
Phycobiliproteins—A Family of Algae-Derived Biliproteins: Productions, Characterization and Pharmaceutical Potentials

Abstract
:1. Introduction
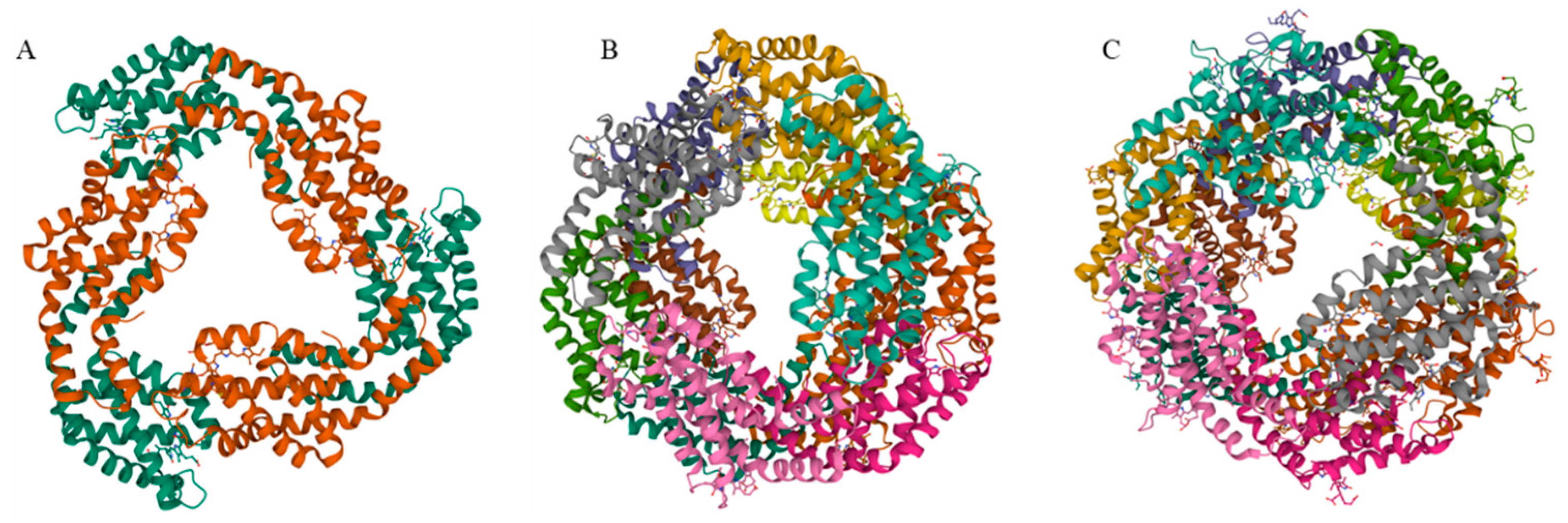
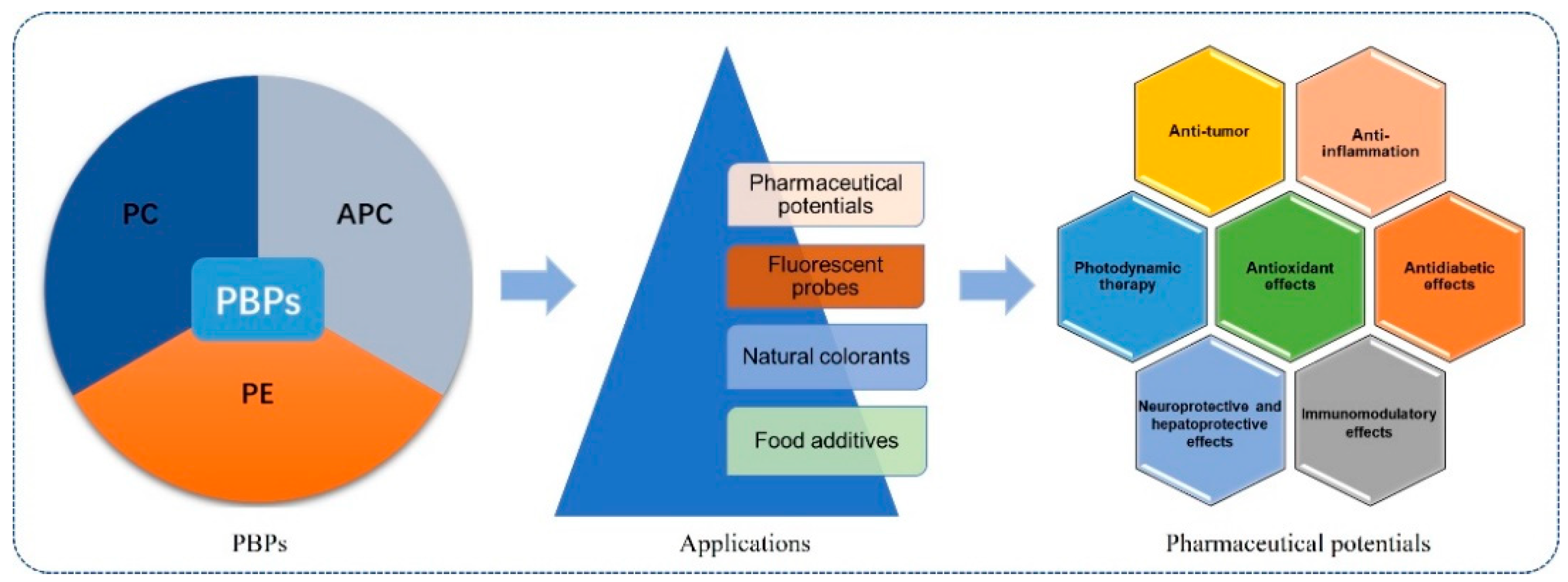
2. Phycobiliproteins
2.1. Allophycocyanin
2.2. Phycocyanin
2.3. Phycoerythrin
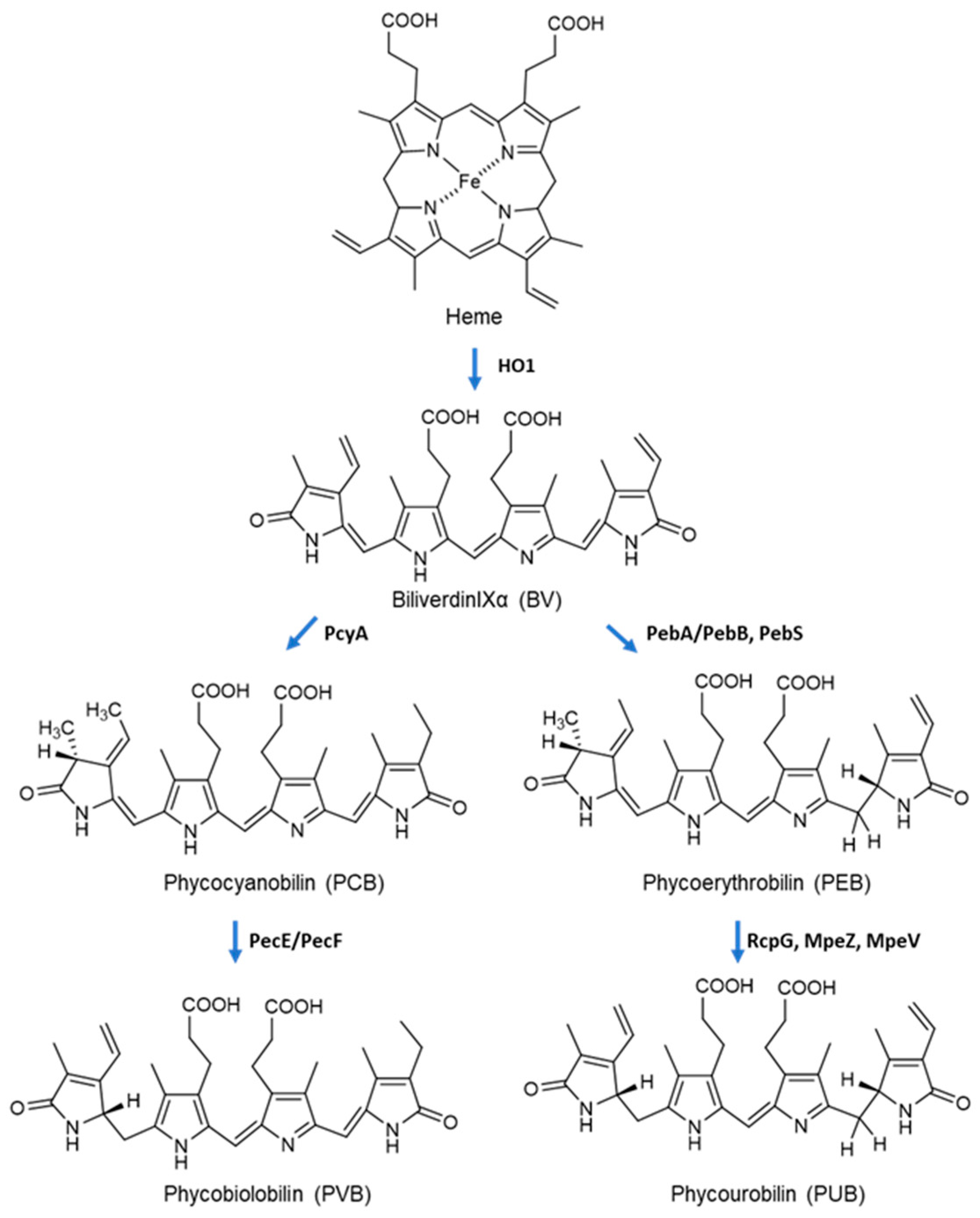
2.4. Biosynthesis of Phycobiliproteins
3. Production of Phycobiliproteins
3.1. Native Phycobiliproteins

{kind=link}
{kind=link}
{kind=link}
| PBPs | Sources for PBP Gene | Hosts | Phycobilins | Lyases | Chromophorylation (%) | Reference |
|---|---|---|---|---|---|---|
| Apo-ApcA, Apo-ApcB | Synechococcos sp. PCC 7002 | E. coli | PCB | - | - | [41] |
| Apo-APC | Asterocapsa nidulans (formerly Anacystis nidulans) | E. coli | PCB | - | - | [42] |
| Apo-CpcA | Anacystis nidufuns R2 | E. coli | PCB | - | - | [43] |
| Holo-CpcA | Synechocystis sp. PCC6803 | E. col | PCB | CpcE/CpcF | ~33.3% | [44] |
| Holo-PecA | Anabaena sp. PCC7120 | E. coli | PVB | PecE/PecF | n.d. | [45] |
| Holo-ApcAB | Synechococcus sp. PCC 7002 | E. coli | PCB | CpcU/CpcS | 71.9% | [46] |
| Holo-ApcAB | Synechocystis sp. PCC 6803 | E. coli | PCB | CpcU/CpcS | n.d. | [47] |
| Holo-ApcAB | Gracilaria chilensis (formerly Agarophyton chilense) | E. coli | PCB | CpcU/CpcS | 52% and 57% | [48] |
| Holo-ApcA | Synechococcus elongatus BP-1 | E. coli | PCB | CpcS | n.d. | [46] |
| Holo-ApcB | Spirulina sp. | E. coli | PCB | CpcS | 81.4% | [49] |
| Holo-ApcA | Synechococcus elongatus BP-1 | E. coli | PCB | CpcS | n.d. | [50] |
| Streptavidin-Holo-ApcA | Synechococcus elongatus BP-1 | E. coli | PCB or PEB | CpcS | n.d. | [51] |
| Streptavidin-Holo-ApcA | Synechococcus elongatus BP-1 | E. coli | PEB | CpcS | Up to 98.6% | [52] |
| Holo-ApcB | Synechococcus elongatus BP-1 | E. coli | PCB | CpcS | n.d. | [53] |
| Holo-ApcF | Synechococcus sp. PCC 7002 | E. coli | PCB | CpcU/CpcS | 68.1% | [46] |
| Holo-CpcA | Synechocystis sp. PCC 6803 | E. coli | PCB | CpcE/CpcF | 48.1% | [46] |
| Holo-CpcB | Synechocystis sp. PCC 6803 | E. coli | PCB | CpcU/CpcS | 37.1 | [46] |
| Holo-CpcB | Synechocystis sp. PCC 6803 | E. coli | PCB | CpcT | 17.4 | [46] |
| Holo-CpcB | Synechococcuselongatus BP-1 | E. coli | PEB, PUB | CpcU, CpcT | n.d. | [54] |
| Holo-CpcA | Synechocystis sp. PCC 6803 Synechococcus sp. PCC 7002 | E. coli | PCB, PEB, PΦB, PUB, PVB, PtVB | CpcE/CpcF PecE/PecF | n.d. | [55] |
| Holo-CpeA | Microchaete diplosiphon (formerly Fremyella diplosiphon) | E. coli | PEB | CpeY | n.d. | [56] |
| Holo-CpeB | Synechococcus sp. RS9916 | E. coli | PUB | MpeV | n.d. | [30] |
| Holo-CpeB | Microchaete diplosiphon (formerly Fremyella diplosiphon) | E. coli | PEB | CpeF | n.d | [29] |
| Holo-CpeB | Prochlorococcus marinus MED4 (Cyanobacteria) | E. coli | PEB | CpeS | n.d. | [57] |
| PcA/PcB | Gracilariopsis lemaneiformis (Rhodophyta) | E. coli | PCB | CpcU/CpcS, CpcE/CpcF, CpcT | n.d. | [58] |
| Holo-MpeA | Synechococcus sp. RS9916 | E. coli | PUB | MpeZ | n.d. | [23] |
| Holo-C-PC equipped with different tags | Anabaena sp. PCC7120 | Anabaena sp. PCC7120 | PCB | - | n.d. | [59] |
| Holo-APC | Cyanophora paradoxa (Glaucophyta) | Synechococcus sp. PCC 7002 | PCB | - | n.d. | [41] |
3.2. Recombinant Phycobiliproteins
4. Pharmaceutical Potentials
4.1. Antioxidant Effects
4.2. Anti-Tumor Effects
4.3. Anti-Inflammatory Effects
4.4. Antidiabetic Effects
4.5. Neuroprotective and Hepatoprotective Effects
4.6. Immunomodulatory Effects
4.7. Photodynamic Therapy
4.8. Other Biological Activities
5. Conclusions and Future Perspectives
Author Contributions
Funding
Conflicts of Interest
Abbreviations
References
- Brown, E.S.; Allsopp, P.J.; Magee, P.J.; Gill, C.I.; Nitecki, S.; Strain, C.R.; McSorley, E.M. Seaweed and human health. Nutr. Rev. 2014, 72, 205–216. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, C.; Gallagher, E.; Tasdemir, D.; Hayes, M. Heart health peptides from macroalgae and their potential use in functional foods. J. Agric. Food Chem. 2011, 59, 6829–6836. [Google Scholar] [CrossRef] [PubMed]
- Mysliwa-Kurdziel, B.; Solymosi, K. Phycobilins and phycobiliproteins used in food industry and medicine. Mini. Rev. Med. Chem. 2017, 17, 1173–1193. [Google Scholar] [CrossRef] [Green Version]
- Ashaolu, T.J.; Samborska, K.; Lee, C.C.; Tomas, M.; Capanoglu, E.; Tarhan, Ö.; Taze, B.; Jafari, S.M. Phycocyanin, a super functional ingredient from algae; properties, purification characterization, and applications. Int. J. Biol. Macromol. 2021, 193, 2320–2331. [Google Scholar] [CrossRef]
- Sidler, W.A. Phycobilisome and Phycobiliprotein Structures; Springer: Zürich, The Netherlands, 1994; pp. 140–216. [Google Scholar]
- Glazer, A.N. Phycobiliproteins—A family of valuable, widely used fluorophores. J. Appl. Phycol. 1994, 6, 105–112. [Google Scholar] [CrossRef]
- Sekar, S.; Chandramohan, M. Phycobiliproteins as a commodity: Trends in applied research, patents and commercialization. J. Appl. Phycol. 2008, 20, 113–136. [Google Scholar] [CrossRef]
- Insights, F.M. Phycocyanin Market: Food & Beverage Application to Hold Close to 85% Value Share throughout the Forecast Period: Global Industry Analysis (2013–2017) & Opportunity Assessment (2018–2028); 2018; Available online: https://www.futuremarketinsights.com/reports/phycocyanin-market/toc (accessed on 20 September 2021).
- Chakdar, H.; Pabbi, S. Cyanobacterial phycobilins: Production, purification, and regulation. In Frontier Discoveries and Innovations in Interdisciplinary Microbiology; Shukla, P., Ed.; Springer: New Delhi, India, 2016; pp. 45–69. [Google Scholar]
- Apt, K.E.; Collier, J.L.; Grossman, A.R. Evolution of the phycobiliproteins. J. Mol. Biol. 1995, 248, 79–96. [Google Scholar] [CrossRef]
- Li, Y.; Lin, Y.; Loughlin, P.C.; Chen, M. Optimization and effects of different culture conditions on growth of Halomicronema hongdechloris—A filamentous cyanobacterium containing chlorophyll f. Front. Plant Sci. 2014, 5, 67. [Google Scholar] [CrossRef] [Green Version]
- Gan, F.; Zhang, S.; Rockwell, N.C.; Martin, S.S.; Lagarias, J.C.; Bryant, D.A. Extensive remodeling of a cyanobacterial photosynthetic apparatus in far-red light. Science 2014, 345, 1312–1317. [Google Scholar] [CrossRef] [Green Version]
- Gan, F.; Bryant, D.A. Adaptive and acclimative responses of cyanobacteria to far-red light. Environ. Microbiol. 2015, 17, 3450–3465. [Google Scholar] [CrossRef]
- Li, Y.; Lin, Y.; Garvey, C.J.; Birch, D.; Corkery, R.W.; Loughlin, P.C.; Scheer, H.; Willows, R.D.; Chen, M. Characterization of red-shifted phycobilisomes isolated from the chlorophyll f-containing cyanobacterium Halomicronema hongdechloris. Biochim. Biophys. Acta 2016, 1857, 107–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soulier, N.; Bryant, D.A. The structural basis of far-red light absorbance by allophycocyanins. Photosynth. Res. 2021, 147, 11–26. [Google Scholar] [CrossRef] [PubMed]
- Debreczeny, M.P.; Sauer, K.; Zhou, J.; Bryan, D.A. Comparison of calculated and experimentally resolved rate constants for excitation energy transfer in C-phycocyanin: I. Monomers. J. Phys. Chem. 1995, 99, 8412–8419. [Google Scholar] [CrossRef]
- Debreczeny, M.P.; Sauer, K.; Zhou, J.; Bryan, D.A. Comparison of calculated and experimentally resolved rate constants for excitation energy transfer in C-phycocyanin: II. Trimers. J. Phys. Chem. 1995, 99, 8420–8431. [Google Scholar] [CrossRef]
- Pagels, F.; Guedes, A.C.; Amaro, H.M.; Kijjoa, A.; Vasconcelos, V. Phycobiliproteins from cyanobacteria: Chemistry and biotechnological applications. Biotechnol. Adv. 2019, 37, 422–443. [Google Scholar] [CrossRef] [PubMed]
- Adir, N.; Dines, M.; Klartag, M.; Mcgregor, A.; Melamed-Frank, M. Assembly and disassembly of phycobilisomes. In Complex Intracellular Structures in Prokaryotes; Springer: Berlin/Heidelberg, Germany, 2006; Volume 2, pp. 47–77. [Google Scholar]
- Everroad, C.; Six, C.; Partensky, F.; Thomas, J.C.; Holtzendorff, J.; Wood, A.M. Biochemical bases of type IV chromatic adaptation in marine Synechococcus spp. J. Bacteriol. 2006, 188, 3345–3356. [Google Scholar] [CrossRef] [Green Version]
- Kehoe, D.M. Chromatic adaptation and the evolution of light color sensing in cyanobacteria. Proc. Natl. Acad. Sci. USA 2010, 107, 9029–9030. [Google Scholar] [CrossRef] [Green Version]
- Scheer, H.; Zhao, K.H. Biliprotein maturation: The chromophore attachment. Mol. Microbiol. 2008, 68, 263–276. [Google Scholar] [CrossRef] [Green Version]
- Shukla, A.; Biswas, A.; Blot, N.; Partensky, F.; Karty, J.A.; Hammad, L.A.; Garczarek, L.; Gutu, A.; Schluchter, W.M.; Kehoe, D.M. Phycoerythrin-specific bilin lyase-isomerase controls blue-green chromatic acclimation in marine Synechococcus. Proc. Natl. Acad. Sci. USA 2012, 109, 20136–20141. [Google Scholar] [CrossRef] [Green Version]
- Zhao, K.; Su, P.; Böhm, S.; Song, B.; Zhou, M.; Bubenzer, C.; Scheer, H. Reconstitution of phycobilisome core-membrane linker, Lcm, by autocatalytic chromophore binding to ApcE. Biochim. Biophys. Acta 2005, 1706, 81–87. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Gasparich, G.E.; Stirewalt, V.L.; de Lorimier, R.; Bryant, D.A. The cpcE and cpcF genes of Synechococcus sp. PCC 7002. Construction and phenotypic characterization of interposon mutants. J. Biol. Chem. 1992, 267, 16138–16145. [Google Scholar] [CrossRef]
- Saunée, N.A.; Williams, S.R.; Bryant, D.A.; Schluchter, W.M. Biogenesis of phycobiliproteins: II. CpcS-I and CpcU comprise the heterodimeric bilin lyase that attaches phycocyanobilin to Cys-82 of beta-phycocyanin and Cys-81 of allophycocyanin subunits in Synechococcus sp. PCC 7002. J. Biol. Chem. 2008, 283, 7513–7522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, G.; Saunée, N.A.; Williams, S.R.; Gallo, E.F.; Schluchter, W.M.; Bryant, D.A. Identification and characterization of a new class of bilin lyase: The cpcT gene encodes a bilin lyase responsible for attachment of phycocyanobilin to Cys-153 on the beta-subunit of phycocyanin in Synechococcus sp. PCC 7002. J. Biol. Chem. 2006, 281, 17768–17778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahmoud, R.M.; Sanfilippo, J.E.; Nguyen, A.A.; Strnat, J.A.; Partensky, F.; Garczarek, L.; Kassem, A.E.N.; Kehoe, D.M.; Schluchter, W.M. Adaptation to blue light in marine Synechococcus requires MpeU, an enzyme with similarity to phycoerythrobilin lyase isomerases. Front. Microbiol. 2017, 8, 243. [Google Scholar] [CrossRef] [Green Version]
- Kronfel, C.M.; Hernandez, C.V.; Frick, J.P.; Hernandez, L.S.; Gutu, A.; Karty, J.A.; Boutaghou, M.N.; Kehoe, D.M.; Cole, R.B.; Schluchter, W.M. CpeF is the bilin lyase that ligates the doubly linked phycoerythrobilin on β-phycoerythrin in the cyanobacterium Fremyella diplosiphon. J. Biol. Chem. 2019, 294, 3987–3999. [Google Scholar] [CrossRef]
- Carrigee, L.A.; Frick, J.P.; Karty, J.A.; Garczarek, L.; Partensky, F.; Schluchter, W.M. MpeV is a lyase isomerase that ligates a doubly linked phycourobilin on the β-subunit of phycoerythrin I and II in marine Synechococcus. J. Biol. Chem. 2021, 296, 100031. [Google Scholar] [CrossRef]
- Román, B.R.; Alvárez-Pez, J.M.; Fernández, A.F.G.; Grima, M.E. Recovery of pure B- Phycoerythrin from the microalga Porphyridium cruentum. J. Biotechnol. 2002, 93, 73–85. [Google Scholar] [CrossRef]
- Khazi, M.I.; Demirel, Z.; Dalay, M.C. Evaluation of growth and phycobiliprotein composition of cyanobacteria isolates cultivated in different nitrogen sources. J. Appl. Phycol. 2018, 30, 1513–1523. [Google Scholar] [CrossRef]
- Syrpas, M.; Bukauskaitė, J.; Ramanauskienė, K.; Karosienė, J.R.; Majienė, D.; Bašinskienė, L.; Venskutonis, P.R. Ultrasound-assisted extraction and assessment of biological activity of phycobiliprotein-rich aqueous extracts from wild cyanobacteria (Aphanizomenon flos-aquae). J. Agric. Food Chem. 2020, 68, 1896–1909. [Google Scholar] [CrossRef]
- Moraes, C.C.; Sala, L.; Cerveira, G.P.; Kalil, S.J. C-phycocyanin extraction from Spirulina platensis wet biomass. Braz. J. Chem. Eng. 2011, 28, 45–49. [Google Scholar] [CrossRef] [Green Version]
- Mogany, T.; Kumari, S.; Swalaha, F.M.; Bux, F. Extraction and characterization of analytical grade C-phycocyanin from Euhalothece sp. J. Appl. Phycol. 2019, 31, 1661–1674. [Google Scholar] [CrossRef]
- Yan, S.; Zhu, L.; Su, H.; Zhang, X.; Chen, X.; Zhou, B.; Zhang, Y. Single-step chromatography for simultaneous purification of C-phycocyanin and allophycocyanin with high purity and recovery from Spirulina (Arthrospira) platensis. J. Appl. Phycol. 2011, 23, 1–6. [Google Scholar] [CrossRef]
- Sorensen, L.; Hantke, A.; Eriksen, N.T. Purification of the photosynthetic pigment C-phycocyanin from heterotrophic Galdieria sulphuraria. J. Sci. Food Agric. 2013, 93, 2933–2938. [Google Scholar] [CrossRef] [PubMed]
- Kumar, D.; Dhar, D.W.; Pabbi, S.; Kumar, N.; Walia, S. Extraction and purification of C-phycocyanin from Spirulina platensis (CCC540). Indian J. Plant Physiol. 2014, 19, 184–188. [Google Scholar] [CrossRef] [Green Version]
- Singh, N.K.; Parmar, A.; Madamwar, D. Optimization of medium components for increased production of C-phycocyanin from Phormidium ceylanicum and its purification by single step process. Bioresour. Technol. 2009, 100, 1663–1669. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Su, H.; Pu, Y.; Chen, J.; Liu, L.; Liu, Q.; Qin, S. Phycobiliproteins: Molecular structure, production, applications, and prospects. Biotechnol. Adv. 2019, 37, 340–353. [Google Scholar] [CrossRef] [PubMed]
- Lorimier, D.R.; Guglielmi, G.; Bryant, D.A.; Stevens, S.J. Functional expression of plastid allophycocyanin genes in a cyanobacterium. J. Bacteriol. 1987, 169, 1830–1835. [Google Scholar] [CrossRef] [Green Version]
- Qin, S.; Tang, Z.; Lin, F.; Sung, L.; Tseng, C. Genomic cloning, expression and recombinant protein purification of α and β subunits of the allophycocyanin gene (apc) from the cyanobacterium Anacystis nidulans UTEX 625. J. App. Phycol. 2004, 16, 483–487. [Google Scholar] [CrossRef]
- Lau, R.H.; Alvarado-Urbina, G.; Lau, P.C. Phycocyanin alpha-subunit gene of Anacystis nidulans R2: Cloning, nucleotide sequencing and expression in Escherichia coli. Gene 1987, 52, 21–29. [Google Scholar] [CrossRef]
- Tooley, A.J.; Cai, Y.A.; Glazer, A.N. Biosynthesis of a fluorescent cyanobacterial C-phycocyanin holo-alpha subunit in a heterologous host. Proc. Natl. Acad. Sci. USA 2001, 98, 10560–10565. [Google Scholar] [CrossRef] [Green Version]
- Tooley, A.J.; Glazer, A.N. Biosynthesis of the cyanobacterial light-harvesting polypeptide phycoerythrocyanin holo-alpha subunit in a heterologous host. J. Bacteriol. 2002, 184, 4666–4671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biswas, A.; Vasquez, Y.M.; Dragomani, T.M.; Kronfel, M.L.; Williams, S.R.; Alvey, R.M.; Bryant, D.A.; Schluchter, W.M. Biosynthesis of cyanobacterial phycobiliproteins in Escherichia coli: Chromophorylation efficiency and specificity of all bilin lyases from Synechococcus sp. strain PCC 7002. Appl. Environ. Microbiol. 2010, 76, 2729–2739. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Chen, Y.; Lu, Y.; Chen, H.; Li, F.; Qin, S. Biosynthesis of fluorescent cyanobacterial allophycocyanin trimer in Escherichia coli. Photosynth. Res. 2010, 105, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Dagnino-Leone, J.; Figueroa, M.; Uribe, E.; Hinrichs, M.V.; Ortiz-López, D.; Martínez-Oyanedel, J.; Bunster, M. Biosynthesis and characterization of a recombinant eukaryotic allophycocyanin using prokaryotic accessory enzymes. MicrobiologyOpen 2020, 9, e989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ge, B.; Tang, Z.; Lin, L.; Ren, Y.; Yang, Y.; Qin, S. Pilot-scale fermentation and purification of the recombinant allophycocyanin over-expressed in Escherichia coli. Biotechnol. Lett. 2005, 27, 783–787. [Google Scholar] [CrossRef]
- Chen, H.; Liu, Q.; Zhao, J.; Jiang, P. Biosynthesis, spectral properties and thermostability of cyanobacterial allophycocyanin holo-α subunits. Int. J. Biol. Macromol. 2016, 88, 88–92. [Google Scholar] [CrossRef]
- Wu, J.; Chen, H.; Zhao, J.; Jiang, P. Fusion proteins of streptavidin and allophycocyanin alpha subunit for immunofluorescence assay. Biochem. Eng. J. 2017, 125, 97–103. [Google Scholar] [CrossRef]
- Chen, H.; Jiang, P. Metabolic engineering of Escherichia coli for efficient biosynthesis of fluorescent phycobiliprotein. Microb. Cell Fact. 2019, 18, 58. [Google Scholar] [CrossRef]
- Chen, H.; Lin, H.; Li, F.; Jiang, P.; Qin, S. Biosynthesis of a stable allophycocyanin beta subunit in metabolically engineered Escherichia coli. J. Biosci. Bioeng. 2013, 115, 485–489. [Google Scholar] [CrossRef]
- Chen, H.; Zheng, C.; Jiang, P.; Ji, G. Biosynthesis of a phycocyanin beta subunit with two noncognate chromophores in Escherichia coli. Appl. Biochem. Biotechnol. 2020, 191, 763–771. [Google Scholar] [CrossRef]
- Alvey, R.M.; Biswas, A.; Schluchter, W.M.; Bryant, D.A. Attachment of noncognate chromophores to CpcA of Synechocystis sp. PCC 6803 and Synechococcus sp. PCC 7002 by heterologous expression in Escherichia coli. Biochemistry 2011, 50, 4890–4902. [Google Scholar] [CrossRef] [PubMed]
- Biswas, A.; Boutaghou, M.N.; Alvey, R.M.; Kronfel, C.M.; Cole, R.B.; Bryant, D.A.; Schluchter, W.M. Characterization of the activities of the CpeY, CpeZ, and CpeS bilin lyases in phycoerythrin biosynthesis in Fremyella diplosiphon strain UTEX 481. J. Biol. Chem. 2011, 286, 35509–35521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiethaus, J.; Busch, A.W.; Kock, K.; Leichert, L.I.; Herrmann, C.; Frankenberg-Dinkel, N. CpeS is a lyase specific for attachment of 3Z-PEB to Cys82 of β-phycoerythrin from Prochlorococcus marinus MED4. J. Biol. Chem. 2010, 285, 37561–37569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, D.; Zang, X.; Guo, Y.; Xiao, D.; Cao, X.; Liu, Z.; Zhang, F.; Jin, Y.; Shi, J.; Wang, Z.; et al. Cloning of pcB and pcA gene from Gracilariopsis lemaneiformis and expression of a fluorescent phycocyanin in heterologous host. Genes 2019, 10, 322. [Google Scholar] [CrossRef] [Green Version]
- Cai, Y.A.; Murphy, J.T.; Wedemayer, G.J.; Glazer, A.N. Recombinant phycobiliproteins. Recombinant C-phycocyanins equipped with affinity tags, oligomerization, and biospecific recognition domains. Anal. Biochem. 2001, 290, 186–204. [Google Scholar] [CrossRef]
- Guan, X.; Zhang, W.; Zhang, X.; Li, Y.; Wang, J.; Lin, H.; Tang, X.; Qin, S. A potent anti-oxidant property: Fluorescent recombinant alpha-phycocyanin of Spirulina. J. Appl. Microbiol. 2009, 106, 1093–1100. [Google Scholar] [CrossRef]
- Qiang, X.; Wang, L.; Niu, J.; Gong, X.; Wang, G. Phycobiliprotein as fluorescent probe and photosensitizer: A systematic review. Int. J. Biol. Macromol. 2021, 193, 1910–1917. [Google Scholar] [CrossRef]
- Ge, B.; Li, Y.; Sun, H.; Zhang, S.; Hu, P.; Qin, S.; Huang, F. Combinational biosynthesis of phycocyanobilin using genetically-engineered Escherichia coli. Biotechnol. Lett. 2013, 35, 689–693. [Google Scholar] [CrossRef]
- Romay, C.; Armesto, J.; Remirez, D.; Gonzalez, R.; Ledon, N.; Garcia, I. Antioxidant and anti-inflammatory properties of C-phycocyanin from blue-green algae. Inflamm. Res. 1998, 47, 36–41. [Google Scholar] [CrossRef]
- Romay, C.; Ledon, N.; Gonzalez, R. Further studies on anti-inflammatory activity of phycocyanin in some animal models of inflammation. Inflamm. Res. 1998, 47, 334–338. [Google Scholar] [CrossRef]
- Bhat, V.B.; Madyastha, K.M. C-phycocyanin: A potent peroxyl radical scavenger in vivo and in vitro. Biochem. Biophys. Res. Commun. 2000, 275, 20–25. [Google Scholar] [CrossRef] [PubMed]
- Sonani, R.R.; Singh, N.K.; Kumar, J.; Thakar, D.; Madamwar, D. Concurrent purification and antioxidant activity of phycobiliproteins from Lyngbya sp. A09DM: An antioxidant and anti-aging potential of phycoerythrin in Caenorhabditis elegans. Process. Biochem. 2014, 49, 1757–1766. [Google Scholar] [CrossRef]
- Hirata, T.; Tanaka, M.; Ooike, M.; Tsunomura, T.; Sakaguchi, M. Antioxidant activities of phycocyanobilin prepared from Spirulina platensis. J. Appl. Phycol. 2000, 12, 435–439. [Google Scholar] [CrossRef]
- Bhat, V.B.; Madyastha, K.M. Scavenging of peroxynitrite by phycocyanin and phycocyanobilin from Spirulina platensis: Protection against oxidative damage to DNA. Biochem. Biophys. Res. Commun. 2001, 285, 262–266. [Google Scholar] [CrossRef]
- Benedetti, S.; Benvenuti, F.; Scoglio, S.; Canestrari, F. Oxygen radical absorbance capacity of phycocyanin and phycocyanobilin from the food supplement Aphanizomenon flos-aquae. J. Med. Food. 2010, 13, 223–2277. [Google Scholar] [CrossRef]
- Pendyala, B.; Patras, A.; Dash, C. Phycobilins as potent food bioactive broad-spectrum inhibitors against proteases of SARS-CoV-2 and other coronaviruses: A preliminary study. Front. Microbiol. 2021, 12, 645713. [Google Scholar] [CrossRef]
- Stocker, R.; Yamamoto, Y.; McDonagh, A.F.; Glazer, A.N.; Ames, B.N. Bilirubin is an antioxidant of possible physiological importance. Science 1987, 235, 1043–1046. [Google Scholar] [CrossRef]
- Ge, B.; Qin, S.; Han, L.; Lin, F.; Ren, Y. Antioxidant properties of recombinant allophycocyanin expressed in Escherichia coli. J. Photochem. Photobiol. B 2006, 84, 175–180. [Google Scholar] [CrossRef]
- Zhang, W.; Guan, X.; Yang, Y.; Ge, B.; Chen, H.; Li, F.; Qin, S. Biosynthesis of fluorescent allophycocyanin alpha-subunits by autocatalysis in Escherichia coli. Biotechnol. Appl. Biochem. 2009, 52, 135–140. [Google Scholar] [CrossRef]
- Pleonsil, P.; Soogarun, S.; Suwanwong, Y. Anti-oxidant activity of holo- and apo-c-phycocyanin and their protective effects on human erythrocytes. Int. J. Biol. Macromol. 2013, 60, 393–398. [Google Scholar] [CrossRef]
- Atmaca, G. Antioxidant effects of sulfur-containing amino acids. Yonsei Med. J. 2004, 45, 776–788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Requejo, R.; Hurd, T.R.; Costa, N.J.; Murphy, M.P. Cysteine residues exposed on protein surfaces are the dominant intramitochondrial thiol and may protect against oxidative damage. FEBS J. 2010, 277, 1465–1480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Z.; Liu, L.; Chen, X.; Wang, J.; Chen, M.; Zhang, Y.Z.; Zhou, B. Factors that affect antioxidant activity of C-phycocyanins from Spirulina platensis. J. Food Biochem. 2005, 29, 313–322. [Google Scholar] [CrossRef]
- Braune, S.; Krüger-Genge, A.; Kammerer, S.; Jung, F.; Küpper, J.H. Phycocyanin from Arthrospira platensis as potential anti-cancer drug: Review of in vitro and in vivo studies. Life 2021, 11, 91. [Google Scholar] [CrossRef] [PubMed]
- Hao, S.; Li, S.; Wang, J.; Yan, Y.; Ai, X.; Zhang, J.; Ren, Y.; Wu, T.; Liu, L.; Wang, C. Phycocyanin exerts anti-proliferative effects through down-regulating TIRAP/NF-κB activity in human non-small cell lung cancer cells. Cells 2019, 8, 588. [Google Scholar] [CrossRef] [Green Version]
- Hao, S.; Li, S.; Wang, J.; Zhao, L.; Yan, Y.; Wu, T.; Zhang, J.; Wang, C. C-Phycocyanin suppresses the in vitro proliferation and migration of non-small-cell lung cancer cells through reduction of RIPK1/NF-κB activity. Mar. Drugs 2019, 17, 362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bottone, C.; Camerlingo, R.; Miceli, R.; Salbitani, G.; Sessa, G.; Pirozzi, G.; Carfagna, S. Antioxidant and anti-proliferative properties of extracts from heterotrophic cultures of Galdieria sulphuraria. Nat. Prod. Res. 2019, 33, 1659–1663. [Google Scholar] [CrossRef] [PubMed]
- Naidu, K.A.; Sarada, R.; Manoj, G.; Khan, M.Y.; Swamy, M.M.; Viswanatha, S.; Murthy, K.N.; Ravishankar, G.A.; Srinivas, L. Toxicity assessment of phycocyanin—A blue colorant from blue green alga Spirulina platensis. Food Biotech. 1999, 13, 51–66. [Google Scholar] [CrossRef]
- Liao, G.; Gao, B.; Gao, Y.; Yang, X.; Cheng, X.; Ou, Y. Phycocyanin inhibits tumorigenic potential of pancreatic cancer cells: Role of apoptosis and autophagy. Sci. Rep. 2016, 6, 34564. [Google Scholar] [CrossRef]
- Jiang, L.; Wang, Y.; Liu, G.; Liu, H.; Zhu, F.; Ji, H.; Li, B. C-Phycocyanin exerts anti-cancer effects via the MAPK signaling pathway in MDA-MB-231 cells. Cancer Cell Int. 2018, 18, 12. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Xu, L.; Ni, C.; Lin, L.; Zhang, C. Inhibitory effect of phycocyanin from Spirulina platensis on the growth of human leukemia K562 cells. J. Appl. Phycol. 2000, 12, 125–130. [Google Scholar] [CrossRef]
- Jiang, L.; Wang, Y.; Zhu, F.; Liu, G.; Liu, H.; Ji, H.; Zheng, S.; Li, B. Molecular mechanism of anti-cancer activity of the nano-drug C-PC/CMC-CD59sp NPs in cervical cancer. J. Cancer. 2019, 10, 92–104. [Google Scholar] [CrossRef] [PubMed]
- Subhashini, J.; Mahipal, S.V.; Reddy, M.C.; Reddy, M.M.; Rachamallu, A.; Reddanna, P. Molecular mechanisms in C-Phycocyanin induced apoptosis in human chronic myeloid leukemia cell line-K562. Biochem. Pharmacol. 2004, 68, 453–462. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Gao, M.; Zhang, X.; Chu, X. Molecular immune mechanism of C-phycocyanin from Spirulina platensis induces apoptosis in HeLa cells in vitro. Biotechnol. Appl. Biochem. 2006, 43, 155–164. [Google Scholar] [PubMed]
- Pattarayan, D.; Rajarajan, D.; Ayyanar, S.; Palanichamy, R.; Subbiah, R. C-phycocyanin suppresses transforming growth factor-beta1-induced epithelial mesenchymal transition in human epithelial cells. Pharmacol. Rep. 2017, 69, 426–431. [Google Scholar] [CrossRef]
- Ji, H.; Liu, G.; Han, J.; Zhu, F.; Dong, X.; Li, B. C-phycocyanin inhibits epithelial-to-mesenchymal transition in Caski cells. Cancer Cell Int. 2020, 20, 292. [Google Scholar] [CrossRef]
- Wang, H.; Liu, Y.; Gao, X.; Carter, C.L.; Liu, Z.R. The recombinant beta subunit of C-phycocyanin inhibits cell proliferation and induces apoptosis. Cancer Lett. 2007, 247, 150–158. [Google Scholar] [CrossRef]
- Gantar, M.; Dhandayuthapani, S.; Rathinavelu, A. Phycocyanin induces apoptosis and enhances the effect of topotecan on prostate cell line LNCaP. J. Med. Food. 2012, 15, 1091–1095. [Google Scholar] [CrossRef]
- Yang, F.; Li, B.; Chu, X.M.; Lv, C.Y.; Xu, Y.J.; Yang, P. Molecular mechanism of inhibitory effects of C-phycocyanin combined with all-trans-retinoic acid on the growth of HeLa cells in vitro. Tumour. Biol. 2014, 35, 5619–5628. [Google Scholar] [CrossRef]
- Remirez, D.; Gonz’alez, A.; Merino, N.; Gonz´alez, R.; Ancheta, O.; Romay, C.; Rodríguez, S. Effect of phycocyanin in zymosan-induced arthritis in mice—Phycocyanin as an antiarthritic compound. Drug Dev. Res. 1999, 48, 70–75. [Google Scholar] [CrossRef]
- Romay, C.; Ledón, N.; González, R. Phycocyanin extract reduces leukotriene B4 levels in arachidonic acid-induced mouse-ear inflammation test. J. Pharm. Pharmacol. 1999, 51, 641–642. [Google Scholar] [CrossRef] [PubMed]
- Reddy, C.M.; Bhat, V.B.; Kiranmai, G.; Reddy, M.N.; Reddanna, P.; Madyastha, K.M. Selective inhibition of cyclooxygenase-2 by C-phycocyanin, a biliprotein from Spirulina platensis. Biochem. Biophys. Res. Commun. 2000, 277, 599–603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bao, X.Q.; Huang, Y.C.; Chen, F. C-Phycocyanin alleviates bladder inflammation and dysfunction in cyclophosphamide-induced cystitis in a mouse model by inhibiting COX-2 and EP4. Evid. Based Complement. Alternat. Med. 2019, 2019, 8424872. [Google Scholar] [CrossRef] [Green Version]
- Soni, B.; Visavadiya, N.P.; Madamwar, D. Attenuation of diabetic complications by C-phycoerythrin in rats: Antioxidant activity of C-phycoerythrin including copper-induced lipoprotein and serum oxidation. Br. J. Nutr. 2009, 102, 102–109. [Google Scholar] [CrossRef] [Green Version]
- Ou, Y.; Lin, L.; Yang, X.; Pan, Q.; Cheng, X. Antidiabetic potential of phycocyanin: Effects on KKAy mice. Pharm. Biol. 2013, 51, 539–544. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Inoguchi, T.; Sasaki, S.; Maeda, Y.; McCarty, M.F.; Fujii, M.; Ikeda, N.; Kobayashi, K.; Sonoda, N.; Takayanagi, R. Phycocyanin and phycocyanobilin from spirulina platensis protect against diabetic nephropathy by inhibiting oxidative stress. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2013, 304, 110–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munawaroh, H.; Gumilar, G.G.; Nurjanah, F.; Yuliani, G.; Show, P.L. In-vitro molecular docking analysis of microalgae extracted phycocyanin as an anti-diabetic candidate. Biochem. Eng. J. 2020, 161, 107666. [Google Scholar] [CrossRef]
- Rimbau, V.; Camins, A.; Romay, C.; González, R.; Pallàs, M. Protective effects of C-phycocyanin against kainic acid-induced neuronal damage in rat hippocampus. Neurosci. Lett. 1999, 276, 75–78. [Google Scholar] [CrossRef]
- Rimbau, V.; Camins, A.; Pubill, D.; Sureda, F.X.; Romay, C.; González, R.; Jiménez, A.; Escubedo, E.; Camarasa, J.; Pallás, M.A. C-phycocyanin protects cerebellar granule cells from low potassium/serum deprivation-induced apoptosis. Naunyn Schmiedebergs Arch. Pharmacol. 2001, 364, 96–104. [Google Scholar] [CrossRef]
- Pentón-Rol, G.; Martínez-Sánchez, G.; Cervantes-Llanos, M.; Lagumersindez-Denis, N.; Acosta-Medina, E.F.; Falcón-Cama, V.; Alonso-Ramírez, R.; Valenzuela-Silva, C.; Rodríguez-Jiménez, E.; Llópiz-Arzuaga, A.; et al. C-Phycocyanin ameliorates experimental autoimmune encephalomyelitis and induces regulatory T cells. Int. Immunopharmacol. 2011, 11, 29–38. [Google Scholar] [CrossRef]
- Marín-Prida, J.; Pent´on-Rol, G.; Rodrigues, F.P.; Alberici, L.C.; Stringhetta, K.; Leopoldino, A.M.; Rosa, M.N. C-Phycocyanin protects SH-SY5Y cells from oxidative injury, rat retina from transient ischemia and rat brain mitochondria from Ca2+/phosphate-induced impairment. Brain Res. Bull. 2012, 89, 159–167. [Google Scholar] [CrossRef] [PubMed]
- Soni, B.; Visavadiya, N.P.; Madamwar, D. Ameliorative action of cyanobacterial phycoerythrin on CCl4-induced toxicity in rats. Toxicology 2008, 248, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Ou, Y.; Zheng, S.; Lin, L.; Jiang, Q.; Yang, X. Protective effect of C-phycocyanin against carbon tetrachloride-induced hepatocyte damage in vitro and in vivo. Chem. Biol. Interact. 2010, 185, 94–100. [Google Scholar] [CrossRef] [PubMed]
- Iijima, N.; Fujjii, N. Antitumor Agent. JP58065216, 26 May 1982. [Google Scholar]
- Nemoto-Kawamura, C.; Hirahashi, T.; Nagai, T.; Yamada, H.; Katoh, T.; Hayashi, O. Phycocyanin enhances secretary IgA antibody response and suppresses allergic IgE antibody response in mice immunized with antigen-entrapped biodegradable microparticles. J. Nutr. Sci. Vitaminol. 2004, 50, 129–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivanova, K.G.; Stankova, K.G.; Nikolov, V.N.; Georgieva, R.T.; Minkova, K.M.; Gigova, L.G.; Rupova, I.T.; Boteva, R.N. The biliprotein C-phycocyanin modulates the early radiation response: A pilot study. Mutat. Res. 2010, 695, 40–45. [Google Scholar] [CrossRef]
- Lee, D.; Nishizawa, M.; Shimizu, Y.; Saeki, H. Anti-inflammatory effects of dulse (Palmaria palmate) resulting from the simultaneous water-extraction of phycobiliproteins and chlorophyll a. Food. Res. Int. 2017, 100, 514–521. [Google Scholar] [CrossRef]
- Chen, H.W.; Yang, T.S.; Chen, M.J.; Chang, Y.C.; Wang, I.C.; Ho, C.L.; Lai, Y.J.; Yu, C.C.; Chou, J.C.; Chao, L.K.; et al. Purification and immunomodulating activity of C-phycocyanin from Spirulina platensis cultured using power plant flue gas. Process. Biochem. 2014, 49, 1337–1344. [Google Scholar] [CrossRef] [Green Version]
- Grover, P.; Bhatnagar, A.; Kumari, N.; Bhatt, A.N.; Nishad, D.K.; Purkayastha, J. C-Phycocyanin a novel protein from Spirulina platensis-in vivo toxicity, antioxidant and immunomodulatory studies. Saudi J. Biol. Sci. 2021, 28, 1853–1859. [Google Scholar] [CrossRef]
- McCarty, M.F. Clinical potential of phycocyanobilin for induction of T regulatory cells in the management of inflammatory disorders. Med. Hypotheses. 2011, 77, 1031–1033. [Google Scholar] [CrossRef]
- Dabrowski, J.M.; Arnaut, L.G. Photodynamic therapy of Cancer: From a local to a systemic treatment. Photochem. Photobiol. Sci. 2015, 14, 1765–1780. [Google Scholar] [CrossRef]
- Mfouo-Tynga, I.S.; Dias, L.D.; Inada, N.M.; Kurachi, C. Biophysical and biological features of third generation photosensitizers used in anticancer photodynamic therapy: Review. Photodiagn. Photodyn. Ther. 2021, 1, 102091. [Google Scholar] [CrossRef] [PubMed]
- Morcos, N.C.; Berns, M.; Henry, W.L. Phycocyanin: Laser activation, cytotoxic effects, and uptake in human atherosclerotic plaque. Lasers Surg. Med. 1988, 8, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Chu, X.; Gao, M.; Li, W. Apoptotic mechanism of MCF-7 breast cells in vivo and in vitro induced by photodynamic therapy with C-phycocyanin. Acta Biochim. Biophys. Sin. 2010, 42, 80–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bharathiraja, S.; Seo, H.; Manivasagan, P.; Santha, M.M.; Park, S.; Oh, J. In vitro photodynamic effect of phycocyanin against breast cancer cells. Molecules 2016, 21, 1470. [Google Scholar] [CrossRef]
- Pu, Y.; Wei, M.; Witkowski, A.; Krzywda, M.; Wang, Y.; Li, W. A hybrid biomaterial of biosilica and C-phycocyanin for enhanced photodynamic effect towards tumor cells. Biochem. Biophys. Res. Commun. 2020, 533, 573–579. [Google Scholar] [CrossRef] [PubMed]
- Huang, B.; Wang, G.C.; Zeng, C.K.; Li, Z.G. The experimental research of R-phycoerythrin subunits on cancer treatment: A new photosensitizer in PDT. Cancer Biother. Radiopharm. 2002, 17, 35–42. [Google Scholar]
- Zhang, S.P.; Pan, J.X.; Han, Z.H.; Zhao, J.Q.; Yao, S.D.; Jiang, L.J. Generation and identification of the transient intermediates of allophycocyanin by laser photolytic and pulse radiolytic techniques. Int. J. Radiat. Biol. 2001, 77, 637–642. [Google Scholar]
- Chueh, C. Method of Allophycocyanin Inhibition of Enterovirus and Influenza Virus Reproduction Resulting in Cytopathic Effect. U.S. Patent 6346408, 12 February 2002. [Google Scholar]
- Shih, S.R.; Tsai, K.N.; Li, Y.S.; Chueh, C.C.; Chan, E.C. Inhibition of enterovirus 71-induced apoptosis by allophycocyanin isolated from a blue-green alga Spirulina platensis. J. Med. Virol. 2003, 70, 119–125. [Google Scholar] [CrossRef]
- Qi, H.; Liu, Y.; Qi, X.; Liang, H.; Chen, H.; Jiang, P.; Wang, D. Dietary recombinant phycoerythrin modulates the gut microbiota of H22 tumor-bearing mice. Mar. Drugs 2019, 26, 665. [Google Scholar] [CrossRef] [Green Version]
- Dev, A.; Mohanbhai, S.J.; Kushwaha, A.C.; Sood, A.; Sardoiwala, M.N.; Choudhury, S.R.; Karmakar, S. κ-carrageenan-C-phycocyanin based smart injectable hydrogels for accelerated wound recovery and real-time monitoring. Acta Biomater. 2020, 109, 121–131. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, H.; Qi, H.; Xiong, P. Phycobiliproteins—A Family of Algae-Derived Biliproteins: Productions, Characterization and Pharmaceutical Potentials. Mar. Drugs 2022, 20, 450. https://doi.org/10.3390/md20070450
Chen H, Qi H, Xiong P. Phycobiliproteins—A Family of Algae-Derived Biliproteins: Productions, Characterization and Pharmaceutical Potentials. Marine Drugs. 2022; 20(7):450. https://doi.org/10.3390/md20070450
Chicago/Turabian StyleChen, Huaxin, Hongtao Qi, and Peng Xiong. 2022. "Phycobiliproteins—A Family of Algae-Derived Biliproteins: Productions, Characterization and Pharmaceutical Potentials" Marine Drugs 20, no. 7: 450. https://doi.org/10.3390/md20070450