
Potential of Microalgae as Functional Foods Applied to Mitochondria Protection and Healthy Aging Promotion


Abstract
:1. Introduction
2. Functions of Mitochondria

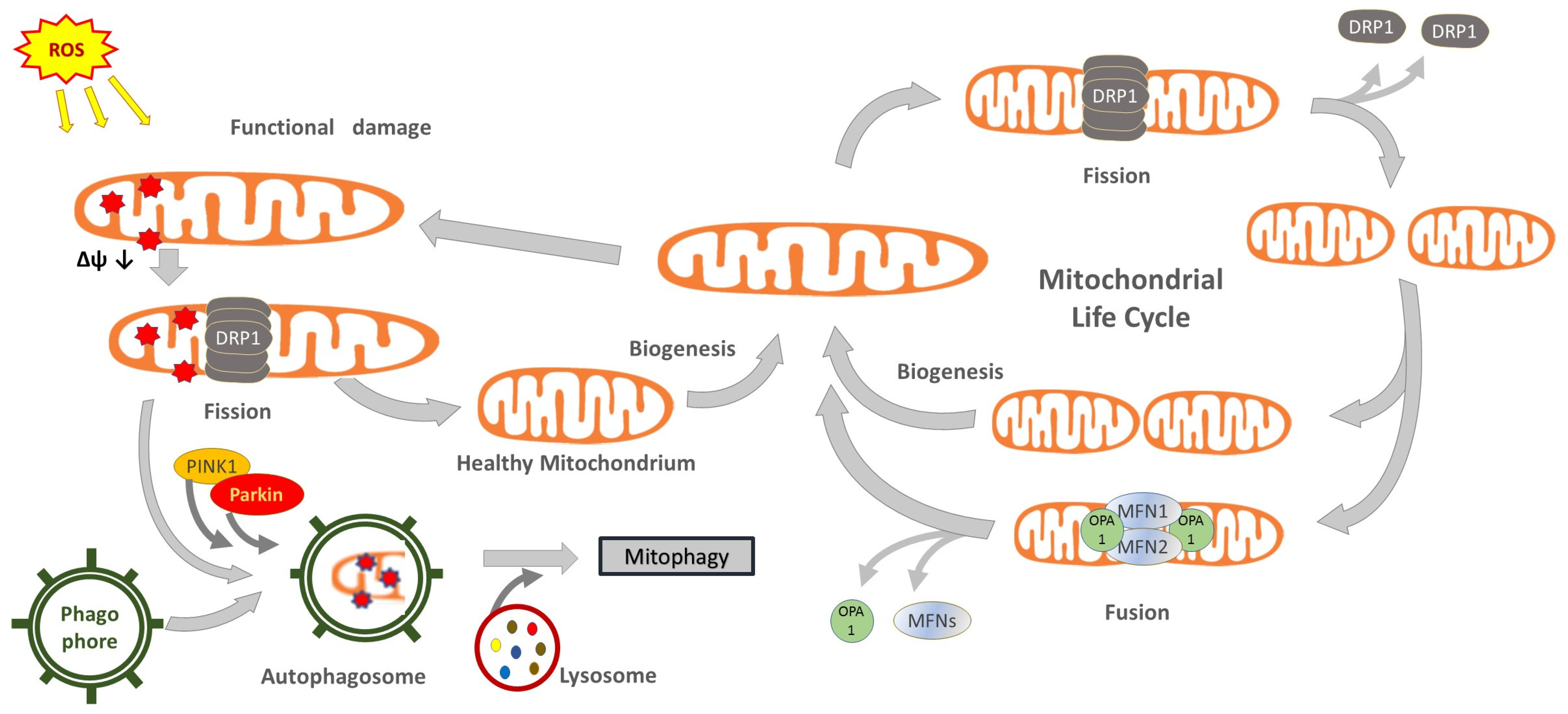{kind=link}
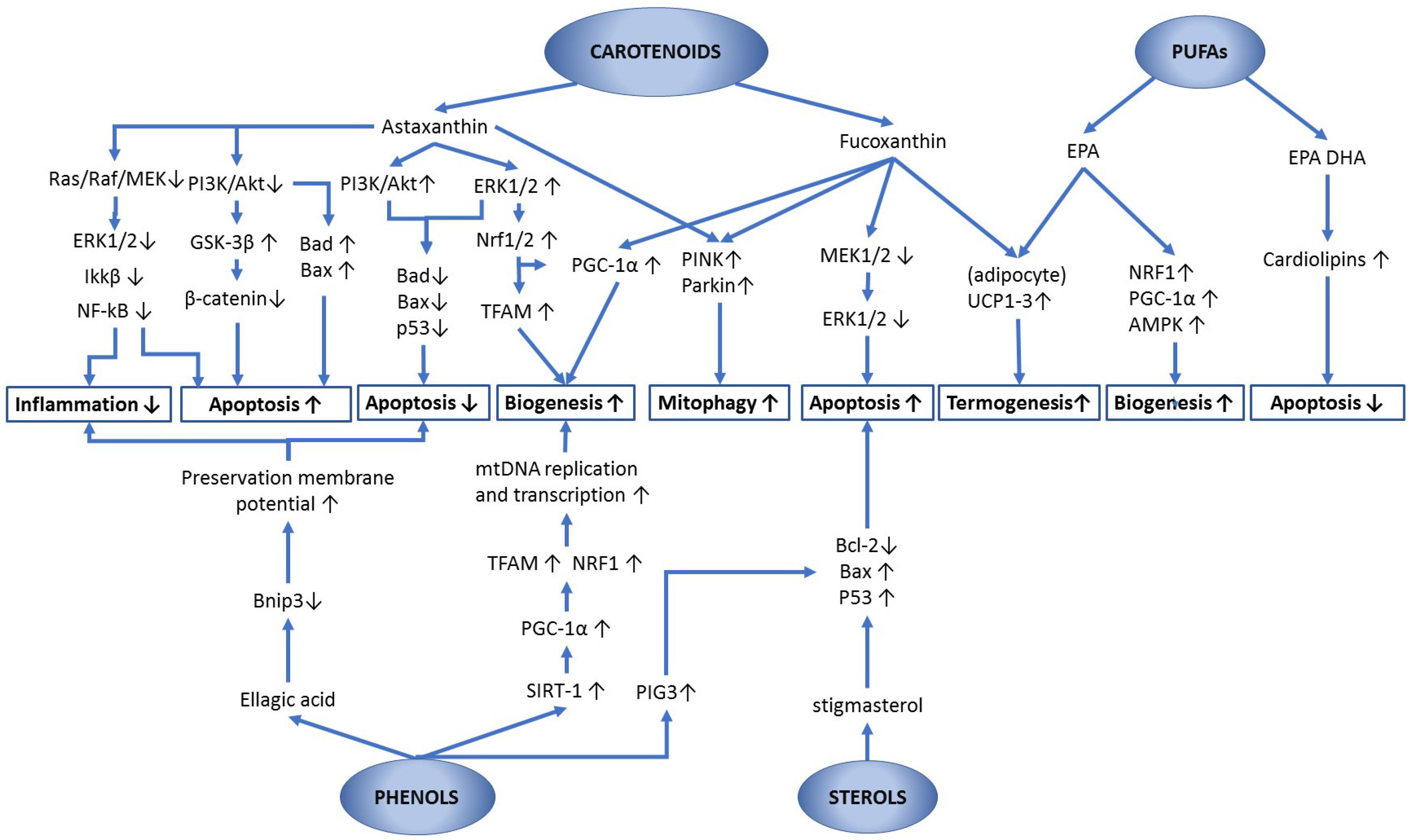{kind=link}
| Modulatory Active Compound | Ref. | Key Regulatory Enzymes or Pathways | Cycle Phase | Description |
|---|---|---|---|---|
| Astaxanthin | [41,42] | PGC-1α, Tfam | Biogenesis | MTs increase by organelle division. The organelles undergo mtDNA replication and subsequent division. |
| EPA | [43] | NRF-1, TFAM, COXIV, SIRT1, PGC-1α | ||
| EPA/DHA, curcumin | [44,45,46] | PGC-1α, NRF1 | ||
| Fucoxanthin | [47] | NRF1, NRF2 | ||
| Quercetin, resveratrol | [45] | Nrf2 | ||
| Salidroside (Rhodiola) | [48] | SIRT1 | ||
| Fucoxanthin, curcumin | [45,47,49] | PGC1α, Tfam | ||
| Fucoxanthin | [47] | Mfns; Opa1 | Fusion | Coordinated fusion of the inner and outer membranes between two organelles aimed to merge intact and slightly dysfunctional MTs. It is particularly useful in case of damaged mtDNA. |
| Omega-3 fatty acids | [46] | Mfns; Opa1 | ||
| Resveratrol | [32] | Mfn2 | ||
| Omega-3 fatty acids | [46] | Drp1, Fis1 | Fission | Separation of the MT into two smaller units. Fission allows the isolation of damaged MT parts for elimination, but it becomes massive in the case of apoptosis. |
| 1H-pyrrole-2-carboxamide compounds (synthetic) | [34] | Drp1 | ||
| Astaxanthin | [42,50] | Drp1 | ||
| Curcumin, astaxanthin, resveratrol, hydroxytyrosol, oleuropein, spermidine | [51] | Modulation of several mitophagy mediators | Mitophagy | Autophagic degradation of irreversibly damaged MTs or part of them. |
| Astaxanthin | [42] | PINK, parkin | ||
| Fucoxanthin | [49] | Pink1, Prk2, Bnip3, BECN1, Atg5 |
| Factor/Protein/Compound Able to Regulate Target Enzyme or Its Gene Expression | Effect | Ref. | Target Enzyme | Target Enzyme Function |
|---|---|---|---|---|
| PPARγ | Up | [71] | Catalase | Hydrogen peroxide (H2O2) decomposition to oxygen (O2) and water (H2O). |
| Oct-1 | Up | [71] | ||
| Astaxanthin | Up | [72] | ||
| DHA (via Nrf2) | Up | [73] | ||
| p53, PIG3 | Down | [74] | ||
| MicroRNA-30b | Up | [75] | ||
| NF-κB, (Sp1), C/EBP, SIRT, FOXO3a, CREB | Up | [73,76,77,78] | Mn-Superoxide dismutase (SOD2) | Manganese enzyme expressed in the inner matrix catalyzes the dismutation of the superoxide radical (O2•−) into ordinary molecular oxygen (O2) and hydrogen peroxide (H2O2). |
| AP-1 | Up | [79] | ||
| AP-2 | Down | [76] | ||
| p53, p50 | Down | [78,80] | ||
| miR-146a | Down | [81] | ||
| Quercetin | Down | [82] | ||
| Curcumin | Up | [83] | ||
| Astaxanthin | Up | [72] | ||
| Sp1, C/EBP, Egr1, Nrf2, NF-κB, ELAV-like proteins, resveratrol | Up | [84,85,86,87] | Cu,Zn-Superoxyde dismutase (SOD1) | Copper-zinc enzyme with the same function as SOD-2 but expressed in the inter-membrane space [77,88]. |
| AP-1 | Down | [78,89] | ||
| Quercetin | Down | [82] | ||
| ARE/EpRE | Up | [90] | Peroxiredoxins (Prx3) | Enzymes are able to catalyze the oxidation of the redox-active cysteine (i.e., peroxidatic cysteine) to a sulfenic acid by the peroxide substrate. |
| Angiotensin II | Down | [91] | ||
| SOD2 | Up | [92] | Thioredoxin (TNX2) | Enzymes are expressed by a nuclear gene and imported into the mitochondrion, which carries out ROS scavenging activity with the concomitant anti-apoptotic effect [93]. |
| Curcumin | Up | [83] | ||
| Resveratrol | Up | [85] | Glutathione peroxidase-1(GPx-1) | A selenocysteine-containing enzyme involved in the reductive detoxification of peroxides. Its expression seems stimulated by the epidermal growth factor (EGF). |
| Genistein | Up | [94] | ||
| Quercetin | Down | [82] | ||
| Resveratrol | Up | [85] | Glutathione (GSH) | It protects the cell from respiration-induced reactive oxygen species and detoxifies lipid hydroperoxides and electrophiles. |
| Quercetin | Down | [45] | ||
| Nrf2/Nrf1 via ARE, AP-1 and NF-kB | Up | [95] | ||
| Procyanidin B2 (upregulation of P1 isoform via nuclear translocation of Nrf2) | Up | [96] | Glutathione-S- transferases (GSTs) | Mitochondrial GSTs display both GSH transferase and peroxidase activities for the detoxification of harmful byproducts [97]. |
| Obtusilactone A (OA) and (−)-sesamin | Down | [98] | Lon proteases | They decompose damaged and misfolded proteins tagged for degradation at their –COOH or –NH3 terminus [57]. Mitochondrial biogenesis in mammalian cells is partly regulated by the matrix Lon protease [99]. |
| Acute stressors, such as heat shock, serum starvation, and oxidative stress (Nrf-2, HIF-1) | Up | [100] | ||
| Only synthetic molecules are known: β-lactones (A2-32-01); Phenyl esters (AV167, TG42, TG53); α-aminoboronic acid | Down | [101] | Clp proteases | Variants of chaperon ATPase subunits (ClpA, ClpC, ClpE etc.) combined with a proteolytic subunit (ClpP) [57]. |
| Only synthetic molecules are known: Acyldepsipeptide analogs (ADEP-41); imipridones (ONC201, ONC212, TR57) | Up | [101] |
Mitochondrial Apoptosis and the Role of BCL-2 Family Proteins
3. Conceptualization of the Functionality of Foods and Implications of Their Active Compounds
- In the EU [117]: β-carotene and a mixture of carotenoids from Dunaliella salina; oil rich in PUFAs from Ulkenia sp.; DHA and EPA ethyl esters oil from Schizochytrium sp.; astaxanthin-rich oleoresin from Haematococcus pluvialis; oil rich in EPA from Phaeodactylum tricornutum;
- National Health Commission (People’s Republic of China): DHA oil from Schizochytrium sp., Ulkenia amoeboida, Crypthecodinium cohnii.
3.1. Protective Features of Crude Extracts of Microalgae
3.2. Activity of Long-Chain Polyunsaturated Fatty Acids (PUFAs)
3.3. Sterols
3.4. Phenolic Compounds
| Species | Total Phenolic Content mg GAE/g Biomass DW | Reference |
|---|---|---|
| Euglena cantabrica | 0.6–12.6 | [157] |
| Demodesmus sp. | 7.7 | [158] |
| Tetraselmis suecica | 4.3 | [151] |
| Dunaliella salina | 4.5 | [158] |
| Haematococcus pluvialis | 1.9 | [151] |
| Nannochloropsis limnetica | 5.8 | [158] |
| Nannochloropsis salina | 6.5 | [158] |
| Galdieria sulphuraria | 1.6–5.3 | [159] |
| Nannochloropsis sp. | 2.2 | [151] |
| Isochrysis sp. | 7.8 | [151] |
| Chaetoceros calcitrans | 2.3 | [151] |
| Porphyridium cruentum | 1 | [151] |
| Phaeodactylum tricornutum | 3.2–6.1 | [151,158] |
| Chlorella sorokiniana | 5.8–5.9 | [158] |
| Auxenochlorella pyrenoidosa | 13.2–25.8 | [160] |
| Arthrospira platensis | 17–43.2 | [160] |
| Arthrospira fusiformis | 47.3–88.5 | [161] |
| Nostoc commune | 0.9 | [162] |
| Phenolic Compound | Euglena cantabrica | Desmodesmus sp. | Tetraselmis suecica | Dunaliella salina | Haematoc. pluvialis | Nannochl. limnetica | Microchl. salina | Galdieria sulphuraria | Nannochloropsis sp. | Diacronema lutheri | Porph. purpureum | Phaeod. tricornutum | Chlorella sorokiniana | Auxenoc. pyrenoidosa | Arthrospira platensis | Arthrospira sp. | Nostoc commune |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Phloroglucinol | + | + | + | + | + | + | + | + | |||||||||
| Pyrocatechol | + | ||||||||||||||||
| Pyrogallol | + | ||||||||||||||||
| Gallic ac. | + | + | + | + | + | + | + | + | + | + | |||||||
| 4-Hydroxy benzoic ac. | + | + | + | + | |||||||||||||
| 3,4-Dihydroxy benzoic ac. | + | + | + | ||||||||||||||
| Protocatechuic ac. | + | + | + | ||||||||||||||
| Quinic ac. | + | ||||||||||||||||
| Salicylic ac. | + | + | + | + | + | ||||||||||||
| Syringic ac. | + | + | + | + | |||||||||||||
| Vanillic ac. | + | + | |||||||||||||||
| Vanillin | + | + | + | ||||||||||||||
| 4-Aminobenzoic ac. | + | ||||||||||||||||
| Caffeic ac. | + | + | + | + | + | + | + | + | |||||||||
| Cinnamic ac. | + | + | + | ||||||||||||||
| Ferulic ac. | + | + | + | + | + | + | + | + | + | + | + | + | |||||
| 2/3/4-Hydroxy-cinnamic ac. | + | ||||||||||||||||
| P-Coumaric ac. | + | + | + | + | + | + | + | + | + | + | + | ||||||
| Chlorogenic ac. | + | + | + | + | + | ||||||||||||
| Phloretin | + | ||||||||||||||||
| Rosmarinic ac. | + | ||||||||||||||||
| Apigenin | + | + | + | + | + | + | |||||||||||
| Catechin hydrate | + | + | + | ||||||||||||||
| Daidzein | + | + | + | ||||||||||||||
| Dihydrokaempferol | + | ||||||||||||||||
| Dihydroquercetin | + | ||||||||||||||||
| Epicatechin | + | + | |||||||||||||||
| Epigallocatechin | + | ||||||||||||||||
| Genistein | + | + | + | ||||||||||||||
| Kaempferol | + | ||||||||||||||||
| Luteolin | + | + | |||||||||||||||
| Naringenin | + | + | |||||||||||||||
| Quercetin | + | + | + | + | + | ||||||||||||
| Ellagic ac. | + | + | |||||||||||||||
| Rutin | + | ||||||||||||||||
| Resveratrol | + | ||||||||||||||||
| References | [150] | [158] | [150,184] | [158] | [184] | [158] | [158] | [172] | [153] | [184] | [184] | [142,150,176] | [158] | [160] | [160,184,185,186] | [153] | [150] |
3.5. Carotenoids
| Major Component | Species | Compound Concentration | Ref. | Total Carotenoid Concentration | Ref. |
|---|---|---|---|---|---|
| β-carotene | Dunaliella salina | Up to 10–13% DW | [201] | up to 29% | [188] |
| Tetraselmis suecica | 0.1% DW | 0.35–1.1% DW | [202] | ||
| Vischeria stellata | 5.9% DW | [203] | 7.7% DW | [203] | |
| Chromochloris zofingiensis | 0.9% DW | [201] | 0.7–0.88% DW | [204,205] | |
| Astaxanthin | Haematococcus pluvialis | 2.3–7.7% DW | [206] | astaxanthin accounts for 85–90% of total carotenoids [207] | |
| Up to 5% DW | [188] | ||||
| Chromochloris zofingiensis | 0.3–0.6% DW | [208] | 0.7% DW | [204] | |
| 0.53–0.6% DW | [188] | 0.7% DW | [204] | ||
| Canthaxanthin | Coelastrella striolata var. multistriata | 4.75% DW | [205] | 5.6% DW | [205] |
| Fucoxanthin | Isochrysis aff. galbana | 1.7–2.1% DW | [209] | 2% DW | [209] |
| Mallomonas sp. SBV13 | 2.6% | [210] | |||
| Isochrysis galbana | 0.22–1.35% DW | [130,211] | 1.76% DW | [211] | |
| Odontella aurita | up to 2.2% DW | [201] | ~1.5% DW | [212] | |
| Phaeodactylum tricornutum | 0.78–1.65% DW | [201,211] | 0.61–1% DW | [151,211] | |
| Lutein | Auxenochlorella protothecoides | 0.54 DW | [201] | 0.8% DW | [213] |
| Chlorella sorokiniana | 0.21–0.32% DW | [158] | 0.4% DW | [214] | |
| Coelastrella sp. | 0.69% DW | [201] | |||
| Desmodesmus sp. | 0.51% DW | [158] | 0.67% DW | [158] | |
| Chromochloris zofingiensis (mutant strain) | 1.38% DW | [215] | 2.74% DW | [215] | |
| Scenedesmus almeriensis | 0.54% DW | [201] | |||
| Violaxanthin | Nannochloropsis sp. | 0.12–0.58% DW | [117] | 0.30–0.86% DW | [117] |
| Zeaxanthin | Chloroidium saccharophila | 1.1% DW | [216] | 1.6% DW | [217] |
| Chloroidium ellipsoideum | 0.42% DW | [216] | |||
| Chromochloris zofingiensis (mutant strain) | 0.7% DW | [215] | 2.74% DW | [215] | |
| Dunaliella salina (mutant strain) | 0.42–0.59% DW | [218] | 1.1–1.28% DW | [218] | |
- Activation of the PI3K/Akt (phosphatidylinositol 3-kinase; AKT-serine/threonine kinase also known as PKB, protein kinase B) survival pathway, which inactivates Bax by phosphorylation and reduces the release of Cyt-c (antiapoptotic effect). Indeed, according to Kale et al. [105], the phosphorylation at residue S184 by Akt inhibits Bax, thus preventing its translocation into the mitochondrion.
- Activation of the p38 MAPK signaling pathway, which promotes the release of Cyt-c and activates the apoptosome (apoptotic effect). Specifically, p38 MAPK can act on the mitochondrial permeability by activating Bim by phosphorylation, which in turn activates Bax or, alternatively, it can phosphorylate p53, which induces the expression of death receptors and can activate members of the BCL-2 family to promote apoptosis [228]. Other proapoptotic mechanisms of action have been hypothesized for p38MAPK, beyond the scope of the present review.
- Stimulation of MEK1/2–ERK1/2 signaling pathway, which in turn activates the pro-survival BCL-2 proteins (antiapoptotic effect), as shortly explained in the first part of this contribution.
3.6. Other Bioactive Compounds
4. Discussion and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bloom, D.E.; Luca, D.L. Chapter 1—The Global Demography of Aging: Facts, Explanations, Future. In Handbook of the Economics of Population Aging; Piggott, J., Woodland, A., Eds.; North-Holland: Amsterdam, The Netherlands, 2016; Volume 1, pp. 3–56. [Google Scholar]
- de Meijer, C.; Wouterse, B.; Polder, J.; Koopmanschap, M. The Effect of Population Aging on Health Expenditure Growth: A Critical Review. Eur. J. Ageing 2013, 10, 353–361. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; He, Y.; Lyu, J.; Yu, C.; Bian, M.; Lee, L. Aging in China: Perspectives on Public Health. Glob. Health J. 2020, 4, 11–17. [Google Scholar] [CrossRef]
- Kaplan, M.; Inguanzo, M. The Social, Economic, and Public Health Consequences of Global Population Aging: Implications for Social Work Practice and Public Policy. J. Soc. Work Glob. Community 2017, 2, 1–12. [Google Scholar] [CrossRef]
- Vancea, M.; Solé-Casals, J. Population Aging in the European Information Societies: Towards a Comprehensive Research Agenda in EHealth Innovations for Elderly. Aging Dis. 2015, 7, 526–539. [Google Scholar] [CrossRef]
- Meydani, M. Nutrition Interventions in Aging and Age-Associated Disease. Ann. N. Y. Acad. Sci. 2001, 928, 226–235. [Google Scholar] [CrossRef] [PubMed]
- Miyazawa, T.; Abe, C.; Burdeos, G.C.; Matsumoto, A.; Toda, M. Food Antioxidants and Aging: Theory, Current Evidence and Perspectives. Nutraceuticals 2022, 2, 181–204. [Google Scholar] [CrossRef]
- García-Montero, C.; Fraile-Martínez, O.; Gómez-Lahoz, A.M.; Pekarek, L.; Castellanos, A.J.; Noguerales-Fraguas, F.; Coca, S.; Guijarro, L.G.; García-Honduvilla, N.; Asúnsolo, A.; et al. Nutritional Components in Western Diet Versus Mediterranean Diet at the Gut Microbiota–Immune System Interplay. Implications for Health and Disease. Nutrients 2021, 13, 699. [Google Scholar] [CrossRef]
- Cordain, L.; Eaton, S.B.; Sebastian, A.; Mann, N.; Lindeberg, S.; Watkins, B.A.; O’Keefe, J.H.; Brand-Miller, J. Origins and Evolution of the Western Diet: Health Implications for the 21st Century. Am. J. Clin. Nutr. 2005, 81, 341–354. [Google Scholar] [CrossRef]
- Castillo, M.; Iriondo-DeHond, A.; Martirosyan, D. Are Functional Foods Essential for Sustainable Health? Ann. Nutr. Food Sci. 2018, 2, 1015. [Google Scholar]
- Milner, J.A. Functional Foods and Health Promotion. J. Nutr. 1999, 129, 1395S–1397S. [Google Scholar] [CrossRef]
- Meléndez-Martínez, A.J.; Böhm, V.; Borge, G.I.A.; Cano, M.P.; Fikselová, M.; Gruskiene, R.; Lavelli, V.; Loizzo, M.R.; Mandić, A.I.; Brahm, P.M.; et al. Carotenoids: Considerations for Their Use in Functional Foods, Nutraceuticals, Nutricosmetics, Supplements, Botanicals, and Novel Foods in the Context of Sustainability, Circular Economy, and Climate Change. Annu. Rev. Food Sci. Technol. 2021, 12, 433–460. [Google Scholar] [CrossRef]
- Barkia, I.; Saari, N.; Manning, S.R. Microalgae for High-Value Products Towards Human Health and Nutrition. Mar. Drugs 2019, 17, 304. [Google Scholar] [CrossRef]
- Basheer, S.; Huo, S.; Zhu, F.; Qian, J.; Xu, L.; Cui, F.; Zou, B. Microalgae in Human Health and Medicine. In Microalgae Biotechnology for Food, Health and High Value Products; Alam, M.A., Xu, J.-L., Wang, Z., Eds.; Springer: Singapore, 2020; pp. 149–174. ISBN 9789811501692. [Google Scholar]
- Del Mondo, A.; Smerilli, A.; Sané, E.; Sansone, C.; Brunet, C. Challenging Microalgal Vitamins for Human Health. Microb. Cell Factories 2020, 19, 201. [Google Scholar] [CrossRef]
- Matos, J.; Cardoso, C.; Bandarra, N.M.; Afonso, C. Microalgae as Healthy Ingredients for Functional Food: A Review. Food Funct. 2017, 8, 2672–2685. [Google Scholar] [CrossRef]
- Abd El-Hack, M.E.; Abdelnour, S.; Alagawany, M.; Abdo, M.; Sakr, M.A.; Khafaga, A.F.; Mahgoub, S.A.; Elnesr, S.S.; Gebriel, M.G. Microalgae in Modern Cancer Therapy: Current Knowledge. Biomed. Pharmacother. 2019, 111, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Andrade, L.M.; Andrade, C.J.; Dias, M.; Nascimento, C.A.O.; Mendes, M.A. Chlorella and Spirulina Microalgae as Sources of Functional Foods, Nutraceuticals, and Food Supplements; an Overview. MOJ Food Process. Technol. 2018, 6, 45–58. [Google Scholar] [CrossRef]
- Guerin, M.; Huntley, M.E.; Olaizola, M. Haematococcus Astaxanthin: Applications for Human Health and Nutrition. Trends Biotechnol. 2003, 21, 210–216. [Google Scholar] [CrossRef]
- Napolitano, G.; Fasciolo, G.; Salbitani, G.; Venditti, P. Chlorella sorokiniana Dietary Supplementation Increases Antioxidant Capacities and Reduces ROS Release in Mitochondria of Hyperthyroid Rat Liver. Antioxidants 2020, 9, 883. [Google Scholar] [CrossRef] [PubMed]
- Wells, M.L.; Potin, P.; Craigie, J.S.; Raven, J.A.; Merchant, S.S.; Helliwell, K.E.; Smith, A.G.; Camire, M.E.; Brawley, S.H. Algae as Nutritional and Functional Food Sources: Revisiting Our Understanding. J. Appl. Phycol. 2017, 29, 949–982. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, S. Chapter Two—Oxidative Stress, Inflammation, and Disease. In Oxidative Stress and Biomaterials; Dziubla, T., Butterfield, D.A., Eds.; Academic Press: Cambridge, MA, USA, 2016; pp. 35–58. ISBN 978-0-12-803269-5. [Google Scholar]
- Rani, V.; Deep, G.; Singh, R.K.; Palle, K.; Yadav, U.C.S. Oxidative Stress and Metabolic Disorders: Pathogenesis and Therapeutic Strategies. Life Sci. 2016, 148, 183–193. [Google Scholar] [CrossRef]
- Hajam, Y.A.; Rani, R.; Ganie, S.Y.; Sheikh, T.A.; Javaid, D.; Qadri, S.S.; Pramodh, S.; Alsulimani, A.; Alkhanani, M.F.; Harakeh, S.; et al. Oxidative Stress in Human Pathology and Aging: Molecular Mechanisms and Perspectives. Cells 2022, 11, 552. [Google Scholar] [CrossRef] [PubMed]
- Murphy, E.; Ardehali, H.; Balaban, R.S.; DiLisa, F.; Dorn, G.W.; Kitsis, R.N.; Otsu, K.; Ping, P.; Rizzuto, R.; Sack, M.N.; et al. Mitochondrial Function, Biology, and Role in Disease. Circ. Res. 2016, 118, 1960–1991. [Google Scholar] [CrossRef]
- Murphy, M.P.; Hartley, R.C. Mitochondria as a Therapeutic Target for Common Pathologies. Nat. Rev. Drug Discov. 2018, 17, 865–886. [Google Scholar] [CrossRef]
- Sukhorukov, V.M.; Dikov, D.; Reichert, A.S.; Meyer-Hermann, M. Emergence of the Mitochondrial Reticulum from Fission and Fusion Dynamics. PLoS Comput. Biol. 2012, 8, e1002745. [Google Scholar] [CrossRef]
- Hock, M.B.; Kralli, A. Transcriptional Control of Mitochondrial Biogenesis and Function. Annu. Rev. Physiol. 2009, 71, 177–203. [Google Scholar] [CrossRef]
- Gureev, A.P.; Shaforostova, E.A.; Popov, V.N. Regulation of Mitochondrial Biogenesis as a Way for Active Longevity: Interaction Between the Nrf2 and PGC-1α Signaling Pathways. Front. Genet. 2019, 10, 435. [Google Scholar] [CrossRef]
- Yun, J.; Finkel, T. Mitohormesis. Cell Metab. 2014, 19, 757–766. [Google Scholar] [CrossRef] [PubMed]
- Westermann, B. Bioenergetic Role of Mitochondrial Fusion and Fission. Biochim. Biophys. Acta BBA Bioenerg. 2012, 1817, 1833–1838. [Google Scholar] [CrossRef]
- Robb, E.L.; Moradi, F.; Maddalena, L.A.; Valente, A.J.F.; Fonseca, J.; Stuart, J.A. Resveratrol Stimulates Mitochondrial Fusion by a Mechanism Requiring Mitofusin-2. Biochem. Biophys. Res. Commun. 2017, 485, 249–254. [Google Scholar] [CrossRef]
- Nicholls, T.J.; Gustafsson, C.M. Separating and Segregating the Human Mitochondrial Genome. Trends Biochem. Sci. 2018, 43, 869–881. [Google Scholar] [CrossRef] [PubMed]
- Mallat, A.; Uchiyama, L.F.; Lewis, S.C.; Fredenburg, R.A.; Terada, Y.; Ji, N.; Nunnari, J.; Tseng, C.C. Discovery and Characterization of Selective Small Molecule Inhibitors of the Mammalian Mitochondrial Division Dynamin, DRP1. Biochem. Biophys. Res. Commun. 2018, 499, 556–562. [Google Scholar] [CrossRef]
- Otera, H.; Ishihara, N.; Mihara, K. New Insights into the Function and Regulation of Mitochondrial Fission. Biochim. Biophys. Acta BBA Mol. Cell Res. 2013, 1833, 1256–1268. [Google Scholar] [CrossRef]
- Kageyama, Y.; Zhang, Z.; Sesaki, H. Mitochondrial Division: Molecular Machinery and Physiological Functions. Curr. Opin. Cell Biol. 2011, 23, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Frank, M.; Duvezin-Caubet, S.; Koob, S.; Occhipinti, A.; Jagasia, R.; Petcherski, A.; Ruonala, M.O.; Priault, M.; Salin, B.; Reichert, A.S. Mitophagy Is Triggered by Mild Oxidative Stress in a Mitochondrial Fission Dependent Manner. Biochim. Biophys. Acta BBA Mol. Cell Res. 2012, 1823, 2297–2310. [Google Scholar] [CrossRef]
- Youle, R.J.; Narendra, D.P. Mechanisms of Mitophagy. Nat. Rev. Mol. Cell Biol. 2011, 12, 9–14. [Google Scholar] [CrossRef]
- Tanaka, K. The PINK1–Parkin Axis: An Overview. Neurosci. Res. 2020, 159, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Ding, W.-X.; Yin, X.-M. Mitophagy: Mechanisms, Pathophysiological Roles, and Analysis. Biol. Chem. 2012, 393, 547–564. [Google Scholar] [CrossRef]
- Liu, P.H.; Aoi, W.; Takami, M.; Terajima, H.; Tanimura, Y.; Naito, Y.; Itoh, Y.; Yoshikawa, T. The Astaxanthin-Induced Improvement in Lipid Metabolism during Exercise Is Mediated by a PGC-1α Increase in Skeletal Muscle. J. Clin. Biochem. Nutr. 2014, 54, 86–89. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Li, S.; Guo, Y.; Yu, H.; Bao, Y.; Xin, X.; Yang, H.; Ni, X.; Wu, N.; Jia, D. Astaxanthin Attenuates Hypertensive Vascular Remodeling by Protecting Vascular Smooth Muscle Cells from Oxidative Stress-Induced Mitochondrial Dysfunction. Oxidative Med. Cell. Longev. 2020, 2020, 4629189. [Google Scholar] [CrossRef]
- Laiglesia, L.M.; Lorente-Cebrián, S.; Prieto-Hontoria, P.L.; Fernández-Galilea, M.; Ribeiro, S.M.R.; Sáinz, N.; Martínez, J.A.; Moreno-Aliaga, M.J. Eicosapentaenoic Acid Promotes Mitochondrial Biogenesis and Beige-like Features in Subcutaneous Adipocytes from Overweight Subjects. J. Nutr. Biochem. 2016, 37, 76–82. [Google Scholar] [CrossRef]
- Flachs, P.; Horakova, O.; Brauner, P.; Rossmeisl, M.; Pecina, P.; Franssen-van Hal, N.; Ruzickova, J.; Sponarova, J.; Drahota, Z.; Vlcek, C.; et al. Polyunsaturated Fatty Acids of Marine Origin Upregulate Mitochondrial Biogenesis and Induce β-Oxidation in White Fat. Diabetologia 2005, 48, 2365–2375. [Google Scholar] [CrossRef] [Green Version]
- Gibellini, L.; Bianchini, E.; De Biasi, S.; Nasi, M.; Cossarizza, A.; Pinti, M. Natural Compounds Modulating Mitochondrial Functions. Evid.-Based Complement. Altern. Med. ECAM 2015, 2015, 527209. [Google Scholar] [CrossRef]
- Lepretti, M.; Martucciello, S.; Burgos Aceves, M.A.; Putti, R.; Lionetti, L. Omega-3 Fatty Acids and Insulin Resistance: Focus on the Regulation of Mitochondria and Endoplasmic Reticulum Stress. Nutrients 2018, 10, 350. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.-T.; Chou, H.-N.; Huang, C. Dietary Fucoxanthin Increases Metabolic Rate and Upregulated MRNA Expressions of the PGC-1alpha Network, Mitochondrial Biogenesis and Fusion Genes in White Adipose Tissues of Mice. Mar. Drugs 2014, 12, 964–982. [Google Scholar] [CrossRef] [PubMed]
- Mao, G.-X.; Xu, X.-G.; Wang, S.-Y.; Li, H.-F.; Zhang, J.; Zhang, Z.-S.; Su, H.-L.; Chen, S.-S.; Xing, W.-M.; Wang, Y.-Z.; et al. Salidroside Delays Cellular Senescence by Stimulating Mitochondrial Biogenesis Partly through a MiR-22/SIRT-1 Pathway. Oxid. Med. Cell. Longev. 2019, 2019, e5276096. [Google Scholar] [CrossRef]
- Li, S.; Ren, X.; Wang, Y.; Hu, J.; Wu, H.; Song, S.; Yan, C. Fucoxanthin Alleviates Palmitate-Induced Inflammation in RAW 264.7 Cells through Improving Lipid Metabolism and Attenuating Mitochondrial Dysfunction. Food Funct. 2020, 11, 3361–3370. [Google Scholar] [CrossRef]
- Zhang, J.; Xu, P.; Wang, Y.; Wang, M.; Li, H.; Lin, S.; Mao, C.; Wang, B.; Song, X.; Lv, C. Astaxanthin Prevents Pulmonary Fibrosis by Promoting Myofibroblast Apoptosis Dependent on Drp1-Mediated Mitochondrial Fission. J. Cell. Mol. Med. 2015, 19, 2215–2231. [Google Scholar] [CrossRef]
- Varghese, N.; Werner, S.; Grimm, A.; Eckert, A. Dietary Mitophagy Enhancer: A Strategy for Healthy Brain Aging? Antioxidants 2020, 9, 932. [Google Scholar] [CrossRef] [PubMed]
- Gleyzer, N.; Vercauteren, K.; Scarpulla, R.C. Control of Mitochondrial Transcription Specificity Factors (TFB1M and TFB2M) by Nuclear Respiratory Factors (NRF-1 and NRF-2) and PGC-1 Family Coactivators. Mol. Cell. Biol. 2005, 25, 1354–1366. [Google Scholar] [CrossRef]
- Nicholls, D.G. Mitochondrial Function and Dysfunction in the Cell: Its Relevance to Aging and Aging-Related Disease. Int. J. Biochem. Cell Biol. 2002, 34, 1372–1381. [Google Scholar] [CrossRef]
- Ortiz, G.G.; Mireles-Ramírez, M.A.; González-Usigli, H.; Macías-Islas, M.A.; Bitzer-Quintero, O.K.; DheniTorres-Sánchez, E.; Sánchez-López, A.L.; Ramírez-Jirano, J.; Ríos-Silva, M.; Torres-Mendoza, B. Mitochondrial Aging and Metabolism: The Importance of a Good Relationship in the Central Nervous System; IntechOpen: London, UK, 2018; ISBN 978-1-78984-266-1. [Google Scholar]
- Srivastava, S. The Mitochondrial Basis of Aging and Age-Related Disorders. Genes 2017, 8, 398. [Google Scholar] [CrossRef] [Green Version]
- Brand, M.D.; Orr, A.L.; Perevoshchikova, I.V.; Quinlan, C.L. The Role of Mitochondrial Function and Cellular Bioenergetics in Ageing and Disease. Br. J. Dermatol. 2013, 169, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Hamon, M.-P.; Bulteau, A.-L.; Friguet, B. Mitochondrial Proteases and Protein Quality Control in Ageing and Longevity. Ageing Res. Rev. 2015, 23, 56–66. [Google Scholar] [CrossRef] [PubMed]
- Sastre, J.; Serviddio, G.; Pereda, J.; Minana, J.B.; Arduini, A.; Vendemiale, G.; Poli, G.; Pallardo, F.V.; Vina, J. Mitochondrial Function in Liver Disease. Front. Biosci.-Landmark 2007, 12, 1200–1209. [Google Scholar] [CrossRef]
- Serviddio, G.; Bellanti, F.; Vendemiale, G.; Altomare, E. Mitochondrial Dysfunction in Nonalcoholic Steatohepatitis. Expert Rev. Gastroenterol. Hepatol. 2011, 5, 233–244. [Google Scholar] [CrossRef] [PubMed]
- Song, B.-J.; Moon, K.-H.; Olsson, N.U.; Salem, N. Prevention of Alcoholic Fatty Liver and Mitochondrial Dysfunction in the Rat by Long-Chain Polyunsaturated Fatty Acids. J. Hepatol. 2008, 49, 262–273. [Google Scholar] [CrossRef]
- Videla, L.A.; Rodrigo, R.; Araya, J.; Poniachik, J. Oxidative Stress and Depletion of Hepatic Long-Chain Polyunsaturated Fatty Acids May Contribute to Nonalcoholic Fatty Liver Disease. Free Radic. Biol. Med. 2004, 37, 1499–1507. [Google Scholar] [CrossRef]
- Hirabara, S.M.; Curi, R.; Maechler, P. Saturated Fatty Acid-Induced Insulin Resistance Is Associated with Mitochondrial Dysfunction in Skeletal Muscle Cells. J. Cell. Physiol. 2010, 222, 187–194. [Google Scholar] [CrossRef]
- Jheng, H.-F.; Tsai, P.-J.; Guo, S.-M.; Kuo, L.-H.; Chang, C.-S.; Su, I.-J.; Chang, C.-R.; Tsai, Y.-S. Mitochondrial Fission Contributes to Mitochondrial Dysfunction and Insulin Resistance in Skeletal Muscle. Mol. Cell. Biol. 2012, 32, 309–319. [Google Scholar] [CrossRef]
- Che, R.; Yuan, Y.; Huang, S.; Zhang, A. Mitochondrial Dysfunction in the Pathophysiology of Renal Diseases. Am. J. Physiol.-Ren. Physiol. 2014, 306, F367–F378. [Google Scholar] [CrossRef]
- Paradies, G.; Paradies, V.; Ruggiero, F.M.; Petrosillo, G. Role of Cardiolipin in Mitochondrial Function and Dynamics in Health and Disease: Molecular and Pharmacological Aspects. Cells 2019, 8, 728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zharikov, S.; Shiva, S. Platelet Mitochondrial Function: From Regulation of Thrombosis to Biomarker of Disease. Biochem. Soc. Trans. 2013, 41, 118–123. [Google Scholar] [CrossRef] [PubMed]
- Jenner, P. Altered Mitochondrial Function, Iron Metabolism and Glutathione Levels in Parkinson’s Disease. Acta Neurol. Scand. 1993, 87, 6–13. [Google Scholar] [CrossRef]
- Turner, C.; Cooper, J.M.; Schapira, A.H.V. Clinical Correlates of Mitochondrial Function in Huntington’s Disease Muscle. Mov. Disord. 2007, 22, 1715–1721. [Google Scholar] [CrossRef]
- Eckert, G.P.; Renner, K.; Eckert, S.H.; Eckmann, J.; Hagl, S.; Abdel-Kader, R.M.; Kurz, C.; Leuner, K.; Muller, W.E. Mitochondrial Dysfunction—A Pharmacological Target in Alzheimer’s Disease. Mol. Neurobiol. 2012, 46, 136–150. [Google Scholar] [CrossRef] [PubMed]
- Catalgol, B.; Batirel, S.; Taga, Y.; Ozer, N. Resveratrol: French Paradox Revisited. Front. Pharmacol. 2012, 3, 141. [Google Scholar] [CrossRef]
- Kodydková, J.; Vávrová, L.; Kocík, M.; Žák, A. Human Catalase, Its Polymorphisms, Regulation and Changes of Its Activity in Different Diseases. Folia Biol. 2014, 60, 153–167. [Google Scholar]
- Lee, D.-H.; Kim, C.-S.; Lee, Y.J. Astaxanthin Protects against MPTP/MPP+-Induced Mitochondrial Dysfunction and ROS Production in Vivo and in Vitro. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2011, 49, 271–280. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, L.; Sun, D.; He, Y.; Jiang, Y.; Cheng, K.-W.; Chen, F. DHA Protects against Monosodium Urate-Induced Inflammation through Modulation of Oxidative Stress. Food Funct. 2019, 10, 4010–4021. [Google Scholar] [CrossRef]
- Kang, M.Y.; Kim, H.-B.; Piao, C.; Lee, K.H.; Hyun, J.W.; Chang, I.-Y.; You, H.J. The Critical Role of Catalase in Prooxidant and Antioxidant Function of P53. Cell Death Differ. 2013, 20, 117–129. [Google Scholar] [CrossRef]
- Haque, R.; Chun, E.; Howell, J.C.; Sengupta, T.; Chen, D.; Kim, H. MicroRNA-30b-Mediated Regulation of Catalase Expression in Human ARPE-19 Cells. PLoS ONE 2012, 7, e42542. [Google Scholar] [CrossRef]
- Xu, Y.; Porntadavity, S.; St Clair, D.K. Transcriptional Regulation of the Human Manganese Superoxide Dismutase Gene: The Role of Specificity Protein 1 (Sp1) and Activating Protein-2 (AP-2). Biochem. J. 2002, 362, 401–412. [Google Scholar] [CrossRef]
- Kim, Y.S.; Gupta Vallur, P.; Phaëton, R.; Mythreye, K.; Hempel, N. Insights into the Dichotomous Regulation of SOD2 in Cancer. Antioxidants 2017, 6, 86. [Google Scholar] [CrossRef]
- Miao, L.; St. Clair, D.K. Regulation of Superoxide Dismutase Genes: Implications in Diseases. Free Radic. Biol. Med. 2009, 47, 344–356. [Google Scholar] [CrossRef]
- Rui, T.; Kvietys, P.R. NFkappaB and AP-1 Differentially Contribute to the Induction of Mn-SOD and ENOS during the Development of Oxidant Tolerance. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2005, 19, 1908–1910. [Google Scholar] [CrossRef]
- Drane, P.; Bravard, A.; Bouvard, V.; May, E. Reciprocal Down-Regulation of P53 and SOD2 Gene Expression–Implication in P53 Mediated Apoptosis. Oncogene 2001, 20, 430–439. [Google Scholar] [CrossRef] [PubMed]
- Ji, G.; Lv, K.; Chen, H.; Wang, T.; Wang, Y.; Zhao, D.; Qu, L.; Li, Y. MiR-146a Regulates SOD2 Expression in H2O2 Stimulated PC12 Cells. PLoS ONE 2013, 8, e69351. [Google Scholar] [CrossRef] [PubMed]
- Röhrdanz, E.; Bittner, A.; Tran-Thi, Q.-H.; Kahl, R. The Effect of Quercetin on the MRNA Expression of Different Antioxidant Enzymes in Hepatoma Cells. Arch. Toxicol. 2003, 77, 506–510. [Google Scholar] [CrossRef]
- Zhang, J.; Bai, K.W.; He, J.; Niu, Y.; Lu, Y.; Zhang, L.; Wang, T. Curcumin Attenuates Hepatic Mitochondrial Dysfunction through the Maintenance of Thiol Pool, Inhibition of MtDNA Damage, and Stimulation of the Mitochondrial Thioredoxin System in Heat-Stressed Broilers. J. Anim. Sci. 2018, 96, 867–879. [Google Scholar] [CrossRef] [PubMed]
- Milani, P.; Amadio, M.; Laforenza, U.; Dell’Orco, M.; Diamanti, L.; Sardone, V.; Gagliardi, S.; Govoni, S.; Ceroni, M.; Pascale, A.; et al. Posttranscriptional Regulation of SOD1 Gene Expression under Oxidative Stress: Potential Role of ELAV Proteins in Sporadic ALS. Neurobiol. Dis. 2013, 60, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Spanier, G.; Xu, H.; Xia, N.; Tobias, S.; Deng, S.; Wojnowski, L.; Forstermann, U.; Li, H. Resveratrol Reduces Endothelial Oxidative Stress by Modulating the Gene Expression of Superoxide Dismutase 1 (SOD1), Glutathione Peroxidase 1 (GPx1) and NADPH Oxidase Subunit (Nox4). J. Physiol. Pharmacol. Off. J. Pol. Physiol. Soc. 2009, 60 (Suppl. 4), 111–116. [Google Scholar]
- Seo, S.J.; Kim, H.T.; Cho, G.; Rho, H.M.; Jung, G. Spl and C/EBP-Related Factor Regulate the Transcription of Human Cu/Zn SOD Gene. Gene 1996, 178, 177–185. [Google Scholar] [CrossRef]
- Minc, E.; de Coppet, P.; Masson, P.; Thiery, L.; Dutertre, S.; Amor-Guéret, M.; Jaulin, C. The Human Copper-Zinc Superoxide Dismutase Gene (SOD1) Proximal Promoter Is Regulated by Sp1, Egr-1, and WT1 via Non-Canonical Binding Sites*. J. Biol. Chem. 1999, 274, 503–509. [Google Scholar] [CrossRef] [Green Version]
- Okado-Matsumoto, A.; Fridovich, I. Subcellular Distribution of Superoxide Dismutases (SOD) in Rat Liver: Cu,Zn-SOD in Mitochondria. J. Biol. Chem. 2001, 276, 38388–38393. [Google Scholar] [CrossRef]
- Riera, H.; Afonso, V.; Collin, P.; Lomri, A. A Central Role for JNK/AP-1 Pathway in the Pro-Oxidant Effect of Pyrrolidine Dithiocarbamate through Superoxide Dismutase 1 Gene Repression and Reactive Oxygen Species Generation in Hematopoietic Human Cancer Cell Line U937. PLoS ONE 2015, 10, e0127571. [Google Scholar] [CrossRef] [PubMed]
- Ishii, T.; Yanagawa, T. Stress-Induced Peroxiredoxins. In Peroxiredoxin Systems: Structures and Functions; Flohé, L., Harris, J.R., Eds.; Subcellular Biochemistry; Springer: Dordrecht, The Netherlands, 2007; pp. 375–384. ISBN 978-1-4020-6051-9. [Google Scholar]
- Lijnen, P.J.; Piccart, Y.; Coenen, T.; Prihadi, J.S. Angiotensin II-Induced Mitochondrial Reactive Oxygen Species and Peroxiredoxin-3 Expression in Cardiac Fibroblasts. J. Hypertens. 2012, 30, 1986–1991. [Google Scholar] [CrossRef] [PubMed]
- Kim, A.; Joseph, S.; Khan, A.; Epstein, C.J.; Sobel, R.; Huang, T.-T. Enhanced Expression of Mitochondrial Superoxide Dismutase Leads to Prolonged in Vivo Cell Cycle Progression and Up-Regulation of Mitochondrial Thioredoxin. Free Radic. Biol. Med. 2010, 48, 1501–1512. [Google Scholar] [CrossRef] [PubMed]
- Li, K.-Y.; Xiang, X.-J.; Song, L.; Chen, J.; Luo, B.; Wen, Q.-X.; Zhong, B.-R.; Zhou, G.-F.; Deng, X.-J.; Ma, Y.-L.; et al. Mitochondrial TXN2 Attenuates Amyloidogenesis via Selective Inhibition of BACE1 Expression. J. Neurochem. 2021, 157, 1351–1365. [Google Scholar] [CrossRef]
- Suzuki, K.; Koike, H.; Matsui, H.; Ono, Y.; Hasumi, M.; Nakazato, H.; Okugi, H.; Sekine, Y.; Oki, K.; Ito, K.; et al. Genistein, a Soy Isoflavone, Induces Glutathione Peroxidase in the Human Prostate Cancer Cell Lines LNCaP and PC-3. Int. J. Cancer 2002, 99, 846–852. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.C. Regulation of Glutathione Synthesis. Mol. Aspects Med. 2009, 30, 42–59. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Ramiro, I.; Ramos, S.; Bravo, L.; Goya, L.; Martín, M.Á. Procyanidin B2 Induces Nrf2 Translocation and Glutathione S-Transferase P1 Expression via ERKs and P38-MAPK Pathways and Protect Human Colonic Cells against Oxidative Stress. Eur. J. Nutr. 2012, 51, 881–892. [Google Scholar] [CrossRef] [PubMed]
- Ribas, V.; García-Ruiz, C.; Fernández-Checa, J.C. Glutathione and Mitochondria. Front. Pharmacol. 2014, 5, 151. [Google Scholar] [CrossRef]
- Wang, H.-M.; Cheng, K.-C.; Lin, C.-J.; Hsu, S.-W.; Fang, W.-C.; Hsu, T.-F.; Chiu, C.-C.; Chang, H.-W.; Hsu, C.-H.; Lee, A.Y.-L. Obtusilactone A and (−)-Sesamin Induce Apoptosis in Human Lung Cancer Cells by Inhibiting Mitochondrial Lon Protease and Activating DNA Damage Checkpoints. Cancer Sci. 2010, 101, 2612–2620. [Google Scholar] [CrossRef]
- Luciakova, K.; Sokolikova, B.; Chloupkova, M.; Nelson, B.D. Enhanced Mitochondrial Biogenesis Is Associated with Increased Expression of the Mitochondrial ATP-Dependent Lon Protease. FEBS Lett. 1999, 444, 186–188. [Google Scholar] [CrossRef] [PubMed]
- Bota, D.A.; Davies, K.J.A. Mitochondrial Lon Protease in Human Disease and Aging: Including an Etiologic Classification of Lon-Related Diseases and Disorders. Free Radic. Biol. Med. 2016, 100, 188–198. [Google Scholar] [CrossRef]
- Nouri, K.; Feng, Y.; Schimmer, A.D. Mitochondrial ClpP Serine Protease-Biological Function and Emerging Target for Cancer Therapy. Cell Death Dis. 2020, 11, 841. [Google Scholar] [CrossRef]
- Peña-Blanco, A.; García-Sáez, A.J. Bax, Bak and beyond—Mitochondrial Performance in Apoptosis. FEBS J. 2018, 285, 416–431. [Google Scholar] [CrossRef] [PubMed]
- Roufayel, R.; Younes, K.; Al-Sabi, A.; Murshid, N. BH3-Only Proteins Noxa and Puma Are Key Regulators of Induced Apoptosis. Life 2022, 12, 256. [Google Scholar] [CrossRef]
- Happo, L.; Strasser, A.; Cory, S. BH3-Only Proteins in Apoptosis at a Glance. J. Cell Sci. 2012, 125, 1081–1087. [Google Scholar] [CrossRef]
- Kale, J.; Osterlund, E.J.; Andrews, D.W. BCL-2 Family Proteins: Changing Partners in the Dance towards Death. Cell Death Differ. 2018, 25, 65–80. [Google Scholar] [CrossRef]
- Cook, S.J.; Stuart, K.; Gilley, R.; Sale, M.J. Control of Cell Death and Mitochondrial Fission by ERK1/2 MAP Kinase Signalling. FEBS J. 2017, 284, 4177–4195. [Google Scholar] [CrossRef] [PubMed]
- Henry, C.J. Functional Foods. Eur. J. Clin. Nutr. 2010, 64, 657–659. [Google Scholar] [CrossRef]
- Domínguez Díaz, L.; Fernández-Ruiz, V.; Cámara, M. The Frontier between Nutrition and Pharma: The International Regulatory Framework of Functional Foods, Food Supplements and Nutraceuticals. Crit. Rev. Food Sci. Nutr. 2020, 60, 1738–1746. [Google Scholar] [CrossRef]
- Kalra, E.K. Nutraceutical-Definition and Introduction. AAPS PharmSci 2003, 5, 27–28. [Google Scholar] [CrossRef] [PubMed]
- Hardy, G. Nutraceuticals and Functional Foods: Introduction and Meaning. Nutrition 2000, 16, 688–689. [Google Scholar] [CrossRef]
- Basu, S.; Thomas, J.; Acharya, S. Prospects for Growth in Global Nutraceutical and Functional Food Markets: A Canadian Perspective. Aust. J. Basic Appl. Sci. 2007, 1, 637–649. [Google Scholar]
- Kwak, N.-S.; Jukes, D.J. Functional Foods. Part 1: The Development of a Regulatory Concept. Food Control 2001, 12, 99–107. [Google Scholar] [CrossRef]
- Guleri, S.; Tiwari, A. Algae and Ageing. In Microalgae Biotechnology for Food, Health and High Value Products; Alam, M.A., Xu, J.-L., Wang, Z., Eds.; Springer: Singapore, 2020; pp. 267–293. ISBN 9789811501692. [Google Scholar]
- Hachicha, R.; Elleuch, F.; Ben Hlima, H.; Dubessay, P.; de Baynast, H.; Delattre, C.; Pierre, G.; Hachicha, R.; Abdelkafi, S.; Michaud, P.; et al. Biomolecules from Microalgae and Cyanobacteria: Applications and Market Survey. Appl. Sci. 2022, 12, 1924. [Google Scholar] [CrossRef]
- Remize, M.; Brunel, Y.; Silva, J.L.; Berthon, J.-Y.; Filaire, E. Microalgae N-3 PUFAs Production and Use in Food and Feed Industries. Mar. Drugs 2021, 19, 113. [Google Scholar] [CrossRef]
- Ampofo, J.; Abbey, L. Microalgae: Bioactive Composition, Health Benefits, Safety and Prospects as Potential High-Value Ingredients for the Functional Food Industry. Foods 2022, 11, 1744. [Google Scholar] [CrossRef] [PubMed]
- Zanella, L.; Vianello, F. Microalgae of the Genus Nannochloropsis: Chemical Composition and Functional Implications for Human Nutrition. J. Funct. Foods 2020, 68, 103919. [Google Scholar] [CrossRef]
- Chen, C.; Tang, T.; Shi, Q.; Zhou, Z.; Fan, J. The Potential and Challenge of Microalgae as Promising Future Food Sources. Trends Food Sci. Technol. 2022, 126, 99–112. [Google Scholar] [CrossRef]
- Bernaerts, T.M.M.; Gheysen, L.; Kyomugasho, C.; Jamsazzadeh Kermani, Z.; Vandionant, S.; Foubert, I.; Hendrickx, M.E.; Van Loey, A.M. Comparison of Microalgal Biomasses as Functional Food Ingredients: Focus on the Composition of Cell Wall Related Polysaccharides. Algal Res. 2018, 32, 150–161. [Google Scholar] [CrossRef]
- Raposo, M.F.D.J.; De Morais, A.M.M.B.; De Morais, R.M.S.C. Carotenoids from Marine Microalgae: A Valuable Natural Source for the Prevention of Chronic Diseases. Mar. Drugs 2015, 13, 5128–5155. [Google Scholar] [CrossRef] [PubMed]
- Dehghani, J.; Movafeghi, A.; Mathieu-Rivet, E.; Mati-Baouche, N.; Calbo, S.; Lerouge, P.; Bardor, M. Microalgae as an Efficient Vehicle for the Production and Targeted Delivery of Therapeutic Glycoproteins against SARS-CoV-2 Variants. Mar. Drugs 2022, 20, 657. [Google Scholar] [CrossRef] [PubMed]
- Torres-Tiji, Y.; Fields, F.J.; Mayfield, S.P. Microalgae as a Future Food Source. Biotechnol. Adv. 2020, 41, 107536. [Google Scholar] [CrossRef]
- Prüser, T.F.; Braun, P.G.; Wiacek, C. Microalgae as a novel food. Potential and legal framework. Ernahrungs Umsch. 2021, 68, 78–85. [Google Scholar]
- Cecchin, M.; Cazzaniga, S.; Martini, F.; Paltrinieri, S.; Bossi, S.; Maffei, M.E.; Ballottari, M. Astaxanthin and Eicosapentaenoic Acid Production by S4, a New Mutant Strain of Nannochloropsis gaditana. Microb. Cell Factories 2022, 21, 117. [Google Scholar] [CrossRef]
- Novoveská, L.; Ross, M.E.; Stanley, M.S.; Pradelles, R.; Wasiolek, V.; Sassi, J.-F. Microalgal Carotenoids: A Review of Production, Current Markets, Regulations, and Future Direction. Mar. Drugs 2019, 17, 640. [Google Scholar] [CrossRef]
- Guiry, M.D.; Guiry, G.M. AlgaeBase World-Wide Electronic Publication, National University of Ireland, Galway. Available online: https://www.algaebase.org (accessed on 23 December 2022).
- Sansone, C.; Galasso, C.; Orefice, I.; Nuzzo, G.; Luongo, E.; Cutignano, A.; Romano, G.; Brunet, C.; Fontana, A.; Esposito, F.; et al. The Green Microalga Tetraselmis suecica Reduces Oxidative Stress and Induces Repairing Mechanisms in Human Cells. Sci. Rep. 2017, 7, 41215. [Google Scholar] [CrossRef]
- Cha, Y.; Kim, T.; Jeon, J.; Jang, Y.; Kim, P.B.; Lopes, C.; Leblanc, P.; Cohen, B.M.; Kim, K.-S. SIRT2 Regulates Mitochondrial Dynamics and Reprogramming via MEK1-ERK-DRP1 and AKT1-DRP1 Axes. Cell Rep. 2021, 37, 110155. [Google Scholar] [CrossRef]
- Nacer, W.; Baba Ahmed, F.Z.; Merzouk, H.; Benyagoub, O.; Bouanane, S. Evaluation of the Anti-Inflammatory and Antioxidant Effects of the Microalgae Nannochloropsis gaditana in Streptozotocin-Induced Diabetic Rats. J. Diabetes Metab. Disord. 2020, 19, 1483–1490. [Google Scholar] [CrossRef]
- Gupta, S.P.; Siddiqi, N.J.; Khan, H.A.; Alrokayan, S.H.; Alhomida, A.S.; Singh, R.K.; Verma, P.K.; Kumar, S.; Acharya, A.; Sharma, B. Phytochemical Profiling of Microalgae Euglena tuba and Its Anticancer Activity in Dalton’s Lymphoma Cells. Front. Biosci.-Landmark 2022, 27, 120. [Google Scholar] [CrossRef] [PubMed]
- Lin, P.-Y.; Tsai, C.-T.; Chuang, W.-L.; Chao, Y.-H.; Pan, I.-H.; Chen, Y.-K.; Lin, C.-C.; Wang, B.-Y. Chlorella sorokiniana Induces Mitochondrial-Mediated Apoptosis in Human Non-Small Cell Lung Cancer Cells and Inhibits Xenograft Tumor Growth In Vivo. BMC Complement. Altern. Med. 2017, 17, 88. [Google Scholar] [CrossRef] [PubMed]
- Abolhasani, M.H.; Safavi, M.; Goodarzi, M.T.; Kassaee, S.M.; Azin, M. Identification and Anti-Cancer Activity in 2D and 3D Cell Culture Evaluation of an Iranian Isolated Marine Microalgae Picochlorum sp. RCC486. DARU J. Pharm. Sci. 2018, 26, 105–116. [Google Scholar] [CrossRef] [PubMed]
- Suh, S.-S.; Kim, S.-M.; Kim, J.E.; Hong, J.-M.; Lee, S.G.; Youn, U.J.; Han, S.J.; Kim, I.-C.; Kim, S. Anticancer Activities of Ethanol Extract from the Antarctic Freshwater Microalga, Botryidiopsidaceae sp. BMC Complement. Altern. Med. 2017, 17, 509. [Google Scholar] [CrossRef]
- Ebrahimi Nigjeh, S.; Yusoff, F.M.; Mohamed Alitheen, N.B.; Rasoli, M.; Keong, Y.S.; bin Omar, A.R. Cytotoxic Effect of Ethanol Extract of Microalga, Chaetoceros calcitrans, and Its Mechanisms in Inducing Apoptosis in Human Breast Cancer Cell Line. BioMed Res. Int. 2012, 2013, e783690. [Google Scholar] [CrossRef]
- Guo, B.; Zhou, Y.; Liu, B.; He, Y.; Chen, F.; Cheng, K.-W. Lipid-Lowering Bioactivity of Microalga Nitzschia laevis Extract Containing Fucoxanthin in Murine Model and Carcinomic Hepatocytes. Pharmaceuticals 2021, 14, 1004. [Google Scholar] [CrossRef]
- Campiche, R.; Sandau, P.; Kurth, E.; Massironi, M.; Imfeld, D.; Schuetz, R. Protective Effects of an Extract of the Freshwater Microalga Scenedesmus rubescens on UV-Irradiated Skin Cells. Int. J. Cosmet. Sci. 2018, 40, 187–192. [Google Scholar] [CrossRef]
- Nawrocka, D.; Kornicka, K.; Śmieszek, A.; Marycz, K. Spirulina platensis Improves Mitochondrial Function Impaired by Elevated Oxidative Stress in Adipose-Derived Mesenchymal Stromal Cells (ASCs) and Intestinal Epithelial Cells (IECs), and Enhances Insulin Sensitivity in Equine Metabolic Syndrome (EMS) Horses. Mar. Drugs 2017, 15, 237. [Google Scholar] [CrossRef] [PubMed]
- Stanley, W.C.; Khairallah, R.J.; Dabkowski, E.R. Update on Lipids and Mitochondrial Function: Impact of Dietary n-3 Polyunsaturated Fatty Acids. Curr. Opin. Clin. Nutr. Metab. Care 2012, 15, 122–126. [Google Scholar] [CrossRef]
- Khairallah, R.J.; Sparagna, G.C.; Khanna, N.; O’Shea, K.M.; Hecker, P.A.; Kristian, T.; Fiskum, G.; Des Rosiers, C.; Polster, B.M.; Stanley, W.C. Dietary Supplementation with Docosahexaenoic Acid, but Not Eicosapentaenoic Acid, Dramatically Alters Cardiac Mitochondrial Phospholipid Fatty Acid Composition and Prevents Permeability Transition. Biochim. Biophys. Acta BBA Bioenerg. 2010, 1797, 1555–1562. [Google Scholar] [CrossRef] [PubMed]
- Singer, P.; Honigmann, G.; Schliack, V. Decrease of Eicosapentaenoic Acid in Fatty Liver of Diabetic Subjects. Prostaglandins Med. 1980, 5, 183–200. [Google Scholar] [CrossRef] [PubMed]
- Wensaas, A.J.; Rustan, A.C.; Just, M.; Berge, R.K.; Drevon, C.A.; Gaster, M. Fatty Acid Incubation of Myotubes From Humans With Type 2 Diabetes Leads to Enhanced Release of β-Oxidation Products Because of Impaired Fatty Acid Oxidation: Effects of Tetradecylthioacetic Acid and Eicosapentaenoic Acid. Diabetes 2009, 58, 527–535. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Chen, X. Eicosapentaenoic Acid Promotes Thermogenic and Fatty Acid Storage Capacity in Mouse Subcutaneous Adipocytes. Biochem. Biophys. Res. Commun. 2014, 450, 1446–1451. [Google Scholar] [CrossRef]
- Zhou, L.; Li, K.; Duan, X.; Hill, D.; Barrow, C.; Dunshea, F.; Martin, G.; Suleria, H. Bioactive Compounds in Microalgae and Their Potential Health Benefits. Food Biosci. 2022, 49, 101932. [Google Scholar] [CrossRef]
- Randhir, A.; Laird, D.W.; Maker, G.; Trengove, R.; Moheimani, N.R. Microalgae: A Potential Sustainable Commercial Source of Sterols. Algal Res. 2020, 46, 101772. [Google Scholar] [CrossRef]
- Fithriani, D.; Ambarwaty, D.; Nurhayati, N. Identification of Bioactive Compounds from Nannochloropsis sp. IOP Conf. Series: Earth Environ. Sci. 2020, 404, 012064. [Google Scholar] [CrossRef]
- Kim, Y.-S.; Li, X.-F.; Kang, K.-H.; Ryu, B.; Kim, S.K. Stigmasterol Isolated from Marine Microalgae Navicula incerta Induces Apoptosis in Human Hepatoma HepG2 Cells. BMB Rep. 2014, 47, 433–438. [Google Scholar] [CrossRef]
- Cilla, A.; Attanzio, A.; Barberá, R.; Tesoriere, L.; Livrea, M.A. Anti-Proliferative Effect of Main Dietary Phytosterols and β-Cryptoxanthin Alone or Combined in Human Colon Cancer Caco-2 Cells through Cytosolic Ca+2–and Oxidative Stress-Induced Apoptosis. J. Funct. Foods 2015, 12, 282–293. [Google Scholar] [CrossRef]
- Mamari, H.H.A. Phenolic Compounds: Classification, Chemistry, and Updated Techniques of Analysis and Synthesis; IntechOpen: London, UK, 2021; ISBN 978-1-83969-347-2. [Google Scholar]
- Alara, O.R.; Abdurahman, N.H.; Ukaegbu, C.I. Extraction of Phenolic Compounds: A Review. Curr. Res. Food Sci. 2021, 4, 200–214. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, S.; Singh, M.; Srivastava, A.; Chavali, M.; Chandrasekhar, K.; Verma, P. Extraction and Characterization of Microalgae-Derived Phenolics for Pharmaceutical Applications: A Systematic Review. J. Basic Microbiol. 2021, 62, 1044–1063. [Google Scholar] [CrossRef]
- Goiris, K.; Muylaert, K.; Fraeye, I.; Foubert, I.; De Brabanter, J.; De Cooman, L. Antioxidant Potential of Microalgae in Relation to Their Phenolic and Carotenoid Content. J. Appl. Phycol. 2012, 24, 1477–1486. [Google Scholar] [CrossRef]
- Rodis, P.S.; Karathanos, V.T.; Mantzavinou, A. Partitioning of Olive Oil Antioxidants between Oil and Water Phases. J. Agric. Food Chem. 2002, 50, 596–601. [Google Scholar] [CrossRef] [PubMed]
- Scaglioni, P.T.; Quadros, L.; de Paula, M.; Furlong, V.B.; Abreu, P.C.; Badiale-Furlong, E. Inhibition of Enzymatic and Oxidative Processes by Phenolic Extracts from Spirulina sp. and Nannochloropsis sp. Food Technol. Biotechnol. 2018, 56, 344–353. [Google Scholar] [CrossRef]
- Goh, S.-H.; Yusoff, F.M.; Loh, S.P. A Comparison of the Antioxidant Properties and Total Phenolic Content in a Diatom, Chaetoceros sp. and a Green Microalga, Nannochloropsis sp. J. Agric. Sci. 2010, 2, p123. [Google Scholar] [CrossRef]
- Parcheta, M.; Świsłocka, R.; Orzechowska, S.; Akimowicz, M.; Choińska, R.; Lewandowski, W. Recent Developments in Effective Antioxidants: The Structure and Antioxidant Properties. Materials 2021, 14, 1984. [Google Scholar] [CrossRef] [PubMed]
- Andriopoulos, V.; Gkioni, M.D.; Koutra, E.; Mastropetros, S.G.; Lamari, F.N.; Hatziantoniou, S.; Kornaros, M. Total Phenolic Content, Biomass Composition, and Antioxidant Activity of Selected Marine Microalgal Species with Potential as Aquaculture Feed. Antioxidants 2022, 11, 1320. [Google Scholar] [CrossRef] [PubMed]
- Muñóz-Almagro, N.; Gilbert-López, B.M.; Carmen, P.-R.; García-Fernandez, Y.; Almeida, C.; Villamiel, M.; Mendiola, J.A.; Ibáñez, E. Exploring the Microalga Euglena cantabrica by Pressurized Liquid Extraction to Obtain Bioactive Compounds. Mar. Drugs 2020, 18, 308. [Google Scholar] [CrossRef]
- Safafar, H.; Van Wagenen, J.; Møller, P.; Jacobsen, C. Carotenoids, Phenolic Compounds and Tocopherols Contribute to the Antioxidative Properties of Some Microalgae Species Grown on Industrial Wastewater. Mar. Drugs 2015, 13, 7339–7356. [Google Scholar] [CrossRef] [PubMed]
- Massa, M.; Buono, S.; Langellotti, A.L.; Martello, A.; Russo, G.L.; Troise, D.A.; Sacchi, R.; Vitaglione, P.; Fogliano, V. Biochemical Composition and in Vitro Digestibility of Galdieria sulphuraria Grown on Spent Cherry-Brine Liquid. New Biotechnol. 2019, 53, 9–15. [Google Scholar] [CrossRef]
- Machu, L.; Misurcova, L.; Vavra Ambrozova, J.; Orsavova, J.; Mlcek, J.; Sochor, J.; Jurikova, T. Phenolic Content and Antioxidant Capacity in Algal Food Products. Molecules 2015, 20, 1118–1133. [Google Scholar] [CrossRef]
- Michael, A.; Kyewalyanga, M.S.; Mtolera, M.S.; Lugomela, C.V. Antioxidants Activity of the Cyanobacterium, Arthrospira (Spirulina) fusiformis Cultivated in a Low-Cost Medium. Afr. J. Food Sci. 2018, 12, 188–195. [Google Scholar] [CrossRef]
- Martinez-Goss, M.R.; Arguelles, E.D.L.R.; Sapin, A.B.; Almeda, R.A. Chemical Composition and In Vitro Antioxidant and Antibacterial Properties of the Edible Cyanobacterium, Nostoc commune Vaucher. Philipp. Sci. Lett. 2021, 14, 25–35. [Google Scholar]
- Lima, G.P.P.; Vianello, F.; Corrêa, C.R.; Campos, R.A.D.S.; Borguini, M.G. Polyphenols in Fruits and Vegetables and Its Effect on Human Health. Food Nutr. Sci. 2014, 5, 1065–1082. [Google Scholar] [CrossRef]
- Chodari, L.; Dilsiz Aytemir, M.; Vahedi, P.; Alipour, M.; Vahed, S.Z.; Khatibi, S.M.H.; Ahmadian, E.; Ardalan, M.; Eftekhari, A. Targeting Mitochondrial Biogenesis with Polyphenol Compounds. Oxid. Med. Cell. Longev. 2021, 2021, e4946711. [Google Scholar] [CrossRef]
- Sandoval-Acuña, C.; Ferreira, J.; Speisky, H. Polyphenols and Mitochondria: An Update on Their Increasingly Emerging ROS-Scavenging Independent Actions. Arch. Biochem. Biophys. 2014, 559, 75–90. [Google Scholar] [CrossRef] [PubMed]
- Naoi, M.; Wu, Y.; Shamoto-Nagai, M.; Maruyama, W. Mitochondria in Neuroprotection by Phytochemicals: Bioactive Polyphenols Modulate Mitochondrial Apoptosis System, Function and Structure. Int. J. Mol. Sci. 2019, 20, 2451. [Google Scholar] [CrossRef]
- Howitz, K.T.; Bitterman, K.J.; Cohen, H.Y.; Lamming, D.W.; Lavu, S.; Wood, J.G.; Zipkin, R.E.; Chung, P.; Kisielewski, A.; Zhang, L.-L.; et al. Small Molecule Activators of Sirtuins Extend Saccharomyces cerevisiae Lifespan. Nature 2003, 425, 191–196. [Google Scholar] [CrossRef]
- Davis, J.M.; Murphy, E.A.; Carmichael, M.D.; Davis, B. Quercetin Increases Brain and Muscle Mitochondrial Biogenesis and Exercise Tolerance. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2009, 296, R1071–R1077. [Google Scholar] [CrossRef] [PubMed]
- Hao, J.; Shen, W.; Yu, G.; Jia, H.; Li, X.; Feng, Z.; Wang, Y.; Weber, P.; Wertz, K.; Sharman, E.; et al. Hydroxytyrosol Promotes Mitochondrial Biogenesis and Mitochondrial Function in 3T3-L1 Adipocytes. J. Nutr. Biochem. 2010, 21, 634–644. [Google Scholar] [CrossRef] [PubMed]
- Signorile, A.; Micelli, L.; De Rasmo, D.; Santeramo, A.; Papa, F.; Ficarella, R.; Gattoni, G.; Scacco, S.; Papa, S. Regulation of the Biogenesis of OXPHOS Complexes in Cell Transition from Replicating to Quiescent State: Involvement of PKA and Effect of Hydroxytyrosol. Biochim. Biophys. Acta 2014, 1843, 675–684. [Google Scholar] [CrossRef]
- Feillet-Coudray, C.; Sutra, T.; Fouret, G.; Ramos, J.; Wrutniak-Cabello, C.; Cabello, G.; Cristol, J.P.; Coudray, C. Oxidative Stress in Rats Fed a High-Fat High-Sucrose Diet and Preventive Effect of Polyphenols: Involvement of Mitochondrial and NAD(P)H Oxidase Systems. Free Radic. Biol. Med. 2009, 46, 624–632. [Google Scholar] [CrossRef] [PubMed]
- Zimermann, J.D.F.; Sydney, E.B.; Cerri, M.L.; de Carvalho, I.K.; Schafranski, K.; Sydney, A.C.N.; Vitali, L.; Gonçalves, S.; Micke, G.A.; Soccol, C.R.; et al. Growth Kinetics, Phenolic Compounds Profile and Pigments Analysis of Galdieria sulphuraria Cultivated in Whey Permeate in Shake-Flasks and Stirred-Tank Bioreactor. J. Water Process Eng. 2020, 38, 101598. [Google Scholar] [CrossRef]
- Hwang, J.M.; Cho, J.S.; Kim, T.H.; Lee, Y.I. Ellagic Acid Protects Hepatocytes from Damage by Inhibiting Mitochondrial Production of Reactive Oxygen Species. Biomed. Pharmacother. 2010, 64, 264–270. [Google Scholar] [CrossRef] [PubMed]
- Firdaus, F.; Zafeer, M.F.; Waseem, M.; Anis, E.; Hossain, M.M.; Afzal, M. Ellagic Acid Mitigates Arsenic-Trioxide-Induced Mitochondrial Dysfunction and Cytotoxicity in SH-SY5Y Cells. J. Biochem. Mol. Toxicol. 2018, 32, e22024. [Google Scholar] [CrossRef]
- Dhingra, A.; Jayas, R.; Afshar, P.; Guberman, M.; Maddaford, G.; Gerstein, J.; Lieberman, B.; Nepon, H.; Margulets, V.; Dhingra, R.; et al. Ellagic Acid Antagonizes Bnip3-Mediated Mitochondrial Injury and Necrotic Cell Death of Cardiac Myocytes. Free Radic. Biol. Med. 2017, 112, 411–422. [Google Scholar] [CrossRef]
- Larrosa, M.; Tomás-Barberán, F.A.; Espín, J.C. The Dietary Hydrolysable Tannin Punicalagin Releases Ellagic Acid That Induces Apoptosis in Human Colon Adenocarcinoma Caco-2 Cells by Using the Mitochondrial Pathway. J. Nutr. Biochem. 2006, 17, 611–625. [Google Scholar] [CrossRef]
- Salimi, A.; Roudkenar, M.H.; Sadeghi, L.; Mohseni, A.; Seydi, E.; Pirahmadi, N.; Pourahmad, J. Ellagic Acid, a Polyphenolic Compound, Selectively Induces ROS-Mediated Apoptosis in Cancerous B-Lymphocytes of CLL Patients by Directly Targeting Mitochondria. Redox Biol. 2015, 6, 461–471. [Google Scholar] [CrossRef]
- Ho, C.-C.; Huang, A.-C.; Yu, C.-S.; Lien, J.-C.; Wu, S.-H.; Huang, Y.-P.; Huang, H.-Y.; Kuo, J.-H.; Liao, W.-Y.; Yang, J.-S.; et al. Ellagic Acid Induces Apoptosis in TSGH8301 Human Bladder Cancer Cells through the Endoplasmic Reticulum Stress- and Mitochondria-Dependent Signaling Pathways. Environ. Toxicol. 2014, 29, 1262–1274. [Google Scholar] [CrossRef]
- Edderkaoui, M.; Odinokova, I.; Ohno, I.; Gukovsky, I.; Go, V.L.W.; Pandol, S.J.; Gukovskaya, A.S. Ellagic Acid Induces Apoptosis through Inhibition of Nuclear Factor Kappa B in Pancreatic Cancer Cells. World J. Gastroenterol. 2008, 14, 3672–3680. [Google Scholar] [CrossRef] [PubMed]
- Duan, J.; Li, Y.; Gao, H.; Yang, D.; He, X.; Fang, Y.; Zhou, G. Phenolic Compound Ellagic Acid Inhibits Mitochondrial Respiration and Tumor Growth in Lung Cancer. Food Funct. 2020, 11, 6332–6339. [Google Scholar] [CrossRef] [PubMed]
- Gorlach, S.; Fichna, J.; Lewandowska, U. Polyphenols as Mitochondria-Targeted Anticancer Drugs. Cancer Lett. 2015, 366, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Cheng, G.; Qiu, H.; Zhu, L.; Ren, Z.; Zhao, W.; Zhang, T.; Liu, L. The P53-Inducible Gene 3 Involved in Flavonoid-Induced Cytotoxicity through the Reactive Oxygen Species-Mediated Mitochondrial Apoptotic Pathway in Human Hepatoma Cells. Food Funct. 2015, 6, 1518–1525. [Google Scholar] [CrossRef]
- de Oliveira, M.R.; Nabavi, S.F.; Manayi, A.; Daglia, M.; Hajheydari, Z.; Nabavi, S.M. Resveratrol and the Mitochondria: From Triggering the Intrinsic Apoptotic Pathway to Inducing Mitochondrial Biogenesis, a Mechanistic View. Biochim. Biophys. Acta BBA Gen. Subj. 2016, 1860, 727–745. [Google Scholar] [CrossRef]
- Goiris, K.; Muylaert, K.; Voorspoels, S.; Noten, B.; De Paepe, D.; E Baart, G.J.; De Cooman, L. Detection of Flavonoids in Microalgae from Different Evolutionary Lineages. J. Phycol. 2014, 50, 483–492. [Google Scholar] [CrossRef] [PubMed]
- Mapoung, S.; Arjsri, P.; Thippraphan, P.; Semmarath, W.; Yodkeeree, S.; Chiewchanvit, S.; Piyamongkol, W.; Limtrakul, P. Photochemoprotective Effects of Spirulina platensis Extract against UVB Irradiated Human Skin Fibroblasts. S. Afr. J. Bot. 2020, 130, 198–207. [Google Scholar] [CrossRef]
- Seghiri, R.; Kharbach, M.; Essamri, A. Functional Composition, Nutritional Properties, and Biological Activities of Moroccan Spirulina Microalga. J. Food Qual. 2019, 2019, e3707219. [Google Scholar] [CrossRef]
- Niranjana, R.; Gayathri, R.; Nimish Mol, S.; Sugawara, T.; Hirata, T.; Miyashita, K.; Ganesan, P. Carotenoids Modulate the Hallmarks of Cancer Cells. J. Funct. Foods 2015, 18, 968–985. [Google Scholar] [CrossRef]
- Le Goff, M.; Le Ferrec, E.; Mayer, C.; Mimouni, V.; Lagadic-Gossmann, D.; Schoefs, B.; Ulmann, L. Microalgal Carotenoids and Phytosterols Regulate Biochemical Mechanisms Involved in Human Health and Disease Prevention. Biochimie 2019, 167, 106–118. [Google Scholar] [CrossRef]
- Zanella, L.; Alam, M.A. Extracts and Bioactives from Microalgae (Sensu Stricto): Opportunities and Challenges for a New Generation of Cosmetics. In Microalgae Biotechnology for Food, Health and High Value Products; Alam, M.A., Xu, J.-L., Wang, Z., Eds.; Springer: Singapore, 2020; pp. 295–349. ISBN 9789811501692. [Google Scholar]
- Lesmana, R.; Yusuf, I.F.; Goenawan, H.; Achadiyani, A.; Khairani, A.F.; Fatimah, S.N.; Supratman, U. Low Dose of β-Carotene Regulates Inflammation, Reduces Caspase Signaling, and Correlates with Autophagy Activation in Cardiomyoblast Cell Lines. Med. Sci. Monit. Basic Res. 2020, 26, e928648-1. [Google Scholar] [CrossRef]
- Yamamoto, H.; Itoh, N.; Kawano, S.; Yatsukawa, Y.; Momose, T.; Makio, T.; Matsunaga, M.; Yokota, M.; Esaki, M.; Shodai, T.; et al. Dual Role of the Receptor Tom20 in Specificity and Efficiency of Protein Import into Mitochondria. Proc. Natl. Acad. Sci. USA 2011, 108, 91–96. [Google Scholar] [CrossRef]
- Weinrich, T.; Xu, Y.; Wosu, C.; Harvey, P.J.; Jeffery, G. Mitochondrial Function, Mobility and Lifespan Are Improved in Drosophila melanogaster by Extracts of 9-Cis-β-Carotene from Dunaliella salina. Mar. Drugs 2019, 17, 279. [Google Scholar] [CrossRef] [PubMed]
- Voutilainen, S.; Nurmi, T.; Mursu, J.; Rissanen, T.H. Carotenoids and Cardiovascular Health. Am. J. Clin. Nutr. 2006, 83, 1265–1271. [Google Scholar] [CrossRef]
- Druesne-Pecollo, N.; Latino-Martel, P.; Norat, T.; Barrandon, E.; Bertrais, S.; Galan, P.; Hercberg, S. Beta-Carotene Supplementation and Cancer Risk: A Systematic Review and Metaanalysis of Randomized Controlled Trials. Int. J. Cancer 2010, 127, 172–184. [Google Scholar] [CrossRef] [PubMed]
- Sowmya Shree, G.; Yogendra Prasad, K.; Arpitha, H.S.; Deepika, U.R.; Nawneet Kumar, K.; Mondal, P.; Ganesan, P. β-Carotene at Physiologically Attainable Concentration Induces Apoptosis and down-Regulates Cell Survival and Antioxidant Markers in Human Breast Cancer (MCF-7) Cells. Mol. Cell. Biochem. 2017, 436, 1–12. [Google Scholar] [CrossRef]
- He, J.; Gu, Y.; Zhang, S. Vitamin A and Breast Cancer Survival: A Systematic Review and Meta-Analysis. Clin. Breast Cancer 2018, 18, e1389–e1400. [Google Scholar] [CrossRef]
- Lee, K.E.; Kwon, M.; Kim, Y.S.; Kim, Y.; Chung, M.G.; Heo, S.C.; Kim, Y. KoreaMed Synapse. Nutr. Res. Pract. 2021, 16, 161–172. [Google Scholar] [CrossRef]
- Li, K.; Zhang, B. The Association of Dietary β-Carotene and Vitamin A Intake on the Risk of Esophageal Cancer: A Meta-Analysis. Rev. Espanola Enfermedades Dig. Organo Of. Soc. Espanola Patol. Dig. 2020, 112, 620–626. [Google Scholar] [CrossRef] [PubMed]
- Tamaki, S.; Mochida, K.; Suzuki, K. Diverse Biosynthetic Pathways and Protective Functions against Environmental Stress of Antioxidants in Microalgae. Plants 2021, 10, 1250. [Google Scholar] [CrossRef]
- Corona, J.C.; Duchen, M.R. PPARγ as a Therapeutic Target to Rescue Mitochondrial Function in Neurological Disease. Free Radic. Biol. Med. 2016, 100, 153–163. [Google Scholar] [CrossRef]
- Sathasivam, R.; Ki, J.-S. A Review of the Biological Activities of Microalgal Carotenoids and Their Potential Use in Healthcare and Cosmetic Industries. Mar. Drugs 2018, 16, 26. [Google Scholar] [CrossRef]
- Patel, A.K.; Albarico, F.P.J.B.; Perumal, P.K.; Vadrale, A.P.; Nian, C.T.; Chau, H.T.B.; Anwar, C.; Wani, H.M.U.D.; Pal, A.; Saini, R.; et al. Algae as an Emerging Source of Bioactive Pigments. Bioresour. Technol. 2022, 351, 126910. [Google Scholar] [CrossRef]
- Li, Z.; Sun, M.; Li, Q.; Li, A.; Zhang, C. Profiling of Carotenoids in Six Microalgae (Eustigmatophyceae) and Assessment of Their β-Carotene Productions in Bubble Column Photobioreactor. Biotechnol. Lett. 2012, 34, 2049–2053. [Google Scholar] [CrossRef] [PubMed]
- Del Campo, J.A.; Rodríguez, H.; Moreno, J.; Vargas, M.Á.; Rivas, J.; Guerrero, M.G. Accumulation of Astaxanthin and Lutein in Chlorella zofingiensis (Chlorophyta). Appl. Microbiol. Biotechnol. 2004, 64, 848–854. [Google Scholar] [CrossRef]
- Abe, K.; Hattori, H.; Hirano, M. Accumulation and Antioxidant Activity of Secondary Carotenoids in the Aerial Microalga Coelastrella striolata var. multistriata. Food Chem. 2007, 100, 656–661. [Google Scholar] [CrossRef]
- Kang, C.D.; Lee, J.S.; Park, T.H.; Sim, S.J. Comparison of Heterotrophic and Photoautotrophic Induction on Astaxanthin Production by Haematococcus pluvialis. Appl. Microbiol. Biotechnol. 2005, 68, 237–241. [Google Scholar] [CrossRef] [PubMed]
- Butler, T.; Golan, Y. Astaxanthin Production from Microalgae. In Microalgae Biotechnology for Food, Health and High Value Products; Springer: Singapore, 2020; pp. 175–242. [Google Scholar] [CrossRef]
- Liu, J.; Sun, Z.; Gerken, H.; Liu, Z.; Jiang, Y.; Chen, F. Chlorella zofingiensis as an Alternative Microalgal Producer of Astaxanthin: Biology and Industrial Potential. Mar. Drugs 2014, 12, 3487–3515. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.M.; Kang, S.-W.; Kwon, O.-N.; Chung, D.; Pan, C.-H. Fucoxanthin as a Major Carotenoid in Isochrysis aff. galbana: Characterization of Extraction for Commercial Application. J. Korean Soc. Appl. Biol. Chem. 2012, 55, 477–483. [Google Scholar] [CrossRef]
- Petrushkina, M.; Gusev, E.; Sorokin, B.; Zotko, N.; Mamaeva, A.; Filimonova, A.; Kulikovskiy, M.; Maltsev, Y.; Yampolsky, I.; Guglya, E.; et al. Fucoxanthin Production by Heterokont Microalgae. Algal Res. 2017, 24, 387–393. [Google Scholar] [CrossRef]
- Di Lena, G.; Casini, I.; Lucarini, M.; Lombardi-Boccia, G. Carotenoid Profiling of Five Microalgae Species from Large-Scale Production. Food Res. Int. 2019, 120, 810–818. [Google Scholar] [CrossRef]
- Li, Z.; Li, A.-F.; Zhang, C.-W. High Performance Liquid Chromatography Analysis and Supercritical Carbon Dioxide Extraction of Pigments from the Diatom Odontella aurita. Nat. Prod. Res. Dev. 2012, 24, 814–818. [Google Scholar]
- Campenni’, L.; Nobre, B.P.; Santos, C.A.; Oliveira, A.C.; Aires-Barros, M.R.; Palavra, A.M.F.; Gouveia, L. Carotenoid and Lipid Production by the Autotrophic Microalga Chlorella protothecoides under Nutritional, Salinity, and Luminosity Stress Conditions. Appl. Microbiol. Biotechnol. 2013, 97, 1383–1393. [Google Scholar] [CrossRef] [PubMed]
- Diprat, A.B.; Silveira Thys, R.C.; Rodrigues, E.; Rech, R. Chlorella sorokiniana: A New Alternative Source of Carotenoids and Proteins for Gluten-Free Bread. LWT 2020, 134, 109974. [Google Scholar] [CrossRef]
- Huang, W.; Lin, Y.; He, M.; Gong, Y.; Huang, J. Induced High-Yield Production of Zeaxanthin, Lutein, and β-Carotene by a Mutant of Chlorella zofingiensis. J. Agric. Food Chem. 2018, 66, 891–897. [Google Scholar] [CrossRef] [PubMed]
- Bourdon, L.; Jensen, A.A.; Kavanagh, J.M.; McClure, D.D. Microalgal Production of Zeaxanthin. Algal Res. 2021, 55, 102266. [Google Scholar] [CrossRef]
- Singh, D.; Puri, M.; Wilkens, S.; Mathur, A.S.; Tuli, D.K.; Barrow, C.J. Characterization of a New Zeaxanthin Producing Strain of Chlorella saccharophila Isolated from New Zealand Marine Waters. Bioresour. Technol. 2013, 143, 308–314. [Google Scholar] [CrossRef]
- Jin, E.; Feth, B.; Melis, A. A Mutant of the Green Alga Dunaliella salina Constitutively Accumulates Zeaxanthin under All Growth Conditions. Biotechnol. Bioeng. 2003, 81, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Nishida, Y.; Nawaz, A.; Hecht, K.; Tobe, K. Astaxanthin as a Novel Mitochondrial Regulator: A New Aspect of Carotenoids, beyond Antioxidants. Nutrients 2022, 14, 107. [Google Scholar] [CrossRef]
- Hoffman, R.; Sultan, L.D.; Saada, A.; Hirschberg, J.; Osterzetser-Biran, O.; Gruenbaum, Y. Astaxanthin Extends Lifespan via Altered Biogenesis of the Mitochondrial Respiratory Chain Complex III. bioRxiv 2019. [CrossRef]
- Huangfu, J.; Liu, J.; Sun, Z.; Wang, M.; Jiang, Y.; Chen, Z.-Y.; Chen, F. Antiaging Effects of Astaxanthin-Rich Alga Haematococcus pluvialis on Fruit Flies under Oxidative Stress. J. Agric. Food Chem. 2013, 61, 7800–7804. [Google Scholar] [CrossRef]
- Sztretye, M.; Dienes, B.; Gönczi, M.; Czirják, T.; Csernoch, L.; Dux, L.; Szentesi, P.; Keller-Pintér, A. Astaxanthin: A Potential Mitochondrial-Targeted Antioxidant Treatment in Diseases and with Aging. Oxid. Med. Cell. Longev. 2019, 2019, e3849692. [Google Scholar] [CrossRef] [PubMed]
- Wolf, A.M.; Asoh, S.; Hiranuma, H.; Ohsawa, I.; Iio, K.; Satou, A.; Ishikura, M.; Ohta, S. Astaxanthin Protects Mitochondrial Redox State and Functional Integrity against Oxidative Stress. J. Nutr. Biochem. 2010, 21, 381–389. [Google Scholar] [CrossRef] [PubMed]
- Peserico, A.; Chiacchiera, F.; Grossi, V.; Matrone, A.; Latorre, D.; Simonatto, M.; Fusella, A.; Ryall, J.G.; Finley, L.W.S.; Haigis, M.C.; et al. A Novel AMPK-Dependent FoxO3A-SIRT3 Intramitochondrial Complex Sensing Glucose Levels. Cell. Mol. Life Sci. CMLS 2013, 70, 2015–2029. [Google Scholar] [CrossRef] [PubMed]
- Sorrenti, V.; Davinelli, S.; Scapagnini, G.; Willcox, B.J.; Allsopp, R.C.; Willcox, D.C. Astaxanthin as a Putative Geroprotector: Molecular Basis and Focus on Brain Aging. Mar. Drugs 2020, 18, 351. [Google Scholar] [CrossRef]
- Kim, S.H.; Kim, H. Inhibitory Effect of Astaxanthin on Oxidative Stress-Induced Mitochondrial Dysfunction-A Mini-Review. Nutrients 2018, 10, 1137. [Google Scholar] [CrossRef] [Green Version]
- Aoi, W.; Naito, Y.; Takanami, Y.; Ishii, T.; Kawai, Y.; Akagiri, S.; Kato, Y.; Osawa, T.; Yoshikawa, T. Astaxanthin Improves Muscle Lipid Metabolism in Exercise via Inhibitory Effect of Oxidative CPT I Modification. Biochem. Biophys. Res. Commun. 2008, 366, 892–897. [Google Scholar] [CrossRef] [PubMed]
- Gräb, J.; Rybniker, J. The Expanding Role of P38 Mitogen-Activated Protein Kinase in Programmed Host Cell Death. Microbiol. Insights 2019, 12, 1178636119864594. [Google Scholar] [CrossRef]
- Ganesan, P.; Matsubara, K.; Sugawara, T.; Hirata, T. Marine Algal Carotenoids Inhibit Angiogenesis by Down-Regulating FGF-2-Mediated Intracellular Signals in Vascular Endothelial Cells. Mol. Cell. Biochem. 2013, 380, 1–9. [Google Scholar] [CrossRef]
- Palozza, P.; Torelli, C.; Boninsegna, A.; Simone, R.; Catalano, A.; Mele, M.C.; Picci, N. Growth-Inhibitory Effects of the Astaxanthin-Rich Alga Haematococcus pluvialis in Human Colon Cancer Cells. Cancer Lett. 2009, 283, 108–117. [Google Scholar] [CrossRef]
- Kavitha, K.; Kowshik, J.; Kishore, T.K.K.; Baba, A.B.; Nagini, S. Astaxanthin Inhibits NF-ΚB and Wnt/β-Catenin Signaling Pathways via Inactivation of Erk/MAPK and PI3K/Akt to Induce Intrinsic Apoptosis in a Hamster Model of Oral Cancer. Biochim. Biophys. Acta 2013, 1830, 4433–4444. [Google Scholar] [CrossRef]
- Li, J.; Dai, W.; Xia, Y.; Chen, K.; Li, S.; Liu, T.; Zhang, R.; Wang, J.; Lu, W.; Zhou, Y.; et al. Astaxanthin Inhibits Proliferation and Induces Apoptosis of Human Hepatocellular Carcinoma Cells via Inhibition of Nf-Κb P65 and Wnt/Β-Catenin in Vitro. Mar. Drugs 2015, 13, 6064–6081. [Google Scholar] [CrossRef] [PubMed]
- Kaltschmidt, B.; Kaltschmidt, C.; Hofmann, T.G.; Hehner, S.P.; Dröge, W.; Schmitz, M.L. The Pro- or Anti-Apoptotic Function of NF-ΚB Is Determined by the Nature of the Apoptotic Stimulus. Eur. J. Biochem. 2000, 267, 3828–3835. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.-L.; Kamata, H.; Karin, M. IKK/NF-ΚB Signaling: Balancing Life and Death—A New Approach to Cancer Therapy. J. Clin. Investig. 2005, 115, 2625–2632. [Google Scholar] [CrossRef]
- Gardaneh, M.; Nayeri, Z.; Akbari, P.; Gardaneh, M.; Tahermansouri, H. Molecular Simulations Identify Target Receptor Kinases Bound by Astaxanthin to Induce Breast Cancer Cell Apoptosis. Arch. Breast Cancer 2020, 7, 72–82. [Google Scholar] [CrossRef]
- Gammone, M.A.; D’Orazio, N. Anti-Obesity Activity of the Marine Carotenoid Fucoxanthin. Mar. Drugs 2015, 13, 2196–2214. [Google Scholar] [CrossRef] [PubMed]
- Maeda, H. Nutraceutical Effects of Fucoxanthin for Obesity and Diabetes Therapy: A Review. J. Oleo Sci. 2015, 64, 125–132. [Google Scholar] [CrossRef]
- Houstis, N.; Rosen, E.D.; Lander, E.S. Reactive Oxygen Species Have a Causal Role in Multiple Forms of Insulin Resistance. Nature 2006, 440, 944–948. [Google Scholar] [CrossRef] [PubMed]
- Ferdous, K.A.; Burnett, G.; Scott, M.; Amjad, E.; Bannerman, S.; Park, H.-A. Neuroprotective Function of Fucoxanthin in Oxidative Stress-Mediated Mitochondrial Dysfunction. Curr. Dev. Nutr. 2022, 6, 787. [Google Scholar] [CrossRef]
- Mondal, A.; Bose, S.; Banerjee, S.; Patra, J.K.; Malik, J.; Mandal, S.K.; Kilpatrick, K.L.; Das, G.; Kerry, R.G.; Fimognari, C.; et al. Marine Cyanobacteria and Microalgae Metabolites—A Rich Source of Potential Anticancer Drugs. Mar. Drugs 2020, 18, 476. [Google Scholar] [CrossRef]
- Gugulothu, P.; Bajhaiya, A.K. Bioactive Compound from Micro Algae and Their Anti-Cancer Properties. Biomed. J. Sci. Tech. Res. 2022, 42, 33928–33931. [Google Scholar] [CrossRef]
- Saxena, A.; Raj, A.; Tiwari, A.; Saxena, A.; Raj, A.; Tiwari, A. Exploring the Anti-Cancer Potential of Microalgae; IntechOpen: London, UK, 2022; ISBN 978-1-80356-024-3. [Google Scholar]
- Brogi, L.; Marchese, M.; Cellerino, A.; Licitra, R.; Naef, V.; Mero, S.; Bibbiani, C.; Fronte, B. β-Glucans as Dietary Supplement to Improve Locomotion and Mitochondrial Respiration in a Model of Duchenne Muscular Dystrophy. Nutrients 2021, 13, 1619. [Google Scholar] [CrossRef]
- Samarakoon, K.W.; Ko, J.-Y.; Lee, J.-H.; Kwon, O.-N.; Kim, S.-W.; Jeon, Y.-J. Apoptotic Anticancer Activity of a Novel Fatty Alcohol Ester Isolated from Cultured Marine Diatom, Phaeodactylum tricornutum. J. Funct. Foods 2014, 6, 231–240. [Google Scholar] [CrossRef]
- Kasashima, K.; Sumitani, M.; Satoh, M.; Endo, H. Human Prohibitin 1 Maintains the Organization and Stability of the Mitochondrial Nucleoids. Exp. Cell Res. 2008, 314, 988–996. [Google Scholar] [CrossRef]
- Tredici, M.R. Mass Production of Microalgae: Photobioreactors. In Handbook of Microalgal Culture: Biotechnology and Applied Phycology; Blackwell Publishing Ltd.: Oxford, UK, 2004; pp. 178–214. [Google Scholar]
- Vishwakarma, R.; Dhaka, V.; Ariyadasa, T.U.; Malik, A. Exploring Algal Technologies for a Circular Bio-Based Economy in Rural Sector. J. Clean. Prod. 2022, 354, 131653. [Google Scholar] [CrossRef]
- Loke Show, P. Global Market and Economic Analysis of Microalgae Technology: Status and Perspectives. Bioresour. Technol. 2022, 357, 127329. [Google Scholar] [CrossRef]
- Brendler, T.; Williamson, E.M. Astaxanthin: How Much Is Too Much? A Safety Review. Phytother. Res. 2019, 33, 3090–3111. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Xing, L.; Lin, H.; Leng, K.; Zhai, Y.; Liu, X. Assessment and Comparison of in Vitro Immunoregulatory Activity of Three Astaxanthin Stereoisomers. J. Ocean Univ. China 2016, 15, 283–287. [Google Scholar] [CrossRef]
- Becker, W. Microalgae in Human and Animal Nutrition. In Handbook of Microalgal Culture; Blackwell: Oxford, UK, 2004; pp. 312–351. [Google Scholar]
- Muhammad, G.; Alam, M.A.; Xiong, W.; Lv, Y.; Xu, J.-L. Microalgae Biomass Production: An Overview of Dynamic Operational Methods. In Microalgae Biotechnology for Food, Health and High Value Products; Alam, M.A., Xu, J.-L., Wang, Z., Eds.; Springer: Singapore, 2020; pp. 415–432. ISBN 9789811501692. [Google Scholar]
- Teuling, E.; Wierenga, P.A.; Agboola, J.O.; Gruppen, H.; Schrama, J.W. Cell Wall Disruption Increases Bioavailability of Nannochloropsis gaditana Nutrients for Juvenile Nile Tilapia (Oreochromis niloticus). Aquaculture 2019, 499, 269–282. [Google Scholar] [CrossRef]
- Machado, L.; Carvalho, G.; Pereira, R.N. Effects of Innovative Processing Methods on Microalgae Cell Wall: Prospects towards Digestibility of Protein-Rich Biomass. Biomass 2022, 2, 80–102. [Google Scholar] [CrossRef]
- Mendes-Pinto, M.M.; Raposo, M.F.J.; Bowen, J.; Young, A.J.; Morais, R. Evaluation of Different Cell Disruption Processes on Encysted Cells of Haematococcus pluvialis: Effects on Astaxanthin Recovery and Implications for Bio-Availability. J. Appl. Phycol. 2001, 13, 19–24. [Google Scholar] [CrossRef]
- Stiefvatter, L.; Lehnert, K.; Frick, K.; Montoya-Arroyo, A.; Frank, J.; Vetter, W.; Schmid-Staiger, U.; Bischoff, S.C. Oral Bioavailability of Omega-3 Fatty Acids and Carotenoids from the Microalgae Phaeodactylum tricornutum in Healthy Young Adults. Mar. Drugs 2021, 19, 700. [Google Scholar] [CrossRef]
- Merchant, R.E.; Andre, C.A. A Review of Recent Clinical Trials of the Nutritional Supplement Chlorella pyrenoidosa in the Treatment of Fibromyalgia, Hypertension, and Ulcerative Colitis. Altern. Ther. Health Med. 2001, 7, 79–91. [Google Scholar]
- Panahi, Y.; Darvishi, B.; Jowzi, N.; Beiraghdar, F.; Sahebkar, A. Chlorella vulgaris: A Multifunctional Dietary Supplement with Diverse Medicinal Properties. Curr. Pharm. Des. 2016, 22, 164–173. [Google Scholar]
- Ebrahimi-Mameghani, M.; Sadeghi, Z.; Abbasalizad Farhangi, M.; Vaghef-Mehrabany, E.; Aliashrafi, S. Glucose Homeostasis, Insulin Resistance and Inflammatory Biomarkers in Patients with Non-Alcoholic Fatty Liver Disease: Beneficial Effects of Supplementation with Microalgae Chlorella vulgaris: A Double-Blind Placebo-Controlled Randomized Clinical Trial. Clin. Nutr. 2017, 36, 1001–1006. [Google Scholar] [CrossRef]
- Panahi, Y.; Ghamarchehreh, M.E.; Beiraghdar, F.; Zare, R.; Jalalian, H.R.; Sahebkar, A. Investigation of the Effects of Chlorella vulgaris Supplementation in Patients with Non-Alcoholic Fatty Liver Disease: A Randomized Clinical Trial. Hepatogastroenterology 2012, 59, 2099–2103. [Google Scholar] [CrossRef] [PubMed]
- Talebi Pour, B.; Jameshorani, M.; Salmani, R.; Chiti, H. The Effect of Chlorella vulgaris vs. Artichoke on Patients with Non-Alcoholic Fatty Liver Disease (NAFLD): A Randomized Clinical Trial. J. Adv. Med. Biomed. Res. 2015, 23, 36–44. [Google Scholar]
- Chiu, H.-F.; Lee, H.-J.; Han, Y.-C.; Venkatakrishnan, K.; Golovinskaia, O.; Wang, C.-K. Beneficial Effect of Chlorella pyrenoidosa Drink on Healthy Subjects: A Randomized, Placebo-Controlled, Double-Blind, Cross-over Clinical Trial. J. Food Biochem. 2021, 45, e13665. [Google Scholar] [CrossRef] [PubMed]
- Spiller, G.A.; Dewell, A. Safety of an Astaxanthin-Rich Haematococcus pluvialis Algal Extract: A Randomized Clinical Trial. J. Med. Food 2003, 6, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Katagiri, M.; Satoh, A.; Tsuji, S.; Shirasawa, T. Effects of Astaxanthin-Rich Haematococcus pluvialis Extract on Cognitive Function: A Randomised, Double-Blind, Placebo-Controlled Study. J. Clin. Biochem. Nutr. 2012, 51, 102–107. [Google Scholar] [CrossRef]
- Kim, J.H.; Chang, M.J.; Choi, H.D.; Youn, Y.-K.; Kim, J.T.; Oh, J.M.; Shin, W.G. Protective Effects of Haematococcus Astaxanthin on Oxidative Stress in Healthy Smokers. J. Med. Food 2011, 14, 1469–1475. [Google Scholar] [CrossRef]
- Kidd, P. Astaxanthin, Cell Membrane Nutrient with Diverse Clinical Benefits and Anti-Aging Potential. Altern. Med. Rev. J. Clin. Ther. 2011, 16, 355–364. [Google Scholar]
- García, Á.; Toro-Román, V.; Siquier-Coll, J.; Bartolomé, I.; Muñoz, D.; Maynar-Mariño, M. Effects of Tetraselmis chuii Microalgae Supplementation on Anthropometric, Hormonal and Hematological Parameters in Healthy Young Men: A Double-Blind Study. Int. J. Environ. Res. Public. Health 2022, 19, 6060. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, A.; Yasuda, K.; Murata, A.; Suzuki, K.; Miura, N. Effects of Euglena gracilis Intake on Mood and Autonomic Activity under Mental Workload, and Subjective Sleep Quality: A Randomized, Double-Blind, Placebo-Controlled Trial. Nutrients 2020, 12, 3243. [Google Scholar] [CrossRef] [PubMed]
- Rao, A.; Briskey, D.; Nalley, J.O.; Ganuza, E. Omega-3 Eicosapentaenoic Acid (EPA) Rich Extract from the Microalga Nannochloropsis Decreases Cholesterol in Healthy Individuals: A Double-Blind, Randomized, Placebo-Controlled, Three-Month Supplementation Study. Nutrients 2020, 12, 1869. [Google Scholar] [CrossRef]
- Karkos, P.D.; Leong, S.C.; Karkos, C.D.; Sivaji, N.; Assimakopoulos, D.A. Spirulina in Clinical Practice: Evidence-Based Human Applications. Evid. Based Complement. Alternat. Med. 2010, 2011, 531053. [Google Scholar] [CrossRef] [PubMed]
- de la Jara, A.; Ruano-Rodriguez, C.; Polifrone, M.; Assunçao, P.; Brito-Casillas, Y.; Wägner, A.M.; Serra-Majem, L. Impact of Dietary Arthrospira (Spirulina) Biomass Consumption on Human Health: Main Health Targets and Systematic Review. J. Appl. Phycol. 2018, 30, 2403–2423. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zanella, L.; Vianello, F. Potential of Microalgae as Functional Foods Applied to Mitochondria Protection and Healthy Aging Promotion. Nutraceuticals 2023, 3, 119-152. https://doi.org/10.3390/nutraceuticals3010010
Zanella L, Vianello F. Potential of Microalgae as Functional Foods Applied to Mitochondria Protection and Healthy Aging Promotion. Nutraceuticals. 2023; 3(1):119-152. https://doi.org/10.3390/nutraceuticals3010010
Chicago/Turabian StyleZanella, Lorenzo, and Fabio Vianello. 2023. "Potential of Microalgae as Functional Foods Applied to Mitochondria Protection and Healthy Aging Promotion" Nutraceuticals 3, no. 1: 119-152. https://doi.org/10.3390/nutraceuticals3010010