
Seaweed as a Natural Source against Phytopathogenic Bacteria
 ,
,  and
and 
Abstract
:1. Introduction
2. Material and Methods
3. Phytopathogenic Bacteria
4. Phytopathogenic Antibacterial Potential of Seaweeds
Putative Mechanisms of Antibacterial Action
5. Seaweed Potential as Plant-Priming Agent
6. Seaweed Elicitors

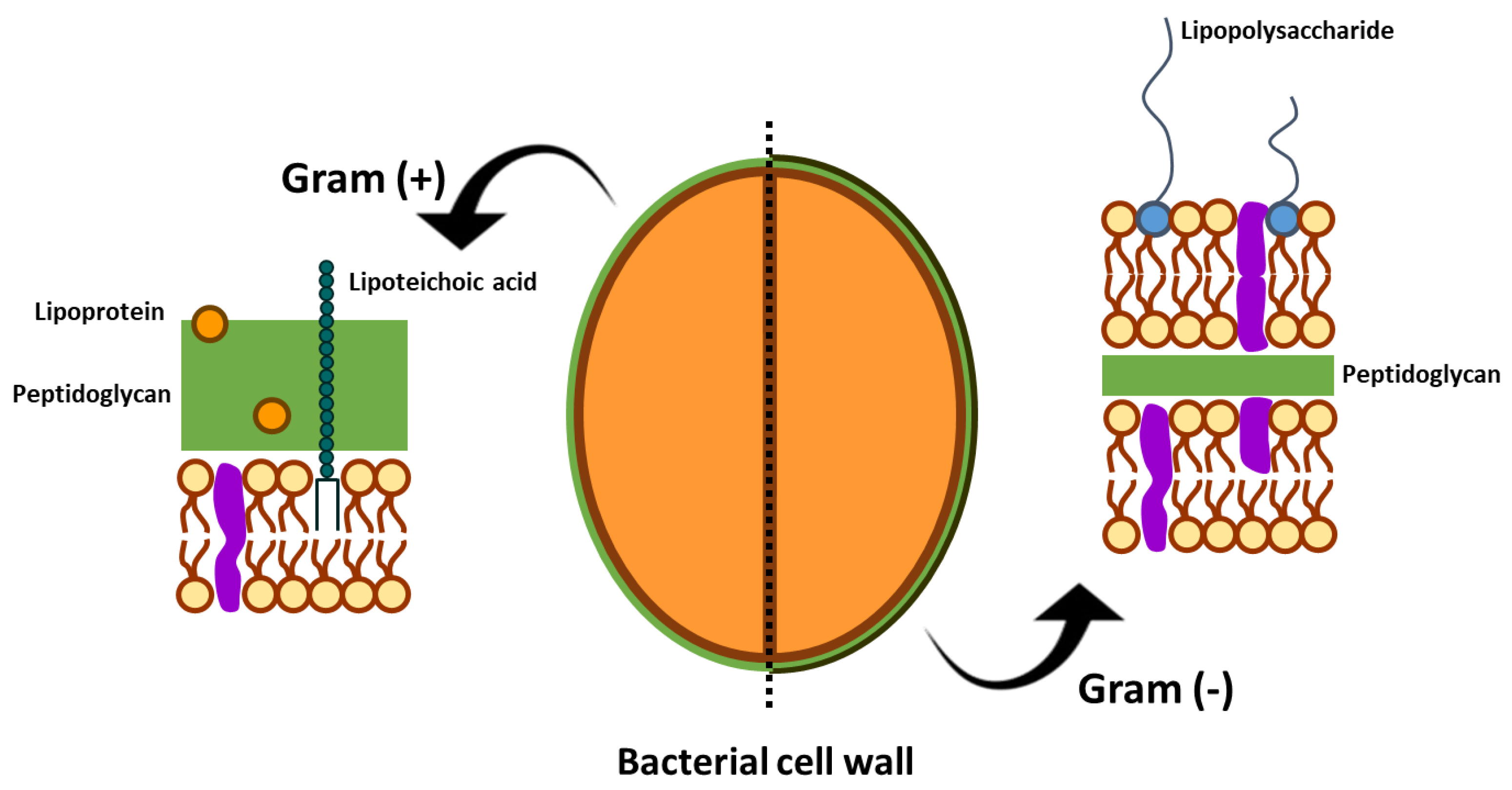{kind=link}
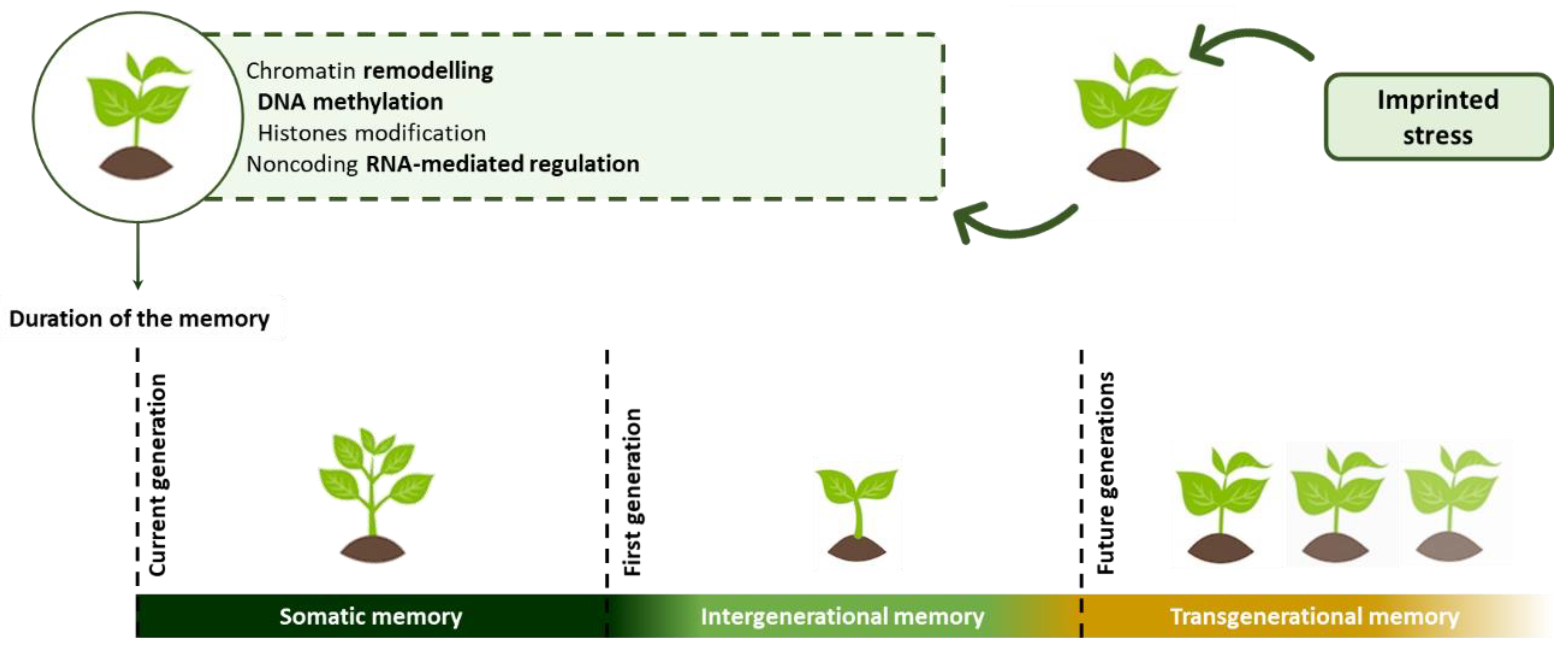{kind=link}
| Species | Seaweed | Extract/Solvent | References |
|---|---|---|---|
| Agrobacterium tumefaciens | Fucus spiralis | Aqueous extract | [130] |
| Cystoseira myriophylloides | Aqueous extract | [130] | |
| Erwinia carotovora subsp. carotovora | Laminaria digitata | Purified laminarin A | [225] |
| Pseudomonas aeruginosa | Ascophyllum nodosum | - | [231] |
| Ascophyllum nodosum | Stella Maris® | [227] | |
| Pseudomonas syringae | Ascophyllum nodosum | Stella Maris® | [227] |
| Pseudomonas syringae pv. tabaci | Cystoseira myriophylloides | Aqueous extract | [71] |
| Fucus spiralis | Aqueous extract | [71] | |
| Laminaria digitata | Aqueous extract | [71] | |
| Pseudomonas syringae pv. tomato | Ascophyllum nodosum | Aqueous extract | [179] |
| Ascophyllum nodosum | Chloroform extract | [179] | |
| Ascophyllum nodosum | Ethyl acetate | [179] | |
| Kappaphycus alvarezii | Aqueous extract | [232] | |
| Staphylococcus aureus | Ascophyllum nodosum | Essential oils | [231] |
| Xanthomonas campestris | Ascophyllum nodosum | Stella Maris® | [227] |
| Xanthomonas campestris pv. malvacearum | Sargassum wightii | Aqueous extract (Dravya) | [233] |
| Xanthomonas campestris pv vesicatoria | Ascophyllum nodosum | Alkaline extract (commercial product) | [6,56] |
| Acanthophora spicifera | Alkaline extract | [234] | |
| Gelidium serrulatum | Alkaline extract | [235] | |
| Sargassum filipendula | Alkaline extract | [235] | |
| Sargassum vulgare | Alkaline extract | [234] | |
| Ulva lactuca | Alkaline extract | [235] | |
| Xanthomonas oryzae pv oryzae | Kappaphycus alvarezii | Aqueous extraction * | [232] |
7. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rahman, S.F.S.A.; Singh, E.; Pieterse, C.M.J.; Schenk, P.M. Emerging Microbial Biocontrol Strategies for Plant Pathogens. Plant Sci. 2017, 267, 102–111. [Google Scholar] [CrossRef] [Green Version]
- Hamed, S.M.; Abd El-Rhman, A.A.; Abdel-Raouf, N.; Ibraheem, I.B.M. Role of Marine Macroalgae in Plant Protection & Improvement for Sustainable Agriculture Technology. Beni-Suef Univ. J. Basic Appl. Sci. 2018, 7, 104–110. [Google Scholar] [CrossRef]
- Martins, P.M.M.; Merfa, M.V.; Takita, M.A.; De Souza, A.A. Persistence in Phytopathogenic Bacteria: Do We Know Enough? Front. Microbiol. 2018, 9, 1099. [Google Scholar] [CrossRef]
- Kannan, V.R.; Bastas, K.K.; Devi, R.S. Scientific and Economic Impact of Plant Pathogenic Bacteria. In Sustainable Approaches to Controlling Plant Pathogenic Bacteria; Kannan, R.V., Bastas, K.K., Eds.; CRC Press: Boca Raton, FL, USA, 2015; pp. 369–392. [Google Scholar] [CrossRef]
- Savary, S.; Ficke, A.; Aubertot, J.-N.; Hollier, C. Crop Losses due to Diseases and Their Implications for Global Food Production Losses and Food Security. Food Secur. 2012, 4, 519–537. [Google Scholar] [CrossRef]
- Ali, N.; Ramkissoon, A.; Ramsubhag, A.; Jayaraj, J. Ascophyllum Extract Application Causes Reduction of Disease Levels in Field Tomatoes Grown in a Tropical Environment. Crop Prot. 2016, 83, 67–75. [Google Scholar] [CrossRef]
- Farag, S.M.A.; Elhalag, K.M.A.; Hagag, M.H.; Khairy, A.S.M.; Ibrahim, H.M.; Saker, M.T.; Messiha, N.A.S. Potato Bacterial Wilt Suppression and Plant Health Improvement after Application of Different Antioxidants. J. Phytopathol. 2017, 165, 522–537. [Google Scholar] [CrossRef]
- Van der Wolf, J.; De Boer, S.H. Phytopathogenic Bacteria. In Principles of Plant-Microbe Interactions; Springer International Publishing: Cham, Switzerland, 2016; pp. 65–77. [Google Scholar] [CrossRef]
- Bellgard, S.E.; Williams, S.E. Response of Mycorrhizal Diversity to Current Climatic Changes. Diversity 2011, 3, 8–90. [Google Scholar] [CrossRef] [Green Version]
- Leducq, J.B.; Charron, G.; Samani, P.; Dubé, A.K.; Sylvester, K.; James, B.; Almeida, P.; Sampaio, J.P.; Hittinger, C.T.; Bell, G.; et al. Local Climatic Adaptation in a Widespread Microorganism. Proc. R. Soc. B Biol. Sci. 2014, 281, 20132472. [Google Scholar] [CrossRef]
- Anderson, P.K.; Cunningham, A.A.; Patel, N.G.; Morales, F.J.; Epstein, P.R.; Daszak, P. Emerging Infectious Diseases of Plants: Pathogen Pollution, Climate Change and Agrotechnology Drivers. Trends Ecol. Evol. 2004, 19, 535–544. [Google Scholar] [CrossRef]
- Harvell, C.D.; Mitchell, C.E.; Ward, J.R.; Altizer, S.; Dobson, A.P.; Ostfeld, R.S.; Samuel, M.D. Climate Warming and Disease Risks for Terrestrial and Marine Biota. Science 2002, 296, 2158–2162. [Google Scholar] [CrossRef] [Green Version]
- Zuluaga, A.P.; Puigvert, M.; Valls, M. Novel Plant Inputs Influencing Ralstonia solanacearum during Infection. Front. Microbiol. 2013, 4, 349. [Google Scholar] [CrossRef] [Green Version]
- Badosa, E.; Ferre, R.; Planas, M.; Feliu, L.; Montesinos, E.; Cabrefiga, J.; Bardajı, E.; Besalu, E. A Library of Linear Undecapeptides with Bactericidal Activity against Phytopathogenic Bacteria. Peptides 2007, 28, 2276–2285. [Google Scholar] [CrossRef]
- De Waard, M.A.; Georgopoulos, S.G.; Hollomon, D.W.; Ishii, H.; Leroux, P.; Ragsdale, N.N.; Schwinn, F.J. Chemical Control of Plant Diseases: Problems and Prospects. Annu. Rev. Phytopathol. 1993, 31, 403–421. [Google Scholar] [CrossRef]
- Hahn, M. The Rising Threat of Fungicide Resistance in Plant Pathogenic Fungi: Botrytis as a Case Study. J. Chem. Biol. 2014, 7, 133–141. [Google Scholar] [CrossRef] [Green Version]
- Ma, Z.; Michailides, T.J. Advances in Understanding Molecular Mechanisms of Fungicide Resistance and Molecular Detection of Resistant Genotypes in Phytopathogenic Fungi. Crop Prot. 2005, 24, 853–863. [Google Scholar] [CrossRef]
- Zhang, Z.; Chen, Y.; Li, B.; Chen, T.; Tian, S. Reactive Oxygen Species: A Generalist in Regulating Development and Pathogenicity of Phytopathogenic Fungi. Comput. Struct. Biotechnol. J. 2020, 18, 3344–3349. [Google Scholar] [CrossRef]
- Glick, B.R.; Bashan, Y. Genetic Manipulation of Plant Growth-Promoting Bacteria to Enhance Biocontrol of Phytopathogens. Biotechnol. 1997, 15, 353–378. [Google Scholar] [CrossRef] [Green Version]
- Vidaver, A.K. Prospects for Control of Phytopathogenic Bacteria by Bacteriophages and Bacteriocins. Annu. Rev. Phytopathol. 1976, 14, 451–465. [Google Scholar] [CrossRef]
- ŽIvković, S.; Stojanović, S.; Ivanović, Ž.; Gavrilović, V.; Popović, T.; Balaž, J. Screening of Antagonistic Activity of Microorganisms against Colletotrichum acutatum and Colletotrichum gloeosporioides. Arch. Biol. Sci. 2010, 62, 611–623. [Google Scholar] [CrossRef]
- Dimkić, I.; Živković, S.; Berić, T.; Ivanović, Ž.; Gavrilović, V.; Stanković, S.; Fira, D. Characterization and Evaluation of Two Bacillus Strains, SS-12.6 and SS-13.1, as Potential Agents for the Control of Phytopathogenic Bacteria and Fungi. Biol. Control 2013, 65, 312–321. [Google Scholar] [CrossRef]
- Von der Weid, I.; Alviano, D.S.; Santos, A.L.S.; Soares, R.M.A.; Alviano, C.S.; Seldin, L. Antimicrobial Activity of Paenibacillus Peoriae Strain NRRL BD-62 against a Broad Spectrum of Phytopathogenic Bacteria and Fungi. J. Appl. Microbiol. 2003, 95, 1143–1151. [Google Scholar] [CrossRef] [PubMed]
- Walsh, U.F.; Morrissey, J.P.; O’Gara, F. Pseudomonas for Biocontrol of Phytopathogens: From Functional Genomics to Commercial Exploitation. Curr. Opin. Biotechnol. 2001, 12, 289–295. [Google Scholar] [CrossRef] [PubMed]
- Cesa-Luna, C.; Baez, A.; Quintero-Hernández, V.; De La Cruz-Enríquez, J.; Castañeda-Antonio, M.D.; Muñoz-Rojas, J. The Importance of Antimicrobial Compounds Produced by Beneficial Bacteria on the Biocontrol of Phytopathogens. Acta Biol. Colomb. 2020, 25, 140–154. [Google Scholar] [CrossRef]
- Elshahat, M.R.; Ahmed, A.A.; Enas, A.H.; Fekria, M.S. Plant Growth Promoting Rhizobacteria and Their Potential for Biocontrol of Phytopathogens. Afr. J. Microbiol. Res. 2016, 10, 486–504. [Google Scholar] [CrossRef] [Green Version]
- Khabbaz, S.E.; Zhang, L.; Cáceres, L.A.; Sumarah, M.; Wang, A.; Abbasi, P.A. Characterisation of Antagonistic Bacillus and Pseudomonas Strains for Biocontrol Potential and Suppression of Damping-off and Root Rot Diseases. Ann. Appl. Biol. 2015, 166, 456–471. [Google Scholar] [CrossRef]
- Tontou, R.; Gaggia, F.; Baffoni, L.; Devescovi, G.; Venturi, V.; Giovanardi, D.; Stefani, E. Molecular Characterisation of an endophyte showing a strong antagonistic activity against Pseudomonas syringae pv. actinidiae. Plant Soil 2016, 405, 97–106. [Google Scholar] [CrossRef]
- Sekhar, A.C.; Thomas, P. Isolation and Identification of Shoot-Tip Associated Endophytic Bacteria from Banana Cv. Grand Naine and Testing for Antagonistic Activity against Fusarium oxysporum f. sp. cubense. Am. J. Plant Sci. 2015, 6, 943–954. [Google Scholar] [CrossRef] [Green Version]
- Huo, Y.; Kang, J.-P.; Kim, Y.-J.; Yang, D.-C. Paraburkholderia Panacihumi Sp. Nov., an Isolate from Ginseng-Cultivated Soil, Is Antagonistic against Root Rot Fungal Pathogen. Arch. Microbiol. 2018, 200, 1151–1158. [Google Scholar] [CrossRef]
- Van der Putten, W.H.; Cook, R.; Costa, S.; Davies, K.G.; Fargette, M.; Freitas, H.; Hol, W.H.G.; Kerry, B.R.; Maher, N.; Mateille, T.; et al. Nematode Interactions in Nature: Models for Sustainable Control of Nematode Pests of Crop Plants? Adv. Agron. 2006, 89, 227–260. [Google Scholar] [CrossRef]
- Ran, L.X.; Liu, C.Y.; Wu, G.J.; van Loon, L.C.; Bakker, P.A.H.M. Suppression of bacterial wilt in Eucalyptus Urophylla by fluorescent Pseudomonas spp. in China. Biol. Control 2005, 32, 111–120. [Google Scholar] [CrossRef]
- Siddiqui, I.A.; Shaukat, S.S. Plant Species, Host Age and Host Genotype Effects on Meloidogyne Incognita Biocontrol by Pseudomonas Fluorescens Strain CHA0 and Its Genetically-Modified Derivatives. J. Phytopathol. 2003, 151, 231–238. [Google Scholar] [CrossRef]
- Notz, R.; Maurhofer, M.; Duffy, B.; Haas, D.; Défago, G. Biotic Factors Affecting Expression of the 2,4-Diacetylphloroglucinol Biosynthesis Gene PhlA in Pseudomonas fluorescens Biocontrol Strain CHA0 in the Rhizosphere. Biol. Control 2001, 91, 873–881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clough, S.E.; Jousset, A.; Elphinstone, J.G.; Friman, V.-P. Combining in vitro and in vivo screening to identify efficient Pseudomonas biocontrol strains against the phytopathogenic bacterium Ralstonia solanacearum. Microbiologyopen 2022, 11, e1283. [Google Scholar] [CrossRef] [PubMed]
- DeGray, G.; Rajasekaran, K.; Smith, F.; Sanford, J.; Daniell, H. Expression of an Antimicrobial Peptide via the Chloroplast Genome to Control Phytopathogenic Bacteria and Fungi. Plant Physiol. 2001, 127, 852–862. [Google Scholar] [CrossRef] [PubMed]
- Leandro, A.; Pereira, L. Diverse Applications of Marine Macroalgae. Mar. Drugs 2020, 18, 17. [Google Scholar] [CrossRef] [Green Version]
- Kiuru, P.; D’Auria, M.V.; Muller, C.D.; Tammela, P.; Vuorela, H.; Yli-Kauhaluoma, J. Exploring Marine Resources for Bioactive Compounds. Planta Med. 2014, 80, 1234–1246. [Google Scholar] [CrossRef]
- Nawaz, A.; Chaudhary, R.; Shah, Z.; Dufossé, L.; Fouillaud, M.; Mukhtar, H.; Haq, I.U. An Overview on Industrial and Medical Applications of Bio-Pigments Synthesized by Marine Bacteria. Microorganisms 2021, 9, 11. [Google Scholar] [CrossRef]
- McLachlan, J. Macroalgae (Seaweeds): Industrial Resources and Their Utilization. Plant Soil 1985, 89, 137–157. [Google Scholar] [CrossRef]
- Olesen, P.E.; Maretzki, A.; Almodovar, L.A. An Investigation of Antimicrobial Substances from Marine Algae. Bot. Mar. 1964, 6, 3–4, 224–232. [Google Scholar] [CrossRef]
- Milledge, J.J.; Nielsen, B.V.; Bailey, D. High-Value Products from Macroalgae: The Potential Uses of the Invasive Brown Seaweed, Sargassum muticum. Rev. Environ. Sci. Biotechnol. 2016, 15, 67–88. [Google Scholar] [CrossRef]
- Tyśkiewicz, K.; Tyśkiewicz, R.; Konkol, M.; Rój, E.; Jaroszuk-Ściseł, J.; Skalicka-Woźniak, K. Antifungal Properties of Fucus vesiculosus L. Supercritical Fluid Extract Against Fusarium culmorum and Fusarium oxysporum. Molecules 2019, 24, 3518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabrita, M.T.; Vale, C.; Rauter, A.P. Halogenated Compounds from Marine Algae. Mar. Drugs 2010, 8, 2301–2317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kladi, M.; Vagias, C.; Roussis, V. Volatile Halogenated Metabolites from Marine Red Algae. Phytochem. Rev. 2004, 3, 337–366. [Google Scholar] [CrossRef]
- Shannon, E.; Abu-Ghannam, N. Antibacterial Derivatives of Marine Algae: An Overview of Pharmacological Mechanisms and Applications. Mar. Drugs 2016, 14, 81. [Google Scholar] [CrossRef] [PubMed]
- Asimakis, E.; Shehata, A.A.; Eisenreich, W.; Acheuk, F.; Lasram, S.; Basiouni, S.; Emekci, M.; Ntougias, S.; Taner, G.; May-Simera, H.; et al. Algae and Their Metabolites as Potential Bio-Pesticides. Microorganisms 2022, 10, 307. [Google Scholar] [CrossRef] [PubMed]
- Ibraheem, I.B.M.; Hamed, S.M.; Abd Elrhman, A.A.; Farag, F.M.; Abdel-Raouf, N. Antimicrobial activities of some brown macroalgae against some soil borne plant pathogens and in vivo management of Solanum Melongena root diseases. Aust. J. Basic Appl. Sci. 2017, 11, 157–168. [Google Scholar]
- Poveda, J.; Díez-Méndez, A. Use of Elicitors from Macroalgae and Microalgae in the Management of Pests and Diseases in Agriculture. Phytoparasitica 2022, 1–35. [Google Scholar] [CrossRef]
- Rafiee, H.; Badi, H.N.; Mehrafarin, A.; Qaderi, A.; Zarinpanjeh, N.; Sekara, A.; Zand, E. Application of Plant Biostimulants as New Approach to Improve the Biological Responses of Medicinal Plants-A Critical Review. J. Med. Plants 2016, 15, 6–39. [Google Scholar]
- Shukla, P.S.; Mantin, E.G.; Adil, M.; Bajpai, S.; Critchley, A.T.; Prithiviraj, B. Ascophyllum Nodosum-Based Biostimulants: Sustainable Applications in Agriculture for the Stimulation of Plant Growth, Stress Tolerance, and Disease Management. Front. Plant Sci. 2019, 10, 655. [Google Scholar] [CrossRef] [Green Version]
- Bahcevandziev, K.; Pereira, L. Seaweeds as Plant Fertilizer, Agricultural Biostimulants and Animal Fodder—A Book Presentation. Acta Hortic. 2021, 1320, 405–412. [Google Scholar] [CrossRef]
- Fernandes, A.L.T.; de Silva, R.O. Avaliação Do Extrato de Algas (Ascophyllum Nodosum) No Desenvolvimento Vegetativo e Produtivo Do Cafeeiro Irrigado Por Gotejamento e Cultivado Em Condições de Cerrado. Enciclopédia Biostera 2011, 7, 147–157. [Google Scholar]
- Jayaraman, J.; Norrie, J.; Punja, Z.K. Commercial Extract from the Brown Seaweed Ascophyllum Nodosum Reduces Fungal Diseases in Greenhouse Cucumber. J. Appl. Phycol. 2011, 23, 353–361. [Google Scholar] [CrossRef]
- Ali, N.; Farrell, A.; Ramsubhag, A.; Jayaraman, J. The Effect of Ascophyllum Nodosum Extract on the Growth, Yield and Fruit Quality of Tomato Grown under Tropical Conditions. J. Appl. Phycol. 2016, 28, 1353–1362. [Google Scholar] [CrossRef]
- Ali, O.; Ramsubhag, A.; Jayaraman, J. Biostimulatory Activities of Ascophyllum Nodosum Extract in Tomato and Sweet Pepper Crops in a Tropical Environment. PLoS ONE 2019, 14, e0216710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, W.; Rayirath, U.P.; Subramanian, S.; Jithesh, M.N.; Rayorath, P.; Hodges, D.M.; Critchley, A.T.; Craigie, J.S.; Norrie, J.; Prithiviraj, B. Seaweed Extracts as Biostimulants of Plant Growth and Development. J. Plant Growth Regul. 2009, 28, 386–399. [Google Scholar] [CrossRef]
- Craigie, J.S. Seaweed Extract Stimuli in Plant Science and Agriculture. J. Appl. Phycol. 2011, 23, 371–393. [Google Scholar] [CrossRef]
- Considine, D.M.; Considine, G.D. Foods and Food Production Encyclopedia; Springer: New York, NY, USA, 1982. [Google Scholar] [CrossRef]
- Agarwal, P.K.; Dangariya, M.; Agarwal, P. Seaweed Extracts: Potential Biodegradable, Environmentally Friendly Resources for Regulating Plant Defence. Algal Res. 2021, 58, 102363. [Google Scholar] [CrossRef]
- Rodrigues, B.; Morais, T.P.; Zaini, P.A.; Campos, C.S.; Almeida-Souza, H.O.; Dandekar, A.M.; Nascimento, R.; Goulart, L.R. Antimicrobial Activity of Epsilon-Poly-ι-lysine against Phytopathogenic Bacteria. Sci. Rep. 2020, 10, 11324. [Google Scholar] [CrossRef]
- Jha, Y.; Subramanian, R.B.; Sahoo, S. Antifungal potential of fenugreek coriander, mint, spinach herbs extracts against Aspergillus niger and Pseudomonas aeruginosa phyto-pathogenic fungi. Allelopath. J. 2014, 34, 325–334. [Google Scholar]
- Mansfield, J.; Genin, S.; Magori, S.; Citovsky, V.; Sriariyanum, M.; Ronald, P.; Dow, M.; Verdier, V.; Beer, S.V.; Machado, M.A.; et al. Top 10 Plant Pathogenic Bacteria in Molecular Plant Pathology. Mol. Plant Pathol. 2012, 13, 614–629. [Google Scholar] [CrossRef] [Green Version]
- Teper, D.; Pandey, S.S.; Wang, N. The HrpG/HrpX Regulon of Xanthomonads—An Insight to the Complexity of Regulation of Virulence Traits in Phytopathogenic Bacteria. Microorganisms 2021, 9, 187. [Google Scholar] [CrossRef] [PubMed]
- Schuster, M.L.; Coyne, D.P. Survival Mechanisms of Phytopathogenic Bacteria. Annu. Rev. Phytopathol. 1974, 12, 199–221. [Google Scholar] [CrossRef]
- Carlson, R.R.; Vidaver, A.K. Taxonomy of Corynebacterium Plant Pathogens, Including a New Pathogen of Wheat, Based on Polyacrylamide Gel Electrophoresis of Cellular Proteins? Int. J. Syst. Bacteriol. 1982, 32, 315–326. [Google Scholar] [CrossRef] [Green Version]
- Thimann, K.V.; Sachs, T. The Role of Cytokinins in the “Fasciation” Disease Caused by Corynebacterium fascians. Am. J. Bot. 1966, 53, 731–739. [Google Scholar] [CrossRef]
- Naher, U.A.; Othman, R.; Shamsuddin, Z.H.J.; Saud, H.M.; Ismail, M.R. Growth Enhancement and Root Colonization of Rice Seedlings by Rhizobium and Corynebacterium spp. Int. J. Agric. Biol. 2009, 11, 586–590. [Google Scholar]
- Fett, W.F.; Dunn, M.F. Exopolysaccharides Produced by Phytopathogenic Pseudomonas syringae Pathovars in Infected Leaves of Susceptible Hosts. Plant Physiol. 1989, 89, 5–9. [Google Scholar] [CrossRef] [Green Version]
- De Vos, P.; Goor, M.; Gillis, M.; De Ley, J. Ribosomal Ribonucleic Acid Cistron Similarities of Phytopathogenic Pseudomonas Species. Int. J. Syst. Bacteriol. 1985, 35, 169–184. [Google Scholar] [CrossRef] [Green Version]
- Esserti, S.; Smaili, A.; Makroum, K.; Belfaiza, M.; Rifai, L.A.; Koussa, T.; Kasmi, I.; Faize, M. Priming of Nicotiana benthamiana antioxidant defences using brown seaweed extracts. J. Phytopathol. 2018, 166, 86–94. [Google Scholar] [CrossRef]
- Caleya, R.F.; Gonzalez-Pascual, B.; García-Olmedo, F.; Carbonero, P. Susceptibility of Phytopathogenic Bacteria to Wheat Purothionins In Vitro. Appl. Microbiol. 1972, 23, 998–1000. [Google Scholar] [CrossRef]
- Subramoni, S.; Nathoo, N.; Klimov, E.; Yuan, Z.C. Agrobacterium tumefaciens responses to plant-derived signaling molecules. Front. Plant Sci. 2014, 5, 322. [Google Scholar] [CrossRef] [Green Version]
- McCullen, C.A.; Binns, A.N. Agrobacterium tumefaciens and Plant Cell Interactions and Activities Required for Interkingdom Macromolecular Transfer. Annu. Rev. Cell Dev. Biol. 2006, 22, 101–127. [Google Scholar] [CrossRef] [Green Version]
- De Cleene, M.; De Ley, J. The Host Range of Crown Gall. Bot. Rev. 1976, 42, 389–466. [Google Scholar] [CrossRef]
- Wattana-Amorn, P.; Charoenwongsa, W.; Williams, C.; Crump, M.P.; Apichaisataienchote, B. Antibacterial Activity of Cyclo (L-Pro-L-Tyr) and Cyclo (D-Pro-L-Tyr) from Streptomyces Sp. Strain 22-4 against Phytopathogenic Bacteria. Nat. Prod. Res. 2016, 30, 1980–1983. [Google Scholar] [CrossRef] [Green Version]
- Louws, F.J.; Bell, J.; Medina-Mora, C.M.; Smart, C.D.; Opgenorth, D.; Ishimaru, C.A.; Hausbeck, M.K.; De Bruijn, F.J.; Fulbright, D.W. Rep-PCR-Mediated Genomic Fingerprinting: A Rapid and Effective Method to Identify Clavibacter michiganensis. Phytopathology 1998, 88, 862–868. [Google Scholar] [CrossRef] [Green Version]
- Elia, S.; Gosselé, F.; Vantomme, R.; Swings, J.; Ley, J.D. Corynebacterium fascians: Phytopathogenicity and Numerical Analysis of Phenotypic Features. J. Phytopathol. 1984, 110, 89–105. [Google Scholar] [CrossRef]
- Thyr, B.D.; Samuel, M.J.; Brown, P.G. New solanaceous host records for Corynebacterium michiganense. Plant Dis. Rep. 1975, 59, 595–598. [Google Scholar]
- Wallis, F.M. Ultrastructural histopathology of tomato plants infected with Corynebacferium michiganense. Physiol. Plant Pathol. 1977, 11, 333–342. [Google Scholar] [CrossRef]
- Van Vaerenbergh, J.P.C.; Chauveau, J.F. Detection of Corynebacterium michiganense in tomato seed lots. EPPO Bull. 1987, 17, 131–138. [Google Scholar]
- Spencer, J.F.T.; Gorin, P.A.J. The occurrence in the host plant of physiological active gums produced by Corynebacterium insidiosum and Corynebacterium sepedonicum. Can. J. Microbiol. 1961, 7, 185–188. [Google Scholar] [CrossRef]
- Nelson, G.A. Corynebacterium sepedonicum in potato: Effect of inoculum concentration on ring rot symptoms and latent infection. Can. J. Plant Pathol. 1982, 4, 37–41. [Google Scholar] [CrossRef]
- Collins, M.D.; Jones, D. Reclassification of Corynebacterium flaccumfaciens, Corynebacterium betae, Corynebacterium oorti and Corynebacterium poinsettiae in the genus Curtobacterium, as Curtobacterium flaccumfaciens comb. nov. J. Gen. Microbiol. 1983, 129, 3545–3548. [Google Scholar] [CrossRef]
- Kieu, N.P.; Aznar, A.; Segond, D.; Rigault, M.; Simond-Côte, E.; Kunz, C.; Soulie, M.-C.; Expert, D.; Dellagi, A. Iron deficiency affects plant defence responses and confers resistance to Dickeya dadantii and Botrytis cinerea. Mol. Plant Pathol. 2012, 13, 816–827. [Google Scholar] [CrossRef] [PubMed]
- Dessert, J.M.; Baker, L.R.; Fobes, J.F. Inheritance of reaction to Pseudomonas lachrymans in pickling cucumber. Euphytica 1982, 31, 847–855. [Google Scholar] [CrossRef]
- Caruso, F.L.; Kuć, J. Induced resistance of cucumber to anthracnose and angular leaf spot by Pseudomonas lachrymans and Colletotrichum lagenarium. Physiol. Plant Pathol. 1979, 14, 191–201. [Google Scholar] [CrossRef]
- Keen, N.T.; Williams, P.H. Effect of nutritional factors on extracellular protease production by Pseudomonas lachrymans. Can. J. Microbiol. 1967, 13, 863–871. [Google Scholar] [CrossRef]
- Gardan, L.; Shafik, H.; Belouin, S.; Broch, R.; Grimont, F.; Grimont, P.A.D. DNA relatedness among the pathovars of Pseudomonas syringae and description of Pseudomonas tremae sp. nov. and Pseudomonas cannabina sp. nov. (Ex Sutic and Dowson 1959). Int. J. Syst. Bacteriol. 1999, 49, 469–478. [Google Scholar] [CrossRef] [Green Version]
- Olczak-Woltman, H.; Masny, A.; Bartoszewski, G.; Płucienniczak, A.; Niemirowicz-Szczytt, K. Genetic diversity of Pseudomonas syringae pv. lachrymans strains isolated from cucumber leaves collected in Poland. Plant Pathol. 2007, 56, 373–382. [Google Scholar] [CrossRef]
- Krejzar, V.; Mertelík, J.; Pánková, I.; Kloudová, K.; Kůdela, V. Pseudomonas Marginalis associated with soft rot of Zantedeschia spp. Plant Prot. Sci. 2008, 44, 85–90. [Google Scholar] [CrossRef] [Green Version]
- Gnanamanickam, S. (Ed.) Plant-Associated Bacteria; Springer: Dordrecht, The Netherlands, 2006. [Google Scholar]
- Kůdela, V.; Krejzar, V.; Pánková, I. Pseudomonas Corrugata and Pseudomonas Marginalis associated with the collapse of tomato plants in rockwool slab hydroponic culture. Plant Prot. Sci. 2010, 46, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Hunter, J.E.; Cigna, J.A. Bacterial Blight Incited in Parnsip by Pseudomonas marginalis and Pseudomonas viridiflava. Phytopathology 1981, 71, 1238–1241. [Google Scholar]
- Membré, J.M.; Burlot, P.M. Effects of Temperature, PH, and NaCl on Growth and Pectinolytic Activity of Pseudomonas marginalis. Appl. Environ. Microbiol. 1994, 60, 2017–2022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neale, H.C.; Hulin, M.T.; Harrison, R.J.; Jackson, R.W.; Arnold, D.L. Transposon Mutagenesis of Pseudomonas Syringae Pathovars Syringae and Morsprunorum to Identify Genes Involved in Bacterial Canker Disease of Cherry. Microor 2021, 9, 1328. [Google Scholar] [CrossRef] [PubMed]
- Crosse, J.E.; Garrett, C.M.E. Pathogenicity of Pseudomonas Morsprunorum in Relation to Host Specificity. J. Gen. Microbiol. 1970, 62, 315–327. [Google Scholar] [CrossRef] [Green Version]
- Latorre, B.A.; Jones, A.L. Pseudomonas morsprunorum, the Cause of Bacterial Canker of Sour Cherry in Michigan, and Its Epiphytic Association with P. syringae. Phytopathology 1979, 69, 335–339. [Google Scholar] [CrossRef]
- Gardan, L.; Bollet, C.; Ghorrah, M.A.B.U.; Grimont, F.; Grimont, P.A.D. DNA Relatedness among the Pathovar Strains of Pseudomonas syringae subsp. savastanoi Janse (1982) and Proposal of Pseudomonas savastunoi Sp. Nov. Int. J. Syst. Bacteriol. 1992, 42, 606–612. [Google Scholar] [CrossRef]
- Penyalver, R.; García, A.; Ferrer, A.; Bertolini, E.; López, M.M. Detection of Pseudomonas savastanoi pv. savastanoi in Olive Plants by Enrichment and PCR. Appl. Environ. Microbiol. 2000, 66, 2673–2677. [Google Scholar] [CrossRef] [Green Version]
- Caballo-Ponce, E.; Meng, X.; Uzelac, G.; Halliday, N.; Cámara, M.; Licastro, D.; Silva, D.P.; Ramos, C.; Venturi, V. Quorum Sensing in Pseudomonas savastanoi pv. savastanoi and Erwinia toletana: Role in Virulence and Interspecies Interactions in the Olive Knot. Appl. Environ. Microbiol. 2018, 84, e00950-18. [Google Scholar]
- Yunis, H.; Bashan, Y.; Okon, Y.; Henis, Y. Weather Dependence, Yield Losses, and Control of Bacterial Speck of Tomato Caused by Pseudomonas tomato. Plant Dis. 1980, 64, 937–939. [Google Scholar] [CrossRef]
- Preston, G.M. Pseudomonas syringae pv. tomato: The right pathogen, of the right plant, at the right time. Mol. Plant Pathol. 2000, 1, 263–275. [Google Scholar] [CrossRef] [Green Version]
- Genin, S.; Denny, T.P. Pathogenomics of the Ralstonia solanacearum Species Complex. Annu. Rev. Phytopathol. 2012, 50, 67–89. [Google Scholar] [CrossRef]
- Jha, Y. Macrophytes as a potential tool for crop production by providing nutrient as well as protection against common phyto pathogen. Highlights Biosci. 2021, 4, 1–5. [Google Scholar] [CrossRef]
- Prithiviraj, B.; Bais, H.P.; Jha, A.K.; Vivanco, J.M. Staphylococcus aureus pathogenicity on Arabidopsis thaliana is mediated either by a direct effect of salicylic acid on the pathogen or by SA-dependent, NPR1-independent host responses. Plant J. 2005, 42, 417–432. [Google Scholar] [CrossRef] [PubMed]
- Andrade, A.E.; Silva, L.P.; Pereira, J.L.; Noronha, E.F.; Reis, F.B.; Bloch, C.; Dos Santos, M.F.; Domont, G.B.; Franco, O.L.; Mehta, A. In vivo proteome analysis of Xanthomonas campestris pv. campestris in the interaction with the host plant Brassica oleracea. FEMS Microbiol. Lett. 2008, 281, 167–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, Q.; Wang, N. High-Throughput Screening and Analysis of Genes of Xanthomonas citri subsp. citri Involved in Citrus Canker Symptom Development. Mol. Plant-Microbe Interact. 2012, 25, 69–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martins, P.M.M.; Andrade, M.D.O.; Benedetti, C.E.; de Souza, A.A. Xanthomonas citri subsp. citri: Host interaction and control strategies. Trop. Plant Pathol. 2020, 45, 213–236. [Google Scholar] [CrossRef]
- Zhang, Y.; Callaway, E.M.; Jones, J.B.; Wilson, M. Visualisation of hrp Gene Expression in Xanthomonas euvesicatoria in the tomato phyllosphere. Eur. J. Plant Pathol. 2009, 124, 379–390. [Google Scholar] [CrossRef]
- Oliveira-Pinto, P.R.; Mariz-Ponte, N.; Torres, A.; Tavares, F.; Fernandes-Ferreira, M.; Sousa, R.M.; Santos, C. Satureja montana L. essential oil, montmorillonite and nanoformulation reduce Xanthomonas euvesicatoria infection, modulating redox and hormonal pathways of tomato plants. Sci. Hortic. 2022, 295, 110861. [Google Scholar] [CrossRef]
- Santos, L.V.S.; Melo, E.A.; Silva, A.M.F.; Félix, K.C.S.; Quezado-Duval, A.M.; Albuquerque, G.M.R.; Gama, M.A.S.; Souza, E.B. Weeds as Alternate Hosts of Xanthomonas euvesicatoria pv. euvesicatoria and X. campestris pv. campestris in vegetable-growing fields in the state of Pernambuco, Brazil. Trop. Plant Pathol. 2020, 45, 484–492. [Google Scholar] [CrossRef]
- Shen, Y.; Ronald, P. Molecular Determinants of Disease and Resistance in Interactions of Xanthomonas oryzae pv. oryzae and Rice. Microbes Infect. 2002, 4, 1361–1367. [Google Scholar] [CrossRef] [Green Version]
- Niño-Liu, D.O.; Ronald, P.C.; Bogdanove, A.J. Xanthomonas oryzae pathovars: Model pathogens of a model crop. Mol. Plant Pathol. 2006, 7, 303–324. [Google Scholar] [CrossRef]
- Karavina, C.; Mandumbu, R.; Parwada, C.; Zivenge, E. Epiphytic Survival of Xanthomonas axonopodis pv. phaseoli (E. F. SM). J. Anim. Plant Sci. 2011, 9, 1161–1168. [Google Scholar]
- Azevedo, J.L.; Araújo, W.L.; Lacava, P.T. The diversity of citrus endophytic bacteria and their interactions with Xylella fastidiosa and host plants. Genet. Mol. Biol. 2016, 39, 476–491. [Google Scholar] [CrossRef] [Green Version]
- Cornara, D.; Cavalieri, V.; Dongiovanni, C.; Altamura, G.; Palmisano, F.; Bosco, D.; Porcelli, F.; Almeida, R.P.P.; Saponari, M. Transmission of Xylella fastidiosa by naturally infected Philaenus spumarius (Hemiptera, Aphrophoridae) to different host plants. J. Appl. Entomol. 2017, 141, 80–87. [Google Scholar] [CrossRef]
- Morán, F.; Barbé, S.; Bastin, S.; Navarro, I.; Bertolini, E.; López, M.M.; Hernández-Suárez, E.; Urbaneja, A.; Tena, A.; Siverio, F.; et al. The Challenge of Environmental Samples for PCR Detection of Phytopathogenic Bacteria: A Case Study of Citrus Huanglongbing Disease. Agronomy 2021, 11, 10. [Google Scholar] [CrossRef]
- Narayanasamy, P. Diagnosis of Bacterial Diseases of Plants. In Microbial Plant Pathogens-Detection and Disease Diagnosis: Bacterial and Phytoplasmal Pathogens; Springer: Dordrecht, The Netherlands, 2011; Volume 2, pp. 233–246. [Google Scholar] [CrossRef]
- Ferrarezi, R.S.; Vincent, C.I.; Urbaneja, A.; Machado, M.A. Editorial: Unravelling Citrus Huanglongbing Disease. Front. Plant Sci. 2020, 11, 10–12. [Google Scholar] [CrossRef]
- Bové, J.M. Huanglongbing: A destructive, newly-emerging, century-old disease of citrus. J. Plant Pathol. 2006, 88, 7–37. [Google Scholar]
- Graça, J.V.; Korsten, L. Citrus Huanglongbing: Review, Present Status and Future Strategies. In Diseases of Fruits and Vegetables Volume I; Springer: Dordrecht, The Netherland, 2004; Volume 1, pp. 229–245. [Google Scholar] [CrossRef]
- Planet, P.; Jagoueix, S.; Bové, J.M.; Garnier, M. Detection and Characterization of the African Citrus Greening Liberobacter by Amplification, Cloning, and Sequencing of the rplKAJL-rpoBC Operon. Curr. Microbiol. 1995, 30, 137–141. [Google Scholar] [CrossRef]
- Jagoueix, S.; Bové, J.M.; Garnier, M. PCR detection of the two ‘Candidatus’ liberobacter species associated with greening disease of citrus. Mol. Cell. Probes 1996, 10, 43–50. [Google Scholar] [CrossRef]
- Do Teixeira, D.C.; Saillard, C.; Eveillard, S.; Danet, J.L.; Costa, P.I.; Ayres, A.J.; Bové, J. ‘Candidatus Liberibacter americanus’, associated with citrus huanglongbing (greening disease) in São Paulo State, Brazil. Int. J. Syst. Evol. Microbiol. 2005, 55, 1857–1862. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Rodríguez, J.; Krüger, K.; Pérez-Hedo, M.; Ruíz-Rivero, O.; Urbaneja, A.; Tena, A. Classical biological control of the African citrus psyllid Trioza erytreae, a major threat to the European citrus industry. Sci. Rep. 2019, 9, 9440. [Google Scholar] [CrossRef] [Green Version]
- Urbaneja-Bernat, P.; Pérez-Rodríguez, J.; Krüger, K.; Catalán, J.; Rizza, R.; Hernández-Suárez, E.; Urbaneja, A.; Tena, A. Host range testing of Tamarixia dryi (Hymenoptera: Eulophidae) sourced from South Africa for classical biological control of Trioza erytreae (Hemiptera: Psyllidae) in Europe. Biol. Control 2019, 135, 110–116. [Google Scholar] [CrossRef]
- Lakhdar, F.; Boujaber, N.; Oumaskour, K.; Assobhei, O.; Etahiri, S. Inhibitive Activity of 17 marine algae from the coast of El Jadida-Morocco against Erwinia chrysanthemi. Int. J. Pharm. Pharm. Sci. 2015, 7, 376–380. [Google Scholar]
- Kumar, K.A.; Rengasamy, R. Evaluation of Antibacterial Potential of Seaweeds Occurring along the Coast of Tamil Nadu, India against the Plant Pathogenic Bacterium Xanthomonas oryzae pv. oryzae (Ishiyama) Dye. Bot. Mar. 2000, 43, 409–415. [Google Scholar]
- Esserti, S.; Smaili, A.; Rifai, L.A.; Koussa, T.; Makroum, K.; Belfaiza, M.; Kabil, E.M.; Faize, L.; Burgos, L.; Alburquerque, N.; et al. Protective effect of three brown seaweed extracts against fungal and bacterial diseases of tomato. J. Appl. Phycol. 2017, 29, 1081–1093. [Google Scholar] [CrossRef]
- Shah, Z.; Badshah, S.L.; Iqbal, A.; Shah, Z.; Emwas, A.-H.; Jaremko, M. Investigation of important biochemical compounds from selected freshwater macroalgae and their role in agriculture. Chem. Biol. Technol. Agric. 2022, 9, 9. [Google Scholar] [CrossRef]
- Jiménez, E.; Dorta, F.; Medina, C.; Ramírez, A.; Ramírez, I.; Peña-Cortés, H. Anti-Phytopathogenic Activities of Macro-Algae Extracts. Mar. Drugs 2011, 9, 739–756. [Google Scholar] [CrossRef] [Green Version]
- Paulert, R.; Júnior, A.S.; Stadnik, M.J.; Pizzolatti, M.G. Antimicrobial properties of extracts from the green seaweed Ulva fasciata Delile against pathogenic bacteria and fungi. Arch. Hydrobiol. Suppl. Algol. Stud. 2007, 123, 123–130. [Google Scholar] [CrossRef]
- Kumar, K.A.; Rengasamy, R. Antibacterial Activities of Seaweed Extracts/Fractions Obtained through a TLC Profile against the Phytopathogenic Bacterium Xanthomonas oryzae pv. oryzae. Bot. Mar. 2000, 43, 417–421. [Google Scholar]
- Arunkumar, K.; Selvapalam, N.; Rengasamy, R. The antibacterial compound sulphoglycerolipid 1-0 palmitoyl-3-0(6′-sulpho-alpha-quinovopyranosyl)-glycerol from Sargassum wightii Greville (Phaeophyceae). Bot. Mar. 2005, 48, 441–445. [Google Scholar] [CrossRef]
- Robles-Centeno, P.O.; Ballantine, D.L.; Gerwick, W.H. Dynamics of antibacterial activity in three species of Caribbean marine algae as a function of habitat and life history. Hydrobiologia 1996, 326–327, 457–462. [Google Scholar] [CrossRef]
- Ballantine, D.L.; Gerwick, W.H.; Velez, S.M.; Alexander, E.; Guevara, P. Antibiotic activity of lipid-soluble extracts from Caribbean marine algae. Hydrobiologia 1987, 151–152, 463–469. [Google Scholar] [CrossRef]
- Rao, P.P.S.; Rao, P.S.; Karmarkar, S.M. Antibacterial Substances from Brown Algae II. Efficiency of Solvents in the Evaluation of Antibacterial Substances from Sargassum Johnstonii Setchell et Gardner. Bot. Mar. 1986, XXIX, 503–507. [Google Scholar]
- Kamenarska, Z.; Serkedjieva, J.; Najdenski, H.; Stefanov, K.; Tsvetkova, I.; Dimitrova-Konaklieva, S.; Popov, S. Antibacterial, antiviral, and cytotoxic activities of some red and brown seaweeds from the Black Sea. Bot. Mar. 2009, 52, 80–86. [Google Scholar] [CrossRef]
- Caccamese, S.; Azzolina, R.; Furnari, G.; Cormaci, M.; Grasso, S. Antimicrobial and Antiviral Activities of Some Marine Algae from Eastern Sicily. Bot. Mar. 1981, XXIV, 365–367. [Google Scholar] [CrossRef]
- Rao, P.S.; Parekh, K.S. Antibacterial Activity of Indian Seaweed Extracts. Bot. Mar. 1981, XXIV, 577–582. [Google Scholar] [CrossRef]
- Akira, S.; Uematsu, S.; Takeuchi, O. Pathogen Recognition and Innate Immunity. Cell 2006, 124, 783–801. [Google Scholar] [CrossRef] [Green Version]
- Delepelaire, P.; Wandersman, C. Characterization, localization and transmembrane organization of the three proteins PrtD, PrtE and PrtF necessary for protease secretion by the Gram-negative bacterium Erwinia chrysanthemi. Mol. Microbiol. 1991, 5, 2427–2434. [Google Scholar] [CrossRef]
- Lyon, G.J.; Novick, R.P. Peptide Signaling in Staphylococcus aureus and other Gram-positive bacteria. Peptides 2004, 25, 1389–1403. [Google Scholar] [CrossRef]
- Paterson, D.L. Resistance in Gram-negative bacteria: Enterobacteriaceae. Am. J. Infect. Control 2006, 34, 20–28. [Google Scholar] [CrossRef]
- Lopes, G.; Sousa, C.; Silva, L.R.; Pinto, E.; Andrade, P.B.; Bernando, J.; Mouga, T.; Valentão, P. Can Phlorotannins Purified Extracts Constitute a Novel Pharmacological Alternative for Microbial Infections with Associated Inflammatory Conditions? PLoS ONE 2012, 7, e31145. [Google Scholar] [CrossRef]
- Jörgensen, N.O.G.; Stepanaukas, R.; Pedersen, A.-G.U.; Hansen, M.; Nybroe, O. Occurrence and degradation of peptidoglycan in aquatic environments. FEMS Microbiol. Ecol. 2003, 46, 269–280. [Google Scholar] [CrossRef]
- Kamenarska, Z.; Ivanova, A.; Stancheva, R.; Stefanov, K.; Dimitrova-Konaklieva, S.; Popov, S. Polar constituents of some Black Sea red and brown algae and their application into chemotaxonomy and chemoevolution. Arch. Hydrobiol. Suppl. Algol. Stud. 2006, 119, 139–154. [Google Scholar] [CrossRef]
- Prabha, V.; Prakash, D.J.; Sudha, P.N. Analysis of bioactive compounds and antimicrobial activity of marine algae Kappaphycus alvarezii using three solvent extracts. Int. J. Pharm. Sci. Res. 2013, 4, 306–310. [Google Scholar]
- Santos, S.A.O.; Trindade, S.S.; Oliveira, C.S.D.; Parreira, P.; Rosa, D.; Duarte, M.F.; Ferreira, I.; Cruz, M.T.; Rego, A.M.; Abreu, M.H.; et al. Lipophilic Fraction of Cultivated Bifurcaria bifurcata R. Ross: Detailed Composition and In Vitro Prospection of Current Challenging Bioactive Properties. Mar. Drugs 2017, 15, 340. [Google Scholar] [CrossRef] [Green Version]
- Kanias, G.D.; Skaltsa, H.; Tsitsa, E.; Loukis, A.; Bitis, J. Study of the correlation between trace elements, sterols and fatty acids in brown algae from the Saronikos Gulf of Greece. Fresenius’ J. Anal. Chem. 1992, 344, 334–339. [Google Scholar] [CrossRef]
- Son, B.W. Glycolipids from Gracilaria verrucosa. Phytochemistry 1990, 29, 307–309. [Google Scholar] [CrossRef]
- Siddhanta, A.K.; Ramavat, B.K.; Chauhan, V.D.; Achari, B.; Dutta, P.K.; Pakrashi, S.C. Sulphonoglycolipid from the Green Alga Enteromorpha flexuosa (Wulf.) J. Ag. Bot. Mar. 1991, 34, 365–367. [Google Scholar] [CrossRef]
- Fusetani, N.; Hashimoto, Y. Structures of Two Water Soluble Hemolysins Isolated from the Green Alga Ulva pertusa. Agric. Biol. Chem. 1975, 39, 2021–2025. [Google Scholar] [CrossRef] [Green Version]
- Araki, S.; Sakurai, T.; Oohusa, T.; Kayama, M.; Sato, N. Characterization of Sulfoquinovosyl Diacylglycerol from Marine Red Algae. Plant Cell Physiol. 1989, 30, 775–781. [Google Scholar]
- Barchan, A.; Bakkali, M.; Arakrak, A.; Pagán, R.; Laglaoui, A. The effects of solvents polarity on the phenolic contents and antioxidant activity of three Mentha species extracts. Int. J. Curr. Microbiol. Appl. Sci. 2014, 3, 399–412. [Google Scholar]
- Mcgaw, L.J.; Jäger, A.K.; Van Staden, J. Antibacterial effects of fatty acids and related compounds from plants. S. Afr. J. Bot. 2002, 68, 417–423. [Google Scholar] [CrossRef] [Green Version]
- Nwabueze, T.U.; Okocha, K.S. Extraction performances of polar and non-polar solvents on the physical and chemical indices of African breadfruit (Treculia africana) seed oil. Afr. J. Food Sci. 2008, 2, 119–125. [Google Scholar]
- Wang, Y.; Xu, Z.; Bach, S.J.; McAllister, T.A. Sensitivity of Escherichia Coli to Seaweed (Ascophyllum Nodosum) Phlorotannins and Terrestrial Tannins. Asian-Australas. J. Anim. Sci. 2009, 22, 238–245. [Google Scholar] [CrossRef]
- Malini, M.; Ponnanikajamideen, M.; Malarkodi, C.; Rajeshkumar, S. Explore the Antimicrobial Potential from Organic Solvents Extract of Brown Seaweed (Sargassum Longifolium) Alleviating to Pharmaceuticals. Int. J. Pharm. Res. 2014, 6, 28–35. [Google Scholar]
- Capillo, G.; Savoca, S.; Costa, R.; Sanfilippo, M.; Rizzo, C.; Lo Giudice, A.; Albergamo, A.; Rando, R.; Bartolomeu, G.; Spanò, N.; et al. New Insights into the Culture Method and Antibacterial Potential of Gracilaria gracilis. Mar. Drugs 2018, 16, 492. [Google Scholar] [CrossRef]
- Bharath, B.; Pavithra, A.N.; Divya, A.; Perinbam, K. Chemical Composition of Ethanolic Extracts from Some Seaweed Species of the South Indian Coastal Zone, Their Antibacterial and Membrane-Stabilizing Activity. Russ. J. Mar. Biol. 2020, 46, 370–378. [Google Scholar] [CrossRef]
- Coronado, A.S.; Dionisio-Sese, M.L. Antimicrobial Property of Crude Ethanolic Extract from Sargassum crassifolium. Asian J. Microbiol. Biotechnol. Environ. Sci. 2014, 16, 471–474. [Google Scholar]
- Susilo, B.; Rohim, A.; Wahyu, M.L. Serial Extraction Technique of Rich Antibacterial Compounds in Sargassum Cristaefolium Using Different Solvents and Testing Their Activity. Curr. Bioact. Compd. 2022, 18, 18–25. [Google Scholar] [CrossRef]
- Prakash, S.; Ahila, N.K.; Ramkumar, V.S.; Ravindran, J.; Kannapiran, E. Antimicrofouling Properties of Chosen Marine Plants: An Eco-Friendly Approach to Restrain Marine Microfoulers. Biocatal. Agric. Biotechnol. 2015, 4, 114–121. [Google Scholar] [CrossRef]
- Nofal, A.; Azzazy, M.; Ayyad, S.; Abdelsalm, E.; Abousekken, M.S.; Tammam, O. Evaluation of the Brown Alga, Sargassum Muticum Extract as an Antimicrobial and Feeding Additives. Braz. J. Biol. 2022, 84, 1–9. [Google Scholar] [CrossRef]
- Banu, A.T.; Ramani, P.S.; Murugan, A. Effect of Seaweed Coating on Quality Characteristics and Shelf Life of Tomato (Lycopersicon Esculentum Mill). Food Sci. Hum. Wellness 2020, 9, 176–183. [Google Scholar] [CrossRef]
- Kumar, P.; Selvi, S.S.; Prabha, A.L.; Kumar, K.P.; Ganeshkumar, R.S.; Govindaraju, M. Synthesis of Silver Nanoparticles from Sargassum Tenerrimum and Screening Phytochemicals for Its Antibacterial Activity. Nano Biomed. Eng. 2012, 4, 12–16. [Google Scholar] [CrossRef]
- Albratty, M.; Alhazmi, H.A.; Meraya, A.M.; Najmi, A.; Alam, M.S.; Rehman, Z.; Moni, S.S. Spectral Analysis and Antibacterial Activity of the Bioactive Principles of Sargassum Tenerrimum J. Agardh Collected from the Red Sea, Jazan, Kingdom of Saudi Arabia. Braz. J. Biol. 2023, 83, 1–10. [Google Scholar] [CrossRef]
- Eahamban, K.; Antonisamy, J.M. Preliminary Phytochemical, UV-VIS, HPLC and Anti-Bacterial Studies on Gracilaria Corticata J. Ag. Asian Pac. J. Trop. Biomed. 2012, 2, S568–S574. [Google Scholar] [CrossRef]
- Radhika, D.; Mohaideen, A. Fourier Transform Infrared Analysis of Ulva Lactuca and Gracilaria Corticata and Their Effect on Antibacterial Activity. Asian J. Pharm. Clin. Res. 2015, 8, 209–212. [Google Scholar]
- Zheng, C.J.; Yoo, J.-S.; Lee, T.-G.; Cho, H.-Y.; Kim, Y.-H.; Kim, W.-G. Fatty Acid Synthesis Is a Target for Antibacterial Activity of Unsaturated Fatty Acids. FEBS Lett. 2005, 579, 5157–5162. [Google Scholar] [CrossRef] [Green Version]
- Pierre, G.; Sopena, V.; Juin, C.; Mastouri, A.; Graber, M.; Maugard, T. Antibacterial Activity of a Sulfated Galactan Extracted from the Marine Alga Chaetomorpha aerea Against Staphylococcus aureus. Biotechnol. Bioprocess Eng. 2011, 16, 937–945. [Google Scholar] [CrossRef]
- He, F.; Yang, Y.; Yang, G.; Yu, L. Studies on Antibacterial Activity and Antibacterial Mechanism of a Novel Polysaccharide from Streptomyces Virginia H03. Food Control 2010, 21, 1257–1262. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Haney, E.F.; Vogel, H.J. The Expanding Scope of Antimicrobial Peptide Structures and Their Modes of Action. Trends Biotechnol. 2011, 29, 464–472. [Google Scholar] [CrossRef]
- Hughes, C.C.; Fenical, W. Antibacterials from the Sea. Chem. Eur. J. 2010, 16, 12512–12525. [Google Scholar] [CrossRef] [Green Version]
- Etahiri, S.; Valérie, B.-P.; Caux, C.; Guyot, M. New Bromoditerpenes from the Red Alga Sphaerococcus coronopifolius. J. Nat. Prod. 2001, 64, 1024–1027. [Google Scholar] [CrossRef]
- Lee, J.-H.; Eom, S.-H.; Lee, E.-H.; Jung, Y.-J.; Kim, H.-J.; Jo, M.-R.; Son, K.-T.; Lee, H.-J.; Kim, J.H.; Lee, M.-S.; et al. In Vitro Antibacterial and Synergistic Effect of Phlorotannins Isolated from Edible Brown Seaweed Eisenia bicyclis against Acne-Related Bacteria. Algae 2014, 29, 47–55. [Google Scholar] [CrossRef]
- Subramanian, S.; Sangha, J.S.; Gray, B.A.; Singh, R.P.; Hiltz, D.; Critchley, A.T.; Prithiviraj, B. Extracts of the marine brown macroalga, Ascophyllum nodosum, Induce Jasmonic Acid Dependent Systemic Resistance in Arabidopsis thaliana against Pseudomonas syringae pv. tomato DC3000 and Sclerotinia sclerotiorum. Eur. J. Plant Pathol. 2011, 131, 237–248. [Google Scholar] [CrossRef]
- Mauch-Mani, B.; Baccelli, I.; Luna, E.; Flors, V. Defense Priming: An Adaptive Part of Induced Resistance. Annu. Rev. Plant Biol. 2017, 68, 485–512. [Google Scholar] [CrossRef] [Green Version]
- Wong, P.T.W. Cross-protection against the wheat and oat take-all fungi by Gaeumannomyces graminis var. graminis. Soil Biol. Biochem. 1975, 7, 189–194. [Google Scholar] [CrossRef]
- Grant, M.; Lamb, C. Systemic Immunity. Curr. Opin. Plant Biol. 2006, 9, 414–420. [Google Scholar] [CrossRef]
- Choudhary, D.K.; Prakash, A.; Johri, B.N. Induced systemic resistance (ISR) in plants: Mechanism of action. Indian J. Microbiol. 2007, 47, 289–297. [Google Scholar] [CrossRef]
- Vidhyasekaran, P. Mycorrhiza induced resistance, a mechanism for management of crop diseases. Curr. Trends Mycorrhizal Res. 1990. [Google Scholar]
- Pozo, M.J.; Azcón-Aguilar, C. Unraveling mycorrhiza-induced resistance. Curr. Opin. Plant Biol. 2007, 10, 393–398. [Google Scholar] [CrossRef]
- Jung, S.C.; Martinez-Medina, A.; Lopez-Raez, J.A.; Pozo, M.J. Mycorrhiza-Induced Resistance and Priming of Plant Defenses. J. Chem. Ecol. 2012, 38, 651–664. [Google Scholar] [CrossRef]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis, 3rd ed.; Academic Press: New York, NY, USA, 2008; p. 694. [Google Scholar] [CrossRef]
- Liu, H.; Able, A.J.; Able, J.A. Priming crops for the future: Rewiring stress memory. Trends Plant Sci. 2021, 27, 699–716. [Google Scholar] [CrossRef] [PubMed]
- Conrath, U.; Thulke, O.; Katz, V.; Schwindling, S.; Kohler, A. Priming as a mechanism in induced systemic resistance of plants. Eur. J. Plant Pathol. 2001, 107, 113–119. [Google Scholar] [CrossRef]
- Fan, Y.; Ma, C.; Huang, Z.; Abid, M.; Jiang, S.; Dai, T.; Zhang, W.; Ma, S.; Jiang, D.; Han, X. Heat Priming During Early Reproductive Stages Enhances Thermo-Tolerance to Post-Anthesis Heat Stress via Improving Photosynthesis and Plant Productivity in Winter Wheat (Triticum aestivum L.). Front. Plant Sci. 2018, 9, 805. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Wang, X.; Zhang, J.; Huang, M.; Cai, J.; Zhou, Q.; Dai, T.; Jiang, D. Salicylic acid and cold priming induce late-spring freezing tolerance by maintaining cellular redox homeostasis and protecting photosynthetic apparatus in wheat. Plant Growth Regul. 2020, 90, 109–121. [Google Scholar] [CrossRef]
- Thakur, A.; Sharma, K.D.; Siddique, K.H.M.; Nayyar, H. Cold priming the chickpea seeds imparts reproductive cold tolerance by reprogramming the turnover of carbohydrates, osmo-protectants and redox components in leaves. Sci. Hortic. 2020, 261, 108929. [Google Scholar] [CrossRef]
- Wojtaszek, P. Oxidative burst: An early plant response to pathogen infection. Biochem. J. 1997, 322, 681–692. [Google Scholar] [CrossRef] [Green Version]
- Olson, P.D.; Varner, J.E. Hydrogen peroxide and lignification. Plant J. 1993, 4, 887–892. [Google Scholar] [CrossRef]
- Alvarez, E.M.; Pennell, R.I.; Meijer, P.-J.; Ishikawa, A.; Dixon, R.A.; Lamb, C. Reactive Oxygen Intermediates Mediate a Systemic Signal Network in the Establishment of Plant Immunity. Cell 1998, 92, 773–784. [Google Scholar] [CrossRef] [Green Version]
- Bruce, T.J.A.; Matthes, M.C.; Napier, J.A.; Pickett, J.A. Stressful “memories” of plants: Evidence and possible mechanisms. Plant Sci. 2007, 173, 603–608. [Google Scholar] [CrossRef]
- Zhang, X. The Epigenetic Landscape of Plants. Science 2008, 320, 489–492. [Google Scholar] [CrossRef]
- Wang, X.; Xin, C.; Cai, J.; Zhou, Q.; Dai, T.; Cao, W.; Jiang, D. Heat Priming Induces Trans-generational Tolerance to High Temperature Stress in Wheat. Front. Plant Sci. 2016, 7, 501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwasaki, M.; Paszkowski, J. Epigenetic memory in plants. EMBO J. 2014, 33, 1987–1998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oberkofler, V.; Pratx, L.; Bäurle, I. Epigenetic regulation of abiotic stress memory: Maintaining the good things while they last. Curr. Opin. Plant Biol. 2021, 61, 102007. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; He, Z. Small DNA Methylation, Big Player in Plant Abiotic Stress Responses and Memory. Front. Plant Sci. 2020, 11, 595603. [Google Scholar] [CrossRef]
- Lang-Mladek, C.; Popova, O.; Kiok, K.; Berlinger, M.; Rakic, B.; Aufsatz, W.; Jonak, C.; Hauser, M.-T.; Lusching, C. Transgenerational Inheritance and Resetting of Stress-Induced Loss of Epigenetic Gene Silencing in Arabidopsis. Mol. Plant 2010, 3, 594–602. [Google Scholar] [CrossRef]
- Avramova, Z. Transcriptional ‘memory’ of a stress: Transient chromatin and memory (epigenetic) marks at stress-response genes. Plant J. 2015, 83, 149–159. [Google Scholar] [CrossRef]
- Lämke, J.; Brzezinka, K.; Altmann, S.; Bäurle, I. A hit-and-run heat shock factor governs sustained histone methylation and transcriptional stress memory. EMBO J. 2016, 35, 162–175. [Google Scholar] [CrossRef]
- Hu, T.; Jin, Y.; Li, H.; Amombo, E.; Fu, J. Stress memory induced transcriptional and metabolic changes of perennial ryegrass (Lolium perenne) in response to salt stress. Physiol. Plant. 2016, 156, 54–69. [Google Scholar] [CrossRef]
- Yakovlev, I.A.; Fossdal, C.G. In Silico Analysis of Small RNAs Suggest Roles for Novel and Conserved miRNAs in the Formation of Epigenetic Memory in Somatic Embryos of Norway Spruce. Front. Physiol. 2017, 8, 674. [Google Scholar] [CrossRef]
- Weeraddana, C.D.S.; Kandasamy, S.; Cutler, G.C.; Shukla, P.S.; Critchley, A.T.; Prithiviraj, B. An alkali-extracted biostimulant prepared from Ascophyllum nodosum alters the susceptibility of Arabidopsis thaliana to the green peach aphid. J. Appl. Phycol. 2021, 33, 3319–3329. [Google Scholar] [CrossRef]
- Stirk, W.A.; Rengasamy, K.R.R.; Kulkarni, M.G.; van Staden, J. Plant Biostimulants from Seaweed: An Overview. In The Chemical Biology of Plant Biostimulants; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2020; pp. 33–56. [Google Scholar] [CrossRef]
- Baltazar, M.; Correia, S.; Guinan, K.J.; Sujeeth, N.; Bragança, R.; Gonçalves, B. Recent Advances in the Molecular Effects of Biostimulants in Plants: An Overview. Biomolecules 2021, 11, 1096. [Google Scholar] [CrossRef] [PubMed]
- Blunden, G.; Wildgoose, P.B. The Effects of Aqueous Seaweed Extract and Kinetin on Potato Yields. J. Sci. Food Agric. 1977, 28, 121–125. [Google Scholar] [CrossRef]
- Arman, M.; Qader, S.A.U. Structural analysis of kappa-carrageenan isolated from Hypnea musciformis (red algae) and evaluation as an elicitor of plant defense mechanism. Carbohydr. Polym. 2012, 88, 1264–1271. [Google Scholar] [CrossRef]
- Radman, R.; Saez, T.; Bucke, C.; Keshavarz, T. Elicitation of plants and microbial cell systems. Biotechnol. Appl. Biochem. 2003, 27, 91–102. [Google Scholar] [CrossRef] [PubMed]
- Nürnberger, T. Signal perception in plant pathogen defense. Cell. Mol. Life Sci. 1999, 55, 167–182. [Google Scholar] [CrossRef]
- Walters, D.; Walsh, D.; Newton, A.; Lyon, G. Induced Resistance for Plant Disease Control: Maximizing the Efficacy of Resistance Elicitors. Phytopathology 2005, 95, 1368–1373. [Google Scholar] [CrossRef]
- Mercier, L.; Lafitte, C.; Borderies, G.; Briand, X.; Esquerré-Tugayé, M.T.; Fournier, J. The algal polysaccharide carrageenans can act as an elicitor of plant defence. New Phytol. 2001, 149, 43–51. [Google Scholar] [CrossRef]
- Shukla, P.S.; Borza, T.; Critchley, A.T.; Prithiviraj, B. Seaweed-Based Compounds and Products for Sustainable Protection against Plant Pathogens. Mar. Drugs 2021, 19, 59. [Google Scholar] [CrossRef]
- Ugarte, R.A.; Sharp, G.; Moore, B. Changes in the brown seaweed Ascophyllum nodosum (L.) Le Jol. plant morphology and biomass produced by cutter rake harvests in southern New Brunswick, Canada. J. Appl. Phycol. 2006, 18, 351–359. [Google Scholar] [CrossRef]
- Jayaraj, J.; Wan, A.; Rahman, M.; Punja, Z.K. Seaweed extract reduces foliar fungal diseases on carrot. Crop Prot. 2008, 27, 1360–1366. [Google Scholar] [CrossRef]
- Cheong, J.-J.; Birberg, W.; Fügedi, P.; Pilotti, A.; Garegg, P.J.; Hong, N.; Ogawa, T.; Hahn, M.G. Structure-Activity Relationships of Oligo-β-glucoside Elicitors of Phytoalexin Accumulation in Soybean. Plant Cell 1991, 3, 127–136. [Google Scholar] [PubMed]
- Miller, K.J.; Hadley, J.A.; Gustine, D. Cyclic β-1,6-1,3-Glucans of Bradyrhizobium japonicum USDA 110 Elicit Isoflavonoid Production in the Soybean (Glycine max) Host. Plant Physiol. 1994, 104, 917–923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tai, A.; Ohsawa, E.; Kawazu, K.; Kobayashi, A. A Minimum Essential Structure of LN-3 Elicitor Activity in Bean Cotyledons. Z. Naturforsch.-Sect. C J. Biosci. 1996, 51, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, A.; Tai, A.; Kanzaki, H.; Kawazu, K. Elicitor-Active Oligosaccharides from Algal Laminaran Stimulate the Production of Antifungal Compounds in Alfalfa. Z. Naturforsch.-Sect. C J. Biosci. 1993, 48, 575–579. [Google Scholar] [CrossRef]
- Takeshi, N. Elicitor Actions of N-Acetylchitooligosaccharides and Laminarioligosaccharides for Chitinase and L-Phenylalanine Ammonia-lyase Induction in Rice Suspension Culture. Biosci. Biotechnol. Biochem. 1997, 61, 975–978. [Google Scholar]
- Nalumpang, S.; Gotoh, Y.; Tsuboi, H.; Gomi, K.; Yamamoto, H.; Akimitsu, K. Functional Characterization of Citrus Polygalacturonase-inhibiting Protein. J. Gen. Plant Pathol. 2002, 68, 118–127. [Google Scholar] [CrossRef]
- Klarzynski, O.; Plesse, B.; Joubert, J.M.; Yvin, J.C.; Kopp, M.; Kloareg, B.; Fritig, B. Linear β-1,3 Glucans Are Elicitors of Defense Responses in Tobacco. Plant Physiol. 2000, 124, 1027–1037. [Google Scholar] [CrossRef]
- Patier, P.; Yvin, J.C.; Kloareg, B.; Liénart, Y.; Rochas, C. Seaweed liquid fertilizer from Ascophyllum nodosum contains elicitors of plant D-glycanases. J. Appl. Phycol. 1993, 5, 343–349. [Google Scholar] [CrossRef]
- Cook, J.; Zhang, J.; Norrie, J.; Blal, B.; Cheng, Z. Seaweed Extract (Stella Maris ®) Activates Innate Immune Responses in Arabidopsis thaliana and Protects Host against Bacterial Pathogens. Mar. Drugs 2018, 16, 221. [Google Scholar] [CrossRef] [Green Version]
- Maury, S.; Geoffroy, P.; Legrand, M. Tobacco O-Methyltransferases Involved in Phenylpropanoid Metabolism. The Different Caffeoyl-Coenzyme A/5-Hydroxyferuloyl-Coenzyme A 3/5-O-Methyltransferase and Caffeic Acid/5-Hydroxyferulic Acid 3/5-O-Methyltransferase Classes Have Distinct Substrate Specificities and Expression Patterns. Plant Physiol. 1999, 121, 215–223. [Google Scholar] [CrossRef] [Green Version]
- Vera, J.; Castro, J.; Gonzalez, A.; Moenne, A. Seaweed Polysaccharides and Derived Oligosaccharides Stimulate Defense Responses and Protection against Pathogens in Plants. Mar. Drugs 2011, 9, 2514–2525. [Google Scholar] [CrossRef] [PubMed]
- Mauch-Mani, B.; Slusarenko, A.J. Production of Salicylic Acid Precursors Is a Major Function of Phenylalanine Ammonia-Lyase in the Resistance of Arabidopsis to Peronospora parasitica. Plant Cell 1996, 8, 203–212. [Google Scholar] [CrossRef] [PubMed]
- Elansary, H.O.; Yessoufou, K.; Shokralla, S.; Mahmoud, E.A.; Skalicka-Wo, K. Enhancing mint and basil oil composition and antibacterial activity using seaweed extracts. Ind. Crops Prod. 2016, 92, 50–56. [Google Scholar] [CrossRef]
- Roy, A.; Ghosh, D.; Kasera, M.; Nori, S.; Vemanna, R.S.; Mohapatra, S.; Narayan, S.S.; Bhattacharjee, S. Kappaphycus alvarezii-derived formulations enhance salicylic acid-mediated anti-bacterial defenses in Arabidopsis thaliana and rice. J. Appl. Phycol. 2022, 34, 679–695. [Google Scholar] [CrossRef]
- Raghavendra, V.B.; Lokesh, S.; Prakash, H.S. Dravya, a Product of Seaweed Extract (Sargassum wightii), Induces Resistance in Cotton against Xanthomonas campestris pv. malvacearum. Phytopathology 2007, 35, 442–449. [Google Scholar] [CrossRef]
- Ali, O.; Ramsubhag, A.; Jayaraman, J. Phytoelicitor activity of Sargassum vulgare and Acanthophora spicifera extracts and their prospects for use in vegetable crops for sustainable crop production. J. Appl. Phycol. 2021, 33, 639–651. [Google Scholar] [CrossRef]
- Ramkissoon, A.; Ramsubhag, A.; Jayaraman, J. Phytoelicitor Activity of Three Caribbean seaweed species on suppression of pathogenic infections in tomato plants. J. Appl. Phycol. 2017, 29, 3235–3244. [Google Scholar] [CrossRef]
- Fertah, M. Isolation and Characterization of Alginate from Seaweed. In Seaweed Polysaccharides; Elsevier: Amsterdam, The Netherlands, 2017; pp. 11–26. [Google Scholar] [CrossRef]
- An, Q.-D.; Zhang, G.-L.; Wu, H.-T.; Zhang, Z.-C.; Zheng, G.-S.; Luan, L.; Murata, Y.; Li, X. alginate-deriving oligosaccharide production by alginase from newly isolated Flavobacterium sp. LXA and its potential application in protection against pathogens. J. Appl. Microbiol. 2009, 106, 161–170. [Google Scholar] [CrossRef]
- Bouissil, S.; El Alaoui-talibi, Z.; Pierre, G.; Michaud, P.; El Modafar, C.; Delattre, C. Use of Alginate Extracted from Moroccan Brown Algae to Stimulate Natural Defense in Date Palm Roots. Molecules 2020, 25, 720. [Google Scholar] [CrossRef] [Green Version]
- Dey, P.; Ramanujam, R.; Venkatesan, G.; Nagarathnam, R. Sodium Alginate Potentiates Antioxidant Defense and PR proteins against early blight disease caused by Alternaria solani in Solanum lycopersicum Linn. PLoS ONE 2019, 14, e0223216. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Howlader, P.; Liu, T.; Sun, X.; Jia, X.; Zhao, X.; Shen, P.; Qin, Y.; Wang, W.; Yin, H. Alginate Oligosaccharide (AOS) induced resistance to Pst DC3000 via salicylic acid-mediated signaling pathway in Arabidopsis thaliana. Carbohydr. Polym. 2019, 225, 115221. [Google Scholar] [CrossRef] [PubMed]
- González, A.; Castro, J.; Vera, J.; Moenne, A. Seaweed Oligosaccharides Stimulate Plant Growth by Enhancing Carbon and Nitrogen Assimilation, Basal Metabolism, and Cell Division. J. Plant Growth Regul. 2013, 32, 443–448. [Google Scholar] [CrossRef] [Green Version]
- Chinchilla, D.; Zipfel, C.; Robatzek, S.; Kemmerling, B.; Nürnberger, T.; Jones, J.D.G.; Felix, G.; Boller, T. A flagellin-induced complex of the receptor FLS2 and BAK1 initiates plant defence. Nature 2007, 448, 497–500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Postel, S.; Küfner, I.; Beuter, C.; Mazzotta, S.; Schwedt, A.; Borlotti, A.; Halter, T.; Kemmerling, B.; Nürnberger, T. The multifunctional leucine-rich repeat receptor kinase BAK1 is implicated in Arabidopsis development and immunity. Eur. J. Cell Biol. 2010, 89, 169–174. [Google Scholar] [CrossRef]
- Kemmerling, B.; Halter, T.; Mazzotta, S.; Mosher, S.; Nürnberger, T. A genome-wide survey for Arabidopsis leucine-rich repeat receptor kinases implicated in plant immunity. Front. Plant Sci. 2011, 2, 88. [Google Scholar] [CrossRef] [Green Version]
- Chandía, N.P.; Matsuhiro, B.; Mejías, E.; Moenne, A. Alginic acids in Lessonia Vadosa: Partial hydrolysis and elicitor properties of the polymannuronic acid fraction. J. Appl. Phycol. 2004, 16, 127–133. [Google Scholar] [CrossRef]
- Laporte, D.; Vera, J.; Chandía, N.P.; Zúñiga, E.A.; Matsuhiro, B.; Moenne, A. Structurally unrelated algal oligosaccharides differentially stimulate growth and defense against tobacco mosaic virus in tobacco plants. J. Appl. Phycol. 2007, 19, 79–88. [Google Scholar] [CrossRef]
- Staswick, P.E.; Tiryaki, I. The Oxylipin Signal Jasmonic Acid Is Activated by an Enzyme That Conjugates It to Isoleucine in Arabidopsis. Plant Cell 2004, 16, 2117–2127. [Google Scholar] [CrossRef] [Green Version]
- Cheng, C.S.; Samuel, D.; Liu, Y.J.; Shyu, J.C.; Lai, S.M.; Lin, K.F.; Lyu, P.C. Binding Mechanism of Nonspecific Lipid Transfer Proteins and Their Role in Plant Defense. Biochemistry 2004, 43, 13628–13636. [Google Scholar] [CrossRef]
- Maldonado, A.M.; Doerner, P.; Dixon, R.A.; Lamb, C.J.; Cameron, R.K. A putative lipid transfer protein involved in systemic resistance signalling in Arabidopsis. Nature 2002, 419, 395–399. [Google Scholar] [CrossRef]
- Gomès, E.; Sagot, E.; Gaillard, C.; Laquitaine, L.; Poinssot, B.; Sanejouand, Y.H.; Delrot, S.; Coutos-Thévenot, P. Nonspecific Lipid-Transfer Protein Genes Expression in Grape (Vitis sp.) Cells in Response to Fungal Elicitor Treatments. Mol. Plant-Microbe Interact. 2003, 16, 456–464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Modafar, C.; Elgadda, M.; El Boutachfaiti, R.; Abouraicha, E.; Zehhar, N.; Petit, E.; El Alaoui-Talibi, Z.; Courtois, B.; Courtois, J. Induction of natural defence accompanied by salicylic acid-dependant systemic acquired resistance in tomato seedlings in response to bioelicitors isolated from green algae. Sci. Hortic. 2012, 138, 55–63. [Google Scholar] [CrossRef]
- Vera, J.; Castro, J.; Contreras, R.A.; González, A.; Moenne, A. Oligo-carrageenans induce a long-term and broad-range protection against pathogens in tobacco plants (var. Xanthi). Physiol. Mol. Plant Pathol. 2012, 79, 31–39. [Google Scholar] [CrossRef]
- Ghanbarzadeh, M.; Golmoradizadeh, A.; Homaei, A. Carrageenans and carrageenases: Versatile polysaccharides and promising marine enzymes. Phytochem. Rev. 2018, 17, 535–571. [Google Scholar] [CrossRef]
- Khotimchenko, M.; Tiasto, V.; Kalitnik, A.; Begun, M.; Khotimchenko, R.; Leonteva, E.; Bryukhovetskiy, I.; Khotimchenko, Y. Antitumor potential of carrageenans from marine red algae. Carbohydr. Polym. 2020, 246, 116568. [Google Scholar] [CrossRef]
- Sangha, J.S.; Ravichandran, S.; Prithiviraj, K.; Critchley, A.T.; Prithiviraj, B. Sulfated macroalgal polysaccharides λ-carrageenan and ι-carrageenan differentially alter Arabidopsis thaliana resistance to Sclerotinia sclerotiorum. Physiol. Mol. Plant Pathol. 2010, 75, 38–45. [Google Scholar] [CrossRef]
- Sangha, J.S.; Khan, W.; Ji, X.; Zhang, J.; Mills, A.A.S.; Critchley, A.T.; Prithiviraj, B. Carrageenans, Sulphated Polysaccharides of Red Seaweeds, Differentially Affect Arabidopsis thaliana Resistance to Trichoplusia ni (Cabbage Looper). PLoS ONE 2011, 6, e26834. [Google Scholar] [CrossRef]
- Bi, F.; Iqbal, S. Studies on Aqueous Extracts of Three Green Algae as an Elicitor of Plant Defence Mechanism. Pak. J. Bot. 1999, 31, 193–198. [Google Scholar]
- El Boutachfaiti, R.; Delattre, C.; Petit, E.; El Gadda, M.; Courtois, B.; Michaud, P.; El Modafar, C.; Courtois, J. Improved isolation of glucuronan from algae and the production of glucuronic acid oligosaccharides using a glucuronan lyase. Carbohydr. Res. 2009, 344, 1670–1675. [Google Scholar] [CrossRef]
- Elboutachfaiti, R. Procédé D’obtention D’oligosaccharides Anioniques (Oligouronides et Oligosaccharides Sulfates) par Dégradation Enzymatique des Polysaccharides D’algues marines. Ph.D. Thesis, Université de Picardie Jules Verne, Amiens, France, 2008. [Google Scholar]
- Lahaye, M.; Robic, A. Structure and Functional Properties of Ulvan, a Polysaccharide from Green Seaweeds. Biomacromolecules 2007, 8, 1765–1774. [Google Scholar] [CrossRef]
- Jaulneau, V.; Lafitte, C.; Jacquet, C.; Fournier, S.; Salamagne, S.; Briand, X.; Esquerré-Tugayé, M.-T.; Dumas, B. Ulvan, a Sulfated Polysaccharide from Green Algae, Activates Plant Immunity through the Jasmonic Acid Signaling Pathway. J. Biomed. Biotechnol. 2010, 2010, 525291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klarzynski, O.; Descamps, V.; Plesse, B.; Yvin, J.-C.; Kloareg, B.; Fritig, B. Sulfated Fucan Oligosaccharides Elicit Defense Responses in Tobacco and Local and Systemic Resistance Against Tobacco Mosaic Virus. Mol. Plant-Microbe Interact. 2003, 16, 115–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hornsey, I.S.; Hide, D. The production of antimicrobial compounds by British marine algae I. Antibiotic-producing marine algae. Br. Phycol. J. 1974, 9, 353–361. [Google Scholar] [CrossRef] [Green Version]
- Morales, J.L.; Cantillo-Ciau, Z.O.; Sánchez-Molina, I.; Mena-Rejón, G.J. Screening of Antibacterial and Antifungal Activities of Six Marine Macroalgae from Coasts of Yucatán Peninsula. Pharm. Biol. 2006, 44, 632–635. [Google Scholar] [CrossRef]
- Freile-Pelegrín, Y.; Morales, J.L. Antibacterial activity in marine algae from the coast of Yucatan, Mexico. Bot. Mar. 2004, 47, 140–146. [Google Scholar] [CrossRef]
- Crasta, P.J.; Raviraja, N.S.; Sridhar, K.R. Antimicrobial activity of some marine algae of southwest coast of India. Indian J. Mar. Sci. 1997, 26, 201–205. [Google Scholar]
- Sivakumar, S.M.; Safhi, M.M. Isolation and screening of bioactive principle from Chaetomorpha antennina against certain bacterial strains. Saudi Pharm. J. 2013, 21, 119–121. [Google Scholar] [CrossRef] [Green Version]
- Rosaline, X.D.; Sakthivelkumar, S.; Rajendran, K.; Janarthanan, S. Screening of selected marine algae from the coastal Tamil Nadu, South India for antibacterial activity. Asian Pac. J. Trop. Biomed. 2012, 2, S140–S146. [Google Scholar] [CrossRef]
- Febles, C.I.; Arias, A.; Hardisson, A.; López, A.S.; Gil-Rodríguez, M.C. Antimicrobial Activity of Extracts from Some Canary Species of Phaeophyta and Chlorophyta. Phyther. Res. 1995, 9, 385–387. [Google Scholar] [CrossRef]
- Rizvi, M.A.; Shameel, M. Pharmaceutical Biology of Seaweeds from the Karachi Coast of Pakistan. Pharm. Biol. 2005, 43, 97–107. [Google Scholar] [CrossRef]
- Andriani, Y.; Syamsumir, D.F.; Yee, T.C.; Harisson, F.S.; Herng, G.M.; Abdullah, S.A.; Orosco, C.A.; Ali, A.M.; Latip, J.; Kikuzaki, H.; et al. Biological Activities of Isolated Compounds from Three Edible Malaysian Red Seaweeds, Gracilaria Changii, G. Manilaensis and Gracilaria sp. Nat. Prod. Commun. 2016, 11, 1117–1120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ambreen, A.; Khan, H.; Tariq, A.; Ruqqia, A.; Sultana, V.; Ara, J. Evaluation of Biochemical Component and Antimicrobial Activity of Some Seaweeeds Occurring at Karachi Coast. Pak. J. Bot. 2012, 44, 1799–1803. [Google Scholar]
- Narasimhan, M.K.; Pavithra, S.K.; Krishnan, V.; Chandrasekaran, M. In vitro Analysis of Antioxidant, Antimicrobial and Antiproliferative Activity of Enteromorpha antenna, Enteromorpha linza and Gracilaria corticata Extracts. Jundishapur J. Nat. Pharm. Prod. 2013, 8, 151–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arulkumar, A.; Rosemary, T.; Paramasivam, S.; Rajendran, R.B. Phytochemical composition, in vitro antioxidant, antibacterial potential and GC-MS analysis of red seaweeds (Gracilaria corticata and Gracilaria edulis) from Palk Bay, India. Biocatal. Agric. Biotechnol. 2018, 15, 63–71. [Google Scholar] [CrossRef]
- Alam, K.; Agua, T.; Maven, H.; Taie, R.; Rao, K.S.; Burrows, I.; Huber, M.E.; Rali, T. Preliminary Screening of Seaweeds, Seagrass and Lemongrass Oil from Papua New Guinea for Antimicrobial and Antifungal Activity. Int. J. Pharmacogn. 1994, 32, 396–399. [Google Scholar] [CrossRef]
- Ogasawara, K.; Yamada, K.; Hatsugai, N.; Imada, C.; Nishimura, M. Hexose Oxidase-Mediated Hydrogen Peroxide as a Mechanism for the Antibacterial Activity in the Red Seaweed Ptilophora subcostata. PLoS ONE 2016, 11, e0149084. [Google Scholar] [CrossRef] [Green Version]
- Perumal, B.; Chitra, R.; Maruthupandian, A.; Viji, M. Nutritional assessment and bioactive potential of Sargassum polycystum C. Agardh (Brown Seaweed). Indian J. Geo-Mar. Sci. 2019, 48, 492–498. [Google Scholar]
- Kumaresan, M.; Anand, K.V.; Govindaraju, K.; Tamilselvan, S.; Kumar, V.G. Seaweed Sargassum wightii mediated preparation of zirconia (ZrO2) nanoparticles and their antibacterial activity against gram positive and gram negative bacteria. Microb. Pathog. 2018, 124, 311–315. [Google Scholar] [CrossRef]
- Achary, A.; Muthalagu, K.; Guru, M.S. Identification of Phytochemicals from Sargassum wightii against Aedes aegypti. Int. J. Pharm. Sci. Rev. Res. 2014, 29, 314–319. [Google Scholar]
- Rebecca, L.J.; Dhanalakshmi, V.; Thomas, T. A comparison between the effects of three algal extracts against pathogenic bacteria. J. Chem. Pharm. Res. 2012, 4, 4859–4863. [Google Scholar]
- Belattmania, Z.; Reani, A.; Barakate, M.; Zrid, R.; Elatouani, S.; Hassouani, M.; Eddaoui, A.; Bentiss, F.; Sabour, B. Antimicrobial, antioxidant and alginate potentials of Dictyopteris polypodioides (Dictyotales, Phaeophyceae) from the Moroccan Atlantic coast. Der Pharma Chem. 2016, 8, 216–226. [Google Scholar]
- Thangaraju, N.; Venkatalakshmi, R.P.; Chinnasamy, A.; Kannaiyan, P. Synthesis of silver nanoparticles and the antibacterial and anticancer activities of the crude extract of Sargassum polycystum C. Agardh. Nano Biomed. Eng. 2012, 4, 89–94. [Google Scholar] [CrossRef] [Green Version]
- Rebecca, L.J.; Dhanalakshmi, V.; Sharmila, S.; Das, M.P. Bactericidal Activity of Stoechospermum sp. Res. J. Pharm. Biol. Chem. Sci. Bactericidal 2013, 4, 1749–1754. [Google Scholar]
- McConnell, O.J.; Fenical, W. Polyhalogenated 1-Octene-3-Ones, Antibacterial Metabolites from the Red Seaweed Bonnemaisonia Asparagoides. Tetrahedron Lett. 1977, 18, 1851–1854. [Google Scholar] [CrossRef]
- Hornsey, I.S.; Hide, D. The Production of Antimicrobial Compounds by British Marine Algae. IV. Variation of Antimicrobial Activity with Algal Generation. Br. Phycol. J. 1985, 20, 21–25. [Google Scholar] [CrossRef]
- Zouaoui, B.; Ghalem, B.R. The Phenolic Contents and Antimicrobial Activities of Some Marine Algae from the Mediterranean Sea (Algeria). Russ. J. Mar. Biol. 2017, 43, 491–495. [Google Scholar] [CrossRef]
- Devi, K.P.; Suganthy, N.; Kesika, P.; Pandian, S.K. Bioprotective properties of seaweeds: In vitro evaluation of antioxidant activity and antimicrobial activity against food borne bacteria in relation to polyphenolic content. BMC Complement. Altern. Med. 2008, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Rajaboopathi, S.; Thambidurai, S. Evaluation of UPF and antibacterial activity of cotton fabric coated with colloidal seaweed extract functionalized silver nanoparticles. J. Photochem. Photobiol. B Biol. 2018, 183, 75–87. [Google Scholar] [CrossRef]
- Sivagnanam, S.P.; Yin, S.; Choi, J.H.; Park, Y.B.; Woo, H.C.; Chun, B.S. Biological Properties of Fucoxanthin in Oil Recovered from Two Brown Seaweeds Using Supercritical CO2 Extraction. Mar. Drugs 2015, 13, 3422–3442. [Google Scholar] [CrossRef]
- Arguelles, E.D.; Monsalud, R.G.; Sapin, A.B. Chemical Composition and In Vitro Antioxidant and Antibacterial Activities of Sargassum vulgare C. Agardh from Lobo, Batangas, Philippines. Int. Soc. Southeast Asian Agric. Sci. 2019, 25, 112–122. [Google Scholar]
- Barreto, M.; Meyer, J.J.M. Isolation and antimicrobial activity of a lanosol derivative from Osmundaria serrata (Rhodophyta) and a visual exploration of its biofilm covering. South Afr. J. Bot. 2006, 72, 521–528. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Z.; Chen, Y.; Yao, F.; Chen, W.; Zhong, S.; Zheng, F.; Shi, G. Antioxidant, Antibacterial and Antischistosomal Activities of Extracts from Grateloupia livida (Harv). Yamada. PLoS ONE 2013, 8, e80413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suresh, M.; Iyapparaj, P.; Anantharaman, P. Antifouling Activity of Lipidic Metabolites Derived from Padina tetrastromatica. Appl. Biochem. Biotechnol. 2016, 179, 805–818. [Google Scholar] [CrossRef] [PubMed]
- Jiménez, J.T.; O´Connell, S.; Lyons, H.; Bradley, B.; Hall, M. Antioxidant, antimicrobial, and tyrosinase inhibition activities of acetone extract of Ascophyllum nodosum. Chem. Pap. 2010, 64, 434–442. [Google Scholar] [CrossRef]
- Čagalj, M.; Skroza, D.; Razola-Díaz, M.D.C.; Verardo, V.; Bassi, D.; Frleta, R.; Mekinić, I.G.; Tabanelli, G.; Šimat, V. Variations in the Composition, Antioxidant and Antimicrobial Activities of Cystoseira compressa during Seasonal Growth. Mar. Drugs 2022, 20, 64. [Google Scholar] [CrossRef]
- Goecke, F.; Labes, A.; Wiese, J.; Imhoff, J.F. Dual effect of macroalgal extracts on growth of bacteria in Western Baltic Sea. Rev. Biol. Mar. Oceanogr. 2012, 47, 75–86. [Google Scholar] [CrossRef]


| Genera | Species | Hosts | References |
|---|---|---|---|
| Agrobacterium * | Agrobacterium tumefaciens * (syn. Rhizobium radiobacter) | Wide range of agriculturally and economically relevant species, including vines, shade and fruit trees, woody ornamental plants, herbaceous perennials, and other monocots and (mainly) dicotyledonous (host list undefined). | [3,61,64,65,66,67,68,69,70,71,72,73,74,75] |
| Clavibacter | Clavibacter michiganensis subsp. michiganensis | Tomato | [3,76,77] |
| Corynebacterium * | Corynebacterium fascians | Wide range of ornamental and consumable vegetal species | [72,78] |
| Corynebacterium michiganense | Solanaceous host plants | [72,79,80,81] | |
| Corynebacterium sepedonicum | Potato | [72,82,83] | |
| Curtobacterium | Curtobacterium flaccumfaciens pv. flaccumfaciens | Bean | [72,84] |
| Curtobacterium flaccumfaciens pv. poinsettiae | Poinsettia | [72,84] | |
| Dickeya | Dickeya dadantii * | Wide range of economically relevant plant species, highlighting the tropical and subtropical species | [4,63,85] |
| Dickeya solani * | Potato | [4,63] | |
| Erwinia * | Erwinia amylovora * | Fruits of diverse hosts (pear, apple), Rosaceae family | [3,4,64,65,66,67,68,69,70,71,72] |
| Pectobacterium | Pectobacterium atrosepticum * | Potato | [4,63] |
| Pectobacterium carotovorum * | Diverse crop species | [4,72] | |
| Pseudomonas * | Pseudomonas aeruginosa | Tobacco, soybean, bean, cucumber, tomato, and other crops | [3,62,64,65,66,67,68,69,70,71] |
| Pseudomonas syringae pv. lachrymans | Cucumber | [72,86,87,88,89,90] | |
| Pseudomonas marginalis | Wide range of vegetables (such as tomato, parsnip) and ornamental plants (e.g., Zantedeschia spp.) | [72,91,92,93,94,95] | |
| Pseudomonas syringae pv. morsprunorum | Stone fruit of Prunus species (cherries, plum, apricots, peaches) | [72,92,96,97,98] | |
| Pseudomonas savastanoi pv. sacastanoi * | Oleaceae family plants and oleander (Nerium oleander) | [72,92,99,100,101] | |
| Pseudomonas syringae * | Prunus species | [3,4,72] | |
| Pseudomonas syringae pv. tomato * | Tomato | [72,102,103] | |
| Ralstonia solanacearum* (syn. Pseudomonas solanacearum) | Wide range of species including solanaceous plants, weeds, crops, shrubs, and trees | [3,4,61,72,76,104] | |
| Staphylococcus | Staphylococcus aureus | Arabidopsis thaliana | [105,106] |
| Xanthomonas * | Xanthomonas axonopodis * | Orange, cassava, tomato, pepper, crucifers, cotton, rice, beans, grapes, and others | [3,4,64,65,66,67,68,69,70,71,76] |
| Xanthomonas campestris * | Cruciferous plants (including species economically important) | [4,72,107] | |
| Xanthomonas citri subsp. citri | Citrus species (including the economical varieties) | [61,108,109] | |
| Xanthomonas euvesicatoria | Solanaceous species | [61,110,111,112] | |
| Xanthomonas oryzae pv. Oryzae * | Rice species | [4,113,114] | |
| Xanthomonas phaseoli | Common bean | [72,115] | |
| Xylella | Xylella fastidiosa * | Olive, citrus species | [3,4,116,117] |
| ‘Candidatus Liberibacter’ * | - | Citrus species | [118] |
| Species | Macroalgae Source | Antibacterial Activity Test | References |
|---|---|---|---|
| Agrobacterium tumefaciens | Cystoseira humilis var. myriophylloides | Agar diffusion technique | [130] |
| Laminaria digitata | |||
| Bacillus subtilis | Cladophora glomerata | Disc diffusion technique | [131] |
| Chara vulgaris | |||
| Spirogyra crassal | |||
| Erwinia carotovora | Lessonia trabeculata | Liquid-dilution method | [132] |
| Ulva lactuca | Agar diffusion technique | [133] | |
| Erwinia chrysanthemi | Bifurcaria bifurcata | Agar diffusion technique | [128] |
| Codium decorticatum | |||
| Cystoseira humilis var. myriophylloides | |||
| Ellisolandia elongata | |||
| Ericaria selaginoides | |||
| Fucus spiralis | |||
| Gelidium corneum | |||
| Gelidium sp | |||
| Gracilaria cervicornis | |||
| Gymnogongrus crenulatus | |||
| Halopitys incurva | |||
| Laminaria digitata | |||
| Osmundea pinnatifida | |||
| Plocamium cartilagineum | |||
| Sargassum vulgare | |||
| Ulva intestinalis | |||
| Ulva sp. | |||
| Pseudomonas syringae | Lessonia trabeculata | Liquid-dilution method | [132] |
| Macrocystis pyrifera | |||
| Sargassum wightii | Disc diffusion technique | [133] | |
| Ralstonia solaneacearum | Brown seaweed | Field studies | [7] |
| Cladophora glomerata | Disc diffusion technique | [131] | |
| Chara vulgaris | |||
| Spirogyra crassal | |||
| Staphylococcus aureus | Cladophora glomerata | Disc diffusion technique | [131] |
| Chara vulgaris | |||
| Spirogyra crassal | |||
| Xanthomonas campestris | Cladophora glomerata | Disc diffusion technique | [131] |
| Chara vulgaris | |||
| Spirogyra crassal | |||
| Ulva lactuca | Agar diffusion assay | [133] | |
| Xanthomonas oryzae pv. oryzae | Chnoospora minima | Agar diffusion assay | [129,134,135] |
| Gracilaria blodgettii | |||
| Gracilaria edulis | |||
| Hypnea musciformis | |||
| Hypnea valentiae | |||
| Padina boergesenii | |||
| Spyridia hypnoides | |||
| Turbinaria conoides | |||
| Ulva flexuosa | |||
| Ulva lactuca | |||
| Sargassum wightii |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vicente, T.F.L.; Félix, C.; Félix, R.; Valentão, P.; Lemos, M.F.L. Seaweed as a Natural Source against Phytopathogenic Bacteria. Mar. Drugs 2023, 21, 23. https://doi.org/10.3390/md21010023
Vicente TFL, Félix C, Félix R, Valentão P, Lemos MFL. Seaweed as a Natural Source against Phytopathogenic Bacteria. Marine Drugs. 2023; 21(1):23. https://doi.org/10.3390/md21010023
Chicago/Turabian StyleVicente, Tânia F. L., Carina Félix, Rafael Félix, Patrícia Valentão, and Marco F. L. Lemos. 2023. "Seaweed as a Natural Source against Phytopathogenic Bacteria" Marine Drugs 21, no. 1: 23. https://doi.org/10.3390/md21010023