An Altered DNA Methylation Status in the Human Umbilical Cord Is Correlated with Maternal Exposure to Polychlorinated Biphenyls

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Subjects
2.3. PCB Analysis
2.4. DNA Methylation Analysis
2.5. Pyrosequencing Analysis
2.6. Data Analysis
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Barker, D.J.; Winter, P.D.; Osmond, C.; Margetts, B.; Simmonds, S.J. Weight in infancy and death from ischaemic heart disease. Lancet 1989, 2, 577–580. [Google Scholar] [CrossRef]
- Barker, D.J.P. Developmental origins of adult health and disease. J. Epidemiol. Community Health 2004, 58, 114–115. [Google Scholar] [CrossRef] [Green Version]
- Sata, F. Developmental Origins of Health and Disease (DOHaD) Cohorts and Interventions: Status and Perspective. In Pre-emptive Medicine: Public Health Aspects of Developmental Origins of Health and Disease; Sata, F., Fukuoka, H., Hanson, M., Eds.; Springer: Singapore, 2019; pp. 53–70. [Google Scholar] [CrossRef]
- Barrett, J.R. Programming the Future Epigenetics in the Context of DOHaD. Environ. Health Perspect. 2017, 125, A72. [Google Scholar] [CrossRef] [PubMed]
- M’hamdi, H.I.; de Beaufort, I.; Jack, B.; Steegers, E.A.P. Responsibility in the age of Developmental Origins of Health and Disease (DOHaD) and epigenetics. J. Dev. Orig. Health Dis. 2018, 9, 58–62. [Google Scholar] [CrossRef] [PubMed]
- Rusiecki, J.A.; Baccarelli, A.; Bollati, V.; Tarantini, L.; Moore, L.E.; Bonefeld-Jorgensen, E.C. Global DNA Hypomethylation Is Associated with High Serum-Persistent Organic Pollutants in Greenlandic Inuit. Environ. Health Perspect. 2008, 116, 1547–1552. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.Y.; Kim, D.S.; Lee, S.K.; Lee, I.K.; Kang, J.H.; Chang, Y.S.; Jacobs, D.R.; Steffes, M.; Lee, D.H. Association of Low-Dose Exposure to Persistent Organic Pollutants with Global DNA Hypomethylation in Healthy Koreans. Environ. Health Perspect. 2010, 118, 370–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, M.H.; Cho, E.R.; Lim, J.E.; Jee, S.H. Association between serum persistent organic pollutants and DNA methylation in Korean adults. Environ. Res. 2017, 158, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Fukata, H.; Mori, C. Epigenetic alteration by the chemical substances, food and environmental factors. Reprod. Med. Biol. 2004, 3, 115–121. [Google Scholar] [CrossRef] [Green Version]
- Waterland, R.A.; Michels, K.B. Epigenetic epidemiology of the developmental origins hypothesis. Annu. Rev. Nutr. 2007, 27, 363–388. [Google Scholar] [CrossRef]
- Gallou-Kabani, C.; Gabory, A.; Tost, J.; Karimi, M.; Mayeur, S.; Lesage, J.; Boudadi, E.; Gross, M.S.; Taurelle, J.; Vige, A.; et al. Sex-and diet-specific changes of imprinted gene expression and DNA methylation in mouse placenta under a high-fat diet. PLoS ONE 2010, 5, e14398. [Google Scholar] [CrossRef]
- Sinclair, K.D.; Allegrucci, C.; Singh, R.; Gardner, D.S.; Sebastian, S.; Bispham, J.; Thurston, A.; Huntley, J.F.; Rees, W.D.; Maloney, C.A.; et al. DNA methylation, insulin resistance, and blood pressure in offspring determined by maternal periconceptional B vitamin and methionine status. Proc. Natl. Acad. Sci. USA 2007, 104, 19351–19356. [Google Scholar] [CrossRef] [Green Version]
- Breitling, L.P.; Yang, R.; Korn, B.; Burwinkel, B.; Brenner, H. Tobacco-smoking-related differential DNA methylation: 27K discovery and replication. Am. J. Hum. Genet. 2011, 88, 450–457. [Google Scholar] [CrossRef]
- Lee, D.H.; Jacobs, D.R.; Porta, M. Hypothesis: A Unifying Mechanism for Nutrition and Chemicals as Lifelong Modulators of DNA Hypomethylation. Environ. Health Perspect. 2009, 117, 1799–1802. [Google Scholar] [CrossRef]
- Chervona, Y.; Costa, M. The control of histone methylation and gene expression by oxidative stress, hypoxia, and metals. Free Radic. Biol. Med. 2012, 53, 1041–1047. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Hernandez, A.; Kuo, C.C.; Rentero-Garrido, P.; Tang, W.Y.; Redon, J.; Ordovas, J.M.; Navas-Acien, A.; Tellez-Plaza, M. Environmental chemicals and DNA methylation in adults: A systematic review of the epidemiologic evidence. Clin. Epigenet. 2015, 7, 55. [Google Scholar] [CrossRef]
- Safe, S.H. Polychlorinated biphenyls (PCBs): Environmental impact, biochemical and toxic responses, and implications for risk assessment. Crit. Rev. Toxicol. 1994, 24, 87–149. [Google Scholar] [CrossRef]
- Longnecker, M.P.; Wolff, M.S.; Gladen, B.C.; Brock, J.W.; Grandjean, P.; Jacobson, J.L.; Korrick, S.A.; Rogan, W.J.; Weisglas-Kuperus, N.; Hertz-Picciotto, I.; et al. Comparison of polychlorinated biphenyl levels across studies of human neurodevelopment. Environ. Health Perspect. 2003, 111, 65–70. [Google Scholar] [CrossRef]
- Yamashita, F.; Hayashi, M. Fetal Pcb Syndrome—Clinical-Features, Intrauterine Growth-Retardation and Possible Alteration in Calcium-Metabolism. Environ. Health Perspect. 1985, 59, 41–45. [Google Scholar] [CrossRef]
- Furue, M.; Uenotsuchi, T.; Urabe, K.; Ishikawa, T.; Kuwabara, M.; Yusho, S.G. Overview of Yusho. J. Dermatol. Sci. 2005, S3–S10. [Google Scholar] [CrossRef]
- Hsu, C.-C.; Yu, M.-L.M.; Chen, Y.-C.J.; Guo, Y.-L.L.; Rogan, W.J. The Yu-cheng Rice Oil Poisoning Incident. In Dioxins and Health; Schecter, A., Ed.; Springer: Boston, MA, USA, 1994; pp. 661–684. [Google Scholar] [CrossRef]
- Rogan, W.J.; Gladen, B.C.; Hung, K.L.; Koong, S.L.; Shih, L.Y.; Taylor, J.S.; Wu, Y.C.; Yang, D.; Ragan, N.B.; Hsu, C.C. Congenital Poisoning by Polychlorinated-Biphenyls and Their Contaminants in Taiwan. Science 1988, 241, 334–336. [Google Scholar] [CrossRef]
- Su, K.Y.; Li, M.C.; Lee, N.W.; Ho, B.C.; Cheng, C.L.; Chuang, Y.C.; Yu, S.L.; Guo, Y.L. Perinatal polychlorinated biphenyls and polychlorinated dibenzofurans exposure are associated with DNA methylation changes lasting to early adulthood: Findings from Yucheng second generation. Environ. Res. 2019, 170, 481–486. [Google Scholar] [CrossRef]
- Rogan, W.J. PCBs and cola-colored babies: Japan, 1968, and Taiwan, 1979. Teratology 1982, 26, 259–261. [Google Scholar] [CrossRef]
- Mitoma, C.; Uchi, H.; Tsukimori, K.; Yamada, H.; Akahane, M.; Imamura, T.; Utani, A.; Furue, M. Yusho and its latest findings-A review in studies conducted by the Yusho Group. Environ. Int. 2015, 82, 41–48. [Google Scholar] [CrossRef]
- Kuratsune, M.; Yoshimura, H.; Hori, Y.; Okumura, M.; Masuda, Y. Yusho: A Human Disaster Caused by PCBs and Related Compounds; Kyushu University Press: Fukuoka, Japan, 1996. [Google Scholar]
- Fukata, H.; Omori, M.; Osada, H.; Todaka, E.; Mori, C. Necessity to measure PCBs and organochlorine pesticide concentrations in human umbilical cords for fetal exposure assessment. Environ. Health Perspect. 2005, 113, 297–303. [Google Scholar] [CrossRef]
- Aylward, L.L.; Hays, S.M.; Kirman, C.R.; Marchitti, S.A.; Kenneke, J.F.; English, C.; Mattison, D.R.; Becker, R.A. Relationships of Chemical Concentrations in Maternal and Cord Blood: A Review of Available Data. J. Toxicol. Environ. Health B 2014, 17, 175–203. [Google Scholar] [CrossRef]
- Weissman, A.; Jakobi, P.; Bronshtein, M.; Goldstein, I. Sonographic Measurements of the Umbilical-Cord and Vessels during Normal Pregnancies. J. Ultrasound Med. 1994, 13, 11–14. [Google Scholar] [CrossRef]
- Sakurai, K.; Shioda, K.; Eguchi, A.; Watanabe, M.; Miyaso, H.; Mori, C.; Shioda, T. DNA methylome of human neonatal umbilical cord: Enrichment of differentially methylated regions compared to umbilical cord blood DNA at transcription factor genes involved in body patterning and effects of maternal folate deficiency or children’s sex. PLoS ONE 2019, 14. [Google Scholar] [CrossRef]
- Sakurai, K.; Miyaso, H.; Eguchi, A.; Matsuno, Y.; Yamamoto, M.; Todaka, E.; Fukuoka, H.; Hata, A.; Mori, C. Chiba study of Mother and Children’s Health (C-MACH): Cohort study with omics analyses. BMJ Open 2016, 6. [Google Scholar] [CrossRef]
- Eguchi, A.; Sakurai, K.; Watanabe, M.; Mori, C. Exploration of potential biomarkers and related biological pathways for PCB exposure in maternal and cord serum: A pilot birth cohort study in Chiba, Japan. Environ. Int. 2017. [Google Scholar] [CrossRef]
- Eguchi, A.; Enomoto, T.; Suzuki, N.; Okuno, M.; Mori, C. Development of Simple Analytical Methods of Polychlorinated Biphenyls in Human Serum by Gas Chromatography Negative Ion Chemical Ionization Quadrupole Mass Spectrometry. Acta Chromatogr. 2017, 29, 503–506. [Google Scholar] [CrossRef]
- Tachibana, K.; Sakurai, K.; Watanabe, M.; Miyaso, H.; Mori, C. Associations between changes in the maternal gut microbiome and differentially methylated regions of diabetes-associated genes in fetuses: A pilot study from a birth cohort study. J. Diabetes Investig. 2017. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. Available online: http://www.R-project.org/ (accessed on 30 July 2019).
- Revelle, W. Psych: Procedures for Psychological, Psychometric, and Personality Research. Available online: https://CRAN.R-project.org/package=psych (accessed on 30 July 2019).
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate—A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Hanna, C.W.; Bloom, M.S.; Robinson, W.P.; Kim, D.; Parsons, P.J.; Saal, F.S.V.; Taylor, J.A.; Steuerwald, A.J.; Fujimoto, V.Y. DNA methylation changes in whole blood is associated with exposure to the environmental contaminants, mercury, lead, cadmium and bisphenol A, in women undergoing ovarian stimulation for IVF. Hum. Reprod. 2012, 27, 1401–1410. [Google Scholar] [CrossRef]
- Manikkam, M.; Tracey, R.; Guerrero-Bosagna, C.; Skinner, M.K. Dioxin (TCDD) Induces Epigenetic Transgenerational Inheritance of Adult Onset Disease and Sperm Epimutations. PLoS ONE 2012, 7. [Google Scholar] [CrossRef]
- Georgiadis, P.; Gavriil, M.; Rantakokko, P.; Ladoukakis, E.; Botsivali, M.; Kelly, R.S.; Bergdahl, I.A.; Kiviranta, H.; Vermeulen, R.C.H.; Spaeth, F.; et al. DNA methylation profiling implicates exposure to PCBs in the pathogenesis of B-cell chronic lymphocytic leukemia. Environ. Int. 2019, 126, 24–36. [Google Scholar] [CrossRef]
- Hany, J.; Lilienthal, H.; Sarasin, A.; Roth-Harer, A.; Fastabend, A.; Dunemann, L.; Lichtensteiger, W.; Winneke, G. Developmental exposure of rats to a reconstituted PCB mixture or Aroclor 1254: Effects on organ weights, aromatase activity, sex hormone levels, and sweet preference behavior. Toxicol. Appl. Pharm. 1999, 158, 231–243. [Google Scholar] [CrossRef]
- Amin, S.; Moore, R.W.; Peterson, R.E.; Schantz, S.L. Gestational and lactational exposure to TCDD or coplanar PCBs alters adult expression of saccharin preference behavior in female rats. Neurotoxicol. Teratol. 2000, 22, 675–682. [Google Scholar] [CrossRef]
- Geller, A.M.; Oshiro, W.M.; Haykal-Coates, N.; Kodavanti, P.R.S.; Bushnell, P.J. Gender-dependent behavioral and sensory effects of a commercial mixture of polychlorinated biphenyls (Aroclor 1254) in rats. Toxicol. Sci. 2001, 59, 268–277. [Google Scholar] [CrossRef]
- Kaya, H.; Hany, J.; Fastabend, A.; Roth-Harer, A.; Winneke, G.; Lilienthal, H. Effects of maternal exposure to a reconstituted mixture of polychlorinated biphenyls on sex-dependent behaviors and steroid hormone concentrations in rats: Dose-response relationship. Toxicol. Appl. Pharm. 2002, 178, 71–81. [Google Scholar] [CrossRef]
- Wang, X.Q.; Fang, J.; Nunez, A.A.; Clemens, L.G. Developmental exposure to polychlorinated biphenyls affects sexual behavior of rats. Physiol. Behav. 2002, 75, 689–696. [Google Scholar] [CrossRef]
- Kodavanti, P.R.S.; Kannan, N.; Yamashita, N.; Derr-Yellin, E.C.; Ward, T.R.; Burgin, D.E.; Tilson, H.A.; Birnbaum, L.S. Differential effects of two lots of Aroclor 1254: Congener-specific analysis and neurochemical end points. Environ. Health Perspect. 2001, 109, 1153–1161. [Google Scholar] [CrossRef]
- Guo, Y.L.; Lai, T.J.; Chen, S.J.; Hsu, C.C. Gender-Related Decrease in Ravens Progressive Matrices Scores in Children Prenatally Exposed to Polychlorinated-Biphenyls and Related Contaminants. Bull. Environ. Contam. Toxicol. 1995, 55, 8–13. [Google Scholar] [CrossRef]
- Shoji, M.; Tanaka, T.; Hosokawa, M.; Reuter, M.; Stark, A.; Kato, Y.; Kondoh, G.; Okawa, K.; Chujo, T.; Suzuki, T.; et al. The TDRD9-MIWI2 complex is essential for piRNA-mediated retrotransposon silencing in the mouse male germline. Dev. Cell 2009, 17, 775–787. [Google Scholar] [CrossRef]
- Arafat, M.; Har-Vardi, I.; Harlev, A.; Levitas, E.; Zeadna, A.; Abofoul-Azab, M.; Dyomin, V.; Sheffield, V.C.; Lunenfeld, E.; Huleihel, M.; et al. Mutation in TDRD9 causes non-obstructive azoospermia in infertile men. J. Med. Genet. 2017, 54, 633–639. [Google Scholar] [CrossRef]
- Amir, R.E.; Van den Veyver, I.B.; Wan, M.; Tran, C.Q.; Francke, U.; Zoghbi, H.Y. Rett syndrome is caused by mutations in X-linked MECP2, encoding methyl-CpG-binding protein 2. Nat. Genet. 1999, 23, 185. [Google Scholar] [CrossRef]
- Anway, M.D.; Cupp, A.S.; Uzumcu, M.; Skinner, M.K. Epigenetic transgenerational actions of endocrine disruptors and male fertility. Science 2005, 308, 1466–1469. [Google Scholar] [CrossRef]
- Zama, A.M.; Uzumcu, M. Fetal and neonatal exposure to the endocrine disruptor methoxychlor causes epigenetic alterations in adult ovarian genes. Endocrinology 2009, 150, 4681–4691. [Google Scholar] [CrossRef]
- Skinner, M.K.; Manikkam, M.; Tracey, R.; Guerrero-Bosagna, C.; Haque, M.; Nilsson, E.E. Ancestral dichlorodiphenyltrichloroethane (DDT) exposure promotes epigenetic transgenerational inheritance of obesity. BMC Med. 2013, 11, 228. [Google Scholar] [CrossRef]
- Alcock, R.E.; Behnisch, P.A.; Jones, K.C.; Hagenmaier, H. Dioxin-like PCBs in the environment-human exposure and the significance of sources. Chemosphere 1998, 37, 1457–1475. [Google Scholar] [CrossRef]
- Jin, W.; Otake, M.; Eguchi, A.; Sakurai, K.; Nakaoka, H.; Watanabe, M.; Todaka, E.; Mori, C. Dietary Habits and Cooking Methods Could Reduce Avoidable Exposure to PCBs in Maternal and Cord Sera. Sci. Rep. 2017, 7, 17357. [Google Scholar] [CrossRef]
- Lin, X.; Tan, J.Y.L.; Teh, A.L.; Lim, I.Y.; Liew, S.J.; MacIsaac, J.L.; Chong, Y.S.; Gluckman, P.D.; Kobor, M.S.; Cheong, C.Y.; et al. Cell type-specific DNA methylation in neonatal cord tissue and cord blood: A 850K-reference panel and comparison of cell types. Epigenetics 2018, 13, 941–958. [Google Scholar] [CrossRef]


{kind=link}
| Male | Female | p-Value | |
|---|---|---|---|
| Gender of newborns (n) | 32 | 43 | |
| Total PCBs (pg/g wet wt) (median [IQR *]) | 72 [52, 120] | 66 [44, 100] | 0.247 |
| Body weight at birth (g) (median [IQR]) | 2981 [2774: 3,164] | 3052 [2898: 3195] | 0.360 |
| Male | Female | p-Value | |
|---|---|---|---|
| Gender of newborns (n) | 27 | 41 | |
| Age (year) (median [IQR *]) | 33.0 [31.5, 36.0] | 32.0 [31.0, 36.0] | 0.431 |
| Height (cm) (median [IQR]) | 159.0 [154.9, 164.7] | 160.0 [156.0, 162.0] | 0.945 |
| Body weight at 10 gestational weeks (kg) (median [IQR]) | 53.0 [48.9, 56.5] | 52.0 [48.0, 56.0] | 0.716 |
| Probe ID | MAP INFO | Strand | Primer Types | Primer Sequence | Anneal (°C) | Product (bp) |
|---|---|---|---|---|---|---|
| cg01083397 | CHR X 153362990 | Upper Strand | Forward * | GAGGGTAGAGAGGAGGGA | 52 | 150 |
| Reverse | AACCAAAAAAAAAAACTATAAATAAAACC | |||||
| Sequence | ACCCAAAAACCAAAATCAAAAA | |||||
| cg06154002 | CHR 14 104394776 | Upper Strand | Forward | GTTTTGATTGGAAGGTTT | 54 | 125 |
| cg06289566 | CHR 14 104394782 | Reverse * | TCCCCAACATCCTCAAAACCCA | |||
| Sequence | GGGAGGGGTTTTTAGGG | |||||
| cg09878117 | CHR X 45649477 | Lower Strand | Forward | GTTAGTTGGGTGGTTTTTTATGTTTTAG | 54 | 138 |
| Reverse * | TCCACCCTATTTTTCCCAAAAC | |||||
| Sequence | TGTTTTTGTTTATTTTTTAGAGTT | |||||
| cg12838902 | CHR 7 5322586 (−200bp) | Lower Strand | Forward | GGGTTTTAGGGATAGGGG | 52 | 106 |
| Reverse * | AAAACCTATCCAAACCTTTATCTAC | |||||
| Sequence | GGGAGGGAGGGAGGGGG | |||||
| cg13368805 | CHR X 47420179 | Upper Strand | Forward * | ATTATAGTTTTTTAGAGTTGGTGAATA | 54 | 117 |
| Reverse | CCAATACCCTAAACACCCTTTTAT | |||||
| Sequence | CTTTAAAATAAATTAAAATCAAAAC |
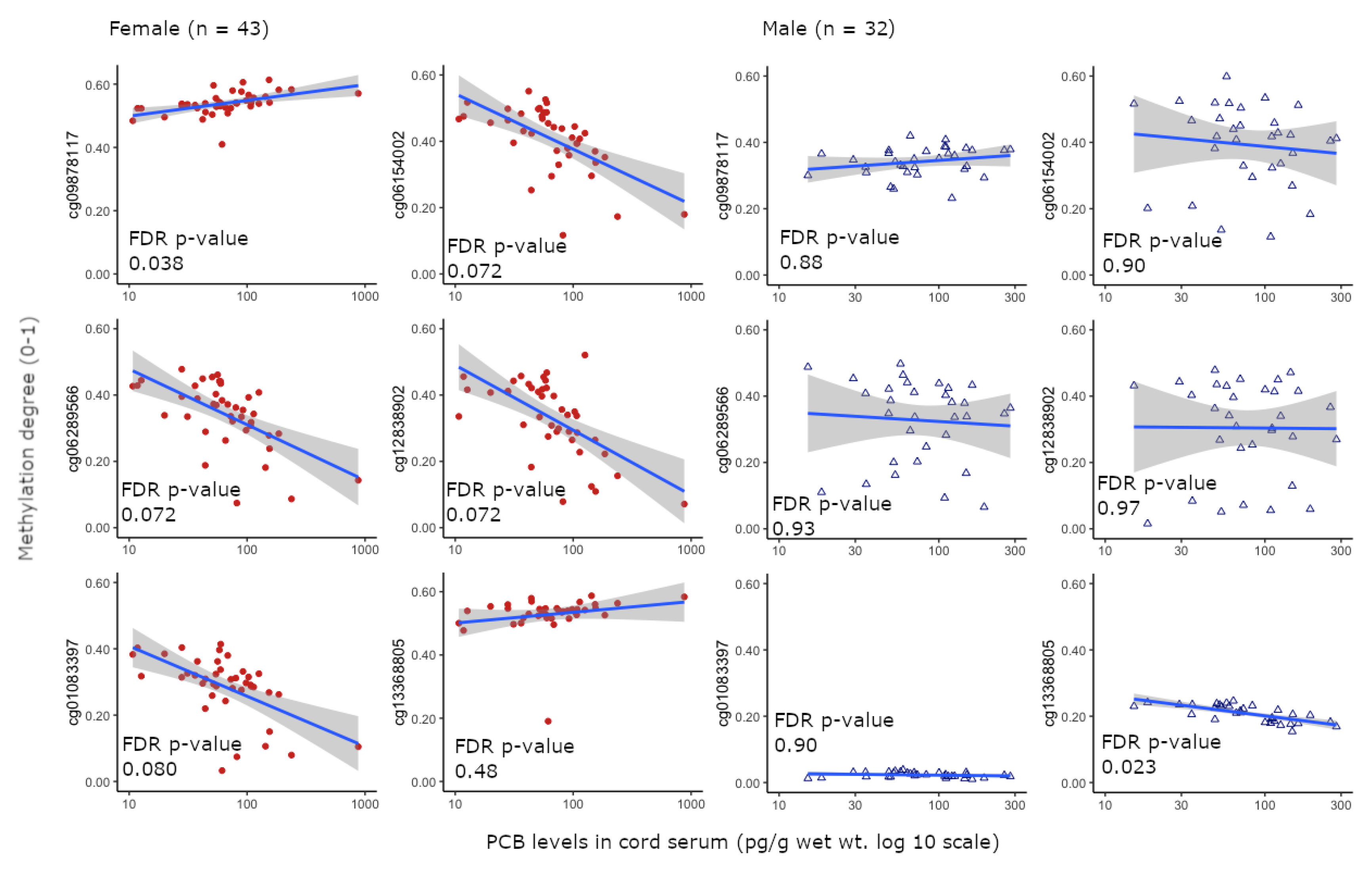
| FDR a | Ilmn ID | UCSC RefGene Name b | UCSC RefGene Group c | SNP ID d | SNP DISTANCE e |
|---|---|---|---|---|---|
| Female: 0.038 (positive) Male: 0.88 | cg09878117 | NA | NA | NA | NA |
| Female: 0.072 (negative) Male: 0.90 | cg06154002 ‡ | TDRD9 | TSS200 | rs530246903; rs113496080 | 24;4 |
| Female: 0.072 (negative) Male: 0.93 | cg06289566 | TDRD9 | TSS200 | rs530246903; rs113496080 | 30;10 |
| Female: 0.072 (negative) Male: 0.97 | cg12838902 | SLC29A4 | 5’UTR;1st exon | NA | NA |
| Female: 0.080 (negative) Male: 0.90 | cg01083397 | MECP2 | Body;5’UTR | NA | NA |
| Female: 0.48 Male: 0.023 (negative) | cg13368805 | ARAF | TSS1500 | NA | NA |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eguchi, A.; Nishizawa-Jotaki, S.; Tanabe, H.; Rahmutulla, B.; Watanabe, M.; Miyaso, H.; Todaka, E.; Sakurai, K.; Kaneda, A.; Mori, C. An Altered DNA Methylation Status in the Human Umbilical Cord Is Correlated with Maternal Exposure to Polychlorinated Biphenyls. Int. J. Environ. Res. Public Health 2019, 16, 2786. https://doi.org/10.3390/ijerph16152786
Eguchi A, Nishizawa-Jotaki S, Tanabe H, Rahmutulla B, Watanabe M, Miyaso H, Todaka E, Sakurai K, Kaneda A, Mori C. An Altered DNA Methylation Status in the Human Umbilical Cord Is Correlated with Maternal Exposure to Polychlorinated Biphenyls. International Journal of Environmental Research and Public Health. 2019; 16(15):2786. https://doi.org/10.3390/ijerph16152786
Chicago/Turabian StyleEguchi, Akifumi, Shino Nishizawa-Jotaki, Hiromi Tanabe, Bahityar Rahmutulla, Masahiro Watanabe, Hidenobu Miyaso, Emiko Todaka, Kenichi Sakurai, Atsushi Kaneda, and Chisato Mori. 2019. "An Altered DNA Methylation Status in the Human Umbilical Cord Is Correlated with Maternal Exposure to Polychlorinated Biphenyls" International Journal of Environmental Research and Public Health 16, no. 15: 2786. https://doi.org/10.3390/ijerph16152786