgrading in a Randomly Packed Gas-Stirred Tank Reactor (GSTR)" Energies 16, no. 7: 3296. https://doi.org/10.3390/en16073296