1. Introduction
The sustainable management of waste activated sludge (WAS) generated from conventional wastewater treatment plants (WWTPs) represents an important environmental, economic, and technical challenge. In Korea, four million tons of WAS was generated in 698 WWTPs in 2020 [
1]. The typical sludge yield coefficient of activated sludge processes is 0.5, and the costs of sludge treatment and disposal account for approximately 50% to 60% of the total operational costs of a WWTP [
2]. Solutions to WAS-associated issues include (a) a reduction in sludge generation in WWTPs through the adoption of paradigm-shifting strategies that advance the role of novel anaerobic and anoxic bio-based mechanisms instead of heavily relying on conventional treatment processes, such as anaerobic membrane biotechnology and anaerobic ammonium oxidation, and (b) the use of sludge as biomass for the production of renewable energy and as CO
2 adsorption materials to compensate for sludge treatment costs. The latter solution is more practical, and various strategies to achieve it are being engineered based on diverse technologies [
3,
4,
5].
Anaerobic digestion (AD) is the main technology for converting organic waste into usable energy via anaerobic biological reactors, which convert organics into CH
4-rich biogas [
6]. The produced biogas can be used for heating and electricity production via generators or CH
4 fuel cells. The AD system can be divided into four sequential food webs (hydrolysis, acidogenesis, acetogenesis, and methanogenesis), and different microbial communities are used for CH
4 conversion. Among these steps, hydrolysis is the rate-limiting step when particulate organic wastes are used. During hydrolysis, complex organic polymers such as carbohydrates, lipids, and proteins are degraded into soluble monomers, such as sugars, long-chain fatty acids, and amino acids, by extracellular microbial enzymes [
7,
8].
WAS includes particulate organic polymers and is mainly composed of excess bacterial biomass with cell walls. Therefore, the AD of WAS is often limited by slow hydrolysis, which leads to poor CH
4 productivity. Only a fraction of the overall available energy is recovered, and the AD system often remains energy-negative [
9]. WAS pretreatment includes thermal, physical, chemical, and biological methods, which can disintegrate the cell walls in WAS and release intracellular material into the liquid phase, where soluble organics are readily available for anaerobic degradation [
10]. However, the energy feasibility of WAS pretreatments, which are critical for the successful employment of full-scale AD systems, is still not optimal [
11]. The reported thermal, physical, and chemical WAS pretreatments are effective but require high electricity loads or hazardous and expensive chemicals [
12].
Biological WAS pretreatment has received interest as a sustainable alternative to energy- and chemical-consuming methods owing to its viability, reliability, and commercial prospects for large-scale production. In this method, fungi, bacteria, or their enzymes are used to convert organic biomass from its native form into a more accessible form for microbial degradation [
13]. The direct use of enzymes as a pretreatment is more efficient and simple for WAS degradation than the use of fungal and bacterial pretreatments, and it can generally be achieved using commercial enzymes (e.g., amylase, cellulase, protease, and lipase) [
14].
The efficiency of an enzymatic pretreatment depends on the chemical composition of the biomass. In general, the cell walls in WAS are mainly composed of heterogeneous organic components mixed with lipids, proteins, and carbohydrates, and each hydrolytic enzyme interacts with a target substrate depending on its exquisite selectivity and specificity associated with the different shapes of enzyme molecules. Therefore, rather than a single hydrolytic enzyme, multiple hydrolytic enzymes can be simultaneously reacted to effectively hydrolyze WAS [
15]. In addition, each enzyme works more efficiently under specific reaction conditions, such as temperature and pH. That is, the catalytic activity of an enzyme during the pretreatment is temperature- and pH-sensitive [
16]. Therefore, when an enzymatic pretreatment is adopted, reaction conditions should be considered to obtain an efficient and reproducible enzymatic reaction. There have been many attempts to use enzymatic pretreatments of organic wastes, including the use of amylase, protease, lipase, and cellulase. However, this method still has inherent limitations for its practical and universal application owing to the high costs of commercial enzymes. For example, in a full-scale AD plant treating organic waste, at least hundreds of kilograms of commercial enzymes are needed to treat hundreds of tons of organic waste daily [
17].
Enzyme cocktails include a mix of cultured microorganisms produced during fermentation, and they have a high potential for the hydrolysis of heterogeneous organic components, which can substantially reduce the operational costs of enzymatic pretreatments. A viable method to obtain an enzyme cocktail is to extract in situ-produced crude hydrolytic extracellular enzymes (CHEEs) from readily biodegradable organic wastes such as food waste (FW), in which diverse enzymes are available for biodegradation [
15,
17]. For example, CHEEs generated from the acid fermentation of FW include a variety of enzymes that can potentially be used as a substitute for commercial enzymes, with technical and economic advantages.
The present work investigates the feasibility of CHEEs as substitutes for commercial enzymes in a series of anaerobic batch tests using pretreated WAS. The CH4 productivity of WAS AD was evaluated in response to the application of enzymatic pretreatment using commercial enzymes. CHEEs extracted from FW fermentation were used as pretreatment agents for WAS, and their performance was compared to that of commercial enzymes. In addition, a mixture of commercial enzymes at different concentrations was employed for WAS pretreatment to quantitatively evaluate the performance of CHEEs on AD. Finally, microbial community changes were monitored to further clarify the microbial action during the batch tests.
2. Materials and Methods
2.1. Preparation of Feedstock, Inoculum, and Enzymes
The WAS used as feedstock was obtained from a Daejeon Metropolitan City WWTP in South Korea and stored in a refrigerator at 4 °C before use. The total chemical oxygen demand (TCOD), total solids (TS), and volatile solids (VS) concentration of the WAS feedstock were 173 ± 4, 145 ± 3, and 122 ± 2 g/L, respectively. The seed sludge was obtained from an anaerobic digester of a brewery wastewater treatment plant located in Cheongju, South Korea, and the samples were prepared after being ground to a diameter of less than 2 mm. The pH and volatile suspended solids (VSS) concentration of the inoculum were 7.5 and 306 ± 23 g/L.
To prepare the CHEEs, FW was fermented. FW collected from a cafeteria of Chungbuk National University in South Korea was ground for homogenization using a blender and then pretreated by heating it at 90 °C for 20 min before its addition to the bioreactor for CHEE production. The FW was added to a bioreactor with a working volume of 2.2 L, and the TCOD of the FW was controlled at 40 g∙COD/L by adding distilled water. FW was fermented without the addition of an inoculum. The initial pH was adjusted to 7.5 ± 0.2 by adding 3N KOH solution, and nitrogen gas (99.999%) was purged into the reactor to maintain the anaerobic condition. Temperature and pH during fermentation were controlled at 35 °C and 5.5 ± 0.2. After fermentation was complete, the mixed solid obtained from the bioreactor was ultrasonicated for 20 min using a horn-type ultrasonic probe with a frequency of 20 kHz (VCX-500-230V-UK, Merck, NJ, USA) for the extraction of CHEEs from microbial cells and centrifuged (FLETA-5, Hanil, Seoul, Republic of Korea) at 2000 rpm for 10 min, and the CHEE-containing supernatant was collected, whereas the solid part was discarded [
18]. The supernatant was immediately filtered using a GF/C filter paper (Whatman, Florham Park, NJ, USA). The total organic acid content of CHEEs was 27,628 mg·COD/L. To prepare the commercial enzymes, four different types of enzymes (namely, α-amylase (BAN 800 MG, 480 KNU-B/g), cellulase (Celluclast BG, 700 EGU/g), lipase (Lipopan 50 BG, 50 KLU/g), and protease (Protamex, 1,5 AU-A/g)) were purchased from Novozymes, Denmark. The working pH and temperature of all commercial enzymes are 6–9 and 30–65 °C.
2.2. Pretreatment and Batch Tests
To investigate the feasibility of CHEEs as pretreatment agents, the CHEEs collected from the fermented FW were added at the amount of 20 mL·CHEE/g·COD of WAS (equivalent to 50 g·COD of WAS/L·CHEE). In addition, some CHEE samples were boiled at 100 °C for 20 min to inactivate the crude enzymes. They were called BCHEE and were added to WAS to clarify the CHEE performance as an enzyme agent. In terms of pretreatment by commercial enzymes, WAS samples were pretreated with a single enzyme (A = α-amylase, P = protease, L = lipase, or C = cellulase) or a cocktail of two or four enzymes. The mixtures of enzymes were prepared using α-amylase + protease (AP) and α-amylase + protease + lipase + cellulase (APCL). The total concentration of each commercial enzyme was controlled at 1 g/g·COD of WAS, and they were then added to WAS. The APCL pretreatment was performed at different concentrations of 0.2, 0.4, 0.8, and 1.6 g/g·COD of WAS to quantitatively evaluate the pretreatment performance of CHEEs. All pretreatments were performed using WAS at a concentration of 50 g·COD/L, pH of 7.0, and temperature of 35 °C, and the samples were stirred at 150 rpm using a shaking incubator (Model SI-300R, HYSC, Seoul, Republic of Korea) for 33 h.
After the enzymatic pretreatment, two sets of anaerobic batch tests were conducted using pretreated WAS to compare the CH4 productivity in response to different pretreatment scenarios. The batch test I was performed to investigate the effect of CHEES and commercial enzymes on CH4 productivity. Batch test II was conducted to evaluate the pretreatment effect of mixed commercial enzymes at different concentrations on CH4 production and quantitatively evaluate the performance of CHEEs on AD compared to CH4 production from the AD of WAS pretreated with commercial enzyme mixtures at different concentrations.
Batch tests were conducted using 250 mL serum bottles. Pretreated WAS, seed sludge, and tap water were added to a serum bottle for a TCOD concentration of 10 g WAS/L and a working volume of 100 mL. The initial pH was adjusted to 7.5 ± 0.2 by adding 3N KOH solution, and nitrogen gas (99.999%) was purged to maintain anaerobic conditions. The samples were stirred at 150 rpm using the shaking incubator (Model SI-300R, HYSC, Seoul, Republic of Korea) for 36 d. During the batch tests, no buffer was added.
2.3. Analytical Methods
For the soluble chemical oxygen demand (SCOD) and organic acid analysis, solid–liquid separation was conducted using a centrifuge (FELTA-5, Hanil, Republic of Korea) and a 0.45 μm membrane filter (Advantec, Durham, NC, USA). The SCOD was measured using closed reflux and colorimetry according to standard methods [
19]. The organic acid was detected using a high-performance liquid chromatograph (HPLC; Ultimate 3000, Thermo Fisher Scientific, Waltham, MA, USA) equipped with a UV–Vis detector (VWD-3400RS) and a 300 mm × 7.8 mm column (Aminex HPX-87H, Bid-rad, Cotati, CA, USA); 0.008 N H
2SO
4 was used as the mobile phase at a flow rate of 0.6 mL/min. The composition of the biogas produced in the batch tests was detected using a gas chromatograph (SRI 310, SRI Instruments, Torrance, CA, USA) equipped with a thermal conductivity detector and a HayeSep T (10 ft × 1/8″) column. N
2 (99.999%) was used as the carrier gas at a flow rate of 10 mL/min.
For the analysis of the microbial community, samples of 50 mL were withdrawn from WAS after the batch test and centrifuged (FLETA-5, Hanil, Seoul, Republic of Korea) at 3000 rpm for 10 min. The settled portion (2 mL) was utilized for DNA extraction with a Fast DNA Spin Kit for soil (QBioGene, Carlsbad, CA, USA) according to the manufacturer’s instructions. The purification of extracted DNA from each sample was further conducted using an UltraClean Microbial DNA Isolation Kit (Mo Bio Laboratories, Carlsbad, CA, USA). Sample amplification was carried out using a GS-FLX Titanium emPCR Kit (454Life Sciences), with a 20 ng aliquot of each sample DNS for a 50 mL PCR reaction. The 16S universal primers of 27F (50 GAGTTTGATCMTGGCTCAG 30) and 518R (50 WTTACCGCGGCTGCTGG 30) were employed to amplify 16S ribosomal RNA (16S rRNA) genes [
20]. A FastStart High Fidelity PCR System (Roche, Basel, Switzerland) was utilized for PCR under the following conditions: 94 °C for 3 min, followed by 35 cycles of 94 °C for 15 s, 55 °C for 45 s, and 72 °C for 1 min, and a final elongation step at 72 °C for 8 min. A purification step was then performed using AMPure beads (Beckman Coulter, Brea, CA, USA). For sequencing, a 454 pyrosequencing genome sequencer FLX Titanium (Roche Applied Science, Penzberg, Germany) was employed according to the manufacturer’s instructions (Macrogen, Seoul, Republic of Korea). The software MOTHUR was used to identify operational taxonomic units (OTUs), assign taxonomy, compare communities, and analyze the sequences. Sequencing errors were reduced by removing sequences with more than one ambiguous base cell and by obtaining sequences that were 300 nt or longer [
21].
2.4. Statistical Analysis
All tests were completed in triplicate, and the experimental results were expressed as mean ± standard deviation. The software Microsoft Office 2018 and SigmaPlot 10.0 were utilized to produce graphs and data.
To calculate CH
4 production, a mass balance was performed based on the headspace measurements of the gas composition and the total biogas volume produced at each time interval. The modified Gompertz equation (Equation (1)) was applied to fit the data of cumulative CH
4, which was used to predict CH
4 production [
22].
where
M(t) is the cumulative CH
4 production (mL) at cultivation time
t (d);
P is the ultimate CH
4 production (mL);
Rm is the gas production rate (mL/d);
λ is the lag period (d); and
e is the exp(1) = 2.71828.
The relationship between CH
4 yield and APCL addition was fitted by Equation (2):
where
is the CH
4 yield;
is the CH
4 yield achieved without APCL addition;
is the APCL amount (g·APCL/g·COD of WAS); and
a and
b are constants.
3. Results and Discussion
3.1. CH4 Production of Pretreated WAS
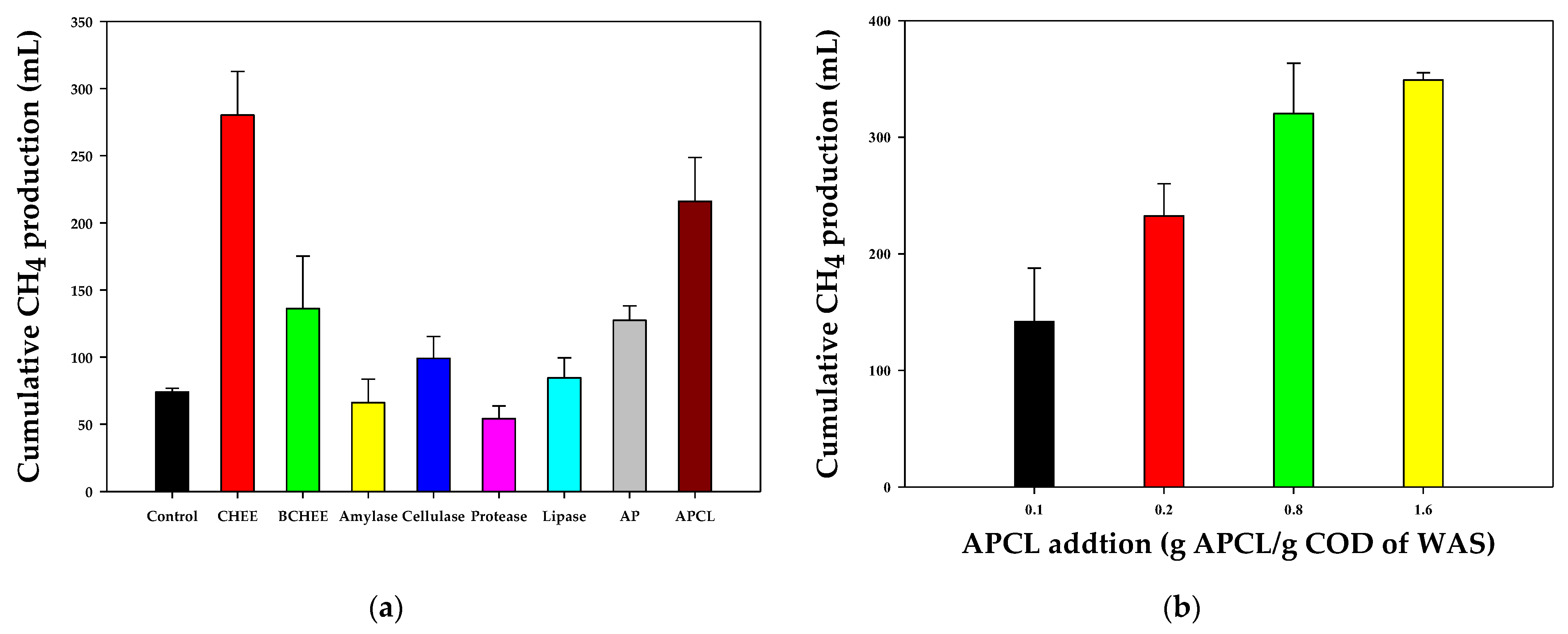
Figure 1a shows the cumulative CH
4 production from WAS under different enzymatic pretreatment scenarios, and
Table 1 summarizes the experimental results of batch test I. The experimental CH
4 production was well fitted by the modified Gompertz equation (R
2 > 0.98), and it clearly demonstrated the effect of the enzymatic pretreatment of WAS on CH
4 production. The final pH values of all samples ranged from 7.3 to 7.4.
Most samples pretreated with commercial enzymes presented higher CH
4 production rates and CH
4 yields than those of the control sample, and their performance varied in response to every single commercial enzyme and different enzyme mixture. The CH
4 production rate and CH
4 yield of samples treated with single commercial enzymes were lower than those of samples treated with mixed commercial enzymes. Cellulase led to the highest CH
4 production rate (65.0 ± 5.0 mL·CH
4/L/day) and CH
4 yield (99.1 ± 18.0 mL·CH
4/g·COD of WAS), which were 120% and 34% higher than those of the control (CH
4 yield of 29.5 ± 0.6 mL·CH
4/L/day; 74.0 ± 2.1 mL·CH
4/g·COD of WAS), respectively. Moreover, protease-pretreated samples harmed CH
4 production, with a CH
4 production rate of 23.5 ± 2.4 mL·CH
4/L/day and CH
4 yield of 54.1 ± 8.8 mL·CH
4/g·COD of WAS. Protease is known to be a very effective enzymatic agent. However, toxic compounds such as ammonia formed from the degradation of proteins by protease often provoke the accumulation of organic acids and the inhibition of microbial metabolism when organic compounds that have a high content of protein are used as feedstock for pretreatment [
23]. The CH
4 yield of amylase-added samples was also low, at 66.1 ± 20.1 mL·CH
4/g·COD of WAS.
Higher enzyme diversity led to higher CH
4 production from WAS. That is, compared to the control, WAS pretreated with a mixture of two enzymes (amylase + protease) led to a gradual increase in both the CH
4 production rate and CH
4 yield, which reached 73.5 mL·CH
4/L/day and 127.3 mL·CH
4/g·COD of WAS, respectively. Moreover, the mixture of four enzymes (amylase + protease + cellulase + lipase) presented the highest CH
4 production rate of 159.5 mL·CH
4/L/day and a CH
4 yield of 216.0 mL·CH
4/g·COD of WAS. This increase was closely related to the chemical composition of WAS cell walls, which were mainly composed of heterogeneous organic components mixed with lipids, proteins, carbohydrates, and cellulose. The multiple hydrolytic enzymes could simultaneously react for the effective hydrolysis of WAS, and thus, more CH
4 was recovered. This result is also in agreement with a previous study reporting that mixed enzymes are more effective for the hydrolysis of organic wastes [
24].
The WAS prepared with CHEEs led to a CH4 production rate of 192.0 ± 4.5 mL·CH4/L/day and a CH4 yield of 211.9 ± 16.4 mL·CH4/g·COD of WAS, which was comparable to the performance of the sample pretreated with the mixture of four commercial enzymes. Based on this result, we speculated that the increased CH4 productivity of CHEEs was derived not only from the direct conversion to CH4 from the soluble organic carbon in the CHEEs but also from the hydrolysis of WAS by crude enzymes extracted from FW. To further clarify the reason for the increased CH4 production and to distinguish the CH4 origin, BCHEEs (which were obtained by boiling CHEEs at 100 °C for 20 min to inactivate crude enzymes) were added to WAS, and further batch tests were conducted. The use of BCHEEs led to a CH4 yield of 136.1 ± 22.6 mL·CH4/g·COD of WAS and a CH4 production rate of 62.0 ± 6.3 mL·CH4/L/day, which were 84% and 110% higher than those of the control. Based on the CH4 yields obtained using the control, CHEEs, and BCHEEs, we estimated the amount of CH4 produced in response to the application of the enzymatic pretreatment using CHEEs (=CH4 CHEE − CH4 BCHEE), which was 75.8 mL·CH4/g·COD of WAS, whereas 62.1 mL·CH4/g·COD of WAS was obtained from the organic carbon of CHEEs (=CH4 BCHEE − CH4 Control).
3.2. Quantitative Evaluation of CHEE Performance
The results of batch test I indicated that CHEEs presented superior characteristics compared to commercial enzymes. CHEEs included a variety of enzymes and worked synergistically to degrade the complex cell walls in WAS and achieved comparable CH4 production to that of APCL, which was a mix of four commercial enzymes (amylase + protease + cellulase + lipase). It was then necessary to determine the mechanisms through which the CHEE enzymatic activity improved CH4 production as compared to the mixture of commercial enzymes. Therefore, we quantitatively evaluated the enzymatic performance of CHEEs by performing batch test II for WAS prepared with APCL at different concentrations (0–1.6 g/g·COD of WAS).
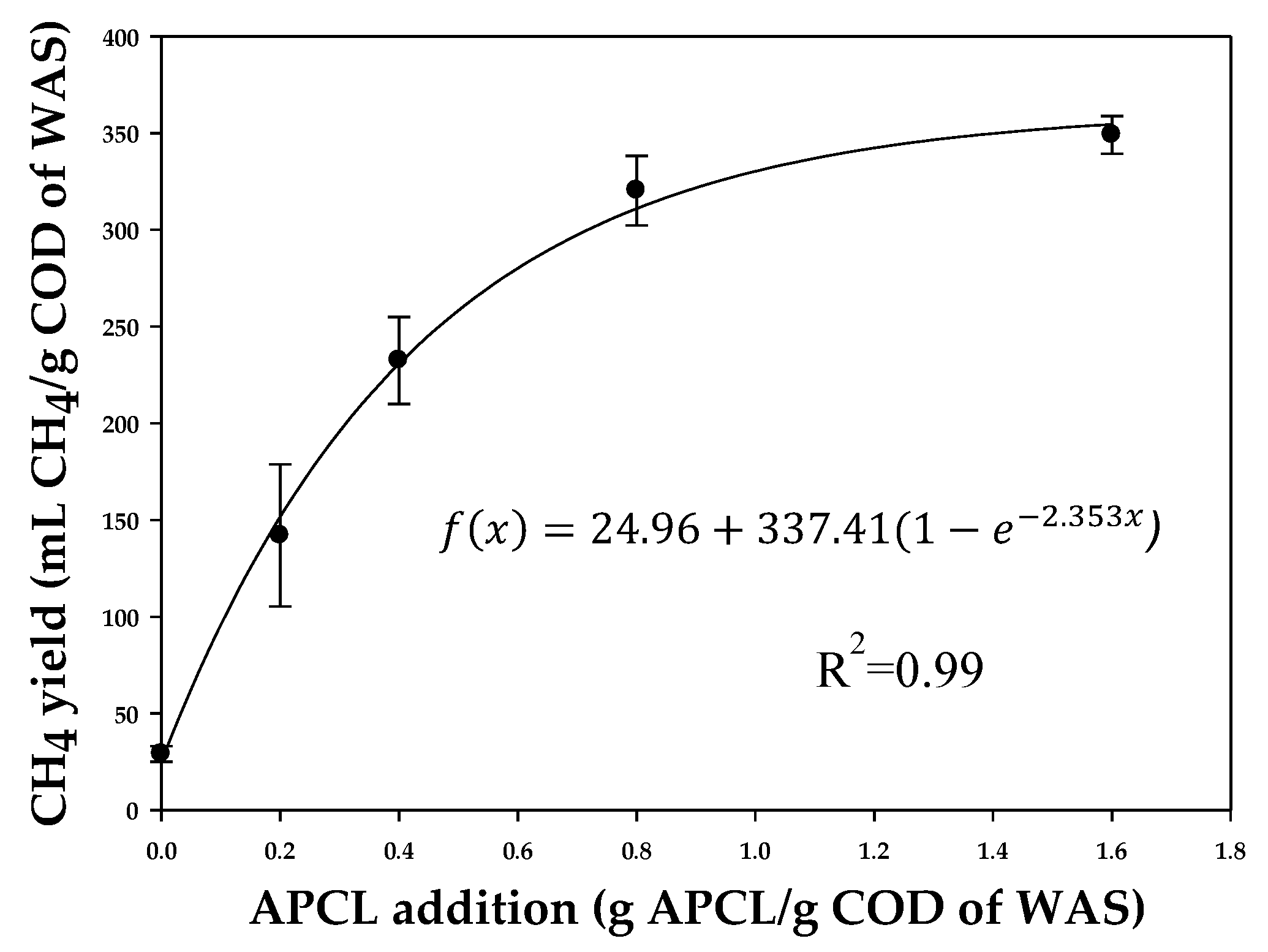
The CH
4 yield of batch test II in response to the WAS pretreatment at different APCL concentrations is shown in
Figure 1b. Overall, higher APCL concentrations led to higher biogas production. For instance, the addition of APCL at 0.4 and 1.6 g·APCL/g·COD of WAS led to CH
4 yields of 232.5 ± 27.6 and 349.1 ± 6.3 mL CH
4/g COD of WAS, respectively. The CH
4 yields at different APCL concentrations were further fitted by Equation (2), and the R
2 values of both curves were over 0.99 (
Figure 2). The results show that the CH
4 yield of WAS pretreated with CHEEs was similar to the CH
4 yield of 0.34 g·APCL/g·COD of WAS.
3.3. Microbial Community and Comprehensive Discussion
The microbial samples collected from batch test I were further analyzed by 16S rRNA gene-targeted next-generation sequencing (NGS) to reveal the variation in CH
4 production in response to the pretreatment according to microbial ecology.
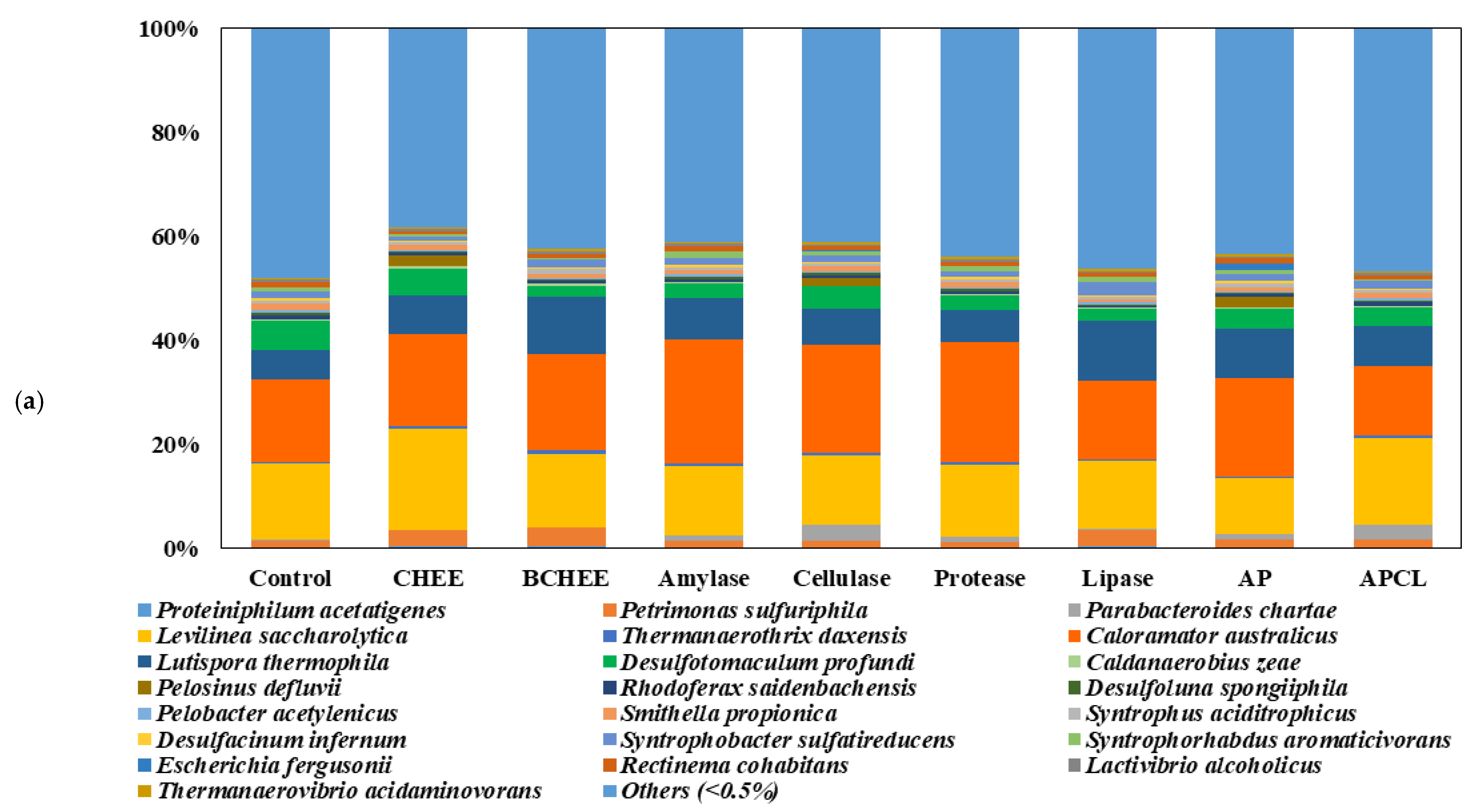
Figure 3a shows apparent changes in the diversity of both bacteria and archaea at the species level. The minor microbial groups, which accounted for less than 0.5%, are included in the category “others.” The bacterial community involved in the hydrolysis and acidogenesis included four main members:
Caloramator australicus (13.36–23.78%),
Levilinea saccharolytica (10.53–19.31%),
Lutispora thermophile (5.50–11.61%), and
Desulfotomaculum profundi (2.10–5.76%). During the batch test fed by non-pretreated WAS, the species
Caloramator australicus (15.9%) and
Levilinea saccharolytica (14.5%) were the most prevalent, followed by
Desulfotomaculum profundi (5.8%) and
Lutispora thermophile (5.5%).
Levilinea saccharolytica and
Caloramator australicus produce acetate from various hydrocarbons [
25]. The share of
Caloramator australicus and
Lutispora thermophile increased when the pretreatments were employed.
Caloramator australicus in the samples pretreated with single amylase, cellulase, and protease represented 23.8%, 20.8%, and 23.2% of the total number of sequences, respectively. The abundance of
Lutispora thermophile increased to 6.1–11.6% of the total number of sequences. Moreover, the abundance of
Levilinea saccharolytica in the samples prepared with CHEEs and APCL increased to 19.3% and 16.5%, respectively, but its share in other samples decreased.
Lutispora thermophile is a fermentative bacterium, reported as the main bacterium during acidogenesis, and its primary end-products from proteinaceous compounds are acetate, iso-butyrate, propionate, and iso-valerate [
26,
27]. It is frequently observed in high abundance during the AD of municipal solid waste. Based on the above results, we attributed the increase in acidogenesis in response to improved WAS hydrolysis by the mixture of enzymes to the increased abundance of
Lutispora thermophile in both CHEE and APCL samples.
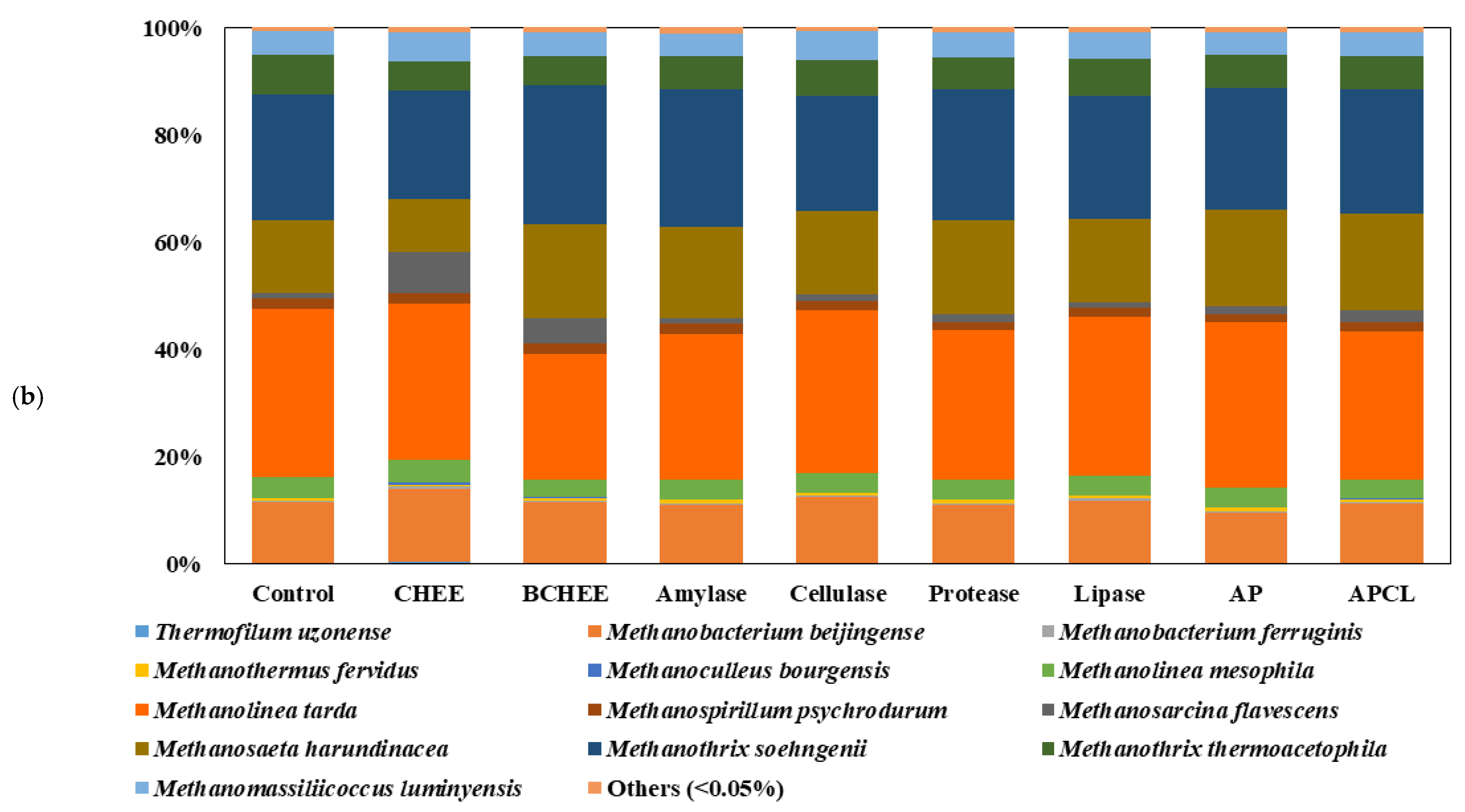
The investigation of the archaeal community conducted in this study was significant because this community is crucial for CH
4 conversion from the end-products of acidogenic bacteria [
28].
Figure 3b shows clear changes in the diversity of the main archaeal species.
Methanolinea tarda,
Methanothrix soehngenii,
Methanosaeta harundinacea, and
Methanobacterium beijingense were the dominant archaea in the samples without pretreatment (control), accounting for 31.4%, 23.5%, 13.5%, and 11.3% of the total archaeal sequences, respectively. However, their populations decreased, and a new bacterial community was developed when CHEEs and BCHEEs were used to pretreat WAS. For example, the abundance of
Methanosarcina flavescens increased, accounting for 7.6% and 4.7% of the total in the samples treated with CHEEs and BCHEEs, respectively.
Methanosarcina spp. uses both acetoclastic and hydrogenotrophic pathways for CH
4 production in the presence of a high concentration of organic acids.
Methanosarcina spp. are also characterized by the utilization of various substrates with a high substrate utilization rate [
29,
30].
Methanosaeta spp. are usually the dominant acetoclastic methanogens when acetate concentrations are low [
31,
32]. Regarding the characteristics of CHEEs and BCHEEs containing a high concentration of organic acids (27,628 mg·COD/L), the increased dominance of
Methanosarcina spp. in the samples was a noteworthy phenomenon that supported the results of enhanced CH
4 production.
This study clearly shows the high activity of CHEEs produced from FW fermentation operated using a mixed culture and their positive effect on CH4 productivity from complex organic polymers. CHEEs present advantages regarding pretreatment conditions, such as pH and temperature. CHEEs were produced under high alkalinity during acid fermentation, and they could self-control the pH of WAS without the addition of external chemicals during the pretreatment. In contrast, a buffer is always needed to control the pH to sustain enzymatic activity when commercial enzymes are used. Furthermore, the CHEEs extracted from FW mesophilic fermentation were stable and showed great catalytic activity in the batch tests for CH4 production. The dominant microbial community in this study presented an optimal growth temperature of approximately 25–40 °C. This study determined the manner through which CHEEs exerted enzymatic activity that improved CH4 production from WAS compared to APCL pretreatment. The CHEEs cultivated in this study were not optimal, and CHEE activity can easily change depending on the operating conditions of fermentation. Therefore, this study clearly shows the high activity of crude enzymes produced from FW fermentation operated using a mixed culture and the positive effect of crude enzymes on the CH4 productivity of complex organic polymers. Thus, further investigation on the optimization of CHEE production conditions to maximize catalytic activity should be conducted.







{kind=link}
{kind=link}
{kind=link}
{kind=link}