The Gut Microbiome from a Biomarker to a Novel Therapeutic Strategy for Immunotherapy Response in Patients with Lung Cancer


Abstract
:1. Introduction
2. Microbiota Profiling of Patients with NSCLC
2.1. Gut Microbiome in NSCLC Patients
2.2. Lung Microbiome in Lung Cancer Patients
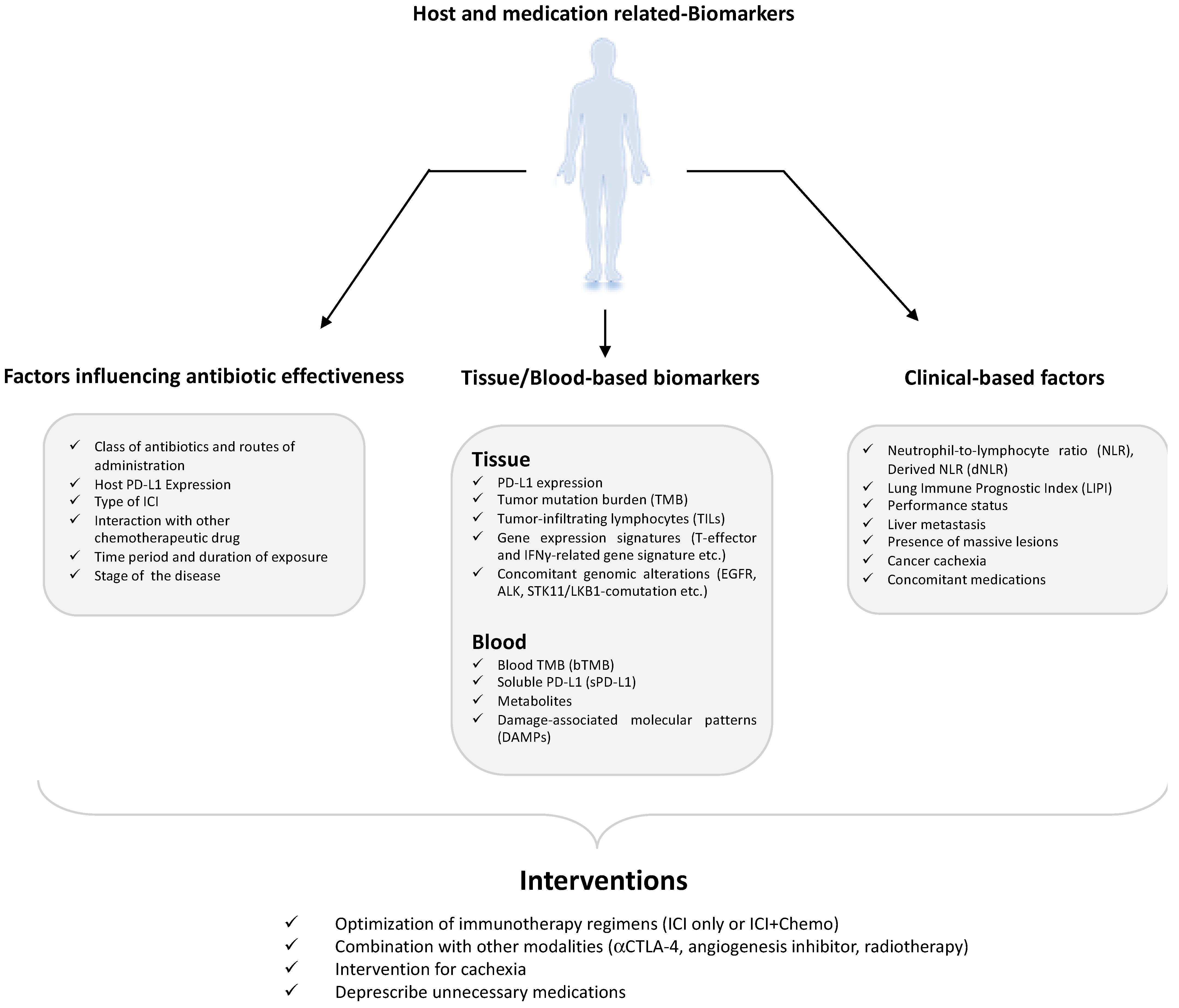
2.3. Impact of Concurrent Medications on ICI Responses in Cancer
3. Modulation of the Gut Microbiome to Improve the Efficacy of Anti-PD-1/PD-L1
3.1. Fecal Microbiota Transplantation
3.2. Probiotics
3.3. Diet Evaluation and Prebiotics
3.4. Other Techniques
3.5. Discussion and Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ALK | Anaplastic lymphoma kinase |
| ATB | Antibiotics |
| BAL | Bronchoalveolar lavage |
| Bcl-2 | B-cell lymphoma 2 |
| Bcl-XL | B-cell lymphoma-extra large |
| CagA | Cytotoxin-associated gene |
| CD8 | Cluster of differentiation 8 |
| CRC | Colorectal cancer |
| CTLA-4 | Cytotoxic T-lymphocyte-associated antigen 4 |
| DNA | Deoxyribonucleic acid |
| DOR | Duration of response |
| EGFR | Epidermal growth factor receptor |
| EMA | European Medicine Agency |
| ERK | Extracellular signal-regulated kinase |
| FFQ | Food Frequency Questionnaire |
| FMT | Fecal Microbiota Therapy |
| FOXP3 | Forkhead box P3 protein |
| GF | Germ-free |
| GM | Gut microbiome |
| IBD | Inflammatory bowel disease |
| ICIs | Immune checkpoint inhibitors |
| IHC | Immunohistochemistry |
| irAEs | Immune-related adverse effects |
| mAb | Monoclonal antibody |
| MDSC | Myeloid-derived suppressor cells |
| MET | Microbial ecosystem therapeutics |
| MHC | Major histocompatibility complex |
| NGS | Next-generation sequencing |
| NK | Natural killer cells |
| NSCLC | Non-small cell lung cancer |
| ORR | Objective response rate |
| OS | Overall survival |
| PD-1 | Programmed cell death protein 1 |
| PD-L1 | Programmed cell death ligand 1 |
| PFS | Progression-free survival |
| PI3K | Phosphoinositide 3-kinase |
| PPI | Proton pump inhibitor |
| QoL | Quality of life |
| RCC | Renal cell carcinoma |
| rCDI | Refractory Clostridium difficile infection |
| rDNA | Ribosomal deoxyribonucleic acid |
| RECIST | Response evaluation criteria in solid tumors |
| RMT | Oral restorative microbiota therapy |
| rRNA | Ribosomal ribonucleic acid |
| SCLC | Small cell lung cancer |
| SPF | Specific pathogen free |
| TLR | Toll-like receptor |
| TKI | Tyrosine kinase inhibitor |
| TP53 | Tumor protein 53 |
| USFDA | United States Food and Drug Administration |
| VEGF | Vascular endothelial growth factor |
References
- Beatty, G.L.; Gladney, W.L. Immune Escape Mechanisms as a Guide for Cancer Immunotherapy. Clin. Cancer Res. 2015, 21, 687–692. [Google Scholar] [CrossRef]
- Reck, M.; Rodríguez-Abreu, D.; Robinson, A.G.; Hui, R.; Csőszi, T.; Fülöp, A.; Gottfried, M.; Peled, N.; Tafreshi, A.; Cuffe, S.; et al. Faculty Opinions recommendation of Pembrolizumab versus Chemotherapy for PD-L1-Positive Non-Small-Cell Lung Cancer. N. Engl. J. Med. 2016, 375, 1823–1833. [Google Scholar] [CrossRef] [PubMed]
- Garassino, M.C.; Gadgeel, S.; Speranza, G.; Felip, E.; Esteban, E.; Dómine, M.; Hochmair, M.J.; Powell, S.F.; Bischoff, H.G.; Peled, N.; et al. Pembrolizumab Plus Pemetrexed and Platinum in Nonsquamous Non–Small-Cell Lung Cancer: 5-Year Outcomes From the Phase 3 KEYNOTE-189 Study. J. Clin. Oncol. 2023, 41, 1992–1998. [Google Scholar] [CrossRef] [PubMed]
- Gandhi, L.; Rodríguez-Abreu, D.; Gadgeel, S.; Esteban, E.; Felip, E.; De Angelis, F.; Domine, M.; Clingan, P.; Hochmair, M.J.; Powell, S.F.; et al. Pembrolizumab plus Chemotherapy in Metastatic Non–Small-Cell Lung Cancer. N. Engl. J. Med. 2018, 378, 2078–2092. [Google Scholar] [CrossRef] [PubMed]
- Lurienne, L.; Cervesi, J.; Duhalde, L.; de Gunzburg, J.; Andremont, A.; Zalcman, G.; Buffet, R.; Bandinelli, P.-A. NSCLC Immunotherapy Efficacy and Antibiotic Use: A Systematic Review and Meta-Analysis. J. Thorac. Oncol. 2020, 15, 1147–1159. [Google Scholar] [CrossRef]
- Paz-Ares, L.; Ciuleanu, T.-E.; Cobo, M.; Schenker, M.; Zurawski, B.; Menezes, J.; Richardet, E.; Bennouna, J.; Felip, E.; Juan-Vidal, O.; et al. First-line nivolumab plus ipilimumab combined with two cycles of chemotherapy in patients with non-small-cell lung cancer (CheckMate 9LA): An international, randomised, open-label, phase 3 trial. Lancet Oncol. 2021, 22, 198–211. [Google Scholar] [CrossRef] [PubMed]
- Elkrief, A.; Derosa, L.; Kroemer, G.; Zitvogel, L.; Routy, B. The negative impact of antibiotics on outcomes in cancer patients treated with immunotherapy: A new independent prognostic factor? Ann. Oncol. 2019, 30, 1572–1579. [Google Scholar] [CrossRef] [PubMed]
- Forde, P.M.; Spicer, J.; Lu, S.; Provencio, M.; Mitsudomi, T.; Awad, M.M.; Felip, E.; Broderick, S.R.; Brahmer, J.R.; Swanson, S.J.; et al. Neoadjuvant Nivolumab plus Chemotherapy in Resectable Lung Cancer. N. Engl. J. Med. 2022, 386, 1973–1985. [Google Scholar] [CrossRef] [PubMed]
- Esfahani, K.; Meti, N.; Miller, W.H.; Hudson, M. Adverse events associated with immune checkpoint inhibitor treatment for cancer. Can. Med Assoc. J. 2019, 191, E40–E46. [Google Scholar] [CrossRef]
- National Institutes of Health (NIH). NIH Human Microbiome Project Defines Normal Bacterial Makeup of the Body. 2015. Available online: https://www.nih.gov/news-events/news-releases/nih-human-microbiome-project-defines-normal-bacterial-makeup-body (accessed on 11 March 2023).
- Rowland, I.; Gibson, G.; Heinken, A.; Scott, K.; Swann, J.; Thiele, I.; Tuohy, K. Gut microbiota functions: Metabolism of nutrients and other food components. Eur. J. Nutr. 2018, 57, 1–24. [Google Scholar] [CrossRef]
- Shreiner, A.B.; Kao, J.Y.; Young, V.B. The gut microbiome in health and in disease. Curr. Opin. Gastroenterol. 2015, 31, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Valitutti, F.; Cucchiara, S.; Fasano, A. Celiac Disease and the Microbiome. Nutrients 2019, 11, 2403. [Google Scholar] [CrossRef]
- Davis, C.D. The Gut Microbiome and Its Role in Obesity. Nutr. Today 2016, 51, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Shi, X.; Yang, J.; Zhao, Y.; Xue, L.; Xu, L.; Cai, J. Gut microbes in cardiovascular diseases and their potential therapeutic applications. Protein Cell 2020, 12, 346–359. [Google Scholar] [CrossRef]
- Zavros, Y.; Merchant, J.L. The immune microenvironment in gastric adenocarcinoma. Nat. Rev. Gastroenterol. Hepatol. 2022, 19, 451–467. [Google Scholar] [CrossRef] [PubMed]
- Garrett, W.S. Cancer and the microbiota. Science 2015, 348, 80–86. [Google Scholar] [CrossRef]
- de Martel, C.; Ferlay, J.; Franceschi, S.; Vignat, J.; Bray, F.; Forman, D.; Plummer, M. Global burden of cancers attributable to infections in 2008: A review and synthetic analysis. Lancet Oncol. 2012, 13, 607–615. [Google Scholar] [CrossRef]
- Hsiao, Y.-C.; Liu, C.-W.; Yang, Y.; Feng, J.; Zhao, H.; Lu, K. DNA Damage and the Gut Microbiome: From Mechanisms to Disease Outcomes. Dna 2023, 3, 13–32. [Google Scholar] [CrossRef]
- Asokan, S.; Cullin, N.; Stein-Thoeringer, C.K.; Elinav, E. CAR-T Cell Therapy and the Gut Microbiota. Cancers 2023, 15, 794. [Google Scholar] [CrossRef]
- Derosa, L.; Routy, B.; Desilets, A.; Daillère, R.; Terrisse, S.; Kroemer, G.; Zitvogel, L. Microbiota-Centered Interventions: The Next Breakthrough in Immuno-Oncology? Cancer Discov. 2021, 11, 2396–2412. [Google Scholar] [CrossRef] [PubMed]
- Routy, B.; Le Chatelier, E.; Derosa, L.; Duong, C.P.; Alou, M.T.; Daillère, R.; Fluckiger, A.; Messaoudene, M.; Rauber, C.; Roberti, M.P.; et al. Gut microbiome influences efficacy of PD-1-based immunotherapy against epithelial tumors. Science 2018, 359, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Gopalakrishnan, V.; Spencer, C.N.; Nezi, L.; Reuben, A.; Andrews, M.C.; Karpinets, T.V.; Prieto, P.A.; Vicente, D.; Hoffman, K.; Wei, S.C.; et al. Gut microbiome modulates response to anti–PD-1 immunotherapy in melanoma patients. Science 2018, 359, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Matson, V.; Fessler, J.; Bao, R.; Chongsuwat, T.; Zha, Y.; Alegre, M.-L.; Luke, J.J.; Gajewski, T.F. The commensal microbiome is associated with anti–PD-1 efficacy in metastatic melanoma patients. Science 2018, 359, 104–108. [Google Scholar] [CrossRef]
- Routy, B.; Gopalakrishnan, V.; Daillère, R.; Zitvogel, L.; Wargo, J.A.; Kroemer, G. The gut microbiota influences anticancer immunosurveillance and general health. Nat. Rev. Clin. Oncol. 2018, 15, 382–396. [Google Scholar] [CrossRef]
- Vétizou, M.; Pitt, J.M.; Daillère, R.; Lepage, P.; Waldschmitt, N.; Flament, C.; Rusakiewicz, S.; Routy, B.; Roberti, M.P.; Duong, C.P.; et al. Anticancer immunotherapy by CTLA-4 blockade relies on the gut microbiota. Science 2015, 350, 1079–1084. [Google Scholar] [CrossRef] [PubMed]
- Fluckiger, A.; Daillère, R.; Sassi, M.; Sixt, B.S.; Liu, P.; Loos, F.; Richard, C.; Rabu, C.; Alou, M.T.; Goubet, A.-G.; et al. Cross-reactivity between tumor MHC class I–restricted antigens and an enterococcal bacteriophage. Science 2020, 369, 936–942. [Google Scholar] [CrossRef]
- Hayase, E.; Jenq, R.R. Role of the intestinal microbiome and microbial-derived metabolites in immune checkpoint blockade immunotherapy of cancer. Genome Med. 2021, 13, 107. [Google Scholar] [CrossRef]
- Routy, B.; Lenehan, J.G.; Miller, W.H.; Jamal, R.; Messaoudene, M.; Daisley, B.A.; Hes, C.; Al, K.F.; Martinez-Gili, L.; Punčochář, M.; et al. Fecal microbiota transplantation plus anti-PD-1 immunotherapy in advanced melanoma: A phase I trial. Nat. Med. 2023, 29, 2121–2132. [Google Scholar] [CrossRef] [PubMed]
- Davar, D.; Dzutsev, A.K.; McCulloch, J.A.; Rodrigues, R.R.; Chauvin, J.-M.; Morrison, R.M.; Deblasio, R.N.; Menna, C.; Ding, Q.; Pagliano, O.; et al. Fecal microbiota transplant overcomes resistance to anti–PD-1 therapy in melanoma patients. Science 2021, 371, 595–602. [Google Scholar] [CrossRef]
- Hanahan, D. Hallmarks of Cancer: New Dimensions. Cancer Discov. 2022, 12, 31–46. [Google Scholar] [CrossRef]
- Derosa, L.; Routy, B.; Zitvogel, L.; Thomas, A.M.; Zalcman, G.; Friard, S.; Mazieres, J.; Audigier-Valette, C.; Moro-Sibilot, D.; Goldwasser, F.; et al. Intestinal Akkermansia muciniphila predicts overall survival in advanced non-small cell lung cancer patients treated with anti-PD-1 antibodies: Results a phase II study. J. Clin. Oncol. 2021, 39, 9019. [Google Scholar] [CrossRef]
- Derosa, L.; Routy, B.; Thomas, A.M.; Iebba, V.; Zalcman, G.; Friard, S.; Mazieres, J.; Audigier-Valette, C.; Moro-Sibilot, D.; Goldwasser, F.; et al. Intestinal Akkermansia muciniphila predicts clinical response to PD-1 blockade in patients with advanced non-small-cell lung cancer. Nat. Med. 2022, 28, 315–324. [Google Scholar] [CrossRef]
- Grenda, A.; Iwan, E.; Chmielewska, I.; Krawczyk, P.; Giza, A.; Bomba, A.; Frąk, M.; Rolska, A.; Szczyrek, M.; Kieszko, R.; et al. Presence of Akkermansiaceae in gut microbiome and immunotherapy effectiveness in patients with advanced non-small cell lung cancer. AMB Express 2022, 12, 86. [Google Scholar] [CrossRef]
- Liu, L.; Xie, W.; Xue, P.; Wei, Z.; Liang, X.; Chen, N. Diagnostic accuracy and prognostic applications of CYFRA 21-1 in head and neck cancer: A systematic review and meta-analysis. PLoS ONE 2019, 14, e0216561. [Google Scholar] [CrossRef]
- Newsome, R.C.; Gharaibeh, R.Z.; Pierce, C.M.; da Silva, W.V.; Paul, S.; Hogue, S.R.; Yu, Q.; Antonia, S.; Conejo-Garcia, J.R.; Robinson, L.A.; et al. Interaction of bacterial genera associated with therapeutic response to immune checkpoint PD-1 blockade in a United States cohort. Genome Med. 2022, 14, 35. [Google Scholar] [CrossRef] [PubMed]
- Dora, D.; Ligeti, B.; Kovacs, T.; Revisnyei, P.; Galffy, G.; Dulka, E.; Krizsán, D.; Kalcsevszki, R.; Megyesfalvi, Z.; Dome, B.; et al. Non-Small Cell Lung Cancer Patients Treated with Anti-PD1 Immunotherapy Show Distinct Microbial Signatures and Metabolic Pathways According to Clinical Outcomes. medRxiv 2022. [Google Scholar] [CrossRef]
- Martini, G.; Ciardiello, D.; Dallio, M.; Famiglietti, V.; Esposito, L.; Della Corte, C.M.; Napolitano, S.; Fasano, M.; Gravina, A.G.; Romano, M.; et al. Gut microbiota correlates with antitumor activity in patients with mCRC and NSCLC treated with cetuximab plus avelumab. Int. J. Cancer 2022, 151, 473–480. [Google Scholar] [CrossRef] [PubMed]
- Katayama, Y.; Yamada, T.; Tanimura, K.; Yoshimura, A.; Takeda, T.; Chihara, Y.; Tamiya, N.; Kaneko, Y.; Uchino, J.; Takayama, K. Impact of bowel movement condition on immune checkpoint inhibitor efficacy in patients with advanced non-small cell lung cancer. Thorac. Cancer 2019, 10, 526–532. [Google Scholar] [CrossRef] [PubMed]
- Hakozaki, T.; Richard, C.; Elkrief, A.; Hosomi, Y.; Benlaïfaoui, M.; Mimpen, I.; Terrisse, S.; Derosa, L.; Zitvogel, L.; Routy, B.; et al. The Gut Microbiome Associates with Immune Checkpoint Inhibition Outcomes in Patients with Advanced Non-Small Cell Lung Cancer. Cancer Immunol. Res. 2020, 8, 1243–1250. [Google Scholar] [CrossRef]
- Shoji, F.; Yamaguchi, M.; Okamoto, M.; Takamori, S.; Yamazaki, K.; Okamoto, T.; Maehara, Y. Gut microbiota diversity and specific composition during immunotherapy in responders with non-small cell lung cancer. Front. Mol. Biosci. 2022, 9, 1040424. [Google Scholar] [CrossRef] [PubMed]
- Krief, J.O.; de Tauriers, P.H.; Dumenil, C.; Neveux, N.; Dumoulin, J.; Giraud, V.; Labrune, S.; Tisserand, J.; Julie, C.; Emile, J.-F.; et al. Role of antibiotic use, plasma citrulline and blood microbiome in advanced non-small cell lung cancer patients treated with nivolumab. J. Immunother. Cancer 2019, 7, 176. [Google Scholar] [CrossRef]
- Jin, Y.; Dong, H.; Xia, L.; Yang, Y.; Zhu, Y.; Shen, Y.; Zheng, H.; Yao, C.; Wang, Y.; Lu, S. The Diversity of Gut Microbiome is Associated With Favorable Responses to Anti–Programmed Death 1 Immunotherapy in Chinese Patients with NSCLC. J. Thorac. Oncol. 2019, 14, 1378–1389. [Google Scholar] [CrossRef]
- Song, P.; Yang, D.; Wang, H.; Cui, X.; Si, X.; Zhang, X.; Zhang, L. Relationship between intestinal flora structure and metabolite analysis and immunotherapy efficacy in Chinese NSCLC patients. Thorac. Cancer 2020, 11, 1621–1632. [Google Scholar] [CrossRef] [PubMed]
- He, D.; Li, X.; An, R.; Wang, L.; Wang, Y.; Zheng, S.; Chen, X.; Wang, X. Response to PD-1-Based Immunotherapy for Non-Small Cell Lung Cancer Altered by Gut Microbiota. Oncol. Ther. 2021, 9, 647–657. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Wang, J.; Sun, Z.; Cao, Y.; Mu, Z.; Ji, X. Commensal microbiota contributes to predicting the response to immune checkpoint inhibitors in non-small-cell lung cancer patients. Cancer Sci. 2021, 112, 3005–3017. [Google Scholar] [CrossRef]
- Masuhiro, K.; Tamiya, M.; Fujimoto, K.; Koyama, S.; Naito, Y.; Osa, A.; Hirai, T.; Suzuki, H.; Okamoto, N.; Shiroyama, T.; et al. Brochoalveolar lavage fluid reveals factors contributing to the efficacy of PD-1 blockade in lung cancer. J. Clin. Investig. 2022, 7, e157915. [Google Scholar] [CrossRef]
- McLean, A.E.; Kao, S.C.; Barnes, D.J.; Wong, K.K.; Scolyer, R.A.; Cooper, W.A.; Kohonen-Corish, M.R. The emerging role of the lung microbiome and its importance in non-small cell lung cancer diagnosis and treatment. Lung Cancer 2022, 165, 124–132. [Google Scholar] [CrossRef]
- Lee, S.H.; Sung, J.Y.; Yong, D.; Chun, J.; Kim, S.Y.; Song, J.H.; Chung, K.S.; Kim, E.Y.; Jung, J.Y.; Kang, Y.A.; et al. Characterization of microbiome in bronchoalveolar lavage fluid of patients with lung cancer comparing with benign mass like lesions. Lung Cancer 2016, 102, 89–95. [Google Scholar] [CrossRef]
- Liu, H.-X.; Tao, L.-L.; Zhang, J.; Zhu, Y.-G.; Zheng, Y.; Liu, D.; Zhou, M.; Ke, H.; Shi, M.-M.; Qu, J.-M. Difference of lower airway microbiome in bilateral protected specimen brush between lung cancer patients with unilateral lobar masses and control subjects. Int. J. Cancer 2017, 142, 769–778. [Google Scholar] [CrossRef] [PubMed]
- Tsay, J.-C.J.; Wu, B.G.; Badri, M.H.; Clemente, J.C.; Shen, N.; Meyn, P.; Li, Y.; Yie, T.-A.; Lhakhang, T.; Olsen, E.; et al. Airway microbiota is associated with upregulation of the PI3K pathway in lung cancer. Am. J. Respir. Crit. Care Med. 2018, 198, 1188–1198. [Google Scholar] [CrossRef] [PubMed]
- Tsay, J.-C.J.; Wu, B.G.; Sulaiman, I.; Gershner, K.; Schluger, R.; Li, Y.; Yie, T.-A.; Meyn, P.; Olsen, E.; Perez, L.; et al. Lower Airway Dysbiosis Affects Lung Cancer Progression. Cancer Discov. 2021, 11, 293–307. [Google Scholar] [CrossRef] [PubMed]
- Greathouse, K.L.; White, J.R.; Vargas, A.J.; Bliskovsky, V.V.; Beck, J.A.; von Muhlinen, N.; Polley, E.C.; Bowman, E.D.; Khan, M.A.; Robles, A.I.; et al. Interaction between the microbiome and TP53 in human lung cancer. Genome Biol. 2018, 19, 123. [Google Scholar] [CrossRef]
- Boesch, M.; Baty, F.; Albrich, W.C.; Flatz, L.; Rodriguez, R.; Rothschild, S.I.; Joerger, M.; Früh, M.; Brutsche, M.H. Local tumor microbial signatures and response to checkpoint blockade in non-small cell lung cancer. OncoImmunology 2021, 10, 1988403. [Google Scholar] [CrossRef]
- Patangia, D.V.; Ryan, C.A.; Dempsey, E.; Ross, R.P.; Stanton, C. Impact of antibiotics on the human microbiome and consequences for host health. Microbiologyopen 2022, 11, e1260. [Google Scholar] [CrossRef] [PubMed]
- Modi, S.R.; Collins, J.J.; Relman, D.A. Antibiotics and the gut microbiota. J. Clin. Investig. 2014, 124, 4212–4218. [Google Scholar] [CrossRef] [PubMed]
- Huemer, F.; Rinnerthaler, G.; Westphal, T.; Hackl, H.; Hutarew, G.; Gampenrieder, S.P.; Weiss, L.; Greil, R. Impact of antibiotic treatment on immune-checkpoint blockade efficacy in advanced non-squamous non-small cell lung cancer. Oncotarget 2018, 9, 16512–16520. [Google Scholar] [CrossRef]
- Derosa, L.; Hellmann, M.D.; Spaziano, M.; Halpenny, D.; Fidelle, M.; Rizvi, H.; Long, N.; Plodkowski, A.J.; Arbour, K.C.; Chaft, J.E.; et al. Negative association of antibiotics on clinical activity of immune checkpoint inhibitors in patients with advanced renal cell and non-small-cell lung cancer. Ann. Oncol. 2018, 29, 1437–1444. [Google Scholar] [CrossRef]
- Cortellini, A.; Cannita, K.; Tiseo, M.; Cortinovis, D.L.; Aerts, J.G.; Baldessari, C.; Giusti, R.; Ferrara, M.G.; D’Argento, E.; Grossi, F.; et al. Post-progression outcomes of NSCLC patients with PD-L1 expression ≥ 50% receiving first-line single-agent pembrolizumab in a large multicentre real-world study. Eur. J. Cancer 2021, 148, 24–35. [Google Scholar] [CrossRef] [PubMed]
- Mei, T.; Gong, Y. EP05.01-005 Impact of Antibiotic Use Before Definitive Concurrent Chemoradiation in Patients with Locally Advanced Non Small Cell Lung Cancer. J. Thorac. Oncol. 2022, 17, S267. [Google Scholar] [CrossRef]
- Fidelle, M.; Rauber, C.; Silva, C.A.C.; Tian, A.-L.; Lahmar, I.; de La Varende, A.-L.M.; Zhao, L.; Thelemaque, C.; Lebhar, I.; Messaoudene, M.; et al. A microbiota-modulated checkpoint directs immunosuppressive intestinal T cells into cancers. Science 2023, 380, eabo2296. [Google Scholar] [CrossRef]
- Suez, J.; Zmora, N.; Zilberman-Schapira, G.; Mor, U.; Dori-Bachash, M.; Bashiardes, S.; Zur, M.; Regev-Lehavi, D.; Brik, R.B.Z.; Federici, S.; et al. Post-Antibiotic Gut Mucosal Microbiome Reconstitution Is Impaired by Probiotics and Improved by Autologous FMT. Cell 2018, 174, 1406–1423.e1. [Google Scholar] [CrossRef] [PubMed]
- Spencer, C.N.; McQuade, J.L.; Gopalakrishnan, V.; McCulloch, J.A.; Vetizou, M.; Cogdill, A.P.; Khan, A.W.; Zhang, X.; White, M.G.; Peterson, C.B.; et al. Dietary fiber and probiotics influence the gut microbiome and melanoma immunotherapy response. Science 2021, 374, 1632–1640. [Google Scholar] [CrossRef] [PubMed]
- Giordan, Q.; Salleron, J.; Vallance, C.; Moriana, C.; Clement-Duchene, C. Impact of Antibiotics and Proton Pump Inhibitors on Efficacy and Tolerance of Anti-PD-1 Immune Checkpoint Inhibitors. Front. Immunol. 2021, 12, 716317. [Google Scholar] [CrossRef]
- Uchiyama, A.A.T.; Silva, P.A.I.A.; Lopes, M.S.M.; Yen, C.T.; Ricardo, E.D.; Mutão, T.; Pimenta, J.R.; Machado, L.M.; Shimba, D.S.; Peixoto, R.D. Proton Pump Inhibitors and Oncologic Treatment Efficacy: A Practical Review of the Literature for Oncologists. Curr. Oncol. 2021, 28, 783–799. [Google Scholar] [CrossRef] [PubMed]
- Derosa, L.; Routy, B.; Fidelle, M.; Iebba, V.; Alla, L.; Pasolli, E.; Segata, N.; Desnoyer, A.; Pietrantonio, F.; Ferrere, G.; et al. Gut Bacteria Composition Drives Primary Resistance to Cancer Immunotherapy in Renal Cell Carcinoma Patients. Eur. Urol. 2020, 78, 195–206. [Google Scholar] [CrossRef] [PubMed]
- Baunwall, S.M.D.; Lee, M.M.; Eriksen, M.K.; Mullish, B.H.; Marchesi, J.R.; Dahlerup, J.F.; Hvas, C.L. Faecal microbiota transplantation for recurrent Clostridioides difficile infection: An updated systematic review and meta-analysis. eClinicalMedicine 2020, 29–30, 100642. [Google Scholar] [CrossRef] [PubMed]
- Porcari, S.; Benech, N.; Valles-Colomer, M.; Segata, N.; Gasbarrini, A.; Cammarota, G.; Sokol, H.; Ianiro, G. Key determinants of success in fecal microbiota transplantation: From microbiome to clinic. Cell Host Microbe 2023, 31, 712–733. [Google Scholar] [CrossRef] [PubMed]
- Baruch, E.N.; Youngster, I.; Ben-Betzalel, G.; Ortenberg, R.; Lahat, A.; Katz, L.; Adler, K.; Dick-Necula, D.; Raskin, S.; Bloch, N.; et al. Fecal microbiota transplant promotes response in immunotherapy-refractory melanoma patients. Science 2021, 371, 602–609. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, T.; Sun, J.; Liu, N. Fecal Microbiota Transplantation and Health Outcomes: An Umbrella Review of Meta-Analyses of Randomized Controlled Trials. Front. Cell. Infect. Microbiol. 2022, 12, 899845. [Google Scholar] [CrossRef] [PubMed]
- Araujo, D.V.; Bernal, M.O.; Tan, T.J.Y.; Heirali, A.A.; Schneeberger, P.H.; Muniz, T.P.; Chen, B.; Stayner, L.-A.; Xia, K.; Hansen, A.R.; et al. First-in-class microbial ecosystem therapeutics 4 (MET4) in metastatic solid cancer patients treated with immunotherapy: MET4-IO. J. Clin. Oncol. 2020, 38, 3098. [Google Scholar] [CrossRef]
- Spreafico, A.; Heirali, A.A.; Araujo, D.V.; Tan, T.J.; Oliva, M.; Schneeberger, P.H.; Chen, B.; Wong, M.K.; Stayner, L.-A.; Hansen, A.R.; et al. First-in-class Microbial Ecosystem Therapeutic 4 (MET4) in combination with immune checkpoint inhibitors in patients with advanced solid tumors (MET4-IO trial). Ann. Oncol. 2023, 34, 520–530. [Google Scholar] [CrossRef]
- Zhao, H.; Li, D.; Liu, J.; Zhou, X.; Han, J.; Wang, L.; Fan, Z.; Feng, L.; Zuo, J.; Wang, Y. Bifidobacterium breve predicts the efficacy of anti-PD-1 immunotherapy combined with chemotherapy in Chinese NSCLC patients. Cancer Med. 2022, 12, 6325–6336. [Google Scholar] [CrossRef] [PubMed]
- Cruellas, M.; Yubero, A.; Zapata, M.; Galvez, E.M.; Gascón, M.; Isla, D.; Lastra, R.; Martínez-Lostao, L.; Ocariz, M.; Pardo, J.; et al. How Could Antibiotics, Probiotics, and Corticoids Modify Microbiota and Its Influence in Cancer Immune Checkpoint Inhibitors: A Review. Infect. Immun. 2021, 89. [Google Scholar] [CrossRef]
- Tomita, Y.; Ikeda, T.; Sakata, S.; Saruwatari, K.; Sato, R.; Iyama, S.; Jodai, T.; Akaike, K.; Ishizuka, S.; Saeki, S.; et al. Association of Probiotic Clostridium butyricum Therapy with Survival and Response to Immune Checkpoint Blockade in Patients with Lung Cancer. Cancer Immunol. Res. 2020, 8, 1236–1242. [Google Scholar] [CrossRef] [PubMed]
- Takada, K.; Shimokawa, M.; Takamori, S.; Shimamatsu, S.; Hirai, F.; Tagawa, T.; Okamoto, T.; Hamatake, M.; Tsuchiya-Kawano, Y.; Otsubo, K.; et al. Clinical impact of probiotics on the efficacy of anti-PD -1 monotherapy in patients with nonsmall cell lung cancer: A multicenter retrospective survival analysis study with inverse probability of treatment weighting. Int. J. Cancer 2021, 149, 473–482. [Google Scholar] [CrossRef]
- Dizman, N.; Meza, L.; Bergerot, P.; Alcantara, M.; Dorff, T.; Lyou, Y.; Frankel, P.; Cui, Y.; Mira, V.; Llamas, M.; et al. Nivolumab plus ipilimumab with or without live bacterial supplementation in metastatic renal cell carcinoma: A randomized phase 1 trial. Nat. Med. 2022, 28, 704–712. [Google Scholar] [CrossRef]
- Takada, K.; Buti, S.; Bersanelli, M.; Shimokawa, M.; Takamori, S.; Matsubara, T.; Takenaka, T.; Okamoto, T.; Hamatake, M.; Tsuchiya-Kawano, Y.; et al. Antibiotic-dependent effect of probiotics in patients with non-small cell lung cancer treated with PD-1 checkpoint blockade. Eur. J. Cancer 2022, 172, 199–208. [Google Scholar] [CrossRef]
- Miura, K.; Sano, Y.; Niho, S.; Kawasumi, K.; Mochizuki, N.; Yoh, K.; Matsumoto, S.; Zenke, Y.; Ikeda, T.; Nosaki, K.; et al. Impact of concomitant medication on clinical outcomes in patients with advanced non-small cell lung cancer treated with immune checkpoint inhibitors: A retrospective study. Thorac. Cancer 2021, 12, 1983–1994. [Google Scholar] [CrossRef] [PubMed]
- Svaton, M.; Zemanova, M.; Zemanova, P.; Kultan, J.; Fischer, O.; Skrickova, J.; Jakubikova, L.; Cernovska, M.; Hrnciarik, M.; Jirousek, M.; et al. Impact of Concomitant Medication Administered at the Time of Initiation of Nivolumab Therapy on Outcome in Non-small Cell Lung Cancer. Anticancer. Res. 2020, 40, 2209–2217. [Google Scholar] [CrossRef]
- Gouez, M.; Raynard, B.; Marijnen, P.; Ho Hio Hen, N.; Fervers, B. Nutrition et activité physique adaptée (APA) pendant et après les traitements du cancer: Bénéfices thérapeutiques, physiopathologie, recommandations, prise en charge clinique. Bull. Cancer 2022, 109, 516–527. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Li, H.; Lv, X.; Hu, L.; Li, W.; Zi, M.; He, Y. Impact of Diets on Response to Immune Checkpoint Inhibitors (ICIs) Therapy against Tumors. Life 2022, 12, 409. [Google Scholar] [CrossRef] [PubMed]
- Zitvogel, L.; Kroemer, G. Boosting the immunotherapy response by nutritional interventions. J. Clin. Investig. 2022, 132, e161483. [Google Scholar] [CrossRef]
- Messaoudene, M.; Pidgeon, R.; Richard, C.; Ponce, M.; Diop, K.; Benlaifaoui, M.; Nolin-Lapalme, A.; Cauchois, F.; Malo, J.; Belkaid, W.; et al. A Natural Polyphenol Exerts Antitumor Activity and Circumvents Anti-PD-1 Resistance through Effects on the Gut Microbiota. Cancer Discov. 2022, 12, 1070–1087. [Google Scholar] [CrossRef] [PubMed]
- Malo, J.; Benlaïfaoui, M.; Diop, K.; Messaoudene, M.; Belkaid, W.; Elkrief, A.; Blais, N.; Tehfe, M.; Florescu, M.; Al-Saleh, A.; et al. CN12 Impact of diet on immune checkpoint blockade: Nurse-directed dietary intervention. Ann. Oncol. 2022, 33, S1356. [Google Scholar] [CrossRef]
- Botticelli, A.; Vernocchi, P.; Marini, F.; Quagliariello, A.; Cerbelli, B.; Reddel, S.; Del Chierico, F.; Di Pietro, F.; Giusti, R.; Tomassini, A.; et al. Gut metabolomics profiling of non-small cell lung cancer (NSCLC) patients under immunotherapy treatment. J. Transl. Med. 2020, 18, 49. [Google Scholar] [CrossRef] [PubMed]
- Vernocchi, P.; Del Chierico, F.; Putignani, L. Gut Microbiota Metabolism and Interaction with Food Components. Int. J. Mol. Sci. 2020, 21, 3688. [Google Scholar] [CrossRef]
- Nomura, M.; Nagatomo, R.; Doi, K.; Shimizu, J.; Baba, K.; Saito, T.; Matsumoto, S.; Inoue, K.; Muto, M. Association of Short-Chain Fatty Acids in the Gut Microbiome with Clinical Response to Treatment With Nivolumab or Pembrolizumab in Patients with Solid Cancer Tumors. JAMA Netw. Open 2020, 3, e202895. [Google Scholar] [CrossRef]
- Gui, Q.; Li, H.; Wang, A.; Zhao, X.; Tan, Z.; Chen, L.; Xu, K.; Xiao, C. The association between gut butyrate-producing bacteria and non-small-cell lung cancer. J. Clin. Lab. Anal. 2020, 34, e23318. [Google Scholar] [CrossRef]
- Sannicolo, S.; Levra, M.G.; Le Gouellec, A.; Aspord, C.; Boccard, J.; Chaperot, L.; Toussaint, B.; Moro-Sibilot, D.; Hannani, D.; Toffart, A.-C. Identification of a predictive metabolic signature of response to immune checkpoint inhibitors in non-small cell lung cancer: METABO-ICI clinical study protocol. Respir. Med. Res. 2021, 80, 100845. [Google Scholar] [CrossRef]
- Hopkins, A.M.; Kichenadasse, G.; Karapetis, C.S.; Rowland, A.; Sorich, M.J. Concomitant Proton Pump Inhibitor Use and Survival in Urothelial Carcinoma Treated with Atezolizumab. Clin. Cancer Res. 2020, 26, 5487–5493. [Google Scholar] [CrossRef]
- Majeed, H.J.; Riquelme, M.V.; Davis, B.C.; Gupta, S.; Angeles, L.; Aga, D.S.; Garner, E.; Pruden, A.; Vikesland, P.J. Evaluation of Metagenomic-Enabled Antibiotic Resistance Surveillance at a Conventional Wastewater Treatment Plant. Front. Microbiol. 2021, 12, 657954. [Google Scholar] [CrossRef]
- Kulkarni, A.; Kumar, M.; Pease, D.F.; Wang, Y.; DeFor, T.E.; Patel, M. Impact of antibiotics and proton pump inhibitors on clinical outcomes of immune check point blockers in advanced non-small cell lung cancers and metastatic renal cell cancer. J. Clin. Oncol. 2019, 37 (Suppl. 15), e20520. [Google Scholar] [CrossRef]
- Qiu, H.; Ma, Q.-G.; Chen, X.-T.; Wen, X.; Zhang, N.; Liu, W.-M.; Wang, T.-T.; Zhang, L.-Z. Different classes of antibiotics exhibit disparate negative impacts on the therapeutic efficacy of immune checkpoint inhibitors in advanced non-small cell lung cancer patients. Am. J. Cancer Res. 2022, 12, 3175–3184. [Google Scholar] [PubMed]
- Galli, G.; Poggi, M.; Fucà, G.; Imbimbo, M.; Proto, C.; Signorelli, D.; Vitali, M.; Zilembo, N.; Ganzinelli, M.; de Braud, F.; et al. Effects of antibiotic use during immunotherapy in metastatic non-small cell lung cancer. Ann. Oncol. 2018, 29, viii531. [Google Scholar] [CrossRef]
- Mielgo-Rubio, X.; Chara, L.; Sotelo-Lezama, M.; Castro, R.L.; Rubio-Martínez, J.; Velastegui, A.; Olier-Garate, C.; Falagan, S.; Gómez-Barreda, I.; Bautista-Sanz, P.; et al. MA10.01 Antibiotic Use and PD-1 Inhibitors: Shorter Survival in Lung Cancer, Especially when Given Intravenously. Type of Infection Also Matters. J. Thorac. Oncol. 2018, 13, S389. [Google Scholar] [CrossRef]
- Huang, J.; Zheng, X.; Kang, W.; Hao, H.; Mao, Y.; Zhang, H.; Chen, Y.; Tan, Y.; He, Y.; Zhao, W.; et al. Metagenomic and metabolomic analyses reveal synergistic effects of fecal microbiota transplantation and anti-PD-1 therapy on treating colorectal cancer. Front. Immunol. 2022, 13, 874922. [Google Scholar] [CrossRef] [PubMed]
- Ochi, N.; Ichihara, E.; Takigawa, N.; Harada, D.; Inoue, K.; Shibayama, T.; Hosokawa, S.; Kishino, D.; Harita, S.; Oda, N.; et al. The effects of antibiotics on the efficacy of immune checkpoint inhibitors in patients with non–small-cell lung cancer differ based on PD-L1 expression. Eur. J. Cancer 2021, 149, 73–81. [Google Scholar] [CrossRef]
- Cortellini, A.; Facchinetti, F.; Derosa, L.; Pinato, D.J. Antibiotic Exposure and Immune Checkpoint Inhibitors in Patients With NSCLC: The Backbone Matters. J. Thorac. Oncol. 2022, 17, 739–741. [Google Scholar] [CrossRef]
- Mino-Kenudson, M.; Schalper, K.; Cooper, W.; Dacic, S.; Hirsch, F.R.; Jain, D.; Lopez-Rios, F.; Tsao, M.S.; Yatabe, Y.; Beasley, M.B.; et al. Predictive Biomarkers for Immunotherapy in Lung Cancer: Perspective from the International Association for the Study of Lung Cancer Pathology Committee. J. Thorac. Oncol. 2022, 17, 1335–1354. [Google Scholar] [CrossRef]
- Bodor, J.N.; Boumber, Y.; Borghaei, H. Biomarkers for immune checkpoint inhibition in non-small cell lung cancer (NSCLC). Cancer 2020, 126, 260–270. [Google Scholar] [CrossRef]
- Mitsuhashi, A.; Okuma, Y. Perspective on immune oncology with liquid biopsy, peripheral blood mononuclear cells, and microbiome with non-invasive biomarkers in cancer patients. Clin. Transl. Oncol. 2018, 20, 966–974. [Google Scholar] [CrossRef]
- Hellmann, M.D.; Ciuleanu, T.-E.; Pluzanski, A.; Lee, J.S.; Otterson, G.A.; Audigier-Valette, C.; Minenza, E.; Linardou, H.; Burgers, S.; Salman, P.; et al. Nivolumab plus Ipilimumab in Lung Cancer with a High Tumor Mutational Burden. N. Engl. J. Med. 2018, 378, 2093–2104. [Google Scholar] [CrossRef] [PubMed]
- Fridman, W.H.; Zitvogel, L.; Sautès–Fridman, C.; Kroemer, G. The immune contexture in cancer prognosis and treatment. Nat. Rev. Clin. Oncol. 2017, 14, 717–734. [Google Scholar] [CrossRef]
- Okuma, Y.; Wakui, H.; Utsumi, H.; Sagawa, Y.; Hosomi, Y.; Kuwano, K.; Homma, S. Soluble Programmed Cell Death Ligand 1 as a Novel Biomarker for Nivolumab Therapy for Non-Small-cell Lung Cancer. Clin. Lung Cancer 2018, 19, 410–417.e1. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.J.; Yu, H.; Khan, M.; Gill, J.; Santhosh, S.; Chatterjee, U.; Iovoli, A.; Farrugia, M.; Mohammadpour, H.; Wooten, K.; et al. Evaluation of Optimal Threshold of Neutrophil-Lymphocyte Ratio and Its Association with Survival Outcomes among Patients with Head and Neck Cancer. JAMA Netw. Open 2022, 5, e227567. [Google Scholar] [CrossRef] [PubMed]
- Auclin, E.; Nolin-Lapalme, A.; Richard, C.; Malo, J.; Tonneau, M.; Benlaifaoui, M.; Ponce, M.; Messaoudene, M.; Okuma, Y.; Hakozaki, T.; et al. Association between lung immune prognostic index, microbiome, and immunotherapy outcomes in non–small cell lung cancer. J. Clin. Oncol. 2022, 40 (Suppl. 16), 9050. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| References | Sample | Type of Study | ICI | Stage of NSCLC | N | Technique | Responders | Non Responders | Notes | Country |
|---|---|---|---|---|---|---|---|---|---|---|
| (Routy et al., 2018) [22] | Feces | Retrospective | Anti-PD-1 | All stages | 153 | WGS | Higher: Akkermansia muciniphila, Ruminococcus spp., Allistepes spp. and Eubacterium spp. | Higher: Parabacteroides distasonis, Bacteriodes nordii | ATB uptake negatively impacts OS, but proton pump inhibitor did not. | France |
| (Derosa et al., 2022) [32,33] | Feces | Prospective | Anti-PD-1 | All stages | 338 | WGS | Higher: A. muciniphila, Eubacterium hallii, and Bifidobacterium adolescentis | Higher: Clostridium innoccuum | Stools with Akkermansia above the 77th percentile is deleterious. | France, Canada |
| (Newsome at al., 2022) [36] | Feces | Prospective | Anti-PD-1/PD-L1 or anti-PD-L1 and anti-CTLA-4 combination | Advanced | 65 | 16S rRNA (V1-V3) | Higher: Ruminococcus, Akkermansia, Blautia, and Faecalibacterium | NA | RNAseq on fecal RNA (N = 10) showed different bacterial transcriptomes within responders and non-responders, such as carbon fixation pathway enriched in prokaryotes in responders while non-responders were enriched in phosphotransferase system. | United States |
| (Martini et al., 2022) [38] | Feces | Prospective | Anti-PD-L1 | Advanced | 10 | 16S rRNA (V4) | Higher: Agathobacter M104/1 and Blautia SR1/5 | NA | All patients received ICI as cetuximab + avelumab combination. | Italy |
| (Katayama et al., 2019) [39] | Feces | Retrospective | Anti-PD-1 | Advanced | 17 | 16S rRNA (V1-V2) | Higher: Lactobacillus, Clostridium, and Syntrophococcus | Higher: Sutterella, Bilophila and Parabacteroides | Patients with higher abundance of Lactobacillus and Clostridium also had longer treatment to TTF. | Japan |
| (Hakozaki et al., 2020) [40] | Feces | Prospective | Anti-PD-1/PD-L1 | Advanced | 70 | 16S rRNA (V3-V4) | Higher: Agathobacter and Ruminococcaceae UCG 13 | Higher: Eggerthellaceae and Barnesiella | ATB use was associated with lower α-diversity. Lactobacillaceae and Raoultella were enriched in patients with no significant irAE. | Japan |
| (Shoji et al., 2021) [41] | Feces and Saliva | Prospective | Anti-PD-1/PD-L1 | Stage II/III/IV | 28 | 16S rRNA (V3-V4) | Higher: Blautia | Higher: RF32 unclassified | Responders had higher α-diversity but lower β-diversity in feces. No significant signal was found from saliva. | Japan |
| (Ouaknine et al., 2018) [42] | Blood | Prospective | Anti-PD-1 | Advanced | 35 | 16S rRNA (V3-V4) | Higher: Peptostreptococcaceae, Lewinella, Paludibaculum, and Holophagae | Higher: Gemmatimonadaceae | Presence of Gemmatimonadaceae at baseline was associated with worse PFS and OS. | France |
| (Jin et al., 2019) [43] | Feces | Prospective | Anti-PD-1 | Advanced | 37 | 16S rRNA (V3-V4) | Higher: Alistipes putredinis, Bifidobacterium longum, and Prevotella copri | Higher: Ruminococcus_unclassified | Responders had higher α-diversity. High α-diversity was associated with enhanced memory T cell and NK cell signatures. | China |
| (Song et al., 2020) [44] | Feces | Prospective | Anti-PD-1 | Advanced | 63 | WGS | Higher: Parabacteroides and Methanobrevibacter | Higher: Veillonella, Selenomonadales, and Negativicutes | Responders had higher β-diversity. Differences in KEGG functional group and metabolic potential of methanol and methane were also noted. | China |
| (He et al., 2021) [45] | Feces | Prospective | Anti-PD-1 | Advanced | 16 | 16S rRNA (V3-V4) | Higher: Escherichia, Shigella, Akkermansia, and Olsenella | Higher: Anaeroglobus | Escherichia-Shigella was positively correlated with IL-12, IFN-γ, and basophils in plasma. Akkermansia was positively correlated with monocytes. | China |
| (Zhang et al., 2021) [46] | Feces and Saliva | Prospective | Anti-PD-1 | Advanced | 75 | 16S rRNA (V3-V4) | Higher (in feces): Desulfovibrio, Actinomycetales, Bifidobacterium, Odoribacteraceae, Anaerostipes, Rikenellaceae, Faecalibacterium, and Alistipes | Higher (in feces): Fusobacterales, Fusobacteriia, Fusobacterium, Fusobacteria, and Fusobacteriaceae | Responders had higher α-diversity in feces. The abundance of Streptococcus in saliva was associated with higher CD8+ T cell density. α-diversity between feces and saliva microbiota was uncorrelated. | China |
| (Masuhiro et al., 2022) [47] | BAL | Prospective | PD-1 | Advanced | 12 | 16S rRNA (V3-V4) | Higher: Bacteriodetes | Higher: Proteobacteria | Responders had higher α-diversity and CXCL9 levels in BAL. | Japan |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duttagupta, S.; Hakozaki, T.; Routy, B.; Messaoudene, M. The Gut Microbiome from a Biomarker to a Novel Therapeutic Strategy for Immunotherapy Response in Patients with Lung Cancer. Curr. Oncol. 2023, 30, 9406-9427. https://doi.org/10.3390/curroncol30110681
Duttagupta S, Hakozaki T, Routy B, Messaoudene M. The Gut Microbiome from a Biomarker to a Novel Therapeutic Strategy for Immunotherapy Response in Patients with Lung Cancer. Current Oncology. 2023; 30(11):9406-9427. https://doi.org/10.3390/curroncol30110681
Chicago/Turabian StyleDuttagupta, Sreya, Taiki Hakozaki, Bertrand Routy, and Meriem Messaoudene. 2023. "The Gut Microbiome from a Biomarker to a Novel Therapeutic Strategy for Immunotherapy Response in Patients with Lung Cancer" Current Oncology 30, no. 11: 9406-9427. https://doi.org/10.3390/curroncol30110681