
Toxicogenomic Effects of Dissolved Saxitoxin on the Early Life Stages of the Longfin Yellowtail (Seriola rivoliana)

Abstract
:
1. Introduction
2. Results
2.1. Hatching Percentages
2.2. Gene Expression Analyses
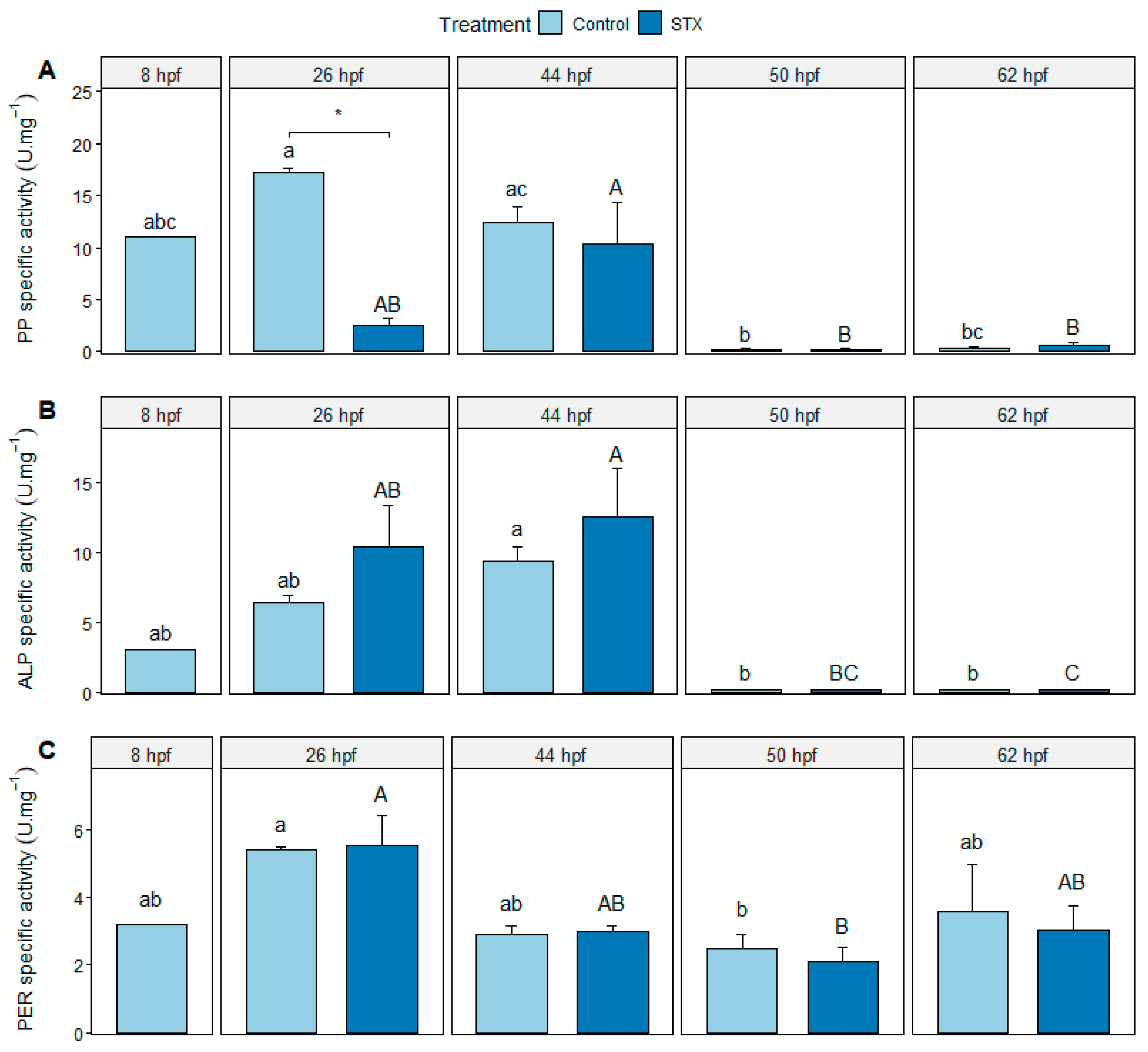
2.3. Enzyme Analyses
3. Discussion
3.1. Reduction of Hatching Percentage due to STX Exposure
3.2. Heat Shock Protein Induction
3.3. Regulation of Voltage-Gated Sodium Channels via Gene Expression
3.4. No STX-Induced Oxidative Stress
3.5. Activation of Lipid Metabolism and Digestion-Related Pathways
3.6. Regulation of Protein Phosphatases
3.7. Implications of Other Factors in Embryonic and Larval Development
4. Materials and Methods
4.1. Acquisition, Extraction, and Quantification of Marine Toxins
4.2. Assessment of Embryonic Development and Larval Viability
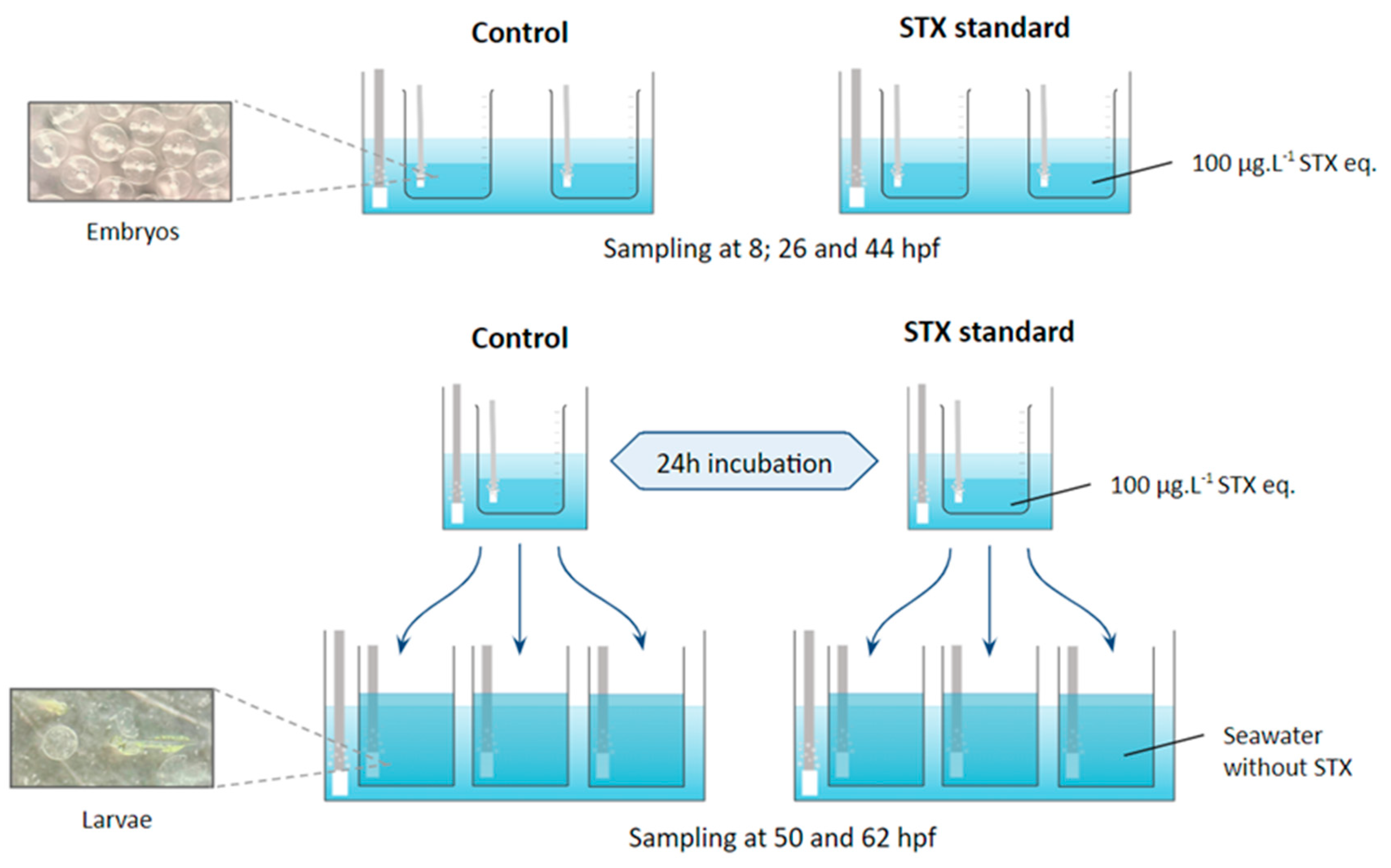
4.2.1. Experimental Protocol
4.2.2. Hatching Percentage
4.3. Gene Expression
4.3.1. Primer Design
4.3.2. RNA Extraction and cDNA Synthesis
4.3.3. Real-Time PCR
4.4. Protein Determination and Enzymatic Assays
4.5. Data Treatment and Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Anderson, D.M.; Cembella, A.D.; Hallegraeff, G.M. Progress in understanding harmful algal blooms: Paradigm shifts and new technologies for research, monitoring, and management. Ann. Rev. Mar. Sci. 2012, 4, 143–176. [Google Scholar] [CrossRef] [PubMed]
- Louzao, M.C.; Vilariño, N.; Vale, C.; Costas, C.; Cao, A.; Raposo-Garcia, S.; Vieytes, M.R.; Botana, L.M. Current Trends and New Challenges in Marine Phycotoxins. Marine Drugs 2022, 20, 198. [Google Scholar] [CrossRef] [PubMed]
- Yasumoto, T.; Murata, M. Marine Toxins. Chem. Rev. 1993, 24, 1897–1909. [Google Scholar] [CrossRef]
- Núñez-Vazquez, E.J.; Garate-Lizarraga, I.; Cordero-Tapia, A.; López-Cortes, D.J.; Hernández-Sandoval, F.E.; Heredia-Tapia, A.; Bustillos-Guzmán, J. Impact of harmful algal blooms on wild and cultured animals in the Gulf of California. J. Environ. Biol. 2011, 32, 413–423. [Google Scholar]
- Núñez-Vázquez, E.J.; Band-Schmidt, C.J.; Hernandez-Sandoval, F.E.; Bustillos-Guzman, J.J.; López-Cortés, D.J.; Cordero-Tapia, A.; Heredia-Tapia, A.; Garcia-Mendoza, E.; Peña-Manjarrez, J.; Ruiz-de la Torre, M.; et al. Impactos de los FAN en la salud pública y animal (silvestres y de cultivo) en el Golfo de California. In Florecimientos Algales Nocivos en México; García-Mendoza, E., Quijano-Sheggia, S.I., Olivos-Ortiz, A., Núñez-Vázquez, E.J., Eds.; CICESE: Mexico City, Mexico, 2016; pp. 196–212. [Google Scholar]
- FAO (Food and Agriculture Organization of the United Nations). Marine Biotoxins. 2. Paralytic Shellfish Poisoning (PSP); FAO: Rome, Italy, 2004; p. 281. [Google Scholar]
- Van Dolah, F. Marine Algal Toxins: Origins, Health Effects, and Their Increased Occurrence. Environ. Health Perspect. 2000, 108, 133–141. [Google Scholar] [CrossRef]
- Falconer, I.R.; Humpage, A.R. Health Risk Assessment of Cyanobacterial (Blue-green Algal) Toxins in Drinking Water. Int. J. Environ. Res. Public Health 2005, 2, 43–50. [Google Scholar] [CrossRef]
- Leal, J.F.; Cristiano, M.L.S. Marine paralytic shellfish toxins: Chemical properties, mode of action, newer analogues, and structure-toxicity relationship. Nat. Prod. Rep. 2022, 39, 33–57. [Google Scholar] [CrossRef]
- Vilariño, N.; Louzao, M.C.; Abal, P.; Cagide, E.; Carrera, C.; Vieytes, M.R.; Botana, L.M. Human Poisoning from Marine Toxins: Unknowns for Optimal Consumer Protection. Toxins 2018, 10, E324. [Google Scholar] [CrossRef]
- Stevens, M.; Peigneur, S.; Tytgat, J. Neurotoxins and their binding areas on voltage-gated sodium channels. Front. Pharmacol. 2011, 2, 71. [Google Scholar] [CrossRef]
- Thottumkara, A.P.; Parsons, W.H.; Du Bois, J. Saxitoxin. Angew. Chem. 2014, 53, 5760–5784. [Google Scholar] [CrossRef]
- Su, Z.; Sheets, M.; Ishida, H.; Li, F.; Barry, W.H. Saxitoxin Blocks L-Type ICa. J. Pharmacol. Exp. Ther. 2004, 308, 324–329. [Google Scholar] [CrossRef]
- Wang, J.; Salata, J.J.; Bennett, P.B. Saxitoxin Is a Gating Modifier of hERG K+ Channels. J. Gen. Physiol. 2003, 121, 583–598. [Google Scholar] [CrossRef] [PubMed]
- Band-Schmidt, C.J.; Duran-Riveroll, L.M.; Bustillos-Guzman, J.J.; Leyva-Valencia, I.; Lopez-Cortes, D.J.; Nunez-Vazquez, E.J.; Ramírez-Rodríguez, D.V. Paralytic toxin producing dinoflagellates in Latin America: Ecology and physiology. Front. Mar. Sci. 2019, 6, 42. [Google Scholar] [CrossRef]
- Schier, A.F.; Talbot, W.S. Molecular genetics of axis formation in zebrafish. Annu. Rev. Genet. 2005, 39, 561–613. [Google Scholar] [CrossRef] [PubMed]
- Espiña, B.; Louzao, M.C.; Cagide, E.; Alfonso, A.; Vieytes, M.R.; Yasumoto, T.; Botana, L.M. The methyl ester of okadaic acid is more potent than okadaic acid in disrupting the actin cytoskeleton and metabolism of primary cultured hepatocytes. Br. J. Pharmacol. 2010, 159, 337–344. [Google Scholar] [CrossRef]
- Guillotin, S.; Delcourt, N. Marine Neurotoxins’ Effects on Environmental and Human Health: An OMICS Overview. Marine Drugs 2021, 20, 18. [Google Scholar] [CrossRef]
- Escoffier, N.; Gaudin, J.; Mezhoud, K.; Huet, H.; Chateau-Joubert, S.; Turquet, J.; Crespeau, F.; Edery, M. Toxicity to medaka fish embryo development of okadaic acid and crude extracts of Prorocentrum dinoflagellates. Toxicon 2007, 49, 1182–1192. [Google Scholar] [CrossRef]
- Lefebvre, K.A.; Trainer, V.L.; Scholz, N.L. Morphological abnormalities and sensorimotor deficits in larval fish exposed to dissolved saxitoxin. Aquat. Toxicol. 2004, 66, 159–170. [Google Scholar] [CrossRef]
- Zhang, N.; Li, H.; Liu, J.; Yang, W. Gene expression profiles in zebrafish (Danio rerio) liver after acute exposure to okadaic acid. Environ. Toxicol. Pharmacol. 2014, 37, 791–802. [Google Scholar] [CrossRef]
- Vasconcelos, V.; Azevedo, J.; Silva, M.; Ramos, V. Effects of marine toxins on the reproduction and early stages development of aquatic organisms. Mar. Drugs 2010, 8, 59–79. [Google Scholar] [CrossRef]
- López-Cortés, D.J.; Núñez-Vázquez, E.J.; Dorantes-Aranda, J.J.; Band-Schmidt, C.J.; Hernandez-Sandoval, F.E.; Bustillos-Guzman, J.J.; Leyva-Valencia, I.; Fernández Herrera, L.J. The State of Knowledge of Harmful Algal Blooms of Margalefidinium polykrikoides (a.k.a. Cochlodinium polykrikoides) in Latin America. Front. Mar. Sci. 2019, 6, 463. [Google Scholar] [CrossRef]
- Núñez-Vázquez, E.J.; Heredia-Tapia, A.; Pérez-Urbiola, J.; Alonso, R.; Arellano-Blanco, J.; Cordero-Tapia, A.; Pérez-Linares, J. Ochoa, J.L Evaluation of dinoflagellate toxicity implicated in recent HAB events in the Gulf of California, México. In Proceedings from HABTech 2003 APEC: A Workshop on Technologies for Monitoring of Harmful Algal Blooms and Marine Biotoxins; Cawtron Report No. 906; Holland, P., Rhodes, L., Brown, L., Eds.; Cawthron Institute: Nelson, New Zealand, 2003; p. 64. [Google Scholar]
- Eschmeyer, W.N.; Herald, E.S.; Hammann, H. A Field Guide to Pacific Coast Fishes: North America; Houghton Mifflin: Boston, MA, USA, 1983. [Google Scholar]
- Cervigón, F. Los peces marinos de Venezuela. In Estación de Investigaciones Marinas de Margarita; Fundación La Salle de Ciencias Naturales: Caracas, Venezuela, 1993. [Google Scholar]
- Perez-Arellano, J.L.; Luzardo, O.P.; Brito, A.P.; Cabrera, M.H.; Zumbado, M.; Carranza, C.; Angel-Moreno, A.; Dickey, R.W.; Boada, L.D. Ciguatera fish poisoning, Canary Islands. Emerg. Infect. Dis. 2005, 11, 1981–1982. [Google Scholar] [CrossRef]
- Otero, P.; Pérez, S.; Alfonso, A.; Vale, C.; Rodríguez, P.; Gouveia, N.N.; Gouveia, N.; Delgado, J.; Vale, P.; Hirama, M.; et al. First toxin profile of ciguateric fish in Madeira Arquipelago (Europe). Anal. Chem. 2010, 82, 6032–6039. [Google Scholar] [CrossRef]
- Boada, L.D.; Zumbado, M.; Luzardo, O.R.; Almeida-Gonzalez, M.; Plakas, S.M.; Granade, H.R.; Abraham, A.; Jester, E.L.E.; Dickey, R.W. Ciguatera fish poisoning on the West Africa Coast: An emerging risk in the Canary Islands (Spain). Toxicon 2010, 56, 1516–1519. [Google Scholar] [CrossRef] [PubMed]
- Roo, J.; Fernández-Palacios, H.; Hernández-Cruz, C.M.; Mesa-Rodríguez, A.; Schuchardt, D.; Izquierdo, M. First results of spawning and larval rearing of longfin yellowtail Seriola rivoliana as a fast-growing candidate for European marine finfish aquaculture diversification. Aquac. Res. 2014, 45, 689–700. [Google Scholar] [CrossRef]
- Mazzola, A.; Favaloro, E.; Sarà, G. Cultivation of the Mediterranean amberjack, Seriola dumerili (Risso, 1810), in submerged cages in the Western Mediterranean Sea. Aquaculture 2000, 181, 257–268. [Google Scholar] [CrossRef]
- Mesa-Rodríguez, A.; Hernandez-Cruz, M.; Socorro, J.A.; Fernández-Palacios, H.; Izquierdo, M.; Roo, J. Skeletal Development and Mineralization Pattern of the Vertebral Column, Dorsal, Anal and Caudal Fin Complex in Seriola rivoliana (Valenciennes, 1833) Larvae. J. Aquac. Res Dev. 2014, 5, 2155–9546. [Google Scholar]
- Yamamoto, T.; Teruya, K.; Hara, T.; Hokazono, H.; Hashimoto, H.; Suzuki, N.; Iwashita, Y.; Matsunari, H.; Furuita, H.; Mushiake, K. Nutritional evaluation of live food organisms and commercial dry feeds used for seed production of amberjack Seriola dumerili. Fish. Sci. 2008, 74, 1096–1108. [Google Scholar] [CrossRef]
- Rountos, K.J.; Kim, J.J.; Hattenrath-Lehmann, T.K.; Gobler, C.J. Effects of the harmful algae, Alexandrium catenella and Dinophysis acuminata, on the survival, growth, and swimming activity of early life stages of forage fish. Mar. Environ. Res. 2019, 148, 46–56. [Google Scholar] [CrossRef] [PubMed]
- Gaillard, S.; Réveillon, D.; Mason, P.L.; Ayache, N.; Sanderson, M.; Smith, J.L.; Giddings, S.; McCarron, P.; Séchet, V.; Hégaret, H.; et al. Mortality and histopathology in sheepshead minnow (Cyprinodon variegatus) larvae exposed to pectenotoxin-2 and Dinophysis acuminata. Aquat. Toxicol. 2023, 257, 106456. [Google Scholar] [CrossRef]
- Brenta, A.; Núñez-Vázquez, E.J.; Asencio-Alcudia, G.; Teles, A.; Salas, J.; Álvarez-González, C.A.; Fernández-Herrera, L.J.; Tovar-Ramírez, D. Dissolved diarrhetic and paralytic shellfish toxins affects viability and lipid metabolism in Seriola rivoliana embryo. Aquatic Toxicol. 2023; submitted. [Google Scholar]
- Le Du, J.; Tovar-Ramírez, D.; Núñez-Vázquez, E.J. Embryotoxic effects of dissolved okadaic acid on the development of Longfin yellowtail Seriola rivoliana. Aquat.Toxicol. 2017, 190, 210–216. [Google Scholar] [CrossRef]
- Grossi, E.; Fernández-Palacios, H.; Abreu, N.; Socorro, J.A.; Roo, J.; Hernandez-Cruz, M.; Schuchardt, D. Organogénesis y morfometría de la fase lecitotrófica de larvas del medregal negro (Seriola rivoliana, Valenciennes,1883). In Proceedings of the XII Congreso Nacional de Acuicultura, Madrid, Spain, 24–26 November 2009. [Google Scholar] [CrossRef]
- Reis Costa, P. Impact and effects of paralytic shellfish poisoning toxins derived from harmful algal blooms to marine fish. Fish Fish. 2014, 17, 226–248. [Google Scholar] [CrossRef]
- Abi-Khalil, C.; Finkelstein, D.S.; Conejero, G.; Du Bois, J.; Destoumieux-Garzon, D.; Rolland, J.L. The paralytic shellfish toxin, saxitoxin, enters the cytoplasm and induces apoptosis of oyster immune cells through a caspase-dependent pathway. Aquat. Toxicol. 2017, 190, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Medhioub, W.; Ramondenc, S.; Vanhove, A.S.; Vergnes, A.; Masseret, E.; Savar, V.; Amzil, Z.; Laabir, M.; Rolland, J.L. Exposure to the Neurotoxic Dinoflagellate, Alexandrium catenella, Induces Apoptosis of the Hemocytes of the Oyster, Crassostrea gigas. Marine Drugs 2013, 11, 4799–4814. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Jia, Z.; Wang, L.; Hu, T. Effect of acute exposure of saxitoxin on development of zebrafish embryos (Danio rerio). Environl. Res. 2020, 185, 109432. [Google Scholar] [CrossRef] [PubMed]
- Iwama, G.; Thomas, P.; Forsyth, R.; Vijayan, M. Heat shock expression in fish. Rev. Fish Biol. Fish. 1998, 8, 35–56. [Google Scholar] [CrossRef]
- Iwama, G.; Afonso, L.; Todgham, A.; Ackerman, P.; Nakano, K. Are hsps suitable for indicating stressed states in fish? J. Exp. Biol. 2004, 207, 15–19. [Google Scholar] [CrossRef]
- Deng, D.-F.; Wang, C.; Lee, S.; Bai, S.; Hung, S.S.O. Feeding rates affect heat shock protein levels in liver of larval white sturgeon (Acipenser transmontanus). Aquaculture 2009, 287, 223–226. [Google Scholar] [CrossRef]
- Wang, J.; Wei, Y.; Li, X.; Cao, H.; Xu, M.; Dai, J. The identification of heat shock protein genes in goldfish (Carassius auratus) and their expression in a complex environment in Gaobeidian Lake, Beijing, China. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2007, 145, 350–362. [Google Scholar] [CrossRef]
- Li, Y.; Sun, B.; Wu, H.; Nie, P. Effects of pure microcystin-LR on the transcription of immune related genes and heat shock proteins in larval stage of zebrafish (Danio rerio). Aquaculture 2009, 289, 154–160. [Google Scholar] [CrossRef]
- Yuan, J.; Gu, Z.; Zheng, Y.; Zhang, Y.; Gao, J.; Chen, S.; Wang, Z. Accumulation, and detoxification dynamics of microcystin-LR and antioxidant responses in male red swamp crayfish Procambarus clarkii. Aquat. Toxicol. 2016, 177, 8–18. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Cheng, J.; Chen, X.; Cheng, S.H.; Mak, Y.L.; Lam, P.K.S.; Chan, L.L.; Wang, M. Early developmental toxicity of saxitoxin on medaka (Oryzias melastigma) embryos. Toxicon 2014, 77, 16–25. [Google Scholar] [CrossRef]
- Barbosa, V.; Santos, M.; Anacleto, P.; Maulvault, A.L.; Pousão-Ferreira, P.; Costa, P.R.; Marques, A. Paralytic Shellfish Toxins and Ocean Warming: Bioaccumulation and Ecotoxicological Responses in Juvenile Gilthead Seabream (Sparus aurata). Toxins 2019, 11, 408. [Google Scholar] [CrossRef] [PubMed]
- Pederzoli, A.; Mola, L. The early stress responses in fish larvae. Acta Histochem. 2016, 118, 443–449. [Google Scholar] [CrossRef] [PubMed]
- Murphy, S.P.; Gorzowski, J.J.; Sarge, K.D.; Phillips, B. Characterization of constitutive HSF2 DNA-binding activity in mouse embryonal carcinoma cells. Mol. Cell. Biol. 1994, 14, 5309–5317. [Google Scholar]
- Bakke, M.J.; Horsberg, T.E. Kinetic properties of saxitoxin in Atlantic salmon (Salmo salar) and Atlantic cod (Gadus morhua). Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2010, 152, 444–450. [Google Scholar] [CrossRef]
- Cestèle, S.; Catterall, W.A. Molecular mechanisms of neurotoxin action on voltage-gated sodium channels. Biochimie 2000, 82, 883–892. [Google Scholar] [CrossRef]
- Mat, A.; Klopp, C.; Payton, L.; Jeziorski, C.; Chalopin, M.; Amzil, Z.; Tran, D.; Wikfors, G.; Hégaret, H.; Soudant, P.; et al. Oyster transcriptome response to Alexandrium exposure is related to saxitoxin load and characterized by disrupted digestion, energy balance, and calcium and sodium signaling. Aquat. Toxicol. 2018, 199, 127–137. [Google Scholar] [CrossRef]
- Black, J.A.; Waxman, S.G. Noncanonical Roles of Voltage-Gated Sodium Channels. Neuron 2013, 80, 280–291. [Google Scholar] [CrossRef]
- Boulekbache, H. Energy Metabolism in Fish Development. Am. Zool. 1981, 21, 377–389. [Google Scholar] [CrossRef]
- Munteanu, C.; Iliuta, A. The role of sodium in the body. Balneo Res. J. 2011, 2, 70–74. [Google Scholar] [CrossRef]
- Zhou, J.; Li, N.; Wang, H.; Mu, C.; Wang, C. Effects of salinity on the ions important and sodium-potassium ATPase in osmoregulation, cortisol, amino acids, digestive and immune enzymes in Scylla paramamosain during indoor overwintering. Aquac. Res. 2020, 51, 4173–4182. [Google Scholar] [CrossRef]
- Van Thiel, J.; Ali Khan, M.; Wouters, R.; Harris, R.; Casewell, N.; Kini, R.; Mackessy, S.; Vonk, F.; Wüster, W.; Richardson, M. Convergent evolution of toxin resistance in animals. Biol. Rev. Camb. Philos. Soc. 2022, 97, 1823–1843. [Google Scholar] [CrossRef]
- Figueroa, D.; Signore, A.; Araneda, O.; Contreras, H.R.; Concha, M.; García, C. Toxicity and differential oxidative stress effects on zebrafish larvae following exposure to toxins from the okadaic acid group. J. Toxicol. Environ. Health Part A 2020, 83, 573–588. [Google Scholar] [CrossRef]
- Reis Costa, P.; Pereira, P.; Guilherme, S.; Barata, M.; Nicolau, L.; Santos, M.A.; Pacheco, M.; Pousão-Ferreira, P. Biotransformation modulation and genotoxicity in white seabream upon exposure to paralytic shellfish toxins produced by Gymnodinium catenatum. Aquat. Toxicol. 2012, 106–107, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Haque, M.N.; Nam, S.-E.; Han, Y.-S.; Park, H.S.; Rhee, J.-S. Chronic exposure to sublethal concentrations of saxitoxin reduces antioxidant activity and immunity in zebrafish but does not affect reproductive parameters. Aquat. Toxicol. 2021, 243, 106070. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Tang, X.; Zhou, B.; Zhou, Z.; Xu, N.; Wang, Y. A ROS-mediated mitochondrial pathway and Nrf2 pathway activation are involved in BDE-47 induced apoptosis in Neuro-2a cells. Chemosphere 2017, 184, 679–686. [Google Scholar] [CrossRef]
- Antonopoulou, E.; Kaitetzidou, E.; Castellana, B.; Panteli, N.; Kyriakis, D.; Vraskou, Y.; Planas, J.V. In Vivo Effects of Lipopolysaccharide on Peroxisome Proliferator-Activated Receptor Expression in Juvenile Gilthead Seabream (Sparus aurata). Biology 2017, 6, 36. [Google Scholar] [CrossRef]
- La Poulsen, L.C.; Siersbæk, M.; Mandrup, S. PPARs: Fatty acid sensors controlling metabolism. Semin. Cell Dev. Biol. 2012, 23, 631–639. [Google Scholar] [CrossRef]
- Wu, X.-Y.; Nie, L.; Lu, X.-J.; Fei, C.-J.; Chen, J. Molecular characterization, expression, and functional analysis of large yellow croaker (Larimichthys crocea) peroxisome proliferator-activated receptor gamma. Fish Shellfish Immunol. 2022, 123, 50–60. [Google Scholar] [CrossRef]
- Teles, A.; Salas-Leiva, J.; Alvarez-González, C.A.; Gisbert, E.; Ibarra-Castro, L.; Urbiola, J.C.P.; Tovar-Ramírez, D. Histological study of the gastrointestinal tract in longfin yellowtail (Seriola rivoliana) larvae. Fish Physiol. Biochem. 2017, 43, 1613–1628. [Google Scholar] [CrossRef]
- Atencio Genes, L.; Moreno, I.; Prieto, A.; Moyano, R.; Molina, A.; Cameán, A. Acute Effects of Microcystins MC-LR and MC-RR on Acid and Alkaline Phosphatase Activities and Pathological Changes in Intraperitoneally Exposed Tilapia Fish (Oreochromis sp.). Toxicol. Pathol. 2008, 36, 449–458. [Google Scholar] [CrossRef]
- Bates, J.M.; Akerlund, J.; Mittge, E.; Guillemin, K. Intestinal Alkaline Phosphatase Detoxifies Lipopolysaccharide and Prevents Inflammation in Zebrafish in Response to the Gut Microbiota. Cell Host Microbe 2007, 2, 371–382. [Google Scholar] [CrossRef]
- Lynes, M.; Narisawa, S.; Millán, J.L.; Widmaier, E.P. Interactions between CD36 and global intestinal alkaline phosphatase in mouse small intestine and effects of high-fat diet. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011, 301, R1738–R1747. [Google Scholar] [CrossRef] [PubMed]
- Pan, H.; Li, L.-Y.; Li, J.-M.; Wang, W.-L.; Limbu, S.M.; Degrace, P.; Li, D.-L.; Du, Z.-Y. Inhibited fatty acid β-oxidation impairs stress resistance ability in Nile tilapia (Oreochromis niloticus). Fish Shellfish Immunol. 2017, 68, 500–508. [Google Scholar] [CrossRef]
- Lefebvre, K.A.; Elder, N.E.; Hershberger, P.K.; Trainer, V.L.; Stehr, C.M.; Scholz, N.L. Dissolved saxitoxin causes transient inhibition of sensorimotor function in larval Pacific herring (Clupea harengus pallasi). Mar. Biol. 2005, 147, 1393–1402. [Google Scholar] [CrossRef]
- Barford, D. Molecular mechanisms of the protein serine/threonine phosphatases. Trends Biochem. Sci. 1996, 21, 407–412. [Google Scholar] [CrossRef] [PubMed]
- Takai, A.; Murata, M.; Torigoe, K.; Isobe, M.; Mieskes, G.; Yasumoto, T. Inhibitory effect of okadaic acid derivatives on protein phosphatases: A study on structure-affinity relationship. Biochem. J. 1992, 284 Pt 2, 539–544. [Google Scholar] [CrossRef]
- Shi, Z.Y.; Chen, X.W.; Gu, Y.F. Cloning, and expression pattern of alkaline phosphatase during the development of Paralichthys olivaceus. Fish Physiol. Biochem. 2011, 37, 411–424. [Google Scholar] [CrossRef]
- Kong, X.; Wang, S.; Jiang, H.; Nie, G.; Li, X. Responses of acid/alkaline phosphatase, lysozyme, and catalase activities and lipid peroxidation to mercury exposure during the embryonic development of goldfish Carassius aratus. Aquat. Toxicol. 2012, 120–121, 119–125. [Google Scholar] [CrossRef]
- O’Loghlen, A.; Pérez-Morgado, M.I.; Salinas, M.; Martín, M.E. Reversible inhibition of the protein phosphatase 1 by hydrogen peroxide. Potential regulation of eIF2 alpha phosphorylation in differentiated PC12 cells. Arch. Biochem. Biophys. 2003, 417, 194–202. [Google Scholar] [CrossRef]
- Sullivan, S.G.; Chiu, D.T.; Errasfa, M.; Wang, J.M.; Qi, J.S.; Stern, A. Effects of H2O2 on protein tyrosine phosphatase activity in HER14 cells. Free Radic. Biol. Med. 1994, 16, 399–403. [Google Scholar]
- Chang, L.; Karin, M. Mammalian MAP kinase signaling cascades. Nature 2001, 410, 37–40. [Google Scholar] [CrossRef] [PubMed]
- Owens, D.M.; Keyse, S.M. Differential regulation of MAP kinase signalling by dual-specificity protein phosphatases. Oncogene 2007, 26, 3203–3213. [Google Scholar] [CrossRef]
- Ding, T.; Zhou, Y.; Long, R.; Chen, C.; Zhao, J.; Cui, P.; Guo, M.; Liang, G.; Xu, L. DUSP8 phosphatase: Structure, functions, expression regulation and the role in human diseases. Cell Biosci. 2019, 9, 70. [Google Scholar] [CrossRef]
- Llewellyn, L. Saxitoxin, a toxic marine natural product that targets a multitude of receptors. Nat. Prod. Rep. 2006, 23, 200–222. [Google Scholar] [CrossRef]
- Alfonso, A.; Louzao, M.C.; Vieytes, M.R.; Botana, L.M. Comparative study of the stability of saxitoxin and neosaxitoxin in acidic solutions and lyophilized samples. Toxicon 1994, 32, 1593–1598. [Google Scholar] [CrossRef]
- Roggatz, C.C.; Fletcher, N.; Benoit, D.M.; Algar, A.C.; Doroff, A.; Wright, B.; Wollenberg Valero, K.C.; Hardege, J.D. Saxitoxin and tetrodotoxin bioavailability increases in future oceans. Nat. Clim. Change 2019, 9, 840–844. [Google Scholar] [CrossRef]
- AOAC. Official Method 959.08 Paralytic Shellfish Poison, Biological Method, Final Action. In Official Methods of Analysis of AOAC International, 16th ed.; Cunniff, P., Ed.; AOAC: Rockville, MD, USA, 1995; pp. 21–22. [Google Scholar]
- Teles, A.; Álvarez-González, C.A.; Llera-Herrera, R.; Gisbert, E.; Salas-Leiva, J.; del Rodríguez-Jaramillo, M.C.; Tovar-Ramírez, D. Debaryomyces hansenii CBS 8339 promotes larval development in Seriola rivoliana. Aquaculture 2023, 560, 738587. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Gee, K.R.; Sun, W.C.; Bhalgat, M.K.; Upson, R.H.; Klaubert, D.H.; Latham, K.A.; Haugland, R.P. Fluorogenic substrates based on fluorinated umbelliferones for continuous assays of phosphatases and beta-galactosidases. Anal. Biochem. 1999, 273, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Gosselin, S.; Fortier, L.; Gagné, J. Vulnerability of marine fish larvae to the toxic dinoflagellate Protogonyaulax tamarensis. Mar. Ecol. Prog. Ser. 1989, 57, 1–10. [Google Scholar] [CrossRef]
- Salierno, J.D.; Snyder, N.S.; Murphy, A.Z.; Poli, M.; Hall, S.; Baden, D.; Kane, A.S. Harmful algal bloom toxins alter c-Fos protein expression in the brain of killifish, Fundulus heteroclitus. Aquat. Toxicol. 2006, 78, 350–357. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, C.A.; de Morais, E.C.; Costa, M.D.; Ribas, J.L.; Guiloski, I.C.; Ramsdorf, W.A. Saxitoxins induce cytotoxicity, genotoxicity and oxidative stress in teleost neurons in vitro. Toxicon 2014, 86, 8–15. [Google Scholar] [CrossRef]
- Silva de Assis, H.C.; da Silva, C.A.; Oba, E.T.; Pamplona, J.H.; Mela, M.; Doria, H.B.; Guiloski, I.C.; Ramsdorf, W.; Cestari, M.M. Hematologic and hepatic responses of the freshwater fish Hoplias malabaricus after saxitoxin exposure. Toxicon 2013, 66, 25–30. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
 | |||||
| Toxin | TEFs 1 | R1 | R2 | R3 | R4 |
| STX | 1.0 | –H | –H | –H |  |
| NeoSTX | 0.92 | –OH | –H | –H | |
| GTX1 | 0.99 | –OH | –H | –HOSO3− | |
| GTX2 | 0.36 | –H | –H | –HOSO3− | |
| GTX3 | 0.64 | –H | –HOSO3− | –H | |
| GTX4 | 0.73 | –OH | –HOSO3− | –H | |
| GTX5 | 0.064 | –H | –H | –H |  |
| GTX6 | – | –OH | –H | –H | |
| C1 | 0.006 | –H | –H | –HOSO3− | |
| C2 | 0.096 | –H | –HOSO3− | –H | |
| C3 | 0.013 | –OH | –H | –HOSO3− | |
| C4 | 0.058 | –OH | –HOSO3− | –H | |
| dcSTX | 0.51 | –H | –H | –H | –OH Decarbamate |
| dcNeoSTX | – | –OH | –H | –H | |
| dcGTX1 | – | –OH | –H | –HOSO3− | |
| dcGTX2 | 0.15 | –H | –H | –HOSO3− | |
| dcGTX3 | 0.38 | –H | –HOSO3− | –H | |
| dcGTX4 | – | –OH | –HOSO3− | –H | |
| Gene Symbol | Target Gene | Primer | Sequence (5′–3′) |
|---|---|---|---|
| 18S | 18S ribosomal RNA | Forward Reverse | CTGAACTGGGGCCATGATTAAGAG GGTATCTGATCGTCGTCGAACCTC |
| HSF2 | Heat shock factor protein 2 | Forward Reverse | TTCATGGTGTTGGACGAGCA TGCTTGAAGTAGGGGTGCTG |
| NaV1.4b | Sodium channel protein type 4 subunit alpha B | Forward Reverse | TCCAGGACAACTCGAAAACC CGAAGTTGATCCAGTGCAGA |
| PPRC1 | Peroxisome proliferator-activated receptor gamma coactivator-related protein 1 | Forward Reverse | AACCCCAGCAAACACCTGAA ACACTTCCCATCTGCTGACG |
| DUSP8 | Dual specificity phosphatase 8 | Forward Reverse | CCTCACAGACAGGACACAACA GCTTTGGTGATGGTTTGACTG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guinle, C.; Núñez-Vázquez, E.J.; Fernández-Herrera, L.J.; Corona-Rojas, D.A.; Tovar-Ramírez, D. Toxicogenomic Effects of Dissolved Saxitoxin on the Early Life Stages of the Longfin Yellowtail (Seriola rivoliana). Mar. Drugs 2023, 21, 597. https://doi.org/10.3390/md21110597
Guinle C, Núñez-Vázquez EJ, Fernández-Herrera LJ, Corona-Rojas DA, Tovar-Ramírez D. Toxicogenomic Effects of Dissolved Saxitoxin on the Early Life Stages of the Longfin Yellowtail (Seriola rivoliana). Marine Drugs. 2023; 21(11):597. https://doi.org/10.3390/md21110597
Chicago/Turabian StyleGuinle, Colleen, Erick Julián Núñez-Vázquez, Leyberth José Fernández-Herrera, Daniela Alejandra Corona-Rojas, and Dariel Tovar-Ramírez. 2023. "Toxicogenomic Effects of Dissolved Saxitoxin on the Early Life Stages of the Longfin Yellowtail (Seriola rivoliana)" Marine Drugs 21, no. 11: 597. https://doi.org/10.3390/md21110597