
Reducing the Risk of Benthic Algae Outbreaks by Regulating the Flow Velocity in a Simulated South–North Water Diversion Open Channel
Abstract
:1. Introduction
2. Methods
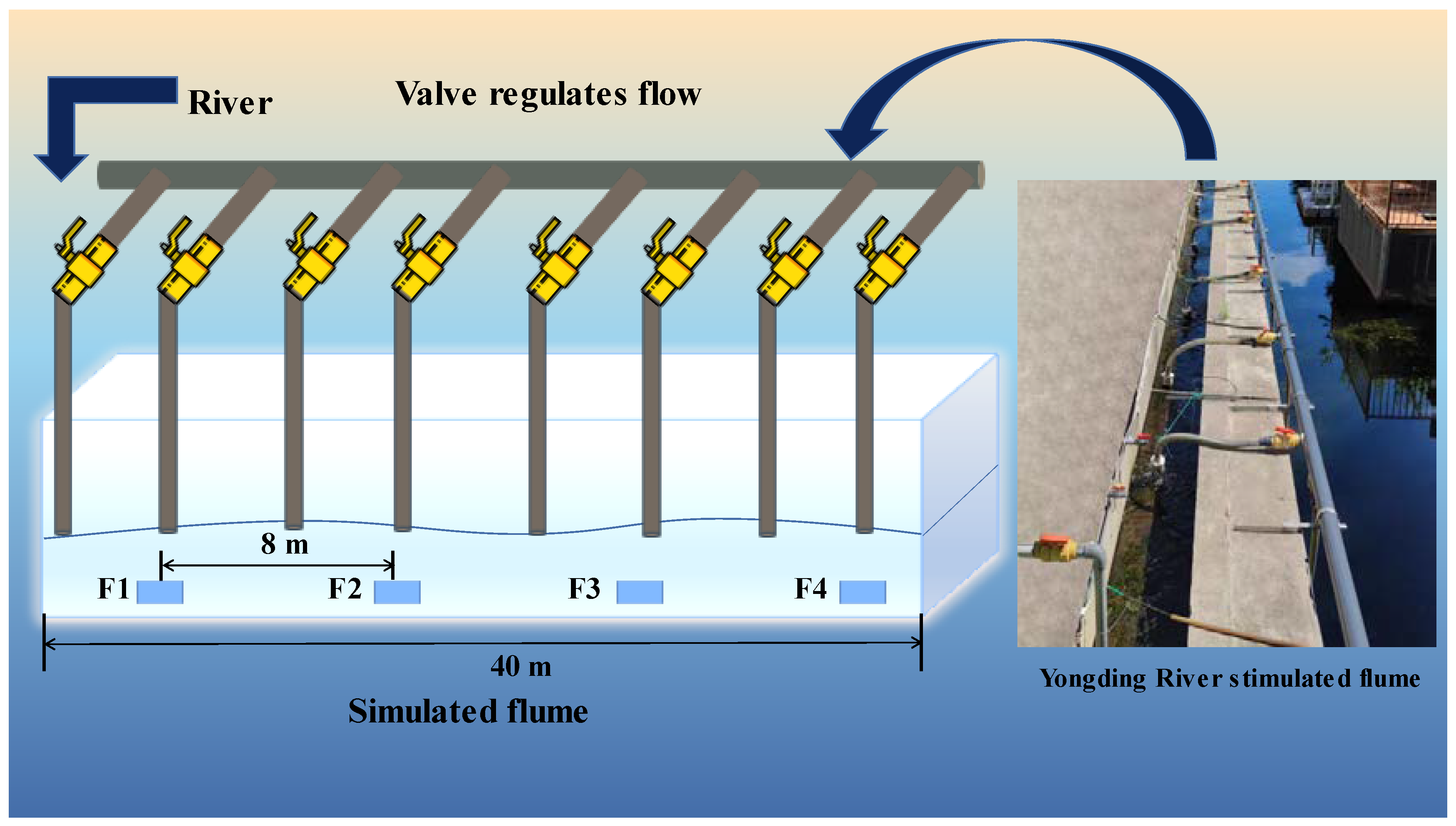
2.1. Experimental Setup and Design
2.2. Sample Extraction and Measurement
2.2.1. Water Sample Collection
2.2.2. Benthic Algae Sample Collection
2.2.3. Physical and Chemical Analysis
2.2.4. Benthic Algae
2.3. Statistical Analysis of Data
3. Results
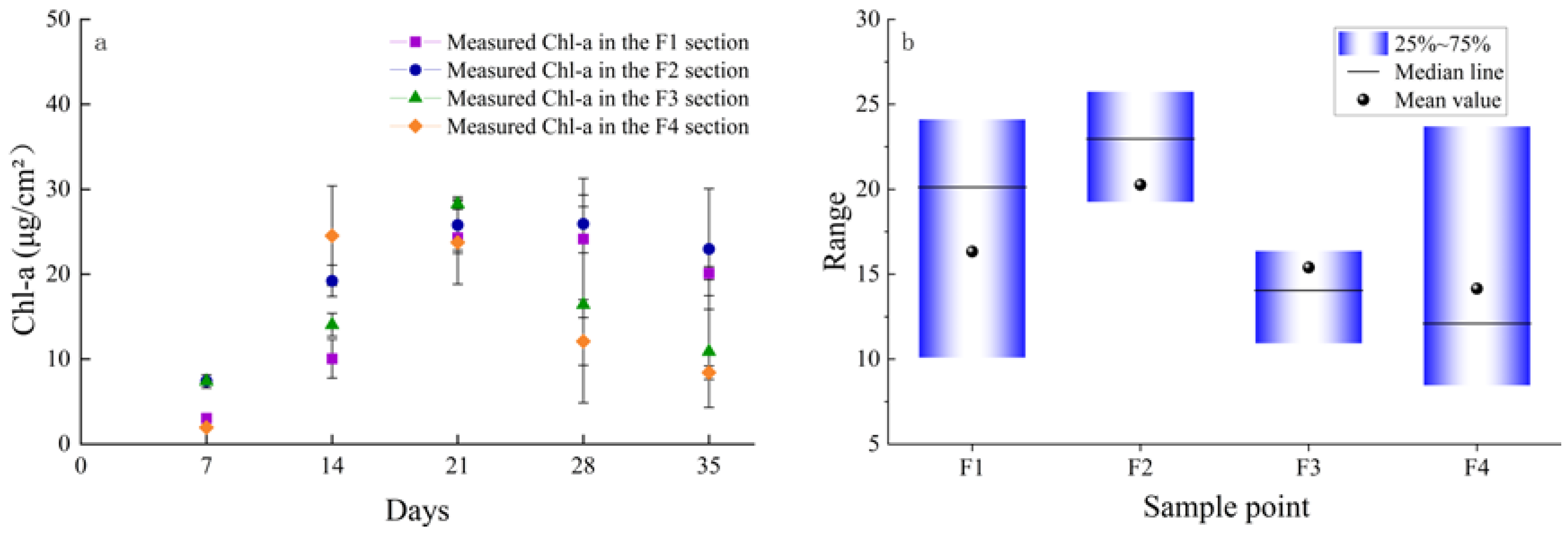
3.1. Evidence for Regime Alterations from Benthic Algal Biomass Levels
3.2. Environmental Metrics Responses to Gradient Flow
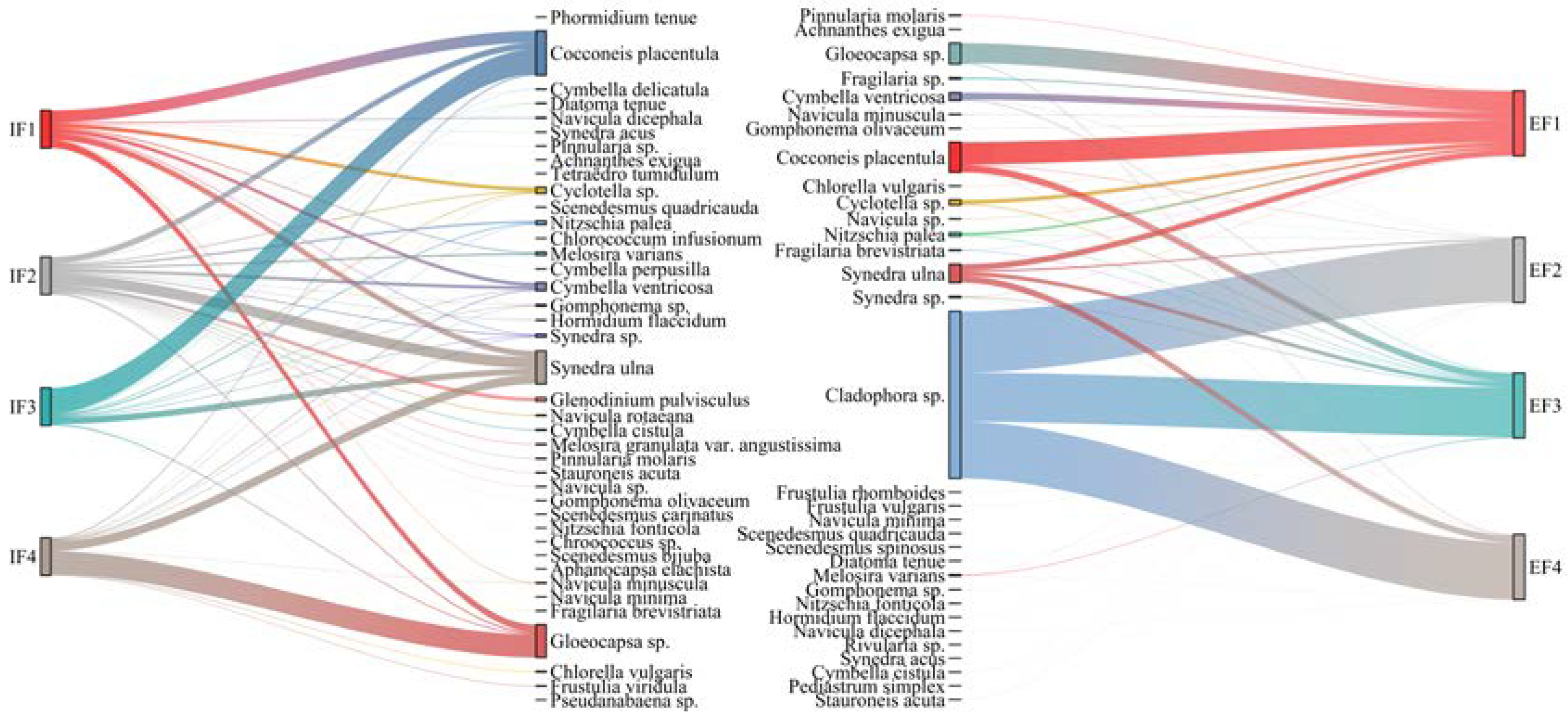
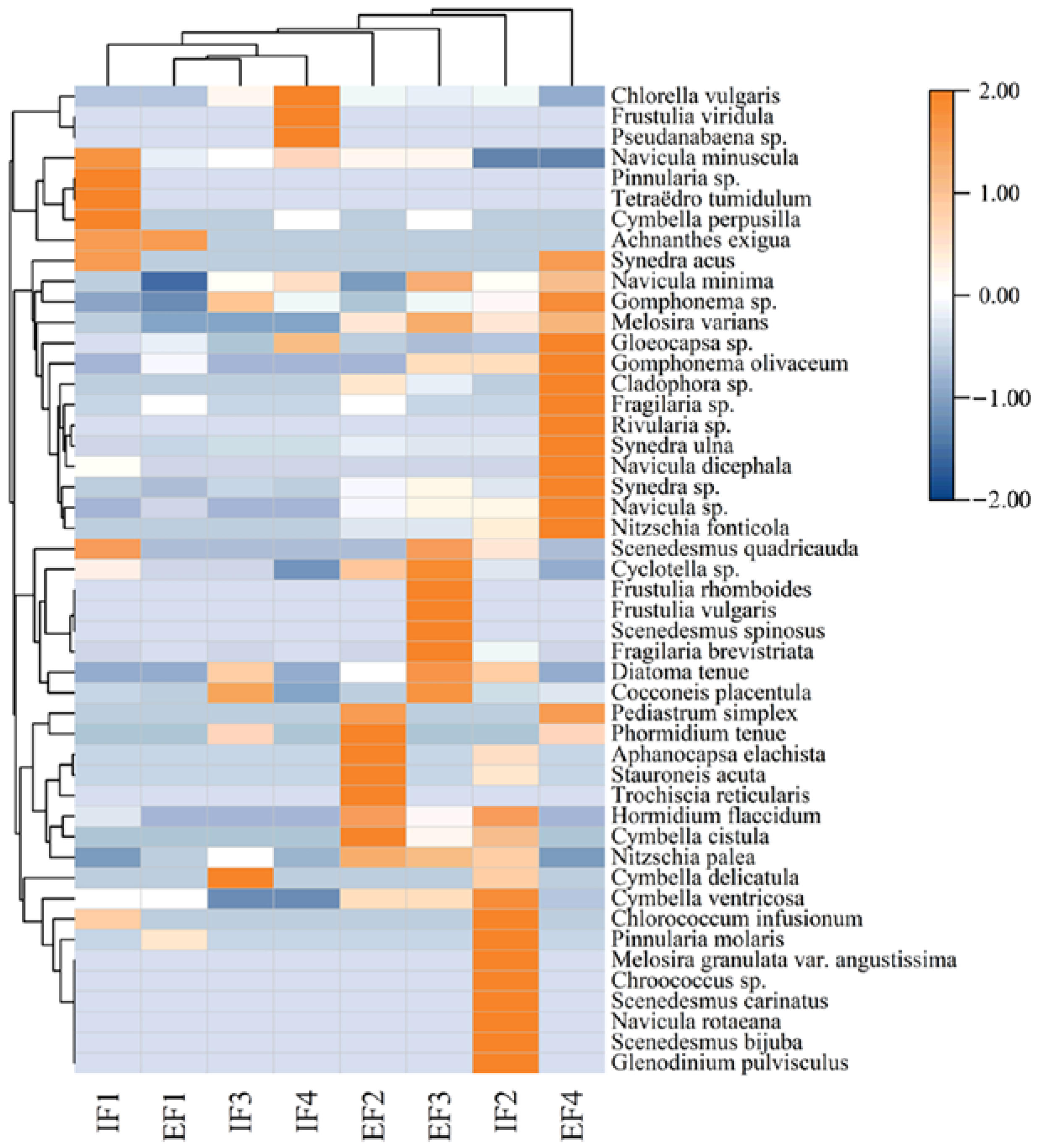
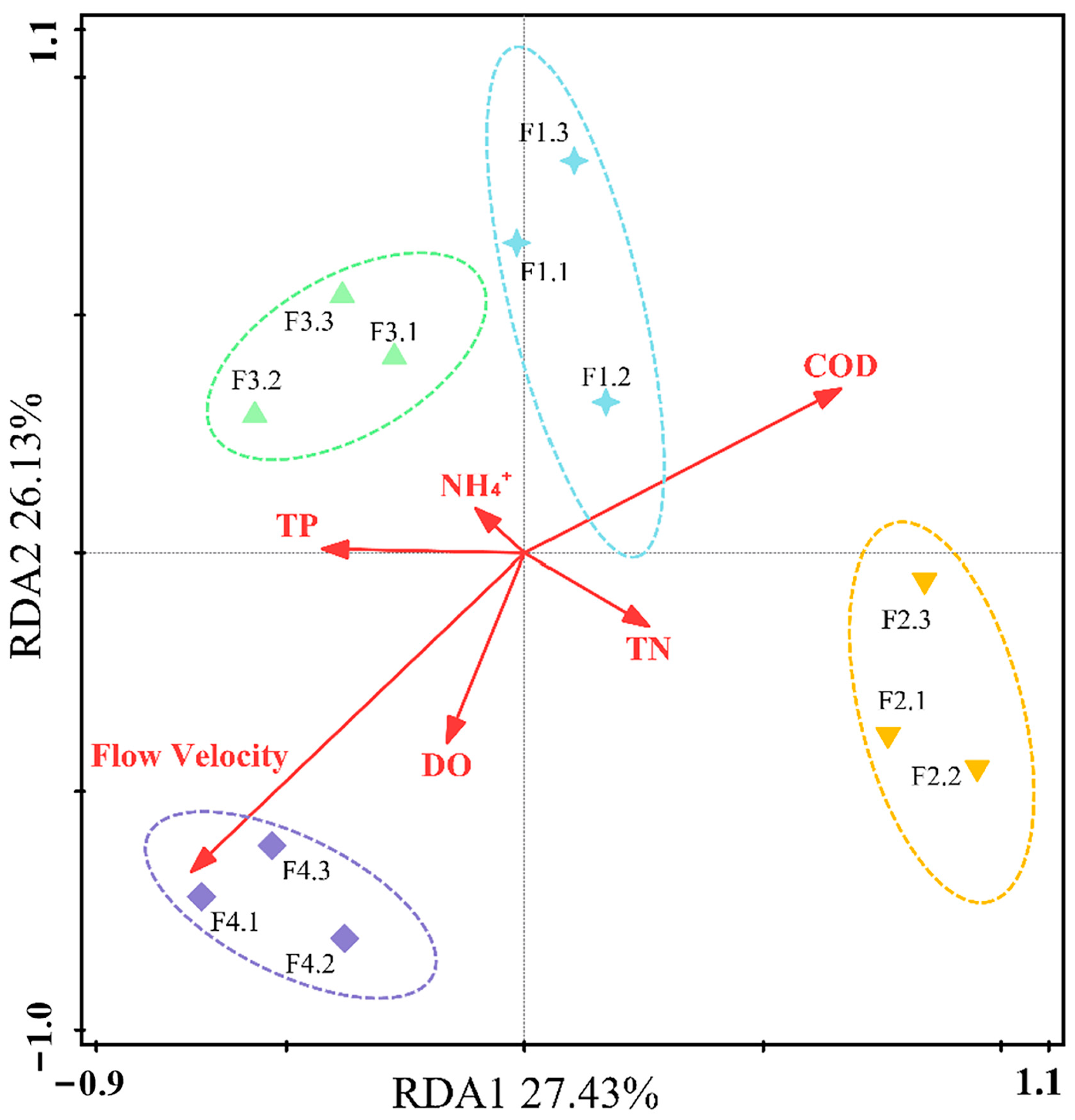
3.3. Relationship between Flow Gradient and Taxonomic Composition of Benthic Algae
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hein, T.; Hauer, C.; Schmid, M.; Stöglehner, G.; Stumpp, C.; Ertl, T.; Graf, W.; Habersack, H.; Haidvogl, G.; Hood-Novotny, R.; et al. The coupled socio-ecohydrological evolution of river systems: Towards an integrative perspective of river systems in the 21st century. Sci. Total Environ. 2021, 801, 149619. [Google Scholar] [CrossRef] [PubMed]
- Duan, S.; Han, F.; Li, F.; Yang, Z. Spatial evaluation of the ecological value importance of national park in Yarlung Tsangpo Grand Canyon. J. Environ. Manag. 2022, 320, 115943. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Ye, A.; Zhang, Y. Changes of total and artificial water bodies in inland China over the past three decades. J. Hydrol. 2022, 613, 128344. [Google Scholar] [CrossRef]
- Fan, X.; Gao, S.; Zhang, Y.; Qin, B.; Xu, H.; Ding, S. Stimulation of high-concentration dissolved nitrogen and reactive phosphorus in Lake Taihu sediments on the initiation and maintenance of cyanobacterial blooms. Sci. Total Environ. 2022, 851, 158088. [Google Scholar] [CrossRef] [PubMed]
- Gu, P.; Zhang, G.; Luo, X.; Xu, L.; Zhang, W.; Li, Q.; Sun, Y.; Zheng, Z. Effects of different fluid fields on the formation of cyanobacterial blooms. Chemosphere 2021, 283, 131219. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Zhang, X.; Huan, W.; Qi, C. Characteristics and Practices of Ecological Flow in Rivers with Flow Reductions Due to Water Storage and Hydropower Projects in China. Water 2018, 10, 1091. [Google Scholar] [CrossRef] [Green Version]
- He, J.; Shi, X. Detection of social-ecological drivers and impact thresholds of ecological degradation and ecological restoration in the last three decades. J. Environ. Manag. 2022, 318, 115513. [Google Scholar] [CrossRef]
- Minshall, G.W.J.B. Autotrophy in Stream Ecosystems. BioScience 1978, 28, 767–771. [Google Scholar] [CrossRef]
- Lock, M.A.; Wallace, R.R.; Costerton, J.W.; Ventullo, R.M.; Charlton, S.E.D.; Calgary, W.J.O. River epilithon: Toward a structural-functional model. Oikos 1984, 42, 10. [Google Scholar] [CrossRef]
- Winterbourn, M.J. Interactions among nutrients, algae and invertebrates in a New Zealand mountain stream. Freshw. Biol. 1990, 23, 463–474. [Google Scholar] [CrossRef]
- Rohlfs, A.-M.; Williams, S.; Rees, G.N.; Lim, R.P.; Werry, L.; Mitrovic, S.M. Experimental dam releases stimulate respiration in an epilithic biofilm community. Hydrobiologia 2018, 820, 175–187. [Google Scholar] [CrossRef]
- Nong, X.; Shao, D.; Zhong, H.; Liang, J. Evaluation of water quality in the South-to-North Water Diversion Project of China using the water quality index (WQI) method. Water Res. 2020, 178, 115781. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Hu, M.; Zhu, Y.; Wang, G.; Xue, B.; Shrestha, S. Phytoplankton community variation and ecological health assessment for impounded lakes along the eastern route of China’s South-to-North Water Diversion Project. J. Environ. Manag. 2022, 318, 115561. [Google Scholar] [CrossRef] [PubMed]
- Nong, X.; Shao, D.; Shang, Y.; Liang, J. Analysis of spatio-temporal variation in phytoplankton and its relationship with water quality parameters in the South-to-North Water Diversion Project of China. Environ. Monit. Assess. 2021, 193, 593. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.-X.; Mi, W.-J.; Li, B.; Liang, J.-K.; Song, G.-F.; Bi, Y.-H. The influences of two hydraulic structures on periphytic algal communities in the middle channel of the South-To-North Water Diversion Project. Acta Hydrobiol. Sin. 2021, 45, 817–825. [Google Scholar]
- Matson, P.G.; Boyer, G.L.; Bridgeman, T.B.; Bullerjahn, G.S.; Kane, D.D.; McKay, R.M.L.; McKindles, K.M.; Raymond, H.A.; Snyder, B.K.; Stumpf, R.P.; et al. Physical drivers facilitating a toxigenic cyanobacterial bloom in a major Great Lakes tributary. Limnol. Oceanogr. 2020, 65, 2866–2882. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Zhang, L.-L.; Li, J.; Chen, M.; Zhang, Y.-W. Mechanism of the influence of hydrodynamics on Microcystis aeruginosa, a dominant bloom species in reservoirs. Sci. Total Environ. 2018, 636, 230–239. [Google Scholar] [CrossRef]
- Tan, Y.; Li, J.; Zhang, L.; Chen, M.; Zhang, Y.; An, R. Mechanism Underlying Flow Velocity and Its Corresponding Influence on the Growth of Euglena gracilis, a Dominant Bloom Species in Reservoirs. Int. J. Environ. Res. Public Health 2019, 16, 4641. [Google Scholar] [CrossRef] [Green Version]
- Irfan, S.; Alatawi, A.M.M. Aquatic Ecosystem and Biodiversity: A Review. Open J. Ecol. 2019, 9, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Marshall, S.J. Hydrology. In Reference Module in Earth Systems and Environmental Sciences; Elsevier: Amsterdam, The Netherlands, 2013. [Google Scholar]
- Stevenson, R.J.; Rollins, S.L. Ecological Assessment with Benthic Algae. In Methods in Stream Ecology, 3rd ed.; Lamberti, G.A., Hauer, F.R., Eds.; Academic Press: Boston, MA, USA, 2017; Chapter 37; pp. 277–292. [Google Scholar]
- EN 14407:2014; Water Quality—Guidance for the Identification and Enumeration of Benthic Diatom Samples from Rivers and Lakes. CEN: Brussels, Belgium, 2014.
- Tornés, E.; Sabater, S. Variable discharge alters habitat suitability for benthic algae and cyanobacteria in a forested Mediterranean stream. Mar. Freshw. Res. 2010, 61, 441–450. [Google Scholar] [CrossRef]
- Battin, T.J.; Kaplan, L.A.; Newbold, J.D.; Cheng, X.; Hansen, C. Effects of current velocity on the nascent architecture of stream microbial biofilms. Appl. Environ. Microbiol. 2003, 69, 5443–5452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, Y.; Hu, H. Freshwater Algae in China: Systematic Classification and Ecology. In Freshwater Algae in China: Systematic Classification and Ecology; Science Press: Beijing, China, 2006. [Google Scholar]
- Spaulding, S.A.; Potapova, M.G.; Bishop, I.W.; Lee, S.S.; Gasperak, T.S.; Jovanoska, E.; Furey, P.C.; Edlund, M.B. Diatoms.org: Supporting taxonomists, connecting communities. Diatom Res. 2021, 36, 291–304. [Google Scholar] [CrossRef] [PubMed]
- Giorgio, A.; De Bonis, S.; Guida, M. Macroinvertebrate and diatom communities as indicators for the biological assessment of river Picentino (Campania, Italy). Ecol. Indic. 2016, 64, 85–91. [Google Scholar] [CrossRef]
- Bennion, H.; Kelly, M.G.; Juggins, S.; Yallop, M.L.; Burgess, A.; Jamieson, J.; Krokowski, J. Assessment of ecological status in UK lakes using benthic diatoms. Freshw. Sci. 2014, 33, 639–654. [Google Scholar] [CrossRef] [Green Version]
- Fore, L.S.; Grafe, C. Using diatoms to assess the biological condition of large rivers in Idaho (USA). Freshw. Biol. 2002, 47, 2015–2037. [Google Scholar] [CrossRef]
- Wilhelm, L.; Besemer, K.; Fragner, L.; Peter, H.; Weckwerth, W.; Battin, T.J. Altitudinal patterns of diversity and functional traits of metabolically active microorganisms in stream biofilms. ISME J. 2015, 9, 2454–2464. [Google Scholar] [CrossRef] [Green Version]
- Schneider, S.C.; Kahlert, M.; Kelly, M.G. Interactions between pH and nutrients on benthic algae in streams and consequences for ecological status assessment and species richness patterns. Sci. Total Environ. 2013, 444, 73–84. [Google Scholar] [CrossRef] [Green Version]
- Stancheva, R.; Sheath, R.G. Benthic soft-bodied algae as bioindicators of stream water quality. Knowl. Manag. Aquat. Ecosyst. 2016, 417, 15. [Google Scholar] [CrossRef] [Green Version]
- Schneider, S.C.; Petrin, Z. Effects of flow regime on benthic algae and macroinvertebrates—A comparison between regulated and unregulated rivers. Sci. Total Environ. 2017, 579, 1059–1072. [Google Scholar] [CrossRef]
- Stubbington, R.; England, J.; Wood, P.J.; Sefton, C.E.M. Temporary streams in temperate zones: Recognizing, monitoring and restoring transitional aquatic-terrestrial ecosystems. Wiley Interdiscip. Rev. Water 2017, 4, e1223. [Google Scholar] [CrossRef] [Green Version]
- Humbert, J.-F.; Shibabaw, T.; Beyene, A.; Awoke, A.; Tirfie, M.; Azage, M.; Triest, L. Diatom community structure in relation to environmental factors in human influenced rivers and streams in tropical Africa. PLoS ONE 2021, 16, e0246043. [Google Scholar] [CrossRef]
- Vadeboncoeur, Y.; Katona, L. Benthic Algae in Lake Littoral Habitats. In Encyclopedia of Inland Waters, 2nd ed.; Mehner, T., Tockner, K., Eds.; Elsevier: Oxford, UK, 2022; pp. 26–37. [Google Scholar]
- Lowe, R.L.; LaLiberte, G.D. Benthic Stream Algae: Distribution and Structure. In Methods in Stream Ecology, 3rd ed.; Hauer, F.R., Lamberti, G.A., Eds.; Academic Press: Boston, MA, USA, 2017; Volume 1, Chapter 11; pp. 193–221. [Google Scholar]
- Biggs, B.J.F. Patterns in Benthic Algae of Streams. In Algal Ecology; Stevenson, R.J., Bothwell, M.L., Lowe, R.L., Eds.; Academic Press: San Diego, CA, USA, 1996; Chapter 2; pp. 31–56. [Google Scholar]
- Graba, M.; Sauvage, S.; Moulin, F.Y.; Urrea, G.; Sabater, S.; Sanchez-Perez, J.M. Interaction between local hydrodynamics and algal community in epilithic biofilm. Water Res. 2013, 47, 2153–2163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horner, R.R.; Welch, E.B. Stream Periphyton Development in Relation to Current Velocity and Nutrients. Can. J. Fish. Aquat. Sci. 1981, 38, 449–457. [Google Scholar] [CrossRef]
- Biggs, B.J.F. Hydraulic habitat of plants in streams. Regul. Rivers Res. Manag. 1996, 12, 131–144. [Google Scholar] [CrossRef]
- Mori, T.; Miyagawa, Y.; Onoda, Y.; Kayaba, Y. Flow-velocity-dependent effects of turbid water on periphyton structure and function in flowing water. Aquat. Sci. 2017, 80, 6. [Google Scholar] [CrossRef]
- Haddadchi, A.; Kuczynski, A.; Hoyle, J.T.; Kilroy, C.; Booker, D.J.; Hicks, M. Periphyton removal flows determined by sediment entrainment thresholds. Ecol. Model. 2020, 434, 109263. [Google Scholar] [CrossRef]
- Davie, A.W.; Mitrovic, S.M. Benthic algal biomass and assemblage changes following environmental flow releases and unregulated tributary flows downstream of a major storage. Mar. Freshw. Res. 2014, 65, 1059–1071. [Google Scholar] [CrossRef]
- King, S.A. Hydrodynamic control of filamentous macroalgae in a sub-tropical spring-fed river in Florida, USA. Hydrobiologia 2014, 734, 27–37. [Google Scholar] [CrossRef]
- Tornes, E.; Colls, M.; Acuna, V.; Sabater, S. Duration of water flow interruption drives the structure and functional diversity of stream benthic diatoms. Sci. Total Environ. 2021, 770, 144675. [Google Scholar] [CrossRef]
- Johnson, R.K.; Angeler, D.G. Effects of agricultural land use on stream assemblages: Taxon-specific responses of alpha and beta diversity. Ecol. Indic. 2014, 45, 386–393. [Google Scholar] [CrossRef]
- Grimm, N.B.; Fisher, S.G. Stability of Periphyton and Macroinvertebrates to Disturbance by Flash Floods in a Desert Stream. J. N. Am. Benthol. Soc. 1989, 8, 293–307. [Google Scholar] [CrossRef]
- Tsai, J.-W.; Chuang, Y.-L.; Wu, Z.-Y.; Kuo, M.-H.; Lin, H.-J. The effects of storm-induced events on the seasonal dynamics of epilithic algal biomass in subtropical mountain streams. Mar. Freshw. Res. 2013, 65, 25–38. [Google Scholar] [CrossRef] [Green Version]
- Algarte, V.M.; Siqueira, N.S.; Ruwer, D.T.; Osório, N.C.; Rodrigues, L. Richness of periphytic algae and its relationship with hydrological attributes. Braz. J. Bot. 2017, 40, 735–740. [Google Scholar] [CrossRef]
- Stevenson, R.J. The Stimulation and Drag of Current. In Algal Ecology; Stevenson, R.J., Bothwell, M.L., Lowe, R.L., Eds.; Academic Press: San Diego, CA, USA, 1996; Chapter 11; pp. 321–340. [Google Scholar]
- Wang, Y.; Wu, N.; Tang, T.; Wang, Y.; Cai, Q. Small run-of-river hydropower dams and associated water regulation filter benthic diatom traits and affect functional diversity. Sci. Total Environ. 2022, 813, 152566. [Google Scholar] [CrossRef] [PubMed]
- John, D.; Rindi, F. Filamentous (Nonconjugating) and Plantlike Green Algae; Academic Press: Cambridge, MA, USA, 2015; pp. 375–427. [Google Scholar]
- Douglas, B. The Ecology of the Attached Diatoms and Other Algae in a Small Stony Stream. J. Ecol. 1958, 46, 295–322. [Google Scholar] [CrossRef]
- Michael, H.; Gerardi, B.L. Green Algae and Diatoms. In The Biology and Troubleshooting of Facultative Lagoons; John Wiley & Sons: Hoboken, NJ, USA, 2015; pp. 85–96. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Environmental Factors | The Initial Stage of the Experiment | The End of the Experiment | ||||||
|---|---|---|---|---|---|---|---|---|
| F1 | F2 | F3 | F4 | F1 | F2 | F3 | F4 | |
| Flow velocity (m/s) | 0.086 | 0.125 | 0.211 | 0.418 | 0.086 | 0.125 | 0.211 | 0.418 |
| T (°C) | 15.84 | 15.84 | 15.84 | 15.84 | 18.40 | 18.40 | 18.40 | 18.40 |
| pH | 7.520 | 7.533 | 7.545 | 7.564 | 7.582 | 7.569 | 7.564 | 7.570 |
| DO (mg/L) | 10.42 | 10.46 | 10.49 | 10.35 | 9.92 | 9.99 | 10.05 | 10.10 |
| TN (mg/L) | 14.34 | 13.25 | 12.85 | 12.77 | 5.05 | 6.20 | 4.60 | 5.25 |
| TP (mg/L) | 0.11 | 0.10 | 0.10 | 0.11 | 0.09 | 0.08 | 0.10 | 0.11 |
| COD (mg/L) | 27 | 34 | 33 | 30 | 31 | 33 | 31 | 26 |
| NH4+ (mg/L) | 0.14 | 0.08 | 0.07 | 0.04 | 0.13 | 0.14 | 0.21 | 0.15 |
| NO3− (mg/L) | 6.60 | 6.45 | 5.25 | 6.17 | 4.30 | 4.10 | 4.55 | 4.45 |
| NO2− (mg/L) | 0.048 | 0.076 | 0.083 | 0.052 | 0.013 | 0.013 | 0.013 | 0.014 |
| Sample | Abundance | Shannon | Simpson | Pielou | Chao1 |
|---|---|---|---|---|---|
| F1 | 15 | 0.17 | 0.05 | 0.06 | 20.00 |
| F2 | 24 | 1.06 | 0.38 | 0.33 | 25.20 |
| F3 | 25 | 1.95 | 0.74 | 0.61 | 30.00 |
| F4 | 19 | 0.38 | 0.14 | 0.13 | 20.50 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, L.; Wu, L.; Liu, X.; Huang, W.; Zhu, D.; Wang, Z.; Guan, R.; Liu, X. Reducing the Risk of Benthic Algae Outbreaks by Regulating the Flow Velocity in a Simulated South–North Water Diversion Open Channel. Int. J. Environ. Res. Public Health 2023, 20, 3564. https://doi.org/10.3390/ijerph20043564
Sun L, Wu L, Liu X, Huang W, Zhu D, Wang Z, Guan R, Liu X. Reducing the Risk of Benthic Algae Outbreaks by Regulating the Flow Velocity in a Simulated South–North Water Diversion Open Channel. International Journal of Environmental Research and Public Health. 2023; 20(4):3564. https://doi.org/10.3390/ijerph20043564
Chicago/Turabian StyleSun, Longfei, Leixiang Wu, Xiaobo Liu, Wei Huang, Dayu Zhu, Zhuowei Wang, Ronghao Guan, and Xingchen Liu. 2023. "Reducing the Risk of Benthic Algae Outbreaks by Regulating the Flow Velocity in a Simulated South–North Water Diversion Open Channel" International Journal of Environmental Research and Public Health 20, no. 4: 3564. https://doi.org/10.3390/ijerph20043564