
The Effects of Enzymes, Species, and Storage of Raw Material on Physicochemical Properties of Protein Hydrolysates from Whitefish Heads
 ,
,
Abstract
:1. Introduction
2. Results and Discussion
2.1. Raw Material
2.2. Dry Matter
2.3. Protein Yield
2.4. Chemical Composition of Dry Powder Hydrolysates
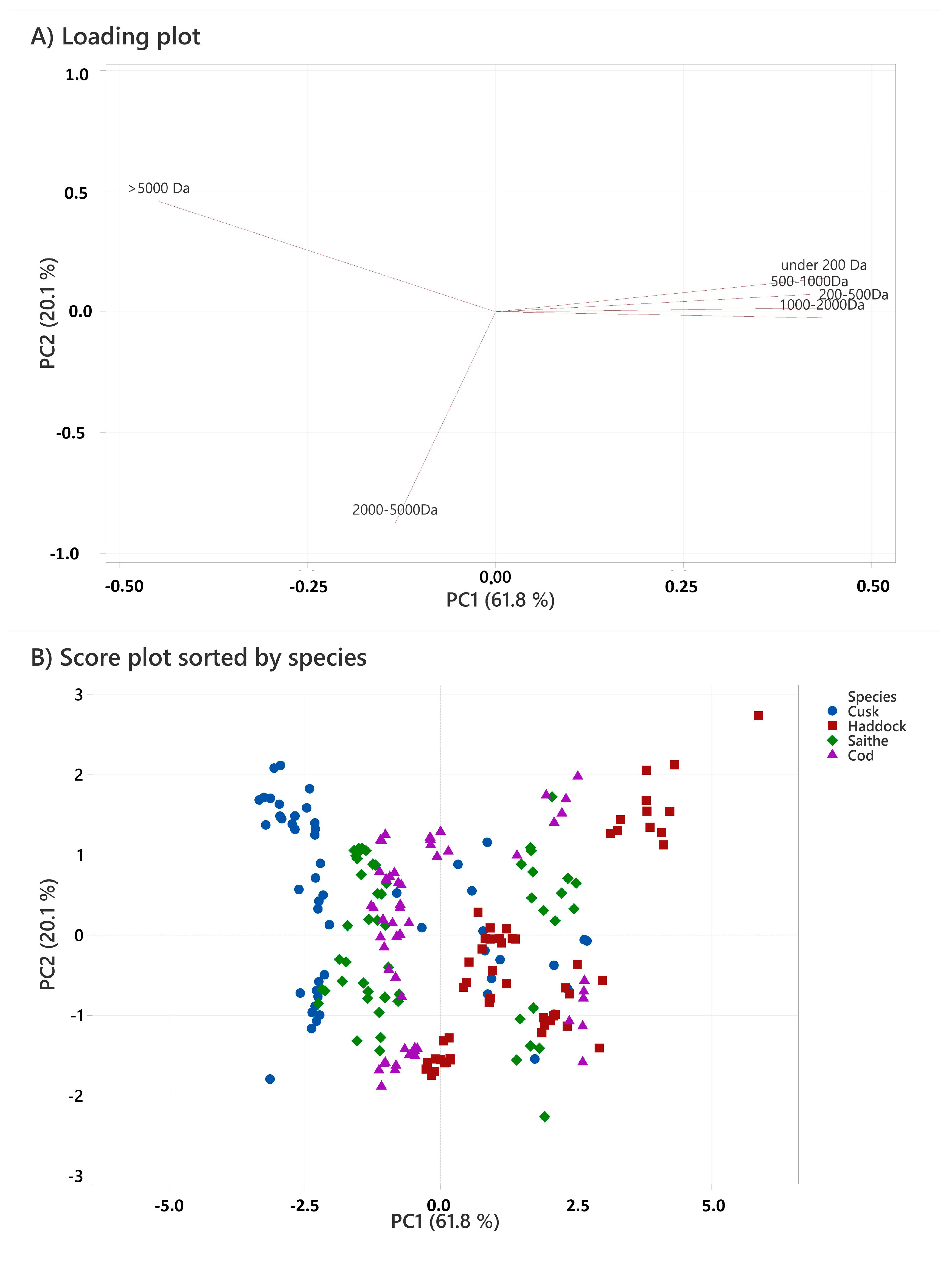
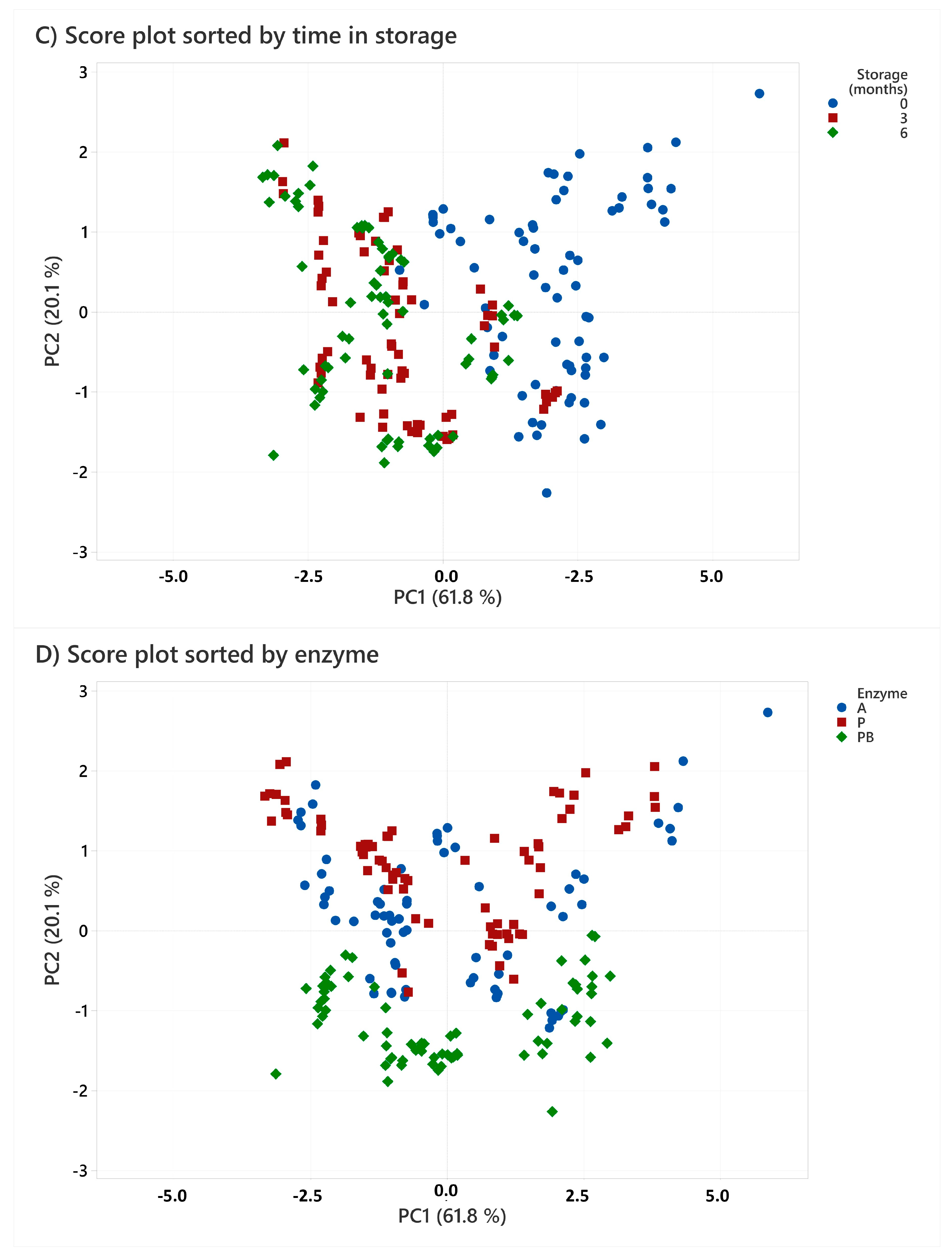
2.5. Molecular Weight Distribution
2.6. Protein Solubility of FPH
3. Materials and Methods
3.1. Raw Material
3.2. Chemicals and Enzymes
3.3. Enzymatic Hydrolysis
3.4. Chemical Characterisation
3.5. Molecular Weight Distribution
3.6. Protein Solubility
3.7. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gao, R.; Yu, Q.; Shen, Y.; Chu, Q.; Chen, G.; Fen, S.; Yang, M.; Yuan, L.; McClements, D.J.; Sun, Q. Production, Bioactive Properties, and Potential Applications of Fish Protein Hydrolysates: Developments and Challenges. Trends Food Sci. Technol. 2021, 110, 687–699. [Google Scholar] [CrossRef]
- Myhre, M.S.; Richardsen, R.; Nystøyl, R.; Strandheim, G. Analyse Marint Restråstoff 2021; SINTEF Ocean AS: New York, NY, USA, 2022; ISBN 978-82-14-07562-5. [Google Scholar]
- Myhre, M.S.; Skavang, P.K.; Remme, J.; Wolff, R.; Carvajal, A. Mapping of Marine Rest Raw Material in the Norwegian Seafood Industry: A Decade of Results. J. Aquat. Food Prod. Technol. 2023, 32, 554–569. [Google Scholar] [CrossRef]
- Fiskeridirektoratet. Fangst Fordelt på Art. Available online: https://www.fiskeridir.no/Yrkesfiske/Tall-og-analyse/Fangst-og-kvoter/Fangst/Fangst-fordelt-paa-art (accessed on 24 October 2022).
- Hjellnes, V.; Rustad, T.; Falch, E. The Value Chain of the White Fish Industry in Norway: History, Current Status and Possibilities for Improvement—A Review. Reg. Stud. Mar. Sci. 2020, 36, 101293. [Google Scholar] [CrossRef]
- Kostveit, T. Jubler over halvert toll på tørrfiskhoder i Nigeria. Available online: https://seafood.no/aktuelt/nyheter/jubler-over-halvert-toll-pa-torrfiskhoder-i-nigeria/ (accessed on 26 October 2022).
- Remme, J.F.; Tveit, G.M.; Bondø, M.; Slizyte, R.; Ólafsdóttir, A.; Jónsdóttir, R.; Geirsdóttir, M.; Carvajal, A.K. Valorisation of Frozen Cod (Gadus morhua) Heads, Captured by Trawl and Longline by the Oceanic Fleet, by Enzymatic Hydrolysis. J. Aquat. Food Prod. Technol. 2022, 31, 483–495. [Google Scholar] [CrossRef]
- Remme, J.; Tveit, G.M.; Toldnes, B.; Slizyte, R.; Carvajal, A.K. Production of Protein Hydrolysates from Cod (Gadus morhua) Heads: Lab and Pilot Scale Studies. J. Aquat. Food Prod. Technol. 2022, 31, 114–127. [Google Scholar] [CrossRef]
- Islam, M.d.S.; Hongxin, W.; Admassu, H.; Noman, A.; Ma, C.; An Wei, F. Degree of Hydrolysis, Functional and Antioxidant Properties of Protein Hydrolysates from Grass Turtle (Chinemys reevesii) as Influenced by Enzymatic Hydrolysis Conditions. Food Sci. Nutr. 2021, 9, 4031–4047. [Google Scholar] [CrossRef]
- Slizyte, R.; Rommi, K.; Mozuraityte, R.; Eck, P.; Five, K.; Rustad, T. Bioactivities of Fish Protein Hydrolysates from Defatted Salmon Backbones. Biotechnol. Rep. 2016, 11, 99–109. [Google Scholar] [CrossRef]
- Siddiqui, S.A.; Schulte, H.; Pleissner, D.; Schönfelder, S.; Kvangarsnes, K.; Dauksas, E.; Rustad, T.; Cropotova, J.; Heinz, V.; Smetana, S. Transformation of Seafood Side-Streams and Residuals into Valuable Products. Foods 2023, 12, 422. [Google Scholar] [CrossRef]
- Badii, F.; Howell, N.K. A Comparison of Biochemical Changes in Cod (Gadus morhua) and Haddock (Melanogrammus aeglefinus) Fillets during Frozen Storage. J. Sci. Food Agric. 2002, 82, 87–97. [Google Scholar] [CrossRef]
- Huidobro, A.; Mohamed, G.F.; Tejada, M. Aggregation of Myofibrillar Proteins in Hake, Sardine, and Mixed Minces during Frozen Storage. J. Agric. Food Chem. 1998, 46, 2601–2608. [Google Scholar] [CrossRef]
- Šližyte, R.; Daukšas, E.; Falch, E.; Storrø, I.; Rustad, T. Yield and Composition of Different Fractions Obtained after Enzymatic Hydrolysis of Cod (Gadus morhua) by-Products. Process Biochem. 2005, 40, 1415–1424. [Google Scholar] [CrossRef]
- Tveit, G.M.; Carvajal, A.K.; Slizyte, R.; Meldstad, F.; Nordtvedt, T.S.; Remme, J.; Rustad, T. Enzymatic Hydrolysis of Cod Heads—Effect of Freezing and Thawing on the Quality and Composition of Protein Hydrolysates. Available online: https://iifiir.org/fr/fridoc/hydrolyse-enzymatique-des-tetes-de-morue-amp-nbsp-effets-de-la-142439 (accessed on 25 November 2022).
- Økland, H.M.W.; Kjerstad, M. Potensiale for Ilandføring Og Utnyttelse Av Fiskehoder. Møreforsking AS, Å2013. 2002. Available online: https://www.moreforsk.no/publikasjoner/rapporter/marin/potensiale-for-ilandforing-og-utnyttelse-av-fiskehoder/1074/435/ (accessed on 18 July 2023).
- Falch, E.; Rustad, T.; Aursand, M. By-Products from Gadiform Species as Raw Material for Production of Marine Lipids as Ingredients in Food or Feed. Process Biochem. 2006, 41, 666–674. [Google Scholar] [CrossRef]
- Spinelli, J.; Koury, B.; Miller, R. Approaches to the utilization of fish for the preparation of protein isolates. Enzymic Modifications of Myofibrillar Fish Proteins. J. Food Sci. 1972, 37, 604–608. [Google Scholar] [CrossRef]
- Kristinsson, H.G.; Rasco, B.A. Fish Protein Hydrolysates: Production, Biochemical, and Functional Properties. Crit. Rev. Food Sci. Nutr. 2000, 40, 43–81. [Google Scholar] [CrossRef]
- Lin, C.-C.; Liang, J.-H. Effect of Antioxidants on the Oxidative Stability of Chicken Breast Meat in a Dispersion System. J. Food Sci. 2002, 67, 530–533. [Google Scholar] [CrossRef]
- He, S.; Franco, C.; Zhang, W. Functions, Applications and Production of Protein Hydrolysates from Fish Processing Co-Products (FPCP). Food Res. Int. 2013, 50, 289–297. [Google Scholar] [CrossRef]
- Sandbakken, I.S.; Five, K.K.; Bardal, T.; Knapp, J.L.; Olsen, R.E. Salmon Hydrolysate as a Protein Source for Atlantic Salmon; Prion Content and Effects on Growth, Digestibility and Gut Health. Aquaculture 2023, 576, 739863. [Google Scholar] [CrossRef]
- Gbogouri, G.A.; Linder, M.; Fanni, J.; Parmentier, M. Influence of Hydrolysis Degree on the Functional Properties of Salmon Byproducts Hydrolysates. J. Food Sci. 2004, 69, C615–C622. [Google Scholar] [CrossRef]
- Volkov, V.; Mezenova, O.; Moersel, J.-T.; Kuehn, S.; Grimm, T.; Höhling, A.; Barabanov, S.; Volkov, K. Hydrolysis Products from Sockeye (Oncorhynchus nerka L.) Heads from the Kamchatka Peninsula Produced by Different Methods: Biological Value. Foods Raw Mater. 2020, 9, 10–18. [Google Scholar] [CrossRef]
- Kvangarsnes, K.; Kendler, S.; Rustad, T.; Aas, G.H. Induced Oxidation and Addition of Antioxidant before Enzymatic Hydrolysisof Heads of Rainbow Trout (Oncorhynchus mykiss)–Effect on the Resulting Oiland Protein Fraction. Heliyon 2021, 7, e06816. [Google Scholar] [CrossRef]
- Abdul-Hamid, A.; Bakar, J.; Bee, G.H. Nutritional Quality of Spray Dried Protein Hydrolysate from Black Tilapia (Oreochromis mossambicus). Food Chem. 2002, 78, 69–74. [Google Scholar] [CrossRef]
- Sathivel, S.; Smiley, S.; Prinyawiwatkul, W.; Bechtel, P.J. Functional and Nutritional Properties of Red Salmon (Oncorhynchus nerka) Enzymatic Hydrolysates. J. Food Sci. 2005, 70, c401–c406. [Google Scholar] [CrossRef]
- Jafarpour, A.; Gomes, R.M.; Gregersen, S.; Sloth, J.J.; Jacobsen, C.; Moltke Sørensen, A.-D. Characterization of Cod (Gadus morhua) Frame Composition and Its Valorization by Enzymatic Hydrolysis. J. Food Compos. Anal. 2020, 89, 103469. [Google Scholar] [CrossRef]
- Picot, L.; Ravallec, R.; Fouchereau-Péron, M.; Vandanjon, L.; Jaouen, P.; Chaplain-Derouiniot, M.; Guérard, F.; Chabeaud, A.; LeGal, Y.; Alvarez, O.M.; et al. Impact of Ultrafiltration and Nanofiltration of an Industrial Fish Protein Hydrolysate on Its Bioactive Properties. J. Sci. Food Agric. 2010, 90, 1819–1826. [Google Scholar] [CrossRef]
- Wijers, M.C.; Pouliot, Y.; Gauthier, S.F.; Pouliot, M.; Nadeau, L. Use of Nanofiltration Membranes for the Desalting of Peptide Fractions from Whey Protein Enzymatic Hydrolysates. Le Lait 1998, 78, 621–632. [Google Scholar] [CrossRef]
- Kim, H.-O.; Li-Chan, E.C.Y. Quantitative Structure-Activity Relationship Study of Bitter Peptides. J. Agric. Food Chem. 2006, 54, 10102–10111. [Google Scholar] [CrossRef]
- Aspevik, T.; Thoresen, L.; Steinsholm, S.; Carlehög, M.; Kousoulaki, K. Sensory and Chemical Properties of Protein Hydrolysates Based on Mackerel (Scomber scombrus) and Salmon (Salmo salar) Side Stream Materials. J. Aquat. Food Prod. Technol. 2021, 30, 176–187. [Google Scholar] [CrossRef]
- Aspevik, T.; Totland, C.; Lea, P.; Oterhals, Å. Sensory and Surface-Active Properties of Protein Hydrolysates Based on Atlantic Salmon (Salmo salar) by-Products. Process Biochem. 2016, 51, 1006–1014. [Google Scholar] [CrossRef]
- Dauksas, E.; Slizyte, R.; Rustad, T.; Storro, I. Bitterness in Fish Protein Hydrolysates and Methods for Removal. J. Aquat. Food Prod. Technol. 2004, 13, 101–114. [Google Scholar] [CrossRef]
- Šližytė, R.; Carvajal, A.K.; Mozuraityte, R.; Aursand, M.; Storrø, I. Nutritionally Rich Marine Proteins from Fresh Herring By-Products for Human Consumption. Process Biochem. 2014, 49, 1205–1215. [Google Scholar] [CrossRef]
- Kim, H.-O.; Li-Chan, E.C.Y. Application of Fourier Transform Raman Spectroscopy for Prediction of Bitterness of Peptides. Appl. Spectrosc. 2006, 60, 1297–1306. [Google Scholar] [CrossRef] [PubMed]
- Leni, G.; Soetemans, L.; Caligiani, A.; Sforza, S.; Bastiaens, L. Degree of Hydrolysis Affects the Techno-Functional Properties of Lesser Mealworm Protein Hydrolysates. Foods 2020, 9, 381. [Google Scholar] [CrossRef] [PubMed]
- Baharuddin, N.A.; Halim, N.R.A.; Sarbon, N.M. Effect of Degree of Hydrolysis (DH) on the Functional Properties and Angiotensin I-Converting Enzyme (ACE) Inhibitory Activity of Eel (Monopterus sp.) Protein Hydrolysate. Int. Food Res. J. 2016, 23, 1424–1431. [Google Scholar]
- Abachi, S.; Pilon, G.; Marette, A.; Bazinet, L.; Beaulieu, L. Beneficial Effects of Fish and Fish Peptides on Main Metabolic Syndrome Associated Risk Factors: Diabetes, Obesity and Lipemia. Crit. Rev. Food Sci. Nutr. 2022, 63, 7896–7944. [Google Scholar] [CrossRef]
- Durand, R.; Fraboulet, E.; Marette, A.; Bazinet, L. Simultaneous Double Cationic and Anionic Molecule Separation from Herring Milt Hydrolysate and Impact on Resulting Fraction Bioactivities. Sep. Purif. Technol. 2019, 210, 431–441. [Google Scholar] [CrossRef]
- Geirsdottir, M.; Hlynsdottir, H.; Thorkelsson, G.; Sigurgisladottir, S. Solubility and Viscosity of Herring (Clupea harengus) Proteins as Affected by Freezing and Frozen Storage. J. Food Sci. 2007, 72, C376–C380. [Google Scholar] [CrossRef]
- Tamura, M.; Mori, N.; Miyoshi, T.; Koyama, S.; Kohri, H.; Okai, H. Practical Debittering Using Model Peptides and Related Compounds. Agric. Biol. Chem. 1990, 54, 41–51. [Google Scholar] [CrossRef]
- Alizadeh, E.; Chapleau, N.; de Lamballerie, M.; Le-Bail, A. Effect of Different Freezing Processes on the Microstructure of Atlantic Salmon (Salmo salar) Fillets. Innov. Food Sci. Emerg. Technol. 2007, 8, 493–499. [Google Scholar] [CrossRef]
- Standal, I.B.; Mozuraityte, R.; Rustad, T.; Alinasabhematabadi, L.; Carlsson, N.-G.; Undeland, I. Quality of Filleted Atlantic Mackerel (Scomber scombrus) During Chilled and Frozen Storage: Changes in Lipids, Vitamin D, Proteins, and Small Metabolites, Including Biogenic Amines. J. Aquat. Food Prod. Technol. 2018, 27, 338–357. [Google Scholar] [CrossRef]
- Ovissipour, M.; Benjakul, S.; Safari, R.; Motamedzadegan, A. Fish protein hydrolysates production from yellowfin tuna Thunnus albacares head using Alcalase and Protamex. Int. Aquat. Res. 2010, 2, 87–95. [Google Scholar]
- Mariotti, F.; Tomé, D.; Mirand, P.P. Converting Nitrogen into Protein--beyond 6.25 and Jones’ Factors. Crit. Rev. Food Sci. Nutr. 2008, 48, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Bligh, E.G.; Dyer, W.J. A Rapid Method of Total Lipid Extraction and Purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein Measurement with the Folin Phenol Reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Species | Storage | Enzyme | Dry Matter | Protein Yield | ||||
|---|---|---|---|---|---|---|---|---|
| (Months) | (Type) | (% of Wet Weight) | (g/100 g Raw Material) | |||||
| Cod | 0 | A | 5.19 | ± | 0.02 d | 6.61 | ± | 0.10 d |
| P | 5.79 | ± | 0.02 bc | 7.15 | ± | 0.09 bcd | ||
| PB | 6.27 | ± | 0.03 a | 8.03 | ± | 0.20 a | ||
| 3 | A | 5.60 | ± | 0.01 c | 7.49 | ± | 0.00 abc | |
| P | 5.62 | ± | 0.04 c | 7.24 | ± | 0.09 abcd | ||
| PB | 5.93 | ± | 0.09 b | 7.82 | ± | 0.29 ab | ||
| 6 | A | 5.80 | ± | 0.00 bc | 7.28 | ± | 0.01 abcd | |
| P | 5.65 | ± | 0.03 c | 6.56 | ± | 0.12 d | ||
| PB | 5.96 | ± | 0.01 b | 6.92 | ± | 0.12 cd | ||
| Cusk | 0 | A | 6.58 | ± | 0.07 a | 8.26 | ± | 0.03 bc |
| P | 6.39 | ± | 0.18 a | 7.86 | ± | 0.21 c | ||
| PB | 6.54 | ± | 0.14 a | 8.01 | ± | 0.50 bc | ||
| 3 | A | 6.58 | ± | 0.02 a | 9.66 | ± | 0.07 a | |
| P | 6.67 | ± | 0.01 a | 9.40 | ± | 0.03 a | ||
| PB | 6.67 | ± | 0.07 a | 9.37 | ± | 0.11 a | ||
| 6 | A | 6.34 | ± | 0.00 a | 8.98 | ± | 0.10 ab | |
| P | 6.34 | ± | 0.10 a | 8.77 | ± | 0.13 abc | ||
| PB | 6.57 | ± | 0.07 a | 8.75 | ± | 0.01 abc | ||
| Haddock | 0 | A | 5.30 | ± | 0.02 a | 6.79 | ± | 0.06 a |
| P | 5.35 | ± | 0.17 a | 6.70 | ± | 0.20 a | ||
| PB | 5.18 | ± | 0.02 a | 6.46 | ± | 0.06 a | ||
| 3 | A | 5.33 | ± | 0.04 a | 7.10 | ± | 0.05 a | |
| P | 4.93 | ± | 0.02 a | 6.40 | ± | 0.08 a | ||
| PB | 5.18 | ± | 0.06 a | 6.48 | ± | 0.32 a | ||
| 6 | A | 5.26 | ± | 0.04 a | 6.63 | ± | 0.02 a | |
| P | 5.08 | ± | 0.08 a | 6.30 | ± | 0.12 a | ||
| PB | 5.13 | ± | 0.11 a | 6.37 | ± | 0.26 a | ||
| Saithe | 0 | A | 5.65 | ± | 0.01 ab | 7.37 | ± | 0.01 a |
| P | 5.68 | ± | 0.06 ab | 7.19 | ± | 0.02 a | ||
| PB | 5.70 | ± | 0.06 ab | 7.10 | ± | 0.04 a | ||
| 3 | A | 5.88 | ± | 0.19 a | 7.58 | ± | 0.17 a | |
| P | 5.42 | ± | 0.04 b | 6.94 | ± | 0.21 a | ||
| PB | 5.81 | ± | 0.03 ab | 7.56 | ± | 0.30 a | ||
| 6 | A | 5.52 | ± | 0.02 ab | 7.18 | ± | 0.07 a | |
| P | 5.41 | ± | 0.02 b | 6.80 | ± | 0.09 a | ||
| PB | 5.55 | ± | 0.02 ab | 7.24 | ± | 0.06 a | ||
| Species | Storage (Months) | Enzyme (Type) | Dry Matter (%) | Ash (%) | Fat (%) | Protein (%) |
|---|---|---|---|---|---|---|
| Cod | 0 | PB | 96.80 ± 1.61 | 10.00 ± 0.06 | 0.50 ± 0.02 | 91.76 ± 0.89 |
| P | 98.21 ± 0.22 | 11.74 ± 0.12 | 0.40 ± 0.01 | 88.58 ± 0.21 | ||
| A | 97.63 ± 0.27 | 10.43 ± 0.39 | 0.32 ± 0.01 | 89.05 ± 0.07 | ||
| 3 | PB | 97.34 ± 0.12 | 11.24 ± 0.16 | 0.48 ± 0.04 | 91.78 ± 0.49 | |
| P | 96.99 ± 0.16 | 12.18 ± 0.24 | 0.48 ± 0.04 | 90.57 ± 0.22 | ||
| A | 96.67 ± 0.02 | 11.71 ± 0.01 | 0.35 ± 0.04 | 90.08 ± 0.24 | ||
| 6 | PB | 98.39 ± 0.03 | 11.86 ± 0.31 | 0.29 ± 0.02 | 80.74 ± 0.77 | |
| P | 98.11 ± 0.30 | 11.31 ± 0.20 | 0.47 ± 0.06 | 83.35 ± 0.37 | ||
| A | 97.73 ± 0.33 | 10.75 ± 0.17 | 0.32 ± 0.01 | 83.46 ± 0.11 | ||
| Cusk | 0 | PB | 98.06 ± 0.15 | 8.58 ± 0.25 | 0.55 ± 0.04 | 88.21 ± 1.37 |
| P | 98.12 ± 0.02 | 8.80 ± 0.13 | 1.42 ± 0.21 | 88.01 ± 0.40 | ||
| A | 98.36 ± 0.17 | 8.07 ± 0.24 | 0.90 ± 0.36 | 89.01 ± 0.81 | ||
| 3 | PB | 99.74 ± 0.06 | 8.96 ± 0.04 | 0.41 ± 0.03 | 97.62 ± 0.88 | |
| P | 99.63 ± 0.17 | 8.96 ± 0.05 | 1.17 ± 0.26 | 98.27 ± 0.14 | ||
| A | 99.57 ± 0.06 | 8.43 ± 0.12 | 0.45 ± 0.14 | 98.09 ± 0.51 | ||
| 6 | PB | 98.53 ± 0.11 | 8.18 ± 0.16 | 0.52 ± 0.06 | 94.57 ± 0.34 | |
| P | 98.78 ± 0.19 | 9.02 ± 0.32 | 0.80 ± 0.02 | 94.19 ± 0.16 | ||
| A | 98.51 ± 0.12 | 8.47 ± 0.05 | 0.62 ± 0.02 | 94.34 ± 0.28 | ||
| Haddock | 0 | PB | 98.05 ± 0.09 | 11.09 ± 0.08 | 0.41 ± 0.05 | 88.80 ± 0.18 |
| P | 98.54 ± 0.17 | 11.31 ± 0.25 | 0.40 ± 0.05 | 88.32 ± 0.56 | ||
| A | 98.63 ± 0.00 | 10.58 ± 0.01 | 0.34 ± 0.10 | 88.05 ± 0.19 | ||
| 3 | PB | 98.27 ± 0.03 | 13.52 ± 0.02 | 0.35 ± 0.02 | 86.46 ± 2.91 | |
| P | 99.24 ± 0.21 | 14.51 ± 0.09 | 0.52 ± 0.10 | 86.99 ± 0.99 | ||
| A | 99.02 ± 0.02 | 13.02 ± 0.00 | 0.45 ± 0.04 | 87.51 ± 0.47 | ||
| 6 | PB | 97.55 ± 0.29 | 13.84 ± 0.79 | 0.41 ± 0.03 | 84.35 ± 0.33 | |
| P | 97.91 ± 0.13 | 13.47 ± 1.54 | 0.55 ± 0.03 | 84.09 ± 0.15 | ||
| A | 98.38 ± 0.19 | 13.06 ± 0.08 | 0.51 ± 0.01 | 85.65 ± 0.25 | ||
| Saithe | 0 | PB | 99.54 ± 0.13 | 11.12 ± 0.23 | 0.30 ± 0.05 | 87.39 ± 0.62 |
| P | 99.66 ± 0.02 | 11.33 ± 0.46 | 0.39 ± 0.02 | 88.10 ± 0.48 | ||
| A | 99.47 ± 0.17 | 10.74 ± 0.05 | 0.36 ± 0.00 | 87.70 ± 0.05 | ||
| 3 | PB | 98.29 ± 0.71 | 11.86 ± 0.17 | 1.27 ± 0.16 | 90.19 ± 0.82 | |
| P | 99.23 ± 0.34 | 13.27 ± 0.13 | 2.38 ± 1.05 | 87.85 ± 1.29 | ||
| A | 98.20 ± 0.97 | 11.68 ± 0.12 | 3.73 ± 3.39 | 88.53 ± 3.52 | ||
| 6 | PB | 98.53 ± 0.16 | 10.89 ± 0.11 | 0.32 ± 0.01 | 89.25 ± 0.03 | |
| P | 98.81 ± 0.05 | 12.49 ± 0.09 | 0.55 ± 0.05 | 87.11 ± 0.04 | ||
| A | 98.77 ± 0.08 | 11.20 ± 0.40 | 0.28 ± 0.03 | 87.54 ± 0.20 |
| Species | Storage Time | Enzyme | >5000 Da | 2000–5000 Da | 1000–2000 Da | 500–1000 Da | 200–500 Da | <200 Da | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Cod | 0 | A | 30.1 | ± | 1.0 | 28.1 | ± | 0.8 | 18.2 | ± | 1.8 | 11.7 | ± | 0.2 | 6.8 | ± | 0.4 | 5.0 | ± | 0.6 |
| P | 21.3 | ± | 1.4 | 23.6 | ± | 1.9 | 26.2 | ± | 2.9 | 13.8 | ± | 0.5 | 8.1 | ± | 0.3 | 6.9 | ± | 1.9 | ||
| PB | 6.7 | ± | 1.8 | 35.5 | ± | 2.3 | 29.0 | ± | 0.7 | 15.3 | ± | 0.1 | 8.6 | ± | 1.6 | 4.9 | ± | 1.5 | ||
| 3 | A | 29.6 | ± | 1.6 | 33.1 | ± | 2.4 | 16.1 | ± | 1.9 | 11.1 | ± | 1.1 | 7.6 | ± | 0.1 | 2.4 | ± | 0.1 | |
| P | 30.4 | ± | 4.8 | 31.8 | ± | 4.7 | 17.4 | ± | 1.3 | 11.0 | ± | 0.9 | 6.9 | ± | 0.7 | 2.5 | ± | 0.1 | ||
| PB | 19.7 | ± | 0.5 | 40.8 | ± | 0.4 | 19.6 | ± | 2.0 | 11.0 | ± | 1.5 | 6.9 | ± | 0.2 | 1.9 | ± | 0.1 | ||
| 6 | A | 29.9 | ± | 1.6 | 32.9 | ± | 1.0 | 20.8 | ± | 0.5 | 7.1 | ± | 0.3 | 6.8 | ± | 0.5 | 2.5 | ± | 0.2 | |
| P | 31.4 | ± | 0.8 | 29.4 | ± | 0.2 | 22.9 | ± | 0.2 | 7.5 | ± | 0.5 | 6.5 | ± | 0.8 | 2.3 | ± | 0.2 | ||
| PB | 20.5 | ± | 0.5 | 41.9 | ± | 0.6 | 23.0 | ± | 0.4 | 6.6 | ± | 0.6 | 5.8 | ± | 0.9 | 2.3 | ± | 0.2 | ||
| Cusk | 0 | A | 18.6 | ± | 3.5 | 33.3 | ± | 2.6 | 21.9 | ± | 3.8 | 15.6 | ± | 1.8 | 8.3 | ± | 0.6 | 2.3 | ± | 0.6 |
| P | 24.5 | ± | 3.5 | 30.6 | ± | 2.8 | 19.2 | ± | 0.9 | 16.4 | ± | 1.2 | 7.3 | ± | 1.9 | 2.0 | ± | 1.4 | ||
| PB | 9.8 | ± | 1.2 | 33.2 | ± | 3.7 | 27.6 | ± | 3.4 | 17.9 | ± | 1.3 | 9.3 | ± | 0.5 | 2.2 | ± | 0.3 | ||
| 3 | A | 36.4 | ± | 1.6 | 32.1 | ± | 1.4 | 17.0 | ± | 0.9 | 8.7 | ± | 0.6 | 4.6 | ± | 0.1 | 1.3 | ± | 0.1 | |
| P | 43.1 | ± | 2.9 | 26.9 | ± | 1.4 | 16.8 | ± | 1.0 | 7.9 | ± | 0.7 | 4.1 | ± | 0.3 | 1.1 | ± | 0.4 | ||
| PB | 30.9 | ± | 0.5 | 38.0 | ± | 0.8 | 18.2 | ± | 0.5 | 7.5 | ± | 0.2 | 4.3 | ± | 0.1 | 1.0 | ± | 0.1 | ||
| 6 | A | 42.8 | ± | 1.7 | 28.5 | ± | 2.4 | 13.1 | ± | 1.2 | 9.4 | ± | 0.7 | 4.9 | ± | 0.1 | 1.3 | ± | 0.1 | |
| P | 46.5 | ± | 1.4 | 26.9 | ± | 1.2 | 14.7 | ± | 0.4 | 6.8 | ± | 0.5 | 4.1 | ± | 0.3 | 1.0 | ± | 0.3 | ||
| PB | 30.2 | ± | 1.1 | 40.5 | ± | 2.4 | 17.6 | ± | 2.5 | 6.7 | ± | 1.7 | 4.2 | ± | 0.3 | 0.9 | ± | 0.2 | ||
| Haddock | 0 | A | 12.9 | ± | 2.1 | 21.4 | ± | 4.5 | 27.5 | ± | 5.5 | 16.0 | ± | 1.0 | 14.0 | ± | 0.3 | 8.2 | ± | 1.5 |
| P | 15.9 | ± | 0.8 | 22.3 | ± | 1.7 | 27.9 | ± | 1.9 | 14.5 | ± | 2.3 | 12.8 | ± | 0.6 | 6.8 | ± | 0.9 | ||
| PB | 8.9 | ± | 2.3 | 36.2 | ± | 2.2 | 23.2 | ± | 2.1 | 14.6 | ± | 1.2 | 9.7 | ± | 0.7 | 7.2 | ± | 1.4 | ||
| 3 | A | 10.1 | ± | 0.3 | 37.4 | ± | 0.5 | 24.2 | ± | 0.8 | 13.6 | ± | 1.0 | 9.1 | ± | 0.1 | 5.6 | ± | 0.1 | |
| P | 20.4 | ± | 1.4 | 32.9 | ± | 1.2 | 22.4 | ± | 0.8 | 10.5 | ± | 0.5 | 8.8 | ± | 0.3 | 4.9 | ± | 0.2 | ||
| PB | 17.6 | ± | 0.7 | 41.2 | ± | 0.8 | 18.7 | ± | 1.2 | 10.3 | ± | 1.1 | 8.5 | ± | 0.2 | 3.8 | ± | 0.2 | ||
| 6 | A | 18.4 | ± | 1.9 | 36.3 | ± | 0.6 | 21.3 | ± | 1.0 | 10.7 | ± | 0.4 | 8.9 | ± | 0.5 | 4.4 | ± | 0.4 | |
| P | 18.6 | ± | 1.2 | 33.3 | ± | 1.3 | 22.0 | ± | 0.3 | 11.0 | ± | 0.2 | 9.4 | ± | 0.3 | 5.6 | ± | 0.1 | ||
| PB | 18.2 | ± | 0.6 | 42.0 | ± | 0.5 | 18.5 | ± | 0.2 | 8.7 | ± | 0.3 | 9.1 | ± | 0.7 | 3.4 | ± | 0.2 | ||
| Saithe | 0 | A | 15.4 | ± | 1.0 | 28.3 | ± | 1.5 | 28.3 | ± | 1.8 | 13.9 | ± | 0.2 | 9.4 | ± | 0.1 | 4.7 | ± | 0.5 |
| P | 21.2 | ± | 1.3 | 26.8 | ± | 2.3 | 23.2 | ± | 1.6 | 14.0 | ± | 0.5 | 9.5 | ± | 0.6 | 5.3 | ± | 0.7 | ||
| PB | 9.2 | ± | 2.8 | 38.1 | ± | 2.4 | 28.2 | ± | 1.0 | 12.4 | ± | 0.7 | 8.7 | ± | 0.8 | 3.4 | ± | 0.4 | ||
| 3 | A | 25.8 | ± | 1.7 | 37.8 | ± | 0.7 | 17.5 | ± | 1.4 | 10.2 | ± | 1.0 | 5.8 | ± | 0.2 | 2.8 | ± | 0.5 | |
| P | 33.7 | ± | 1.9 | 30.1 | ± | 0.7 | 18.0 | ± | 0.6 | 10.3 | ± | 0.5 | 5.4 | ± | 0.3 | 2.5 | ± | 0.2 | ||
| PB | 24.8 | ± | 1.8 | 40.3 | ± | 1.5 | 16.3 | ± | 0.5 | 10.4 | ± | 0.5 | 5.9 | ± | 0.3 | 2.2 | ± | 0.1 | ||
| 6 | A | 30.3 | ± | 2.5 | 33.7 | ± | 2.3 | 18.0 | ± | 1.8 | 9.1 | ± | 1.7 | 6.5 | ± | 0.6 | 2.2 | ± | 0.3 | |
| P | 35.7 | ± | 0.9 | 28.8 | ± | 0.4 | 18.5 | ± | 1.5 | 8.6 | ± | 1.6 | 6.0 | ± | 0.2 | 2.4 | ± | 0.2 | ||
| PB | 31.0 | ± | 0.7 | 37.8 | ± | 1.7 | 16.0 | ± | 3.3 | 7.8 | ± | 1.3 | 5.7 | ± | 0.2 | 1.7 | ± | 0.2 | ||
| Species | Enzyme | Storage Time (Months) | ||
|---|---|---|---|---|
| 0 | 3 | 6 | ||
| Cod | PB | 100 ± 0.1 a,A | 99.5 ± 0.9 a,AB | 100 ± 0.1 a,A |
| P | 100 ± 0.1 a,A | 100 ± 0.1 a,B | 99.8 ± 0.6 a,A | |
| A | 100 ± 0.1 a,A | 97.1 ± 2.7 a,A | 99.9 ± 0.2 a,A | |
| Haddock | PB | 99.4 ± 1.0 a,A | 99.5 ± 1.3 a,A | 95.1 ± 1.0 b,A |
| P | 99.9 ± 0.2 a,A | 100 ± 0.1 a,A | 95.9 ± 2.3 b,A | |
| A | 99.6 ± 0.9 a,A | 99.6 ± 0.6 a,A | 93.4 ± 1.7 b,A | |
| Saithe | PB | 99.7 ± 0.7 a,A | 99.9 ± 0.2 a,A | 94.4 ± 0.8 b,A |
| P | 99.7 ± 0.7 a,A | 99.8 ± 0.5 a,A | 94.7 ± 0.9 b,A | |
| A | 100 ± 0.1 a,A | 100 ± 0.1 a,A | 89.6 ± 2.2 b,B | |
| Tusk | PB | 100 ± 0.1 a,A | 97.3 ± 2.7 b,A | 87.1 ± 1.2 c,A |
| P | 99.0 ± 0.5 a,A | 100 ± 0.1 b,A | 85.9 ± 0.6 c,A | |
| A | 97.8 ± 2.0 a,B | 98.7 ± 1.4 a,A | 84.1 ± 4.0 b,B | |
| Species | Enzymes | Storage Times |
|---|---|---|
| Cod | Alcalase | 0 months |
| Cusk | Papain and Bromelain | 3 months |
| Haddock | Protamex | 6 months |
| Saithe |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Remme, J.F.; Korsnes, S.; Steen, S.; Durand, R.; Kvangarsnes, K.; Stangeland, J. The Effects of Enzymes, Species, and Storage of Raw Material on Physicochemical Properties of Protein Hydrolysates from Whitefish Heads. Mar. Drugs 2023, 21, 587. https://doi.org/10.3390/md21110587
Remme JF, Korsnes S, Steen S, Durand R, Kvangarsnes K, Stangeland J. The Effects of Enzymes, Species, and Storage of Raw Material on Physicochemical Properties of Protein Hydrolysates from Whitefish Heads. Marine Drugs. 2023; 21(11):587. https://doi.org/10.3390/md21110587
Chicago/Turabian StyleRemme, Jannicke Fugledal, Sigurd Korsnes, Stine Steen, Rachel Durand, Kristine Kvangarsnes, and Janne Stangeland. 2023. "The Effects of Enzymes, Species, and Storage of Raw Material on Physicochemical Properties of Protein Hydrolysates from Whitefish Heads" Marine Drugs 21, no. 11: 587. https://doi.org/10.3390/md21110587