
Citrus aurantium ‘Crispifolia’ Essential Oil: A Promise for Nutraceutical Applications
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Isolation of Essential Oil
2.2. GC-MS Analysis
2.3. Bacterial Strains
2.4. Antibacterial Tests
2.5. MIC Experiment
2.6. Antibiofilm Inhibition Test
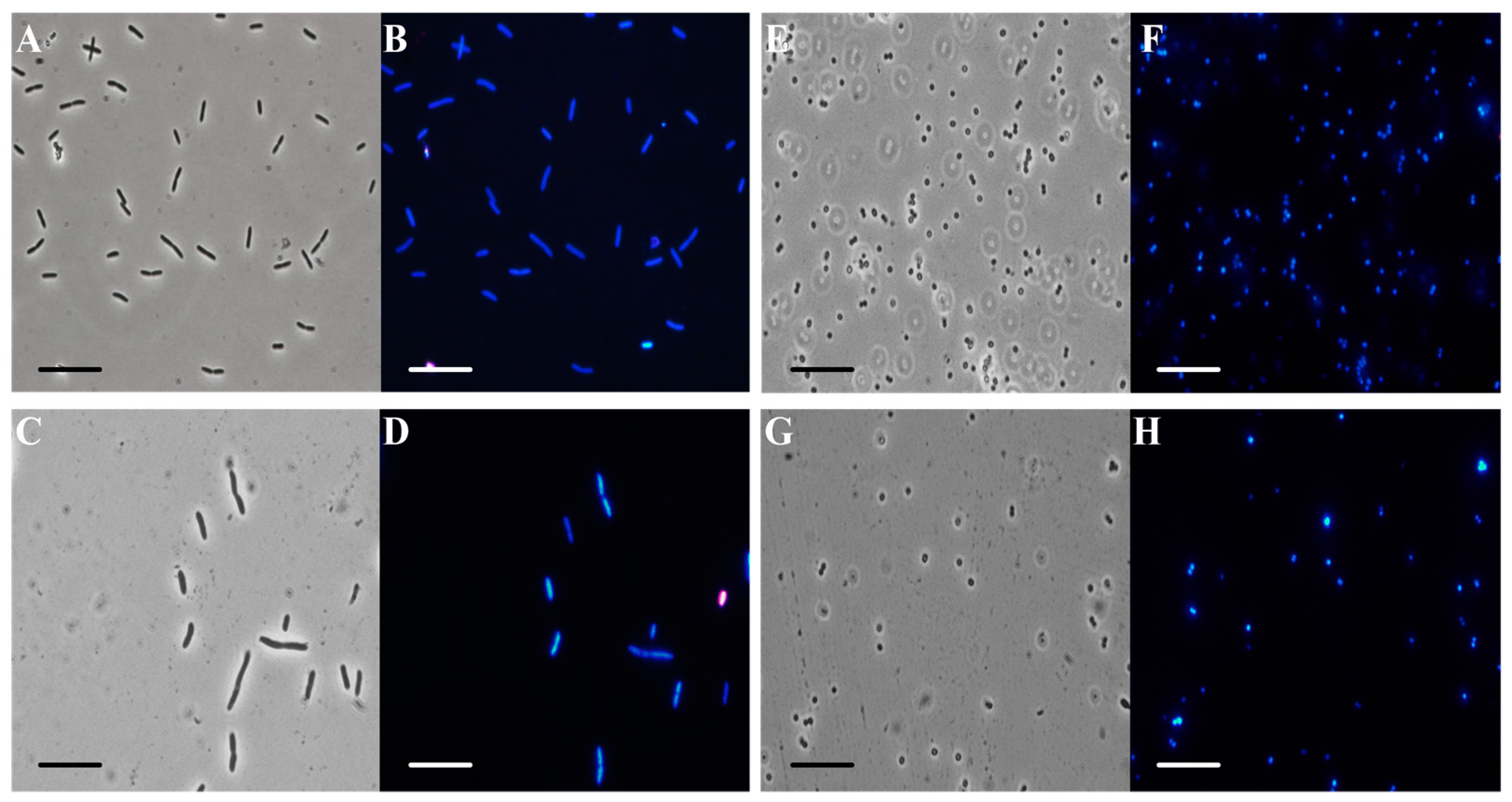
2.7. Fluorescence Microscopy Experiment
2.8. ABTS Scavenging Properties
2.9. Hydrogen Peroxide Scavenging Test
2.10. Cytotoxicity Tests
2.11. ROS Generation and Antioxidant Enzyme Activity against Polymorphonuclear Leukocytes (PMN)
2.12. Statistical Analysis
3. Results and Discussion
3.1. Composition of the Essential Oil
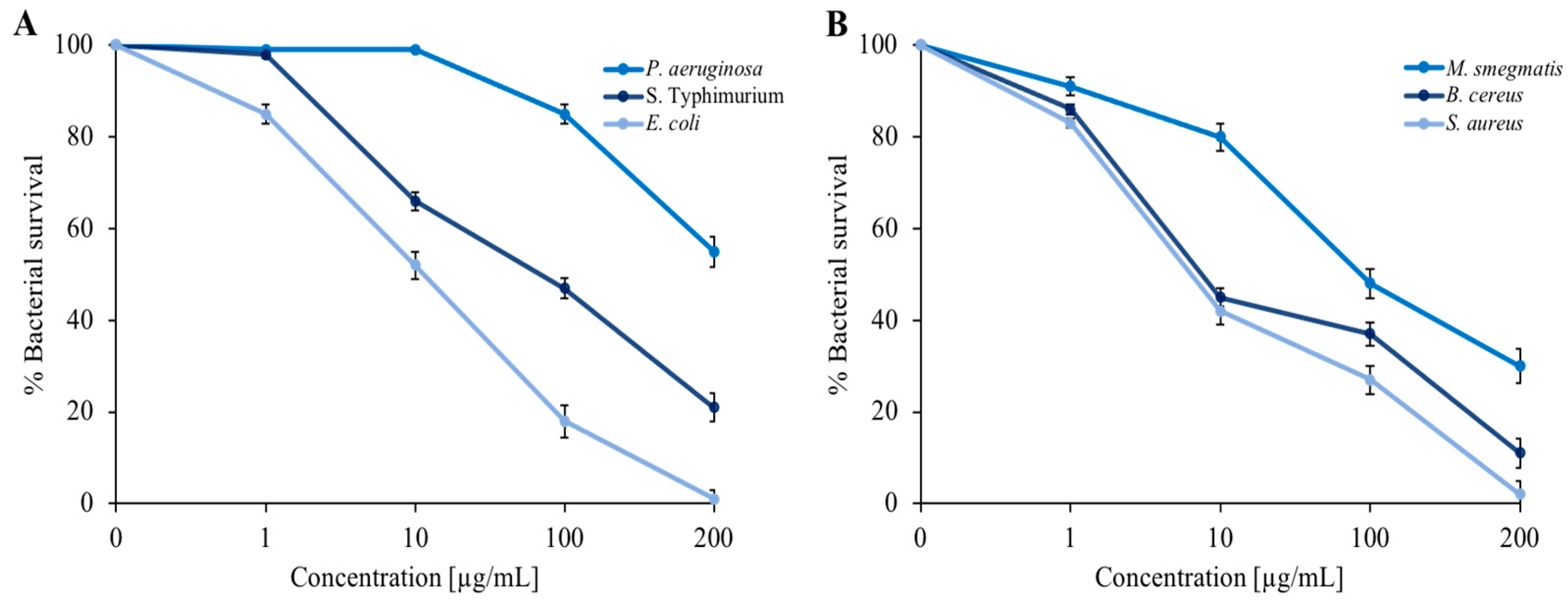
3.2. Antimicrobial Activity Tests
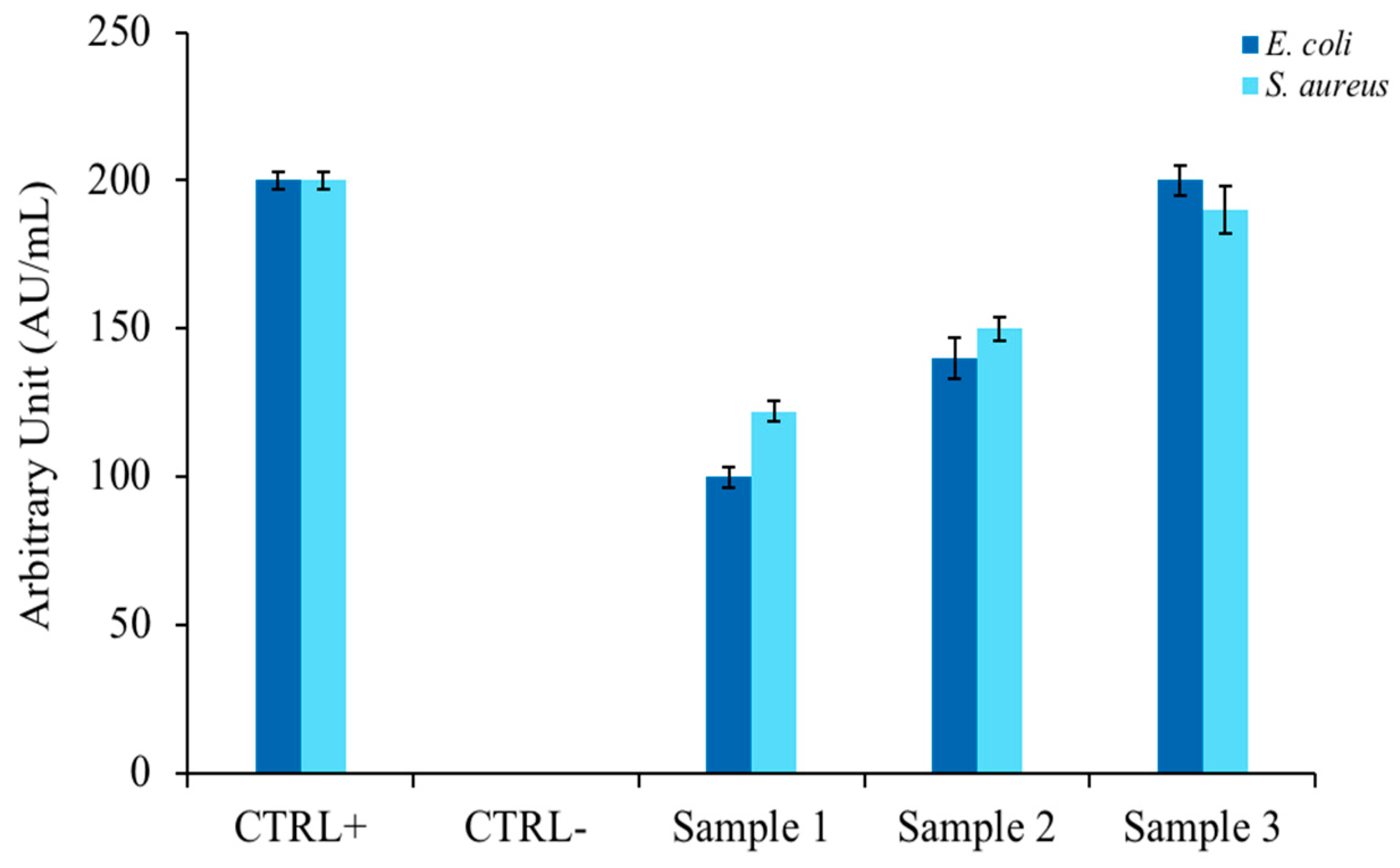
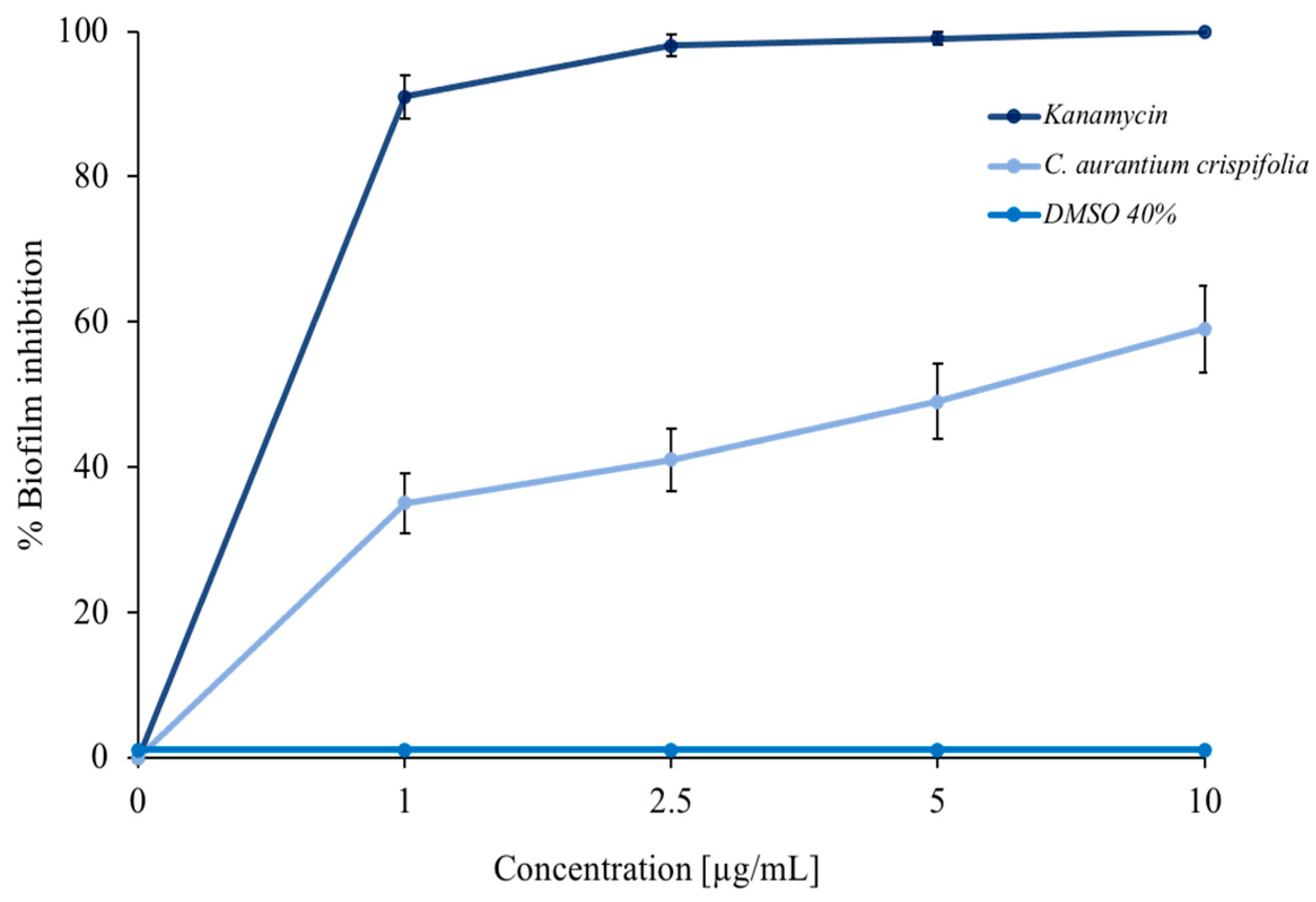
3.3. Antibiofilm Tests
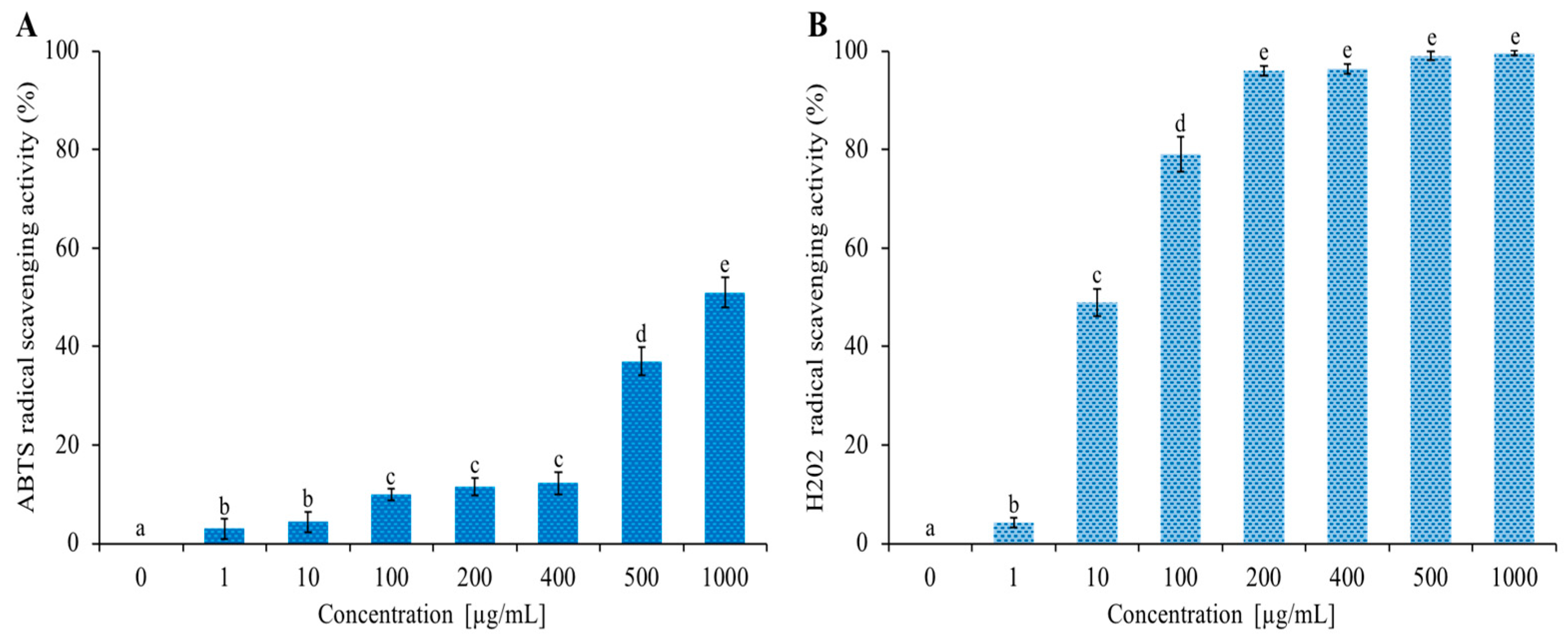
3.4. Antioxidant Activity
3.5. Cytotoxic Activity
3.6. ROS Generation and SOD and CAT Activity in PMN
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Sample Availability
References
- Swingle, W.T.; Reece, P.C.; Reuther, W.; Webber, H.J.; Batchelor, L.D. The Citrus Industry; University of California Press: Berkeley, CA, USA, 1967; pp. 190–430. [Google Scholar]
- Ferrer, V.; Costantino, G.; Paoli, M.; Paymal, N.; Quinton, C.; Ollitrault, P.; Tomi, F.; Luro, F. Intercultivar diversity of sour orange (Citrus aurantium L.) based on genetic markers, phenotypic characteristics, aromatic compounds and sensorial analysis. Agronomy 2021, 11, 1084. [Google Scholar] [CrossRef]
- Wu, G.A.; Prochnik, S.; Jenkins, J.; Salse, J.; Hellsten, U.; Murat, F.; Perrier, X.; Ruiz, M.; Scalabrin, S.; Terol, J.; et al. Sequencing of diverse mandarin, pummelo and orange genomes reveals complex history of admixture during Citrus domestication. Nat. Biotechnol. 2014, 32, 656–662. [Google Scholar] [CrossRef]
- Scora, R.W. On the history and origin of Citrus. Bull. Torrey Bot. Club 1975, 102, 369–375. [Google Scholar] [CrossRef]
- Barrett, H.C.; Rhodes, A.M. A numerical taxonomic study of affinity relationships in cultivated Citrus and its close relatives. Syst. Bot. 1976, 1, 105–136. [Google Scholar] [CrossRef]
- Fang, D.Q. Intra- and intergeneric relationships of Poncirus polyandar: Investigation by leaf isozymes. J. Wuhan Bot. Res. 1993, 11, 34–40. [Google Scholar]
- Nicolosi, E.; Deng, Z.N.; Gentile, A.; La Malfa, S.; Continella, G.; Tribulato, E. Citrus phylogeny and genetic origin of important species as investigated by molecular markers. Theor. Appl. Genet. 2000, 100, 1155–1166. [Google Scholar] [CrossRef]
- Bayer, R.J.; Mabberley, D.J.; Morton, C.; Miller, C.H.; Sharma, I.K.; Pfeil, B.E.; Rich, S.; Hitchcock, R.; Sykes, S. A molecular phylogeny of the orange subfamily (Rutaceae: Aurantioideae) using nine cpDNA sequences. Amer. J. Bot. 2009, 96, 668–685. [Google Scholar] [CrossRef]
- Malik, M.N.; Scora, R.W.; Soost, R.K. Studies on the origin of the lemon. Hilgardia 1974, 42, 361–382. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Xie, R.; Lu, Z.; Zhou, Z. The origin of cultivated Citrus as inferred from internal transcribed spacer and chloroplast DNA sequence and amplified fragment length polymorphism fingerprints. J. Amer. Soc. Hort. Sci. 2010, 135, 341–350. [Google Scholar] [CrossRef] [Green Version]
- USDA Foreign Agricultural Service 2017. Available online: https://www.fas.usda.gov (accessed on 15 December 2022).
- Bonesi, M.; Loizzo, M.R.; Leporini, M.; Tenuta, M.C.; Passalacqua, N.G.; Tundis, R. Comparative evaluation of petitgrain oils from six Citrus species alone and in combination as potential functional anti-radicals and antioxidant agents. Plant Biosyst. 2018, 152, 986–993. [Google Scholar] [CrossRef]
- Russo, C.; Maugeri, A.; Lombardo, G.E.; Musumeci, L.; Barreca, D.; Rapisarda, A.; Cirmi, S.; Navarra, M. The second life of Citrus fruit waste: A valuable source of bioactive compounds. Molecules 2021, 26, 5991. [Google Scholar] [CrossRef] [PubMed]
- Gioffrè, G.; Ursino, D.; Labate, M.L.C.; Giuffrè, A.M. The peel essential oil composition of bergamot fruit (Citrus bergamia, Risso) of Reggio Calabria (Italy): A review. Emir. J. Food Agric. 2020, 32, 835–845. [Google Scholar] [CrossRef]
- Rowshan, V.; Najafian, S. Changes of peel essential oil composition of Citrus aurantium L. during fruit maturation in Iran. J. Essent. Oil Bear. 2015, 18, 1006–1012. [Google Scholar] [CrossRef]
- Giuffrè, A.M.; Nobile, R. Citrus bergamia, Risso: The peel, the juice and the seed oil of the bergamot fruit of Reggio Calabria (South Italy). Emir. J. Food Agric. 2020, 32, 522–532. [Google Scholar] [CrossRef]
- Ferhat, M.A.; Boukhatem, M.N.; Hazzit, M.; Meklati, B.Y.; Chemat, F. Cold pressing, hydrodistillation and microwave dry distillation of Citrus essential oil from Algeria: A comparative study. Electron. J. Biol. 2016, S1, 30–41. [Google Scholar]
- Teneva, D.; Denkova-Kostova, R.; Goranov, B.; Hristova-Ivanova, Y.; Slavchev, A.; Denkova, Z.; Kostov, G. Chemical composition, antioxidant activity and antimicrobial activity of essential oil from Citrus aurantium L Zest against some pathogenic microorganisms. Z. Naturforsch. C 2019, 74, 105–111. [Google Scholar] [CrossRef] [Green Version]
- Lagha, R.; Ben Abdallah, F.; AL-Sarhan, B.; Al-Sodany, Y. Antibacterial and biofilm inhibitory activity of medicinal plant essential oils against Escherichia coli isolated from UTI patients. Molecules 2019, 24, 1161. [Google Scholar] [CrossRef] [Green Version]
- European Pharmacopoeia 10.3. 2020. Determination of Essential Oils in Herbal Drugs, 2.8.12., 307. Available online: https://www.edqm.eu/ (accessed on 10 December 2022).
- Bancheva, S.; Badalamenti, N.; Bruno, M. The essential oil composition of the endemic plant species Centaurea vandasii and chemotaxonomy of section Phalolepis (Asteraceae). Nat. Prod. Res. 2021. [Google Scholar] [CrossRef]
- Bauer, A.W.; Kirby, W.M.; Sherris, J.C.; Turck, M. Antibiotic susceptibility testing by a standardized single disk method. Am. J. Clin. Pathol. 1966, 45, 493–496. [Google Scholar] [CrossRef]
- Di Napoli, M.; Varcamonti, M.; Basile, A.; Bruno, M.; Maggi, F.; Zanfardino, A. Anti-Pseudomonas aeruginosa activity of hemlock (Conium maculatum, Apiaceae) essential oil. Nat. Prod. Res. 2019, 33, 3436–3440. [Google Scholar] [CrossRef]
- Badalamenti, N.; Russi, S.; Bruno, M.; Maresca, V.; Vaglica, A.; Ilardi, V.; Zanfardino, A.; Di Napoli, M.; Varcamonti, M.; Cianciullo, P.; et al. Dihydrophenanthrenes from a Sicilian accession of Himantoglossum robertianum (Loisel.) P. delforge showed antioxidant, antimicrobial, and antiproliferative activities. Plants 2021, 10, 2776. [Google Scholar] [CrossRef] [PubMed]
- Pota, G.; Zanfardino, A.; Di Napoli, M.; Cavasso, D.; Varcamonti, M.; D’Errico, G.; Pezzella, A.; Luciani, G.; Vitiello, G. Bioinspired antibacterial pva/melanin-TiO2 hybrid nanoparticles: The role of poly-vinyl-alcohol on their self-assembly and biocide activity. Colloids Surf. B Biointerfaces 2021, 202, 111671. [Google Scholar] [CrossRef]
- Di Napoli, M.; Maresca, V.; Varcamonti, M.; Bruno, M.; Badalamenti, N.; Basile, A.; Zanfardino, A. (+)-(E)-Chrysanthenyl acetate: A molecule with interesting biological properties contained in the Anthemis secundiramea (Asteraceae) Flowers. Appl. Sci. 2020, 10, 6808. [Google Scholar] [CrossRef]
- Di Napoli, M.; Maresca, V.; Sorbo, S.; Varcamonti, M.; Basile, A.; Zanfardino, A. Proteins of the fruit pulp of Acca sellowiana have antimicrobial activity directed against the bacterial membranes. Nat. Prod. Res. 2021, 35, 2942–2946. [Google Scholar] [CrossRef]
- Napolitano, A.; Di Napoli, M.; Castagliuolo, G.; Badalamenti, N.; Cicio, A.; Bruno, M.; Piacente, S.; Maresca, V.; Cianciullo, P.; Capasso, L.; et al. The chemical composition of the aerial parts of Stachys spreitzenhoferi (Lamiaceae) growing in Kythira Island (Greece), and their antioxidant, antimicrobial, and antiproliferative properties. Phytochemistry 2022, 203, 113373. [Google Scholar] [CrossRef]
- Beers, R.; Sizer, I. A spectrophotometric method for measuring the breakdown of hydrogen peroxide by catalase. J. Biol. Chem. 1952, 195, 133–140. [Google Scholar] [CrossRef]
- Vitiello, G.; Zanfardino, A.; Tammaro, O.; Di Napoli, M.; Caso, M.F.; Pezzella, A.; Varcamonti, M.; Silvestri, B.; D’Errico, G.; Costantini, A.; et al. Bioinspired hybrid eumelanin-TiO2 antimicrobial nanostructures: The key role of organo-inorganic frameworks in tuning eumelanin’s biocide action mechanism through membrane interaction. RSC Adv. 2018, 8, 28275–28283. [Google Scholar] [CrossRef] [Green Version]
- Zanfardino, A.; Bosso, A.; Gallo, G.; Pistorio, V.; Di Napoli, M.; Gaglione, R.; Dell’Olmo, E.; Varcamonti, M.; Notomista, E.; Arciello, A.; et al. Human apolipoprotein E as a reservoir of cryptic bioactive peptides: The case of ApoE 133–167. J. Pept. Sci. 2018, 24, e3095. [Google Scholar] [CrossRef]
- Manna, A.; Saha, P.; Sarkar, A.; Mukhopadhyay, D.; Bauri, A.K.; Kumar, D.; Das, P.; Chattopadhyay, S.; Chatterjee, M. Malabaricone-A induces a redox imbalance that mediates apoptosis in U937 Cell Line. PLoS ONE 2012, 7, e36938. [Google Scholar] [CrossRef]
- Barbosa, P.O.; Pala, D.; Silva, C.T.; de Souza, M.O.; do Amaral, J.F.; Vieira, R.A.L.; de Freitas Folly, G.A.; Volp, A.C.P.; de Freitas, R.N. Açai (Euterpe oleracea Mart.) pulp dietary intake improves cellular antioxidant enzymes and biomarkers of serum in healthy women. Nutrition 2016, 32, 674–680. [Google Scholar] [CrossRef]
- Badalamenti, N.; Bruno, M.; Schicchi, R.; Geraci, A.; Leporini, M.; Gervasi, L.; Tundis, R.; Loizzo, M.R. Chemical compositions and antioxidant activities of essential oils, and their combinations, obtained from flavedo by-product of seven cultivars of Sicilian Citrus aurantium L. Molecules 2022, 27, 1580. [Google Scholar] [CrossRef] [PubMed]
- Hsouna, A.B.; Hamdi, N.; Halima, N.B.; Abdelkafi, S. Characterization of essential oil from Citrus aurantium L. flowers: Antimicrobial and antioxidant activities. J. Oleo Sci. 2013, 62, 763–772. [Google Scholar] [CrossRef]
- Spernovasilis, N.; Psichogiou, M.; Poulakou, G. Skin manifestations of Pseudomonas aeruginosa infections. Curr. Opin. Infect. Dis. 2021, 34, 72–79. [Google Scholar] [CrossRef]
- Serra, R.; Grande, R.; Butrico, L.; Rossi, A.; Settimio, U.F.; Caroleo, B.; Amato, B.; Gallelli, L.; de Franciscis, S. Chronic wound infections: The role of Pseudomonas aeruginosa and Staphylococcus aureus. Expert Rev. Anti Infect. Ther. 2015, 13, 605–613. [Google Scholar] [CrossRef]
- Affhan, S.; Dachang, W.; Xin, Y.; Shang, D. Lactic acid bacteria protect human intestinal epithelial cells from Staphylococcus aureus and Pseudomonas aeruginosa infections. Genet. Mol. Res. 2015, 14, 17044–17058. [Google Scholar] [CrossRef]
- Andreoli, G.; Merla, C.; Valle, C.D.; Corpus, F.; Morganti, M.; D’incau, M.; Colmegna, S.; Marone, P.; Fabbi, M.; Barco, L.; et al. foodborne salmonellosis in italy: Characterization of Salmonella enterica Serovar Typhimurium and monophasic variant 4,[5],12:I− isolated from salami and human patients. J. Food Protect. 2017, 80, 632–639. [Google Scholar] [CrossRef]
- Schoeni, J.L.; Lee Wong, A.C. Bacillus cereus food poisoning and its toxins. J. Food Protect. 2005, 68, 636–648. [Google Scholar] [CrossRef]
- Nazzaro, F.; Fratianni, F.; De Martino, L.; Coppola, R.; De Feo, V. Effect of essential oils on pathogenic bacteria. Pharmaceuticals 2013, 6, 1451–1474. [Google Scholar] [CrossRef]
- Di Napoli, M.; Castagliuolo, G.; Badalamenti, N.; Maresca, V.; Basile, A.; Bruno, M.; Varcamonti, M.; Zanfardino, A. Antimicrobial, antibiofilm, and antioxidant properties of essential oil of Foeniculum vulgare Mill. leaves. Plants 2022, 11, 3573. [Google Scholar] [CrossRef]
- Borotová, P. Biological activity of Citrus aurantium var. bergamia essential oil as a food spoilage inhibitor. Sci. Pap. Anim. Sci. Biotechnol. 2021, 54, 117–120. [Google Scholar]
- González-Mas, M.C.; Rambla, J.L.; López-Gresa, M.P.; Blázquez, M.A.; Granell, A. Volatile compounds in Citrus essential oils: A comprehensive review. Front. Plant Sci. 2019, 10, 12. [Google Scholar] [CrossRef]
- Badalamenti, N.; Maresca, V.; Di Napoli, M.; Bruno, M.; Basile, A.; Zanfardino, A. Chemical composition and biological activities of Prangos ferulacea essential oils. Molecules 2022, 27, 7430. [Google Scholar] [CrossRef]
- Graßmann, J. Terpenoids as plant antioxidants. Vit Horm. 2005, 72, 505–535. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
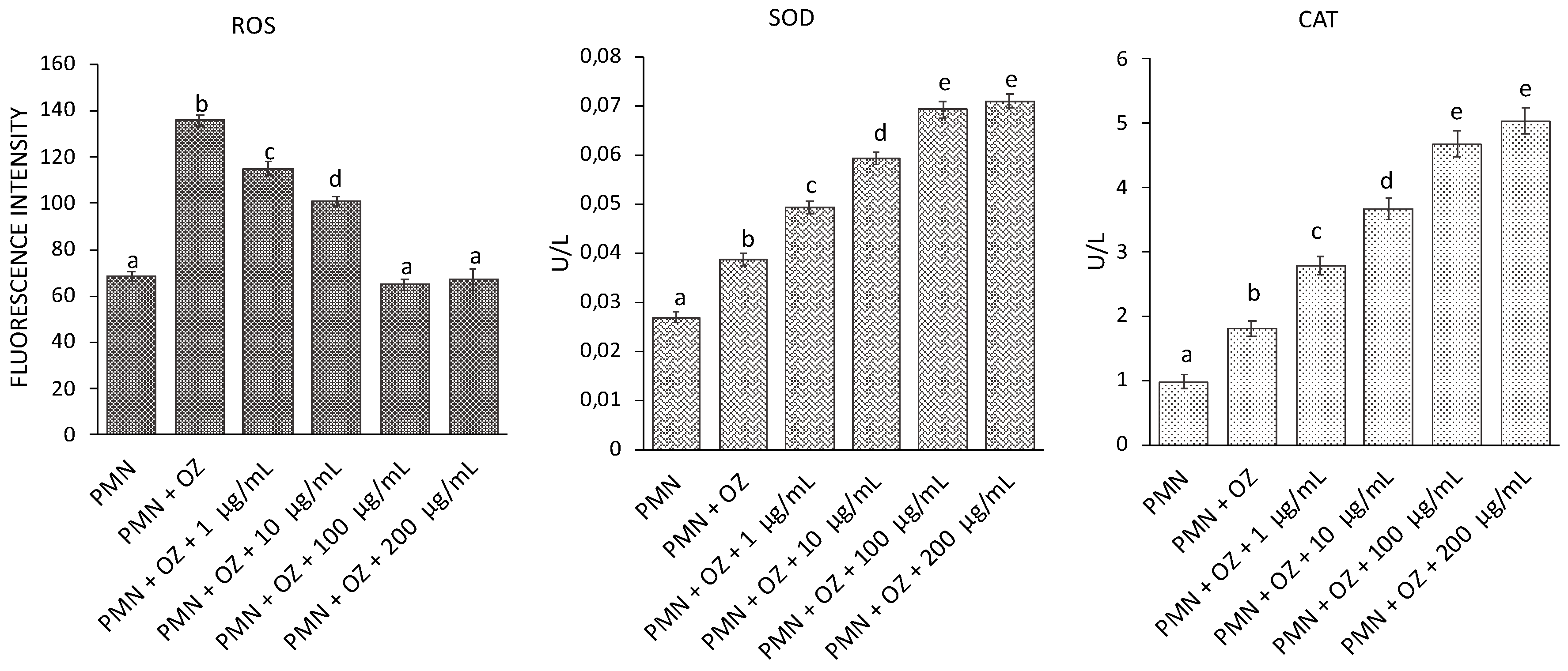{kind=link}
| No. | Compounds | LRIexp A | LRIlit B | A C (%) |
|---|---|---|---|---|
| 1 | β-Pinene | 977 | 981 | 0.91 d |
| 2 | β-Myrcene | 990 | 994 | 1.95 f |
| 3 | Limonene | 1031 | 1028 | 33.35 f |
| 4 | n-Octanol | 1070 | 1078 | 1.59 a |
| 5 | β-Linalool | 1098 | 1101 | 7.69 a |
| 6 | α-Terpineol | 1189 | 1194 | 7.06 a |
| 7 | cis-Geraniol | 1229 | 1235 | 1.63 a |
| 8 | β-Citral | 1238 | 1242 | 1.88 a |
| 9 | Bergamol | 1256 | 1258 | 6.77 a |
| 10 | trans-Geraniol | 1259 | 1267 | 3.36 a |
| 11 | Neryl acetate | 1365 | 1366 | 6.28 a |
| 12 | Geranyl acetate | 1386 | 1392 | 10.12 a |
| 13 | n-Decyl acetate | 1402 | 1406 | 1.44 a |
| 14 | Caryophyllene | 1419 | 1423 | 1.00 a |
| 15 | Germacrene D | 1480 | 1485 | 3.48 a |
| 16 | trans-Nerolidol | 1550 | 1554 | 4.57 a |
| Monoterpene Hydrocarbons | 36.21 | |||
| Oxygenated Monoterpenes | 44.79 | |||
| Sesquiterpene Hydrocarbons | 4.48 | |||
| Oxygenated Sesquiterpenes | 4.57 | |||
| Others | 3.03 | |||
| Total | 93.08 | |||
| Strains | MIC100 [µg/mL] |
|---|---|
| E. coli DH5α | 200 |
| P. aeruginosa PAO1 ATCC 15692 | >200 |
| S. Typhimurium ATCC14028 | >200 |
| S. aureus ATCC6538P | 200 |
| B. cereus ATCC10987 | >200 |
| M. smegmatis MC2155 | >200 |
| Sample | IC50 ABTS (µg/mL) | Sample | IC50 H2O2 (µg/mL) |
|---|---|---|---|
| C. aurantium ‘Crispifolia’ EO | 1000 | C. aurantium ‘Crispifolia’ EO | 10 |
| Ascorbic acid | 0.03 | Resveratrol | 0.05 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Napoli, M.; Castagliuolo, G.; Badalamenti, N.; Maresca, V.; Basile, A.; Bruno, M.; Varcamonti, M.; Zanfardino, A. Citrus aurantium ‘Crispifolia’ Essential Oil: A Promise for Nutraceutical Applications. Nutraceuticals 2023, 3, 153-164. https://doi.org/10.3390/nutraceuticals3010011
Di Napoli M, Castagliuolo G, Badalamenti N, Maresca V, Basile A, Bruno M, Varcamonti M, Zanfardino A. Citrus aurantium ‘Crispifolia’ Essential Oil: A Promise for Nutraceutical Applications. Nutraceuticals. 2023; 3(1):153-164. https://doi.org/10.3390/nutraceuticals3010011
Chicago/Turabian StyleDi Napoli, Michela, Giusy Castagliuolo, Natale Badalamenti, Viviana Maresca, Adriana Basile, Maurizio Bruno, Mario Varcamonti, and Anna Zanfardino. 2023. "Citrus aurantium ‘Crispifolia’ Essential Oil: A Promise for Nutraceutical Applications" Nutraceuticals 3, no. 1: 153-164. https://doi.org/10.3390/nutraceuticals3010011