
Oxidative Stability of Side-Streams from Cod Filleting—Effect of Antioxidant Dipping and Low-Temperature Storage
 ,
,
Abstract
:1. Introduction
2. Results
2.1. Crude Composition: Dry Matter, Protein and Oil Contents of Cod Solid Side-Streams
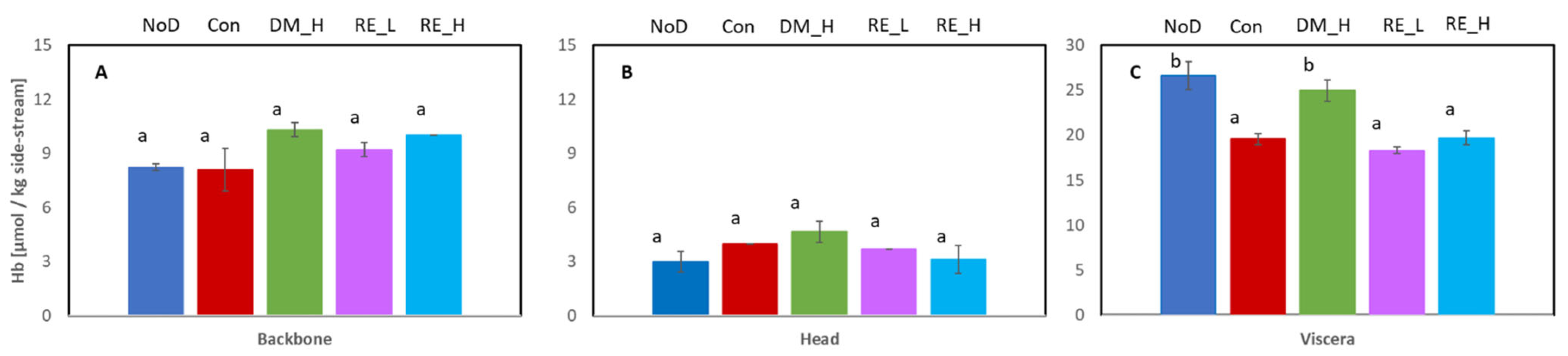
2.2. Initial Hb Content in the Different Cod Side-Streams
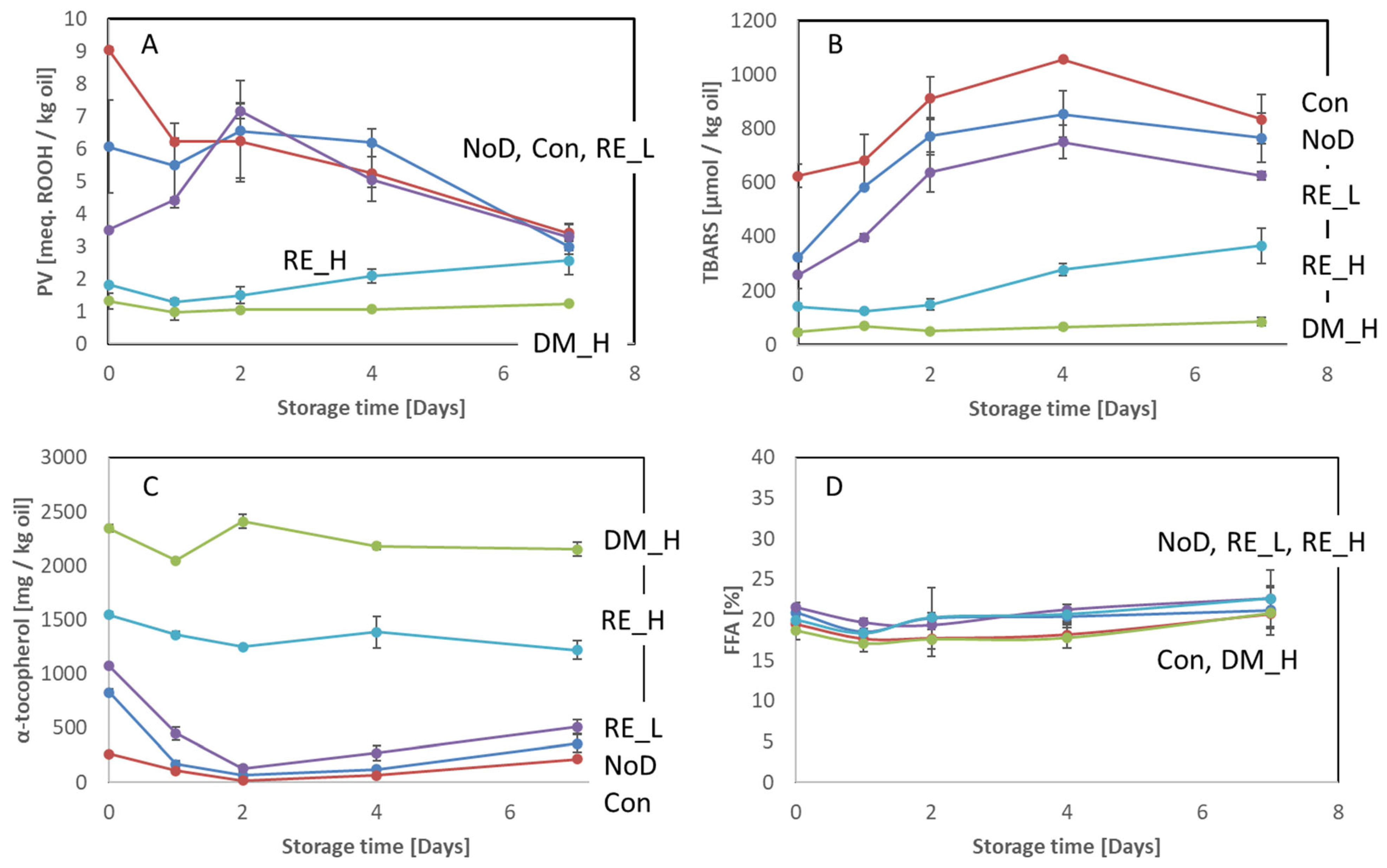
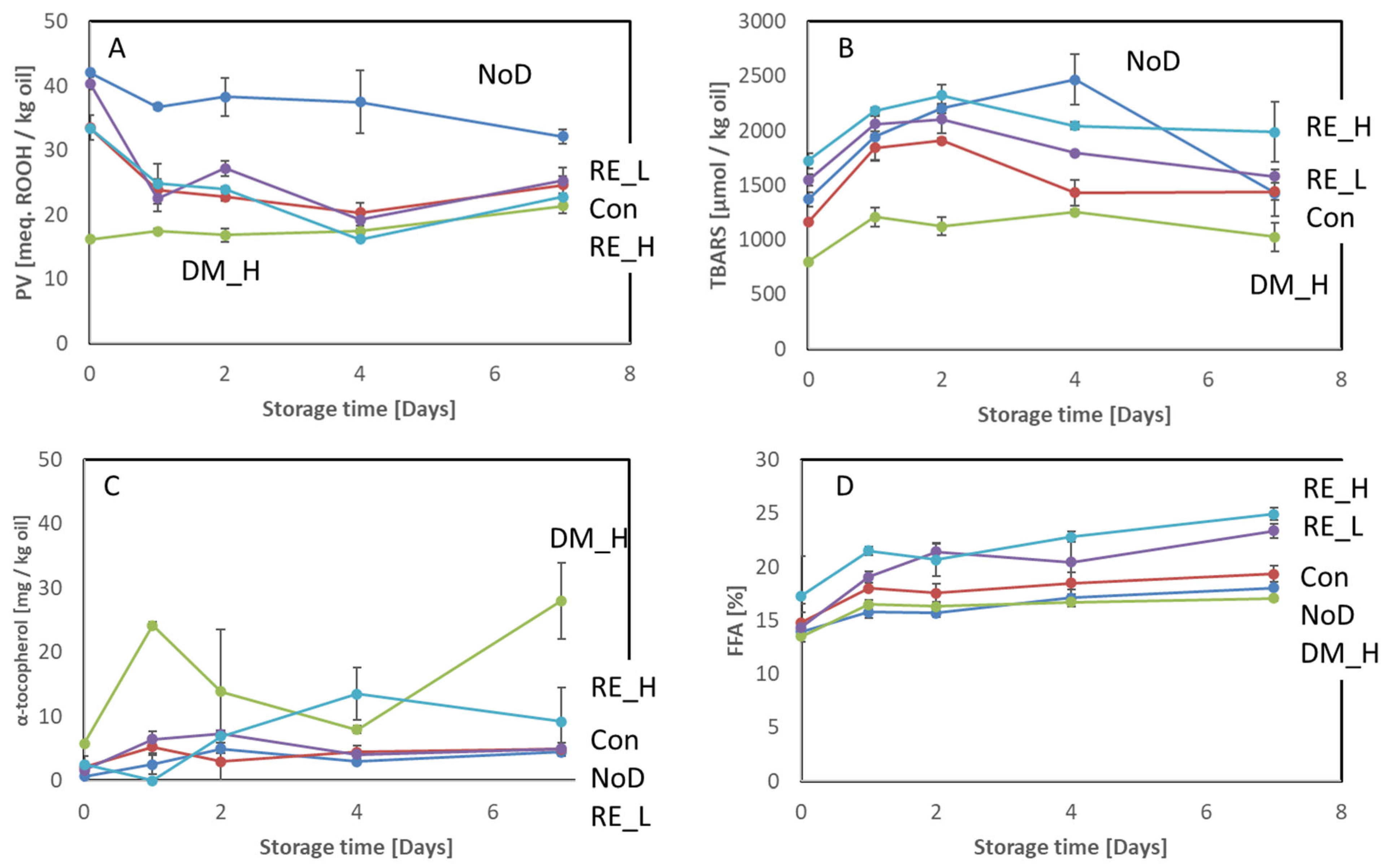
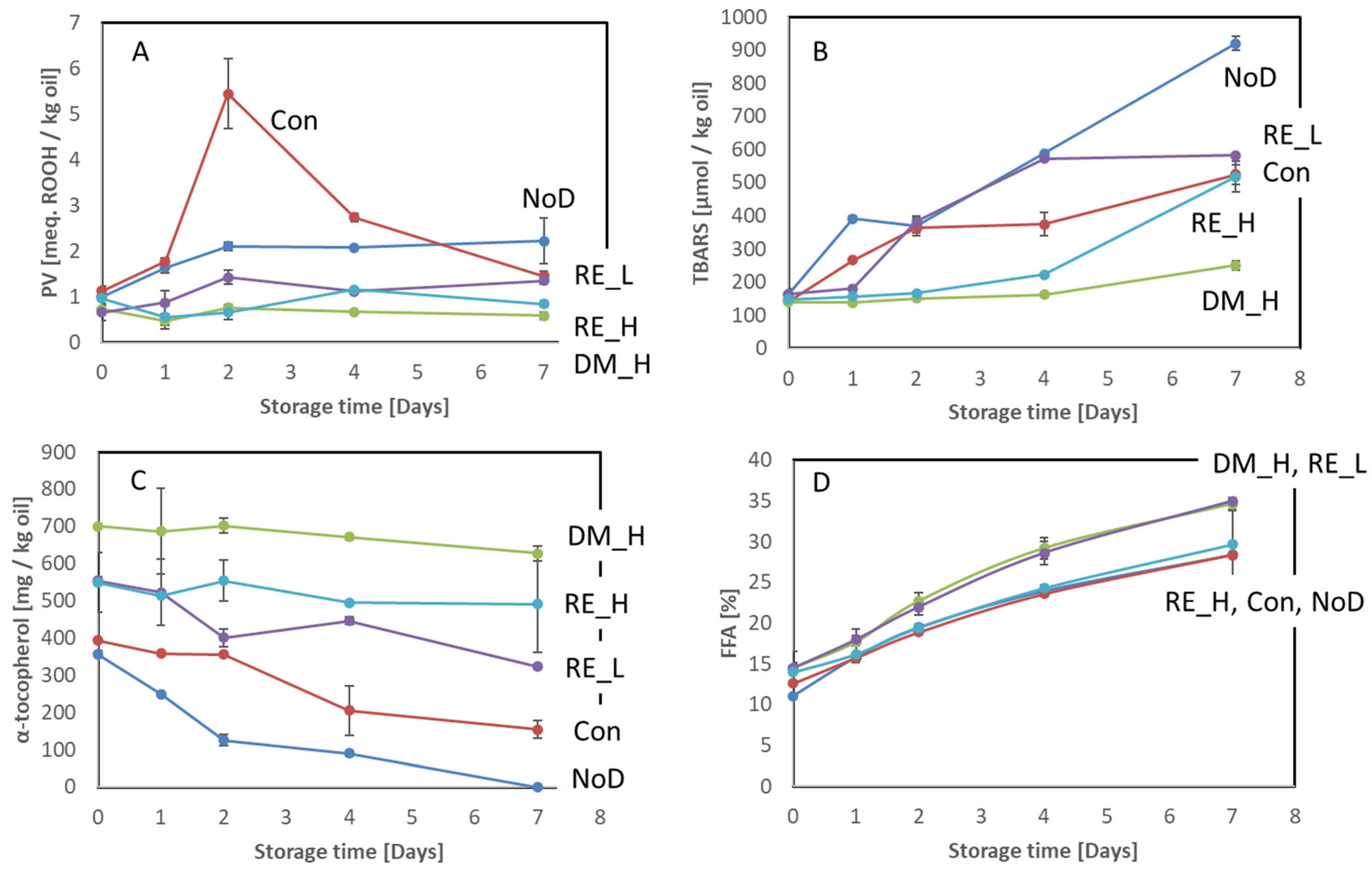
2.3. Effect of the Antioxidant Dipping on Lipid Oxidation
2.3.1. Backbone
2.3.2. Head
2.3.3. Viscera
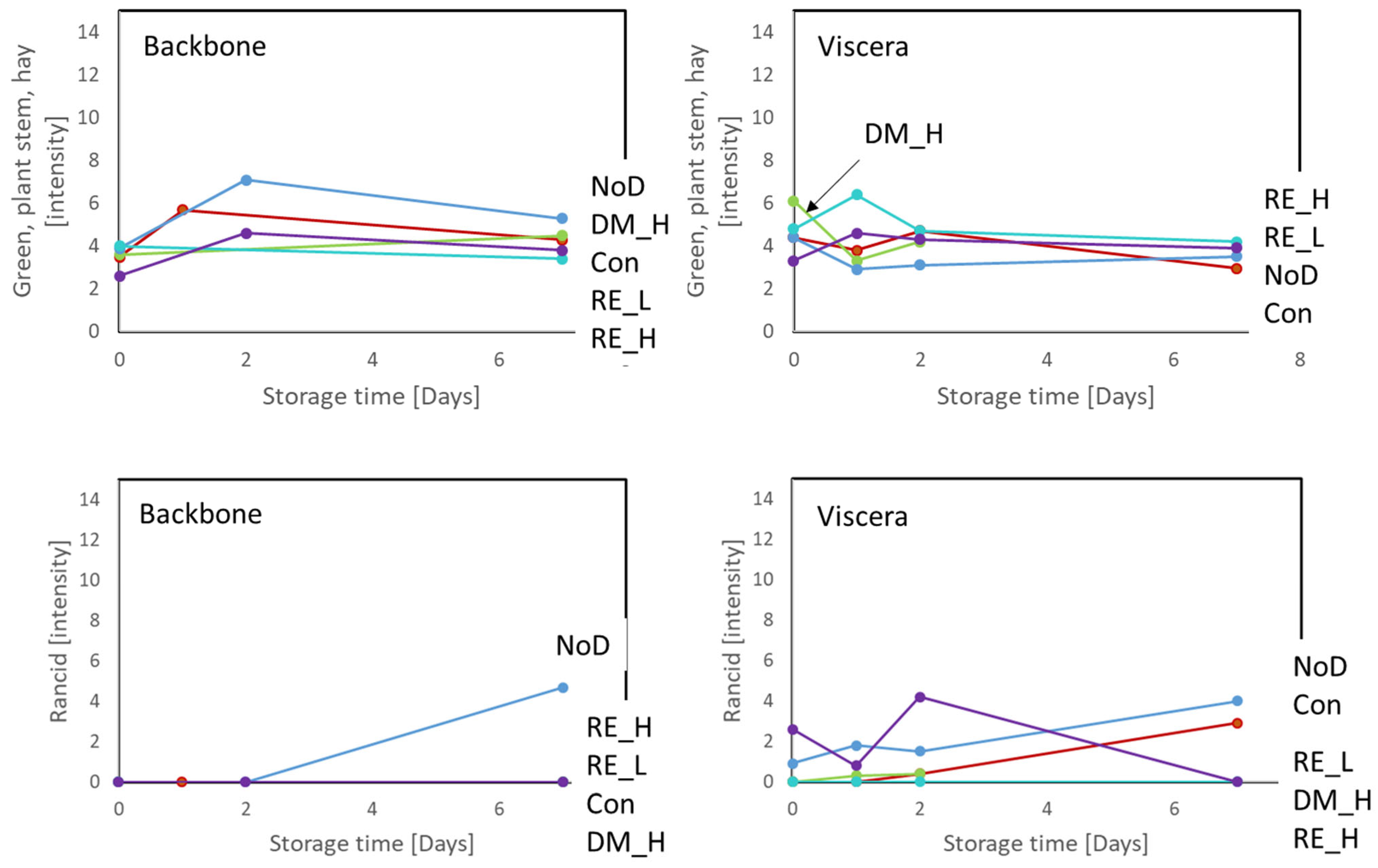
2.4. Sensory Evaluation of the Antioxidant Preserved Cod Side-Streams
3. Discussion
3.1. Effect of the Antioxidant Preservation Measured on Oxidative Deterioration
3.2. Effect of Storage Temperature on the Oxidative Stability of the Antioxidant Treated Cod Side-Streams
4. Materials and Methods
4.1. Materials
Cod Solid Side-Streams
4.2. Antioxidant Dipping
4.3. Storage Conditions and Sampling
4.4. Dry Matter
4.5. Oil Content
4.6. Peroxide Value (PV)
4.7. Tocopherols
4.8. Fatty Acid Methyl Esters (FAME)
4.9. Determination of Thiobarbituric Acid Reactive Substances (TBARS)
4.10. Free Fatty Acids (FFA)
4.11. Hemoglobin Content
4.12. Crude Protein Content
4.13. Sensory Evaluation
4.14. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. The State of World Fisheries and Aquaculture 2022. Towards Blue Transformation; FAO: Rome, Italy, 2022. [Google Scholar] [CrossRef]
- Jacobsen, C.; Warncke, S.A.; Hansen, S.H.; Sørensen, A.-D.M. Fish liver discards as a source of long chain omega-3 polyunsaturated fatty acids. Foods 2022, 11, 905. [Google Scholar] [CrossRef] [PubMed]
- Routray, W.; Dave, D.; Ramakrishnan, V.V.; Murphy, W. Study of drying kinetics of salmon processing by-products at different temperatures and the quality of extracted fish oil. Dry. Technol. 2017, 35, 1981–1993. [Google Scholar] [CrossRef]
- Abdollahi, M.; Rezaei, M.; Jafarpour, A.; Undeland, I. Sequential extraction of gel-forming proteins, collagen and collagen hydrolysate from gutted silver carp (Hypophthalmichthys molitrix), a biorefinery approach. Food Chem. 2018, 242, 568–578. [Google Scholar] [CrossRef] [PubMed]
- Abdollahi, M.; Undeland, I. Physicochemical and gel-forming properties of protein isolated from salmon, cod and herring by-products using the pH-shift method. LWT—Food Sci. Technol. 2019, 101, 678–684. [Google Scholar] [CrossRef]
- Remme, J.; Tveit, G.M.; Toldnes, B.; Slizute, R.; Carvajal, A.K. Production of protein hydrolysates from cod (Gadus morhua) heads: Lab and pilot scale studies. J. Aquat. Food Prod. Technol. 2022, 31, 114–127. [Google Scholar] [CrossRef]
- Remme, J.F.; Tveit, G.M.; Bondø, M.; Slizyte, R.; Ólafsdóttir, A.; Jónsdóttir, R.; Geirsdóttir, M.; Carvajal, A. Valorization of frozen cod (Gadus morhua) heads, captured by trawl and longline by the oceanic fleet, by enzymatic hydrolysis. J. Aquat. Food Prod. Technol. 2022, 31, 483–495. [Google Scholar] [CrossRef]
- Gildberg, A.; Arnesen, J.A.; Carlehög, M. Utilisation of cod backbone by biochemical fractionation. Process Biochem. 2002, 38, 475–480. [Google Scholar] [CrossRef]
- Jafarpour, A.; Gomes, R.M.; Gregersen, S.; Sloth, J.J.; Jacobsen, C.; Sørensen, A.-D.M. Characterization of Cod (Gadus morhua) frame composition and its valorization by enzymatic hydrolysis. J. Food Compos. Anal. 2020, 89, 103469. [Google Scholar] [CrossRef]
- Aspevik, T.; Totland, C.; Lea, P.; Oterhals, Å. Sensory and surface-active properties of protein hyrolysates based on Atlantic salmon (Salmo salar) by-products. Process Biochem. 2016, 51, 1006–1014. [Google Scholar] [CrossRef]
- Šližytė, R.; Mozuraitytė, R.; Martínez-Alvarez, O.; Falch, E.; Foucheereau-Peron, M.; Rustad, T. Functional, bioactive and antioxidative properties of hydrolysates obtained from cod (Gadus morhua) backbones. Process Biochem. 2009, 44, 668–677. [Google Scholar] [CrossRef]
- Bower, C.K.; Hietala, K.A. Acidification methods for stabilization and storage of salmon by-products. J. Aquat. Food Prod. 2008, 17, 459–478. [Google Scholar] [CrossRef]
- Wu, H.; Ghimani, S.; Undeland, I. Stabilization of herring (Clupea harengus) by-products against lipid oxidation by rinsing and incubation with antioxidant solution. Food Chem. 2020, 316, 126337. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Sajib, M.; Undeland, I. Controlling hemeglobin-mediated lipid oxidation in herring (Clupea harengus) co-products via incubation or dipping in a recyclable antioxidant solution. Food Control 2021, 125, 107963. [Google Scholar] [CrossRef]
- Wu, H.; Forghani, B.; Abdollahi, M.; Undeland, I. Lipid oxidation in sorted herring (Clupea harengus) filleting co-products from two seasons and its relationship to composition. Food Chem. 2022, 373, 131523. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Forghani, B.; Sajib, M.; Undeland, I. A recyclable dipping strategy to stabilize herring (Clupea harengus) co-products during ice storage. Food Bioprocess Technol. 2021, 14, 2207–2218. [Google Scholar] [CrossRef]
- Wu, H.; Xiao, S.; Yin, J.; Zhang, J.; Richards, M.P. Mechanisms involved in the inhibitory effects of free fatty acids on lipid peroxidation in turkey muscle. Food Chem. 2021, 342, 128333. [Google Scholar] [CrossRef]
- Tatiyaborworntham, N.; Oz, F.; Richards, M.P.; Wu, H. Paradoxical effects of lipolysis on the lipid oxidation in meat and meat products. Food Chem. X 2022, 14, 100317. [Google Scholar] [CrossRef] [PubMed]
- Labuza, T.P.; Dugan, L.R., Jr. Kinetics of lipid oxidation in foods. Crit. Rev. Food Sci. Nutr. 1971, 2, 355–405. [Google Scholar] [CrossRef]
- Shewfelt, R.L. Fish muscle lipolysis—A review. J. Food Biochem. 1981, 5, 79–100. [Google Scholar] [CrossRef]
- Hanaoka, K.; Toyomizu, M. The effects of dehydration and concentration of components on phospholipid decomposition in frozen fish muscle. Sci. Bull. Fac. Agric.-Kyushu Univ. Jpn. 1979, 33, 225–229. [Google Scholar]
- Wu, H.; Abdollahi, M.; Undeland, I. Effect of recovery technique, antioxidant addition and compositional features on lipid oxidation in protein enriched products from cod- salmon and herring backbones. Food Chem. 2021, 360, 129973. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Axelsson, J.; Kuhlin, M.; Fristedt, R.; Undeland, I. Pilot-Scale Antioxidant Dipping of Herring (Clupea harengus) Co-products to Allow Their Upgrading to a High-Quality Mince for Food Production. ACS Sustain. Chem. Eng. 2023, 11, 4727–4737. [Google Scholar] [CrossRef] [PubMed]
- Sajib, M.; Wu, H.; Fristedt, R.; Undeland, I. Hemoglobin-mediated lipid oxidation of herring filleting co-products during ensilaging and its inhibition by pre-incubation in antioxidant solutions. Sci. Rep. 2021, 11, 19492. [Google Scholar] [CrossRef] [PubMed]
- Jacobsen, C.; Let, M.B.; Sørensen, A.-D.M.; Horn, A.F.; Timm-Heinrich, M.; Nielsen, N.S. Applications of natural antioxidants in omega-3 enriched foods. Electron. J. Environ. Agric. Food Chem. 2008, 7, 3288–3295. [Google Scholar]
- Wu, H.; Richards, M.P.; Undeland, I. Lipid oxidation and antioxidant delivery systems in muscle food. Com. Rev. Food Sci. Food Saf. 2022, 21, 1275–1299. [Google Scholar] [CrossRef] [PubMed]
- Hraš, A.R.; Hadolin, M.; Knez, Ž.; Bauman, D. Comparison of antioxidative and synergistic effects of rosemary extract with α-tocopherol, ascorbyl palmitate and citric acid in sunflower oil. Food Chem. 2000, 71, 229–233. [Google Scholar] [CrossRef]
- Iglesias, J.; Pazos, E.; Andersen, M.L.; Skibsted, L.H.; Medina, I. Caffeic acid as antioxidant in fish muscle: Mechanism of synergism with endogenous ascorbic acid and α-tocopherol. J. Agric. Food Chem. 2009, 57, 675–681. [Google Scholar] [CrossRef]
- Wada, S.; Fang, X. The synergistic antioxidant effect of rosemary extract and α-tocopherol in sardine oil model system and frozen-crushed fish meat. J. Food Process. Preserv. 1992, 16, 263–274. [Google Scholar] [CrossRef]
- Xie, J.; VanAlstyne, P.; Uhlir, A.; Yang, X. A review on rosemary as natural antioxidant solution. Eur. J. Lipid Sci. Technol. 2017, 119, 1600439. [Google Scholar] [CrossRef]
- Larson, K.J.; Undeland, I. Effect of caffeic acid on haemoglobin-mediated lipid and protein oxidation in washed cod mince during ice and frozen storage. J. Sci. Agric. 2010, 90, 2531–2540. [Google Scholar] [CrossRef]
- Karlsdottir, M.; Arason, S.; Thorarindsdottir, K.; Nguyen, M.V.; Kristinsson, H. Lipid degradation of cod liver during frozen storage influenced by temperature, packaging method, and seasonal variation. J. Aquat. Food Prod. Technol. 2016, 25, 802–810. [Google Scholar] [CrossRef]
- Shumilina, E.; Slizyte, R.; Mozuraityte, R.; Dykyy, A.; Stein, T.A.; Dikiy, A. Quality changes of salmon by-products during storage: Assessment and quantification by NMR. Food Chem. 2016, 211, 803–811. [Google Scholar] [CrossRef] [PubMed]
- Sveinsdóttir, H.I.; Karlsdóttir, M.G.; Arason, S.; Stefánsson, G.; Sone, I.; Skåra, T.; Rustad, T.; Larsson, K.; Undeland, I.; Gudjónsdóttir, M. Effect of antioxidants on the sensory quality and physicochemical stability of Atlantic mackerel (Scomber scombrus) fillets during frozen storage. Food Chem. 2020, 321, 126744. [Google Scholar] [CrossRef] [PubMed]
- Ghirmai, S.; Eriksson, L.; Wu, H.; Axelsson, M.; Undeland, I. Improving the stability of red blood cells in Rainbow Trout (Oncorhynchus mykiss) and Herring (Clupea harengus): Potential solutions for post-mortem fish handling to minimize lipid oxidation. Food Bioprocess Technol. 2020, 13, 1344–1355. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Iverson, S.J.; Lang, S.L.C.; Cooper, M.H. Comparison of the Bligh and Dyer and Folch methods for total lipid determination in a broad range of marine tissue. Lipids 2001, 36, 1283–1287. [Google Scholar] [CrossRef] [PubMed]
- Shantha, N.C.; Decker, E.A. Rapid, sensitive, iron-based spectrophotometric methods for determination of peroxide values of food lipids. J. AOAC Int. 1994, 77, 421–424. [Google Scholar] [CrossRef]
- AOCS. AOCS Official Method Ce 8-89. In Official Methods and Recommended Practice of the AOCS; AOCS: Champaign, IL, USA, 1997. [Google Scholar]
- Safafar, H.; Hass, M.Z.; Møller, P.; Holdt, S.L.; Jacobsen, C. High-EPA biomass from Nannochloropsis salina cultivated in a flat-panel photo-bioreactor on a process water-enriched growth medium. Mar. Drugs 2016, 14, 144. [Google Scholar] [CrossRef]
- AOCS. AOCS Official Method Ce 1b-89. In Official Methods and Recommended Practice of the AOCS; AOCS: Champaign, IL, USA, 1998. [Google Scholar]
- Hornsey, H.C. The colour of cooked cured pork. I.—Estimation of the Nitric oxide-Haem Pigments. J. Sci. Food Agric. 1956, 7, 534–540. [Google Scholar] [CrossRef]
- Mariotti, M.; Tomé, D.; Mirand, P.P. Converting nitrogen into protein—Beyond 6.25 and Jones’ factors. Crit. Rev. Food Sci. Nutr. 2008, 48, 177–184. [Google Scholar] [CrossRef]
- ISO 5496:2006; Sensory Analysis—Methodology—Initiation and Training of Assessors in the Detection and Recognition of Odours. International Organization for Standardization: Geneva, Switzerland; 2006.
- ISO 8586:2012; Sensory Analysis—General Guidance for the Selection, Training and Monitoring of Selected Assessors and Expert Sensory Assessors. International Organization for Standardization: Geneva, Switzerland; 2012.
- ISO 8589:2007/Amd.1:2014; Sensory Analysis—General Guidance for the Design of Test Rooms. International Organization for Standardization: Geneva, Switzerland; 2014.
- NMKL. General Guidelines for Quality Assuring Sensory Laboratories; NMKL Procedure No. 6; NMKL: Bergen, Norway, 2023. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compositional Parameter | Backbone | Head | Viscera |
|---|---|---|---|
| Dry matter (%) | 23.2 ± 1.8 a | 23.4 ± 1.6 a | 34.9 ± 5.3 b |
| Oil (% ww) | 1.2 ± 0.1 a | 5.6 ± 0.5 a | 22.5 ± 5.1 b |
| EPA (% of total fatty acids) | 15.2 ± 1.2 b | 10.5 ± 1.4 a | 9.4 ± 0.9 a |
| DHA (% of total fatty acids | 20.8 ± 0.7 b | 11.0 ± 1.2 a | 11.9 ± 0.8 a |
| Protein (% ww) | 15.3 ± 0.7 c | 13.6 ± 0.6 b | 10.3 ± 0.7 a |
| Treatments | Codes | Backbone (B) | Head (H) | Viscera (V) |
|---|---|---|---|---|
| No dipping | NoD | B_NoD | H_NoD | V_NoD |
| 0.9% NaCl | Con | B_Con | H_Con | V_Con |
| 0.9% NaCl and 2% Duralox MANC-213 | DM_H | B_DM_H | H_DM_H | V_DM_H |
| 0.9% NaCl and 0.05% rosemary extract | RE_L | B_RE_L | H_RE_L | V_RE_L |
| 0.9% NaCl and 0.2% rosemary extract | RE_H | B_RE_H | H_RE_H | V_RE_H |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sørensen, A.-D.M.; Wu, H.; Hyldig, G.; Bøknæs, N.; Mejlholm, O.; Undeland, I.; Jacobsen, C. Oxidative Stability of Side-Streams from Cod Filleting—Effect of Antioxidant Dipping and Low-Temperature Storage. Mar. Drugs 2023, 21, 591. https://doi.org/10.3390/md21110591
Sørensen A-DM, Wu H, Hyldig G, Bøknæs N, Mejlholm O, Undeland I, Jacobsen C. Oxidative Stability of Side-Streams from Cod Filleting—Effect of Antioxidant Dipping and Low-Temperature Storage. Marine Drugs. 2023; 21(11):591. https://doi.org/10.3390/md21110591
Chicago/Turabian StyleSørensen, Ann-Dorit Moltke, Haizhou Wu, Grethe Hyldig, Niels Bøknæs, Ole Mejlholm, Ingrid Undeland, and Charlotte Jacobsen. 2023. "Oxidative Stability of Side-Streams from Cod Filleting—Effect of Antioxidant Dipping and Low-Temperature Storage" Marine Drugs 21, no. 11: 591. https://doi.org/10.3390/md21110591