
Isolation, Characterization and Immunomodulatory Activity Evaluation of Chrysolaminarin from the Filamentous Microalga Tribonema aequale
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Comparison of Chrysolaminarin Production in Six Tribonema Species
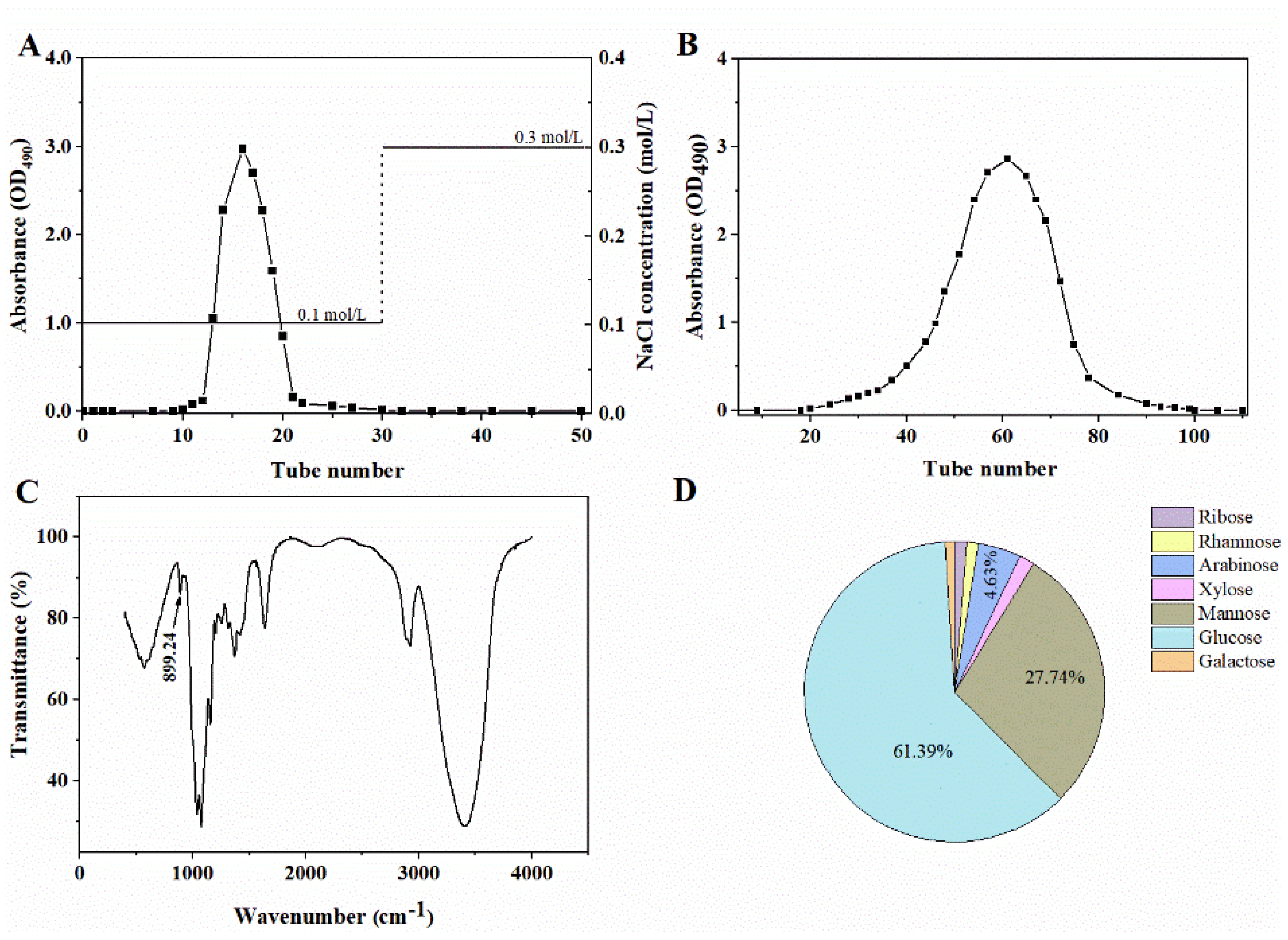
2.2. Preparation and Characterization of Chrysolaminarin from T. aequale
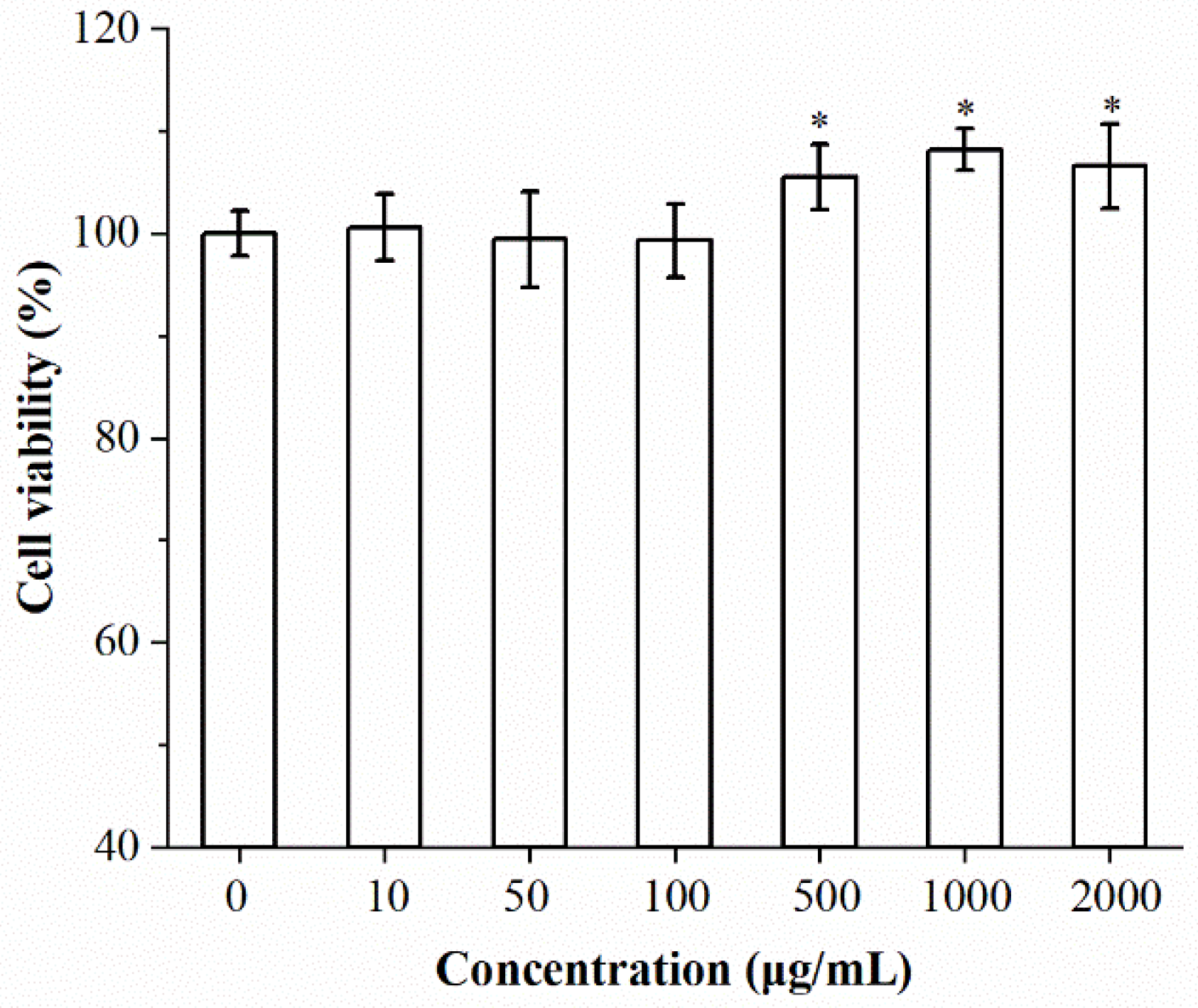
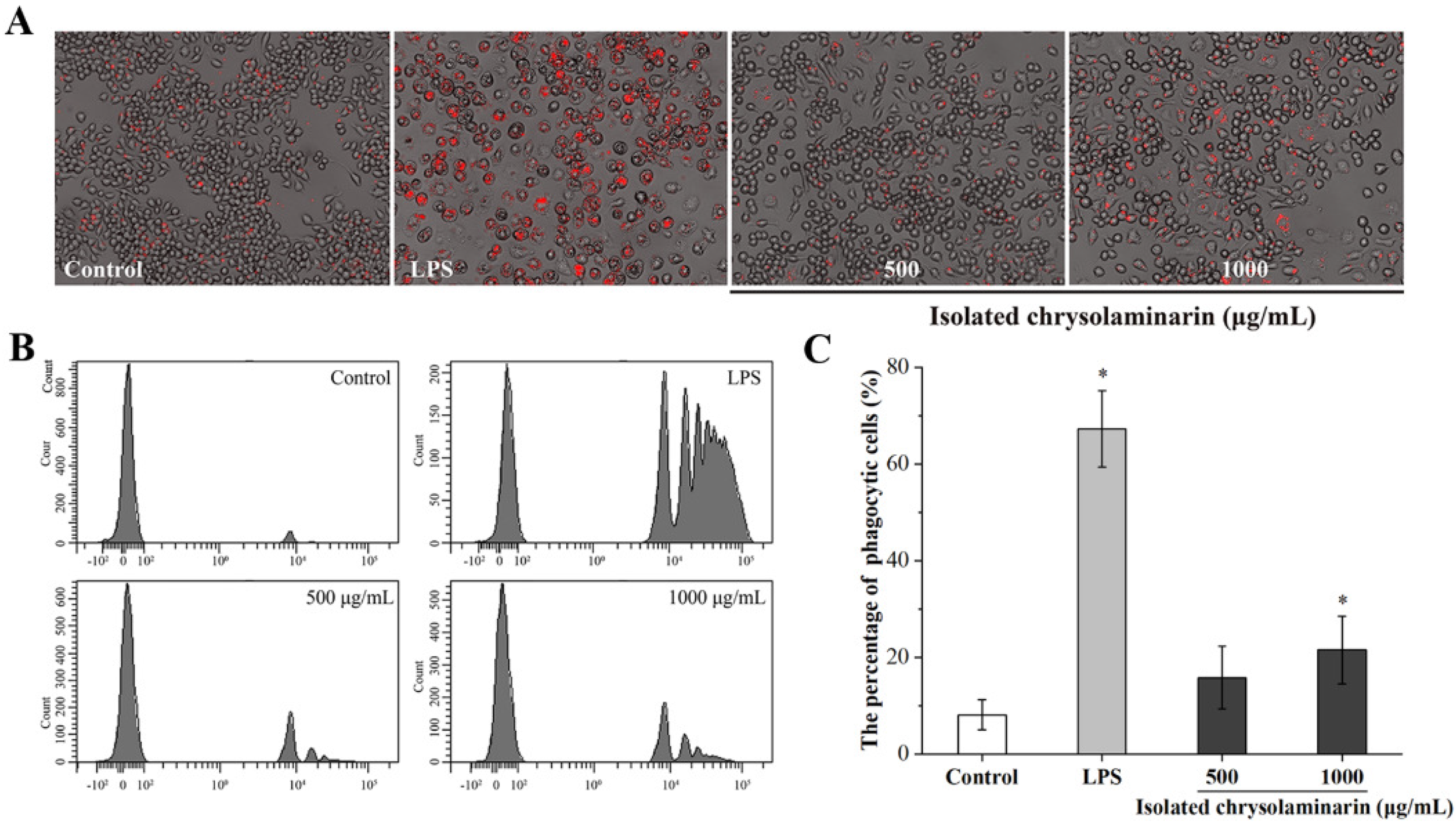
2.3. Effects of Isolated Chrysolaminarin on Macrophages Viability and Phagocytic Activity
2.4. Effects of Isolated Chrysolaminarin on mRNA Expression of Selected Cytokines
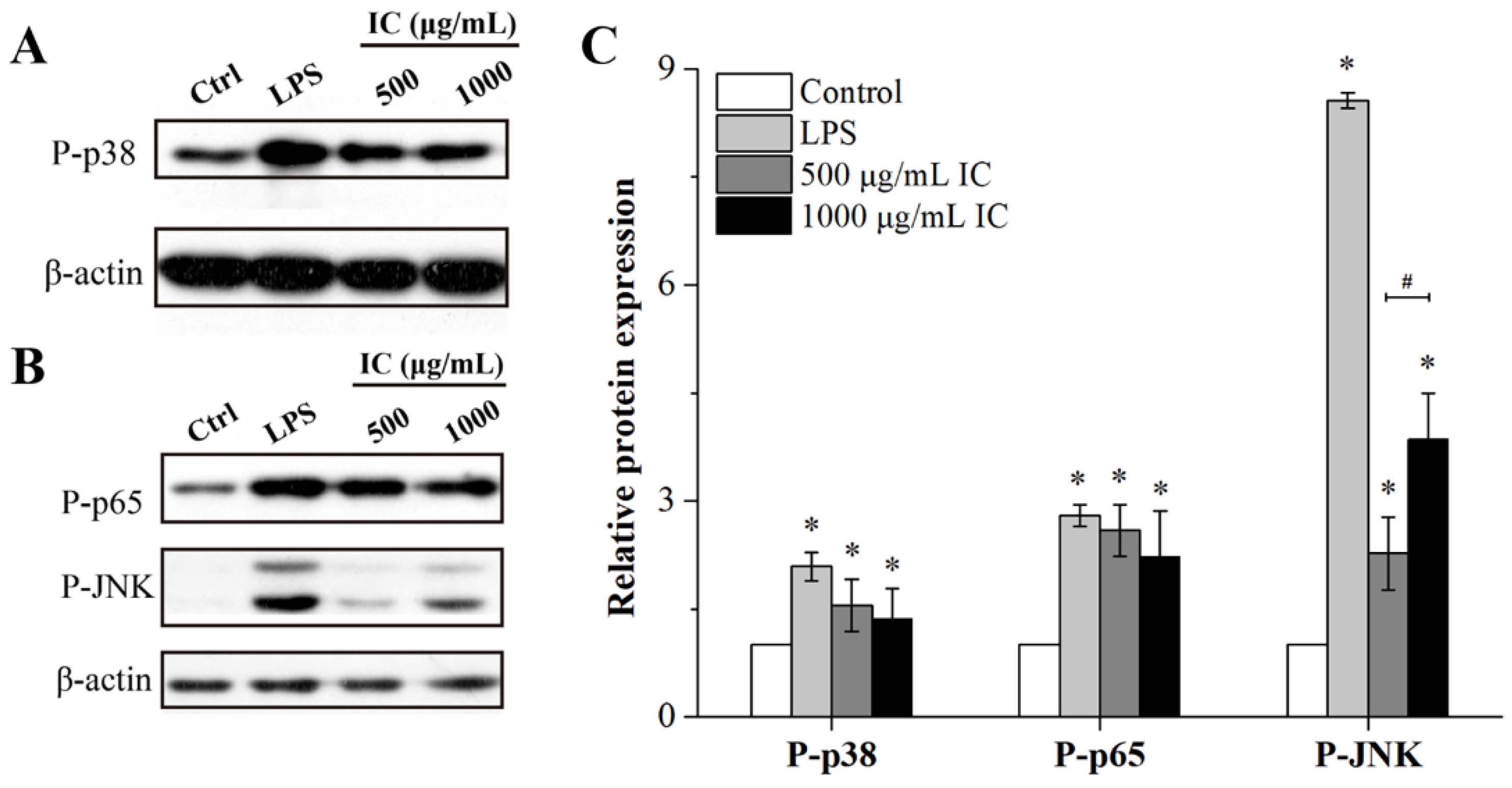
2.5. Effects of Isolated Chrysolaminarin on the MAPK and NF-κB Signaling Pathways
3. Materials and Methods
3.1. Materials
3.2. Microalgae Strains and Culture Conditions
3.3. Determination of Biomass Dry Weight and Chrysolaminarin Content
3.4. Extraction and Isolation of Chrysolaminarin
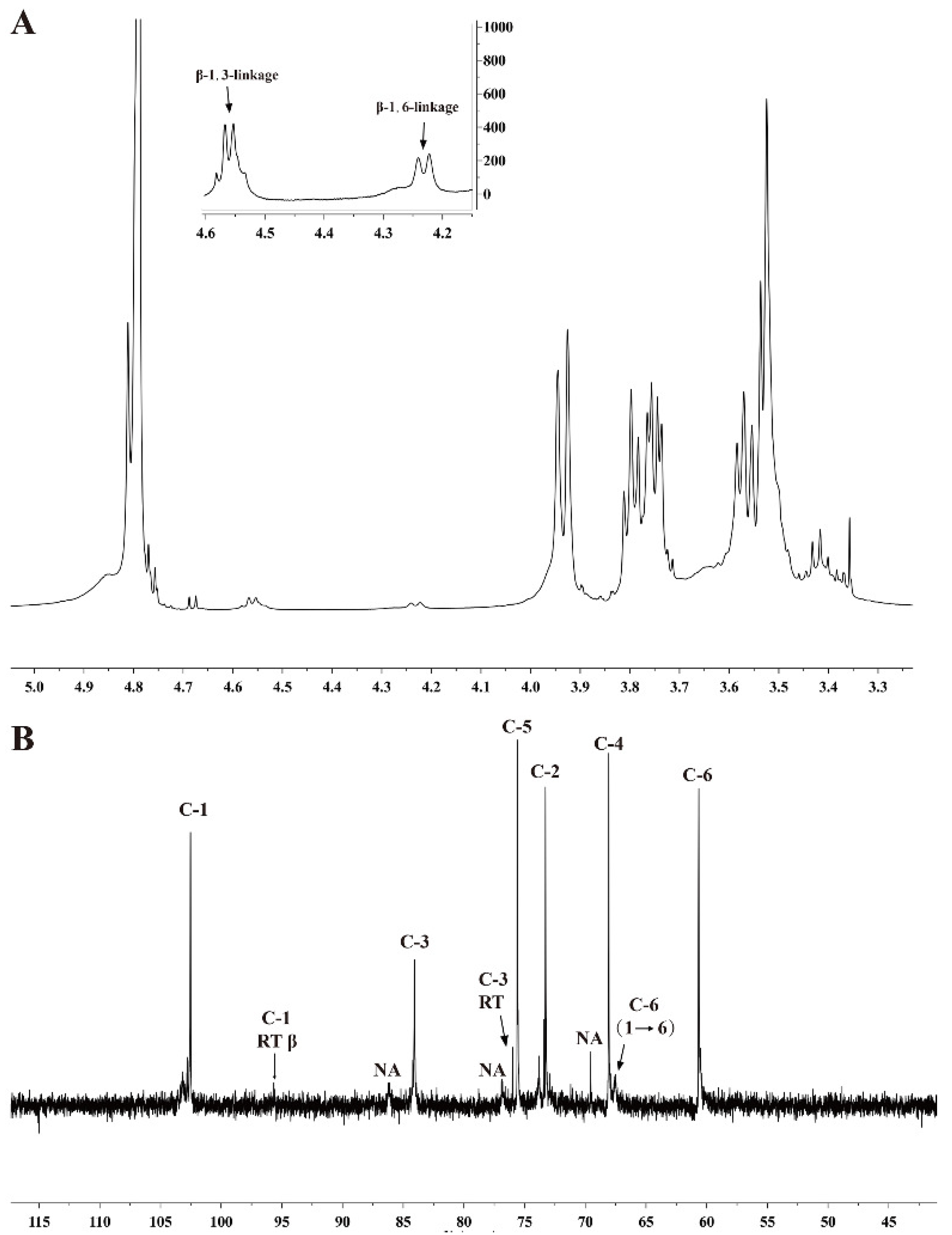
3.5. Characterization of Chrysolaminarin
3.6. Immunomodulatory Activity In Vitro
3.6.1. Cell Culture
3.6.2. RAW 264.7 Cell Viability Assay
3.6.3. Phagocytosis Assay
3.6.4. Quantitative Real-Time PCR for Cytokine Gene Expression in RAW264.7 Cells
3.6.5. Western Blot Analysis
3.7. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, M.; Chen, Y.X.; Zhang, Q.T. A Review of energy consumption in the acquisition of bio-feedstock for microalgae biofuel production. Sustainability 2021, 13, 8873. [Google Scholar] [CrossRef]
- Subhash, G.V.; Rajvanshi, M.; Krishna-Kumar, G.R.; Sagaram, S.; Prasad, V.; Govindachary, S.; Dasgupta, S. Challenges in microalgal biofuel production: A perspective on techno economic feasibility under biorefinery stratagem. Bioresour. Technol. 2022, 343, 126155. [Google Scholar] [CrossRef] [PubMed]
- Gifuni, I.; Pollio, A.; Safi, C.; Marzocchella, A.; Olivieri, G. Current bottlenecks and challenges of the microalgal biorefinery. Trends Biotechnol. 2019, 37, 242–252. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.F.; Gao, B.Y.; Su, M.; Dai, C.M.; Huang, L.F.; Zhang, C.W. Integrated biorefinery strategy for tofu wastewater biotransformation and biomass valorization with the filamentous microalga Tribonema minus. Bioresour. Technol. 2019, 292, 121938. [Google Scholar] [CrossRef]
- Moreira, J.B.; da Silva Vaz, B.; Cardias, B.B.; Cruz, C.G.; de Almeida, A.C.A.; Costa, J.A.V.; de Morais, M.G. Microalgae polysaccharides: An alternative source for food production and sustainable agriculture. Polysaccharides 2022, 3, 441–457. [Google Scholar] [CrossRef]
- Chen, X.L.; Song, L.; Wang, H.; Liu, S.; Yu, H.H.; Wang, X.Q.; Liu, T.Z.; Li, P.C. Partial characterization, the immune modulation and anticancer activities of sulfated polysaccharides from filamentous microalgae Tribonema sp. Molecules 2019, 24, 322. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.Y.; Wang, H.; Guo, G.L.; Pu, Y.F.; Yan, B.L. The isolation and antioxidant activity of polysaccharides from the marine microalgae Isochrysis galbana. Carbohyd. Polym. 2014, 113, 22–31. [Google Scholar] [CrossRef]
- Tounsi, L.; Hentati, F.; Hlima, H.B.; Barkallah, M.; Smaoui, S.; Fendri, I.; Miichaud, P.; Abdelkafi, S. Microalgae as feedstock for bioactive polysaccharides. Int. J. Biol. Macromol. 2022, 221, 1238–1250. [Google Scholar] [CrossRef]
- Kim, M.; Yim, J.H.; Kim, S.Y.; Kim, H.S.; Lee, W.G.; Kim, S.J.; Kang, P.S.; Lee, C.K. In vitro inhibition of influenza A virus infection by marine microalga-derived sulfated polysaccharide p-KG03. Antiviral Res. 2012, 93, 253–259. [Google Scholar] [CrossRef]
- Costa, J.A.V.; Lucas, B.F.; Alvarenga, A.G.P.; Moreira, J.B.; de Morais, M.G. Microalgae polysaccharides: An overview of production, characterization, and potential applications. Polysaccharides 2021, 2, 759–772. [Google Scholar] [CrossRef]
- Arad, S.M.; Levy-Ontman, O. Red microalgal cell-wall polysaccharides: Biotechnological aspects. Curr. Opin. Biotechnol. 2010, 21, 358–364. [Google Scholar] [CrossRef] [PubMed]
- Carballo, C.; Chronopoulou, E.G.; Letsiou, S.; Maya, C.; Labrou, N.E.; Infante, C.; Power, D.M.; Manuel, M. Antioxidant capacity and immunomodulatory effects of a chrysolaminarin-enriched extract in Senegalese sole. Fish Shellfish Immunol. 2018, 82, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Pulz, O.; Gross, W. Valuable products from biotechnology of microalgae. Appl. Microbiol. Biotechnol. 2004, 65, 635–648. [Google Scholar] [CrossRef] [PubMed]
- Kusaikin, M.I.; Ermakova, S.P.; Shevchenko, N.M.; Isakov, V.V.; Gorshkov, A.G.; Vereshchagin, A.L.; Grachev, M.A.; Zvyagintseva, T.N. Structural characteristics and antitumor activity of a new chrysolaminaran from the diatom alga Synedra acus. Chem. Nat. Compd. 2010, 46, 1–4. [Google Scholar] [CrossRef]
- Xia, S.; Gao, B.Y.; Li, A.F.; Xiong, J.H.; Ao, Z.Q.; Zhang, C.W. Preliminary characterization, antioxidant properties and production of chrysolaminarin from marine diatom Odontella aurita. Mar. Drugs 2014, 12, 4883–4897. [Google Scholar] [CrossRef] [Green Version]
- Zuccarello, G.C.; Lokhorst, G.M. Molecular phylogeny of the genus Tribonema (Xanthophyceae) using rbc L gene sequence data: Monophyly of morphologically simple algal species. Phycologia 2005, 44, 384–392. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, Y.; Zhou, W.J.; Noppol, L.; Liu, T.Z. Mechanism and enhancement of lipid accumulation in filamentous oleaginous microalgae Tribonema minus under heterotrophic condition. Biotechnol. Biofuels 2018, 11, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Long, J.J.; Jia, J.; Gong, Y.C.; Han, D.X.; Hu, Q. Assessment of eicosapentaenoic acid (EPA) production from filamentous microalga Tribonema aequale: From laboratory to pilot-scale study. Mar. Drugs 2022, 20, 343. [Google Scholar] [CrossRef]
- Huo, S.H.; Wang, H.Y.; Chen, J.; Hu, X.J.; Zan, X.Y.; Zhang, C.S.; Qian, J.Y.; Zhu, F.F.; Ma, H.L.; Elshobary, M. A preliminary study on polysaccharide extraction, purification, and antioxidant properties of sugar-rich filamentous microalgae Tribonema minus. J. Appl. Phycol. 2022, 34, 2755–2767. [Google Scholar] [CrossRef]
- Wang, F.F.; Gao, B.Y.; Dai, C.M.; Su, M.; Zhang, C.W. Comprehensive utilization of the filamentous oleaginous microalga Tribonema utriculosum for the production of lipids and chrysolaminarin in a biorefinery concept. Algal Res. 2020, 50, 101973. [Google Scholar] [CrossRef]
- Necchi, O. Heterokonts (Xanthophyceae and Chrysophyceae) in rivers. In River Algae; Springer: Cham, Switzerland, 2016; pp. 153–158. [Google Scholar]
- Zhang, W.Y.; Wang, F.F.; Gao, B.Y.; Huang, L.D.; Zhang, C.W. An integrated biorefinery process: Stepwise extraction of fucoxanthin, eicosapentaenoic acid and chrysolaminarin from the same Phaeodactylum tricornutum biomass. Algal Res. 2018, 32, 193–200. [Google Scholar] [CrossRef]
- Wu, W.L.; Zhu, Y.T.; Zhang, L.; Yang, R.W.; Zhou, Y.H. Extraction, preliminary structural characterization, and antioxidant activities of polysaccharides from Salvia miltiorrhiza Bunge. Carbohydr. Polym. 2012, 87, 1348–1353. [Google Scholar] [CrossRef]
- Zhang, Y.X.; Dai, L.; Kong, X.W.; Chen, L.W. Characterization and in vitro antioxidant activities of polysaccharides from Pleurotus ostreatus. Int. J. Biol. Macromol. 2012, 51, 259–265. [Google Scholar] [CrossRef] [PubMed]
- Zhan, Q.P.; Wang, Q.; Lin, R.; He, P.; Lai, F.; Zhang, M.M.; Wu, H. Structural characterization and immunomodulatory activity of a novel acid polysaccharide isolated from the pulp of Rosa laevigata Michx fruit. Int. J. Biol. Macromol. 2020, 145, 1080–1090. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Shen, X.Y.; Yin, K.Y.; Miao, C.Q.; Sun, Y.L.; Mao, S.M.; Liu, D.M.; Sheng, J.W. Structural characteristics and immune-enhancing activity of fractionated polysaccharides from Athyrium Multidentatum (Doll.) Ching. Int. J. Biol. Macromol. 2022, 205, 76–89. [Google Scholar] [CrossRef]
- Guan, X.F.; Wang, Q.; Lin, B.; Sun, M.L.; Zheng, Q.; Huang, J.Q.; Lai, G.T. Structural characterization of a soluble polysaccharide SSPS1 from soy whey and its immunoregulatory activity in macrophages. Int. J. Biol. Macromol. 2022, 217, 131–141. [Google Scholar] [CrossRef]
- Zou, J.; Secombes, C.J. The function of fish cytokines. Biology 2016, 5, 23. [Google Scholar] [CrossRef]
- Lechner, M.; Lirk, P.; Rieder, J. Inducible nitric oxide synthase (iNOS) in tumor biology: The two sides of the same coin. Semin. Cancer Biol. 2006, 15, 277–289. [Google Scholar] [CrossRef]
- Lee, J.S.; Kwon, D.S.; Lee, K.R.; Park, J.M.; Ha, S.J.; Hong, E.K. Mechanism of macrophage activation induced by polysaccharide from Cordyceps militaris culture broth. Carbohydr. Polym. 2015, 120, 29–37. [Google Scholar] [CrossRef]
- Liu, X.X.; Li, J.; Peng, X.H.; Lv, B.; Wang, P.; Zhao, X.M.; Yu, B. Geraniin inhibits LPS-induced THP-1 macrophages switching to M1 phenotype via SOCS1/NF-κB pathway. Inflammation 2016, 39, 1421–1433. [Google Scholar] [CrossRef]
- Gao, B.Y.; Yang, J.; Lei, X.Q.; Xia, S.; Li, A.F.; Zhang, C.W. Characterization of cell structural change, growth, lipid accumulation, and pigment profile of a novel oleaginous microalga, Vischeria stellata (Eustigmatophyceae), cultured with different initial nitrate supplies. J. Appl. Psychol. 2016, 28, 821–830. [Google Scholar] [CrossRef]
- Wang, F.F.; Chen, J.M.; Zhang, C.W.; Gao, B.Y. Resourceful treatment of cane sugar industry wastewater by Tribonema minus towards the production of valuable biomass. Bioresour. Technol. 2020, 316, 123902. [Google Scholar] [CrossRef] [PubMed]
- Palanisamy, S.; Vinosha, M.; Marudhupandi, T.; Rajasekar, P.; Prabhu, N.M. Isolation of fucoidan from Sargassum polycystum brown algae: Structural characterization, in vitro antioxidant and anticancer activity. Int. J. Biol. Macromol. 2017, 102, 405–412. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.M.; Guan, Q.N.; Guo, Y.H.; Du, C.G. Reduction of inflammatory hyperplasia in the intestine in colon cancer-prone mice by water-extract of Cistanche deserticola. Phytother. Res. 2012, 26, 812–819. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, F.; Yang, R.; Guo, Y.; Zhang, C. Isolation, Characterization and Immunomodulatory Activity Evaluation of Chrysolaminarin from the Filamentous Microalga Tribonema aequale. Mar. Drugs 2023, 21, 13. https://doi.org/10.3390/md21010013
Wang F, Yang R, Guo Y, Zhang C. Isolation, Characterization and Immunomodulatory Activity Evaluation of Chrysolaminarin from the Filamentous Microalga Tribonema aequale. Marine Drugs. 2023; 21(1):13. https://doi.org/10.3390/md21010013
Chicago/Turabian StyleWang, Feifei, Rundong Yang, Yuhao Guo, and Chengwu Zhang. 2023. "Isolation, Characterization and Immunomodulatory Activity Evaluation of Chrysolaminarin from the Filamentous Microalga Tribonema aequale" Marine Drugs 21, no. 1: 13. https://doi.org/10.3390/md21010013