
Asperopiperazines A and B: Antimicrobial and Cytotoxic Dipeptides from a Tunicate-Derived Fungus Aspergillus sp. DY001



Abstract
:1. Introduction
2. Results and Discussion
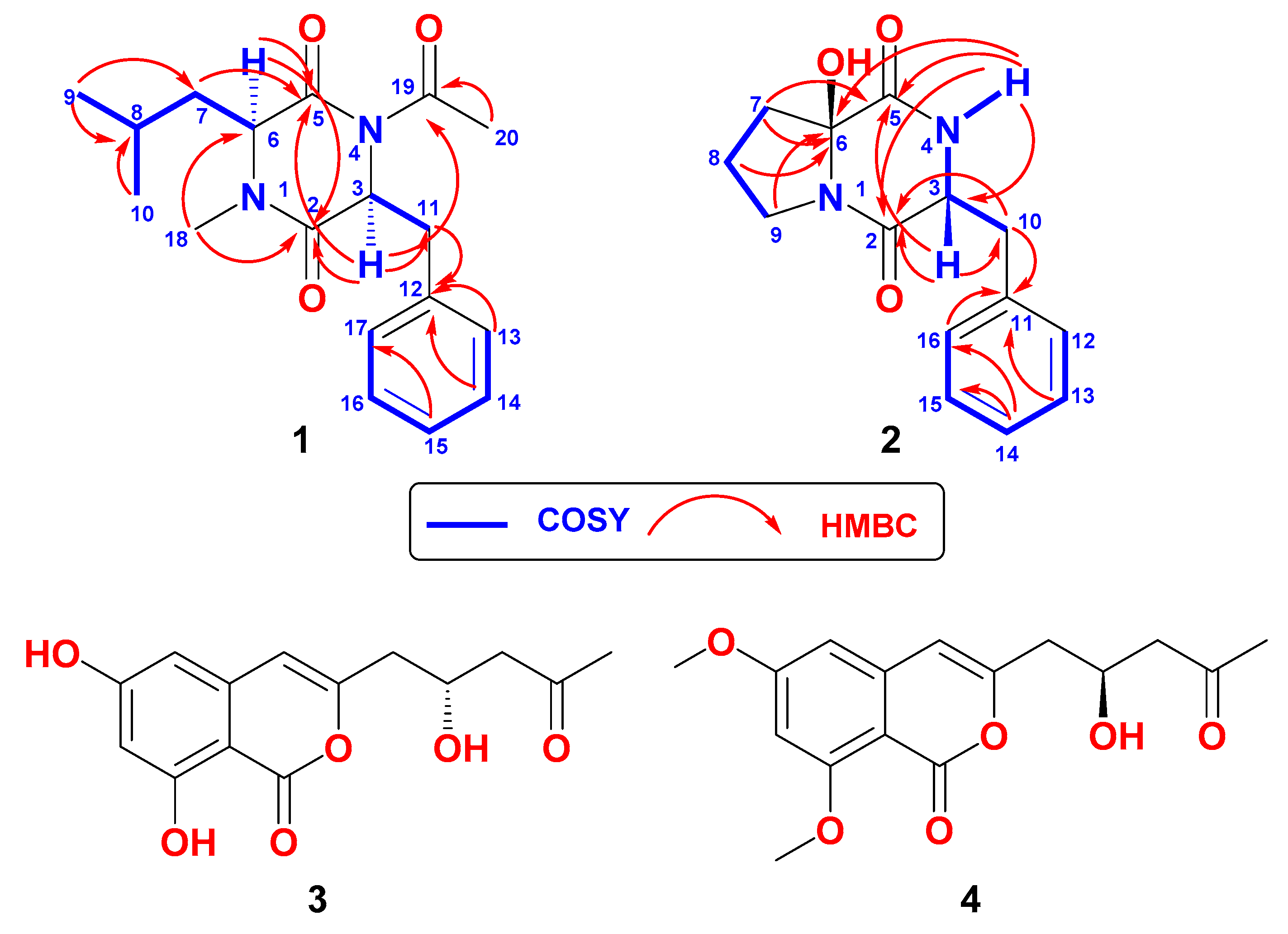
2.1. Structure of Asperopiperazine A (1)
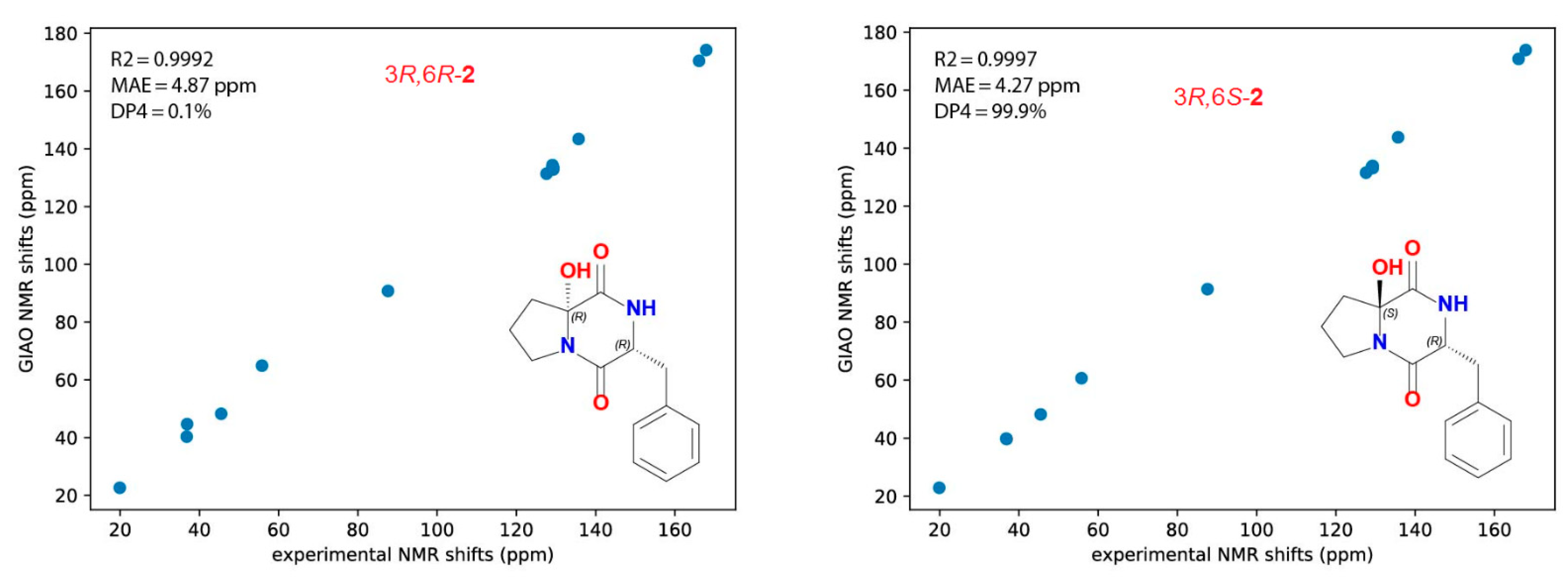
2.2. Structure of Asperopiperazine B (2)
2.3. Biological Activities of the Compounds
3. Materials and Methods
3.1. General Experimental Procedures
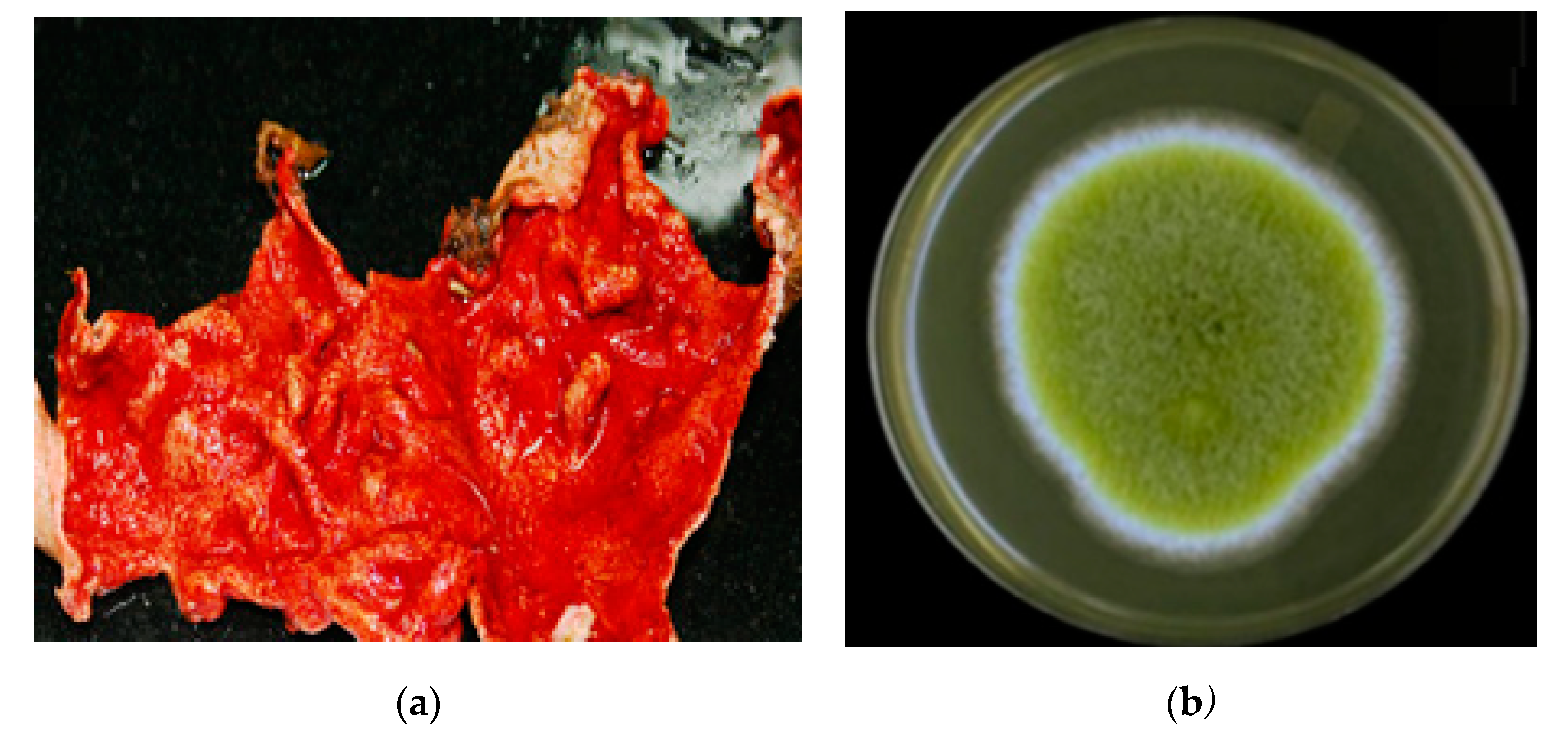
3.2. The Host Organism, Didemnum Species
3.3. Purification of the Fungal Isolate
3.4. Purification of gDNA from Fungal Isolates
3.5. Fungal ITS-rDNA Fragments’ Amplification
3.6. Sequence of ITS-rDNA Fungal Region
3.7. Characterization of the Fungal Isolate
3.8. Large-Scale Culture of Aspergillus sp. DY001
3.9. Purification of Compounds 1–4
3.10. Spectral Data of the Compounds
3.10.1. Asperopiperazine A (1)
3.10.2. Asperopiperazine B (2)
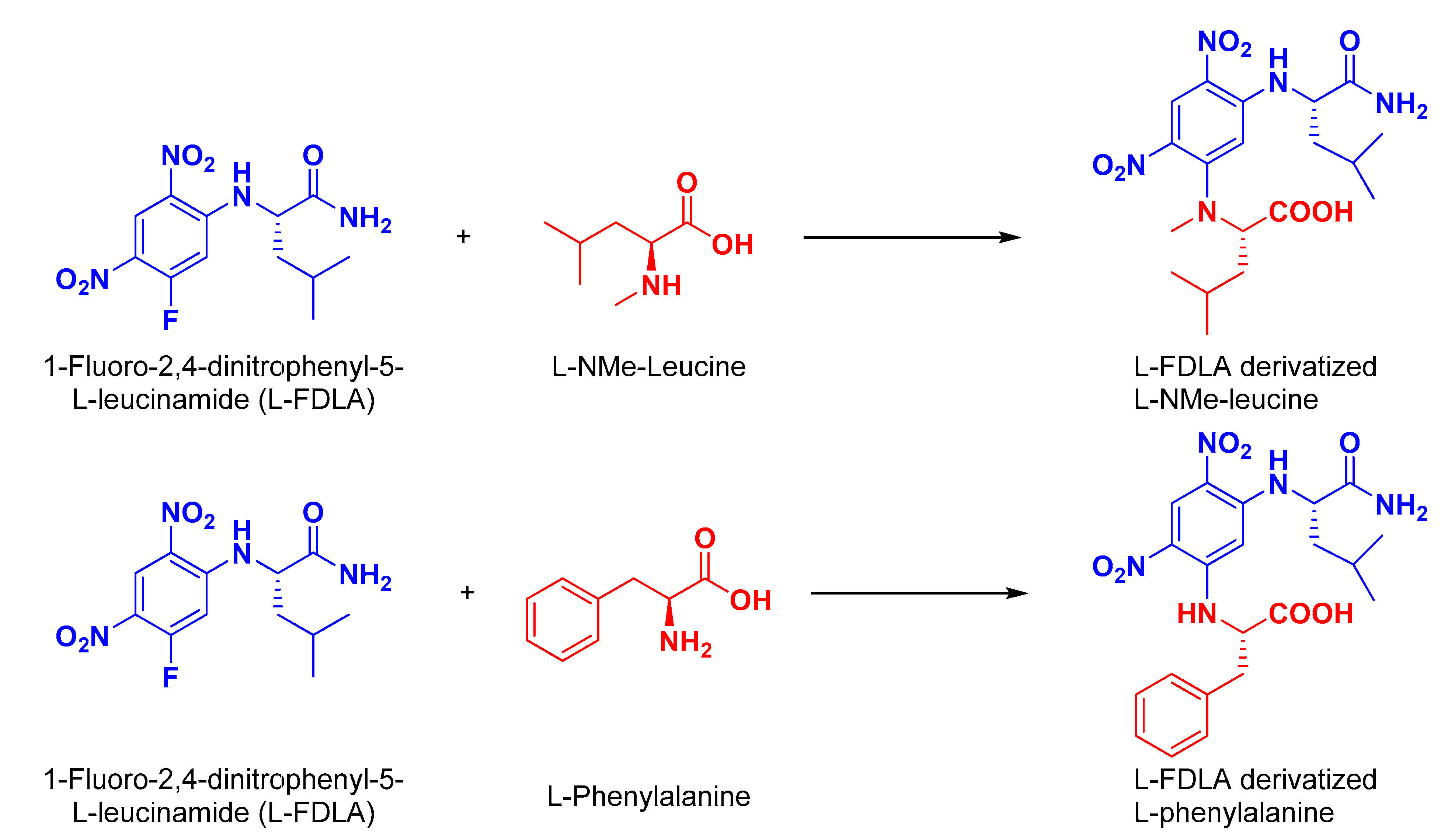
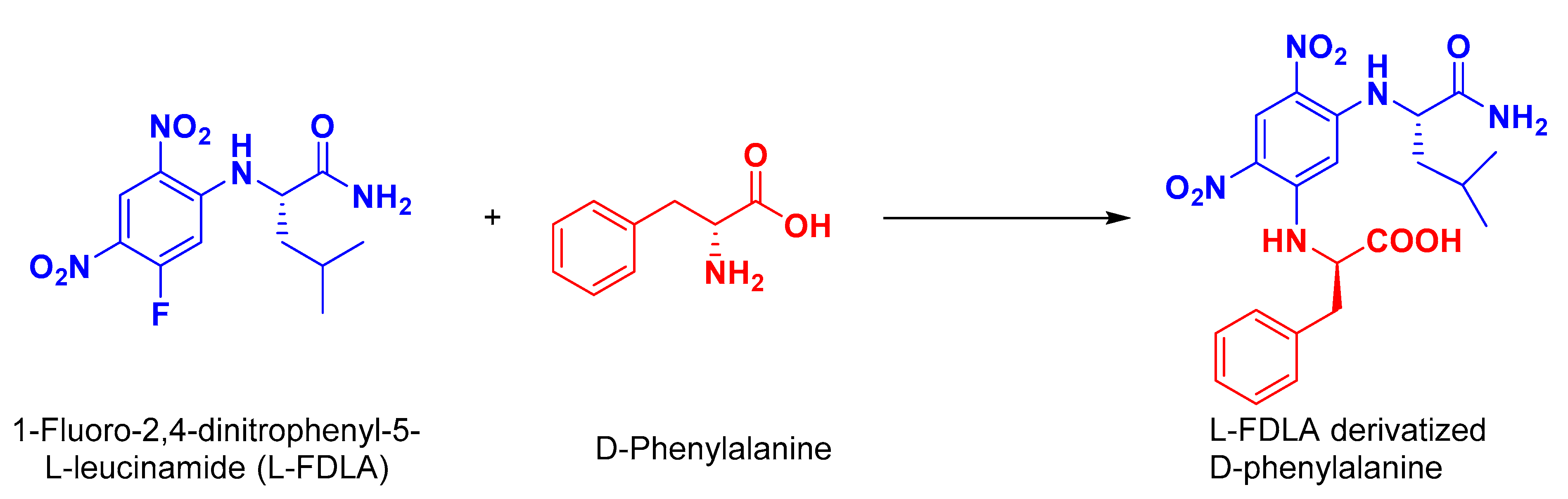
3.11. Absolute Stereochemistry of 1 and 2
3.11.1. Preparation of l-FDLA Derivatives for HPLC Analyses
3.11.2. HPLC Analysis of FDLA Derivatives
3.11.3. Computational Details
3.12. Biological Evaluation of the Compounds
3.12.1. Evaluation of the Antimicrobial Effects of 1–4
Disk Diffusion Assay
Evaluation of the MIC Values
3.12.2. Evaluation of the Growth Inhibition Effects of the Compounds
Culture of Cell Lines
MTT Assay
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pereira, F. Have marine natural product drug discovery efforts been productive and how can we improve their efficiency? Expert Opin. Drug Dis. 2019, 14, 717–722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, F.; Aires-de-Sousa, J. Computational methodologies in the exploration of marine natural product leads. Mar. Drugs 2018, 16, 236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the 30 years from 1981 to 2010. J. Nat. Prod. 2012, 75, 311–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newman, D.J.; Giddings, L.-A. Natural products as leads to antitumor drug. Phytochem. Rev. 2014, 13, 123–137. [Google Scholar] [CrossRef]
- Youssef, D.T.A.; Almagthali, H.; Shaala, L.A.; Schmidt, E.W. Secondary metabolites of the genus Didemnum: A comprehensive review of chemical diversity and pharmacological properties. Mar. Drugs 2020, 18, 307. [Google Scholar] [CrossRef]
- Bailly, C. Anticancer properties of lamellarins. Mar. Drugs 2015, 13, 1105–1123. [Google Scholar] [CrossRef] [Green Version]
- Oku, N.; Matsunaga, S.; Fusetani, N. Shishijimicins A−C, novel enediyne antitumor antibiotics from the ascidian Didemnum proliferum. J. Am. Chem. Soc. 2003, 125, 2044–2045. [Google Scholar] [CrossRef]
- Schmidt, E.W.; Donia, M.S. Life in cellulose houses: Symbiotic bacterial biosynthesis of ascidian drugs and drug leads. Curr. Opin. Biotechnol. 2010, 21, 827–833. [Google Scholar] [CrossRef] [Green Version]
- Ireland, C.M.; Fernandez, A. Cyclic Peptide Antitumor Agent from an Ascidian. Ph.D. Thesis, University of Utah, Salt Lake City, UT, USA, 1998. [Google Scholar]
- Smith, T.E.; Pond, C.D.; Pierce, E.; Harmer, Z.P.; Kwan, J.; Zachariah, M.M.; Harper, M.K.; Wyche, T.P.; Matainaho, T.K.; Bugni, T.S. Accessing chemical diversity from the uncultivated symbionts of small marine animals. Nat. Chem. Biol. 2018, 14, 179. [Google Scholar] [CrossRef]
- Ruegger, A.; Kuhn, M.; Lichti, H.; Loosli, H.R.; Huguenin, R.; Quiquerez, C.; von Wartburg, A. Cyclosporin A, a peptide metabolite from Trichoderma polysporum (Link ex Pers.) Rifai, with a remarkable immunosuppressive activity. Helv. Chim. Acta 1976, 59, 1075–1092. [Google Scholar] [CrossRef]
- Endo, A.; Monacolin, K. A new hypocholesterolemic agent produced by a Monascus species. J. Antibiot. 1979, 32, 852–854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abraham, E.P.; Newton, G.G.; Crawford, K.; Burton, H.S.; Hale, C.W. Cephalosporin N: A new type of penicillin. Nature 1953, 171, 343. [Google Scholar] [CrossRef] [PubMed]
- Nesbitt, B.F.; O’Kelly, J.; Sargeant, K.; Sheridan, A. Aspergillus flavus and turkey X disease. Toxic metabolites of Aspergillus flavus. Nature 1962, 195, 1062–1063. [Google Scholar] [CrossRef] [PubMed]
- Flieger, M.; Wurst, M.; Shelby, R. Ergot alkaloids sources, structures, and analytical methods. Folia Microbiol. 1997, 42, 3–29. [Google Scholar]
- Freeman, G.G.; Gill, J.E. Alkaline hydrolysis of trichothecin. Nature 1950, 166, 698–699. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Wu, W.; Liu, X.; Zaleta-Pinet, D.A.; Clark, B.R. Bioactive compounds isolated from marine-derived microbes in China: 2009–2018. Mar. Drugs 2019, 17, 339. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Zheng, Y.-Y.; Shao, C.-L.; Wang, C.-Y. Metabolites from marine invertebrates and their symbiotic microorganisms: Molecular diversity discovery, mining, and application. Mar. Life Sci. Technol. 2019, 1, 60–94. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Tang, S.; Cao, S. Antimicrobial compounds from marine fungi. Phytochem. Rev. 2021, 20, 85–117. [Google Scholar] [CrossRef]
- Wu, B.; Wiese, J.; Labes, A.; Kramer, A.; Schmaljohann, R.; Imhoff, J.F. Lindgomycin, an unusual antibiotic polyketide from a marine fungus of the Lindgomycetaceae. Mar. Drugs 2015, 13, 4617–4632. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Meng, W.; Cao, C.; Wang, J.; Shan, W.; Wang, Q. Antibacterial and antifungal compounds from marine fungi. Mar. Drugs 2015, 13, 3479–3513. [Google Scholar] [CrossRef]
- Orfali, R.; Aboseada, M.A.; Abdel-Wahab, N.M.; Hassan, H.M.; Perveen, S.; Ameen, F.; Abdelmohsen, U.R. Recent updates on the bioactive compounds of the marine-derived genus Aspergillus. RSC Adv. 2021, 11, 17116–17150. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, H.; Gigant, B.; Yu, Y.; Wu, Y.; Chen, X.; Lai, Q.; Yang, Z.; Chen, Q.; Yang, J. Structures of a diverse set of colchicine binding site inhibitors in complex with tubulin provide a rationale for drug discovery. FEBS J. 2016, 83, 102–111. [Google Scholar] [CrossRef] [PubMed]
- Prota, A.E.; Danel, F.; Bachmann, F.; Bargsten, K.; Buey, R.M.; Pohlmann, J.; Reinelt, S.; Lane, H.; Steinmetz, M.O. The novel microtubule-destabilizing drug BAL27862 binds to the colchicine site of tubulin with distinct effects on microtubule organization. J. Mol. Biol. 2014, 426, 1848–1860. [Google Scholar] [CrossRef] [PubMed]
- Blayney, D.W.; Zhang, Q.; Feng, J.; Zhao, Y.; Bondarenko, I.; Vynnychenko, I.; Kovalenko, N.; Nair, S.; Ibrahim, E.; Udovista, D.P.; et al. Efficacy of Plinabulin vs Pegfilgrastim for prevention of chemotherapy-induced neutropenia in adults with non–small cell lung cancer. A Phase 2 Randomized Clinical Trial. JAMA Oncol. 2020, 6, e204429. [Google Scholar] [CrossRef]
- Huggett, B. Innovation’ nation. Nat. Biotechnol. 2019, 37, 1264–1276. [Google Scholar] [CrossRef] [Green Version]
- Shaala, L.A.; Alzughaibi, T.; Genta-Jouve, G.; Youssef, D.T.A. Fusaripyridines A and B; Highly oxygenated antimicrobial alkaloid dimers featuring an unprecedented 1,4-bis(2-hydroxy-1,2-dihydropyridin-2-yl)butane-2,3-dione core from the marine fungus Fusarium sp. LY019. Mar. Drugs 2021, 19, 505. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, A.; Ono, Y.; Fujii, I.; Sankawa, U.; Mayorga, M.E.; Timberlake, W.E.; Ebizuka, Y. Product identification of polyketide synthase coded by Aspergillus nidulans wA gene. Tetrahedron Lett. 1998, 39, 7733–7736. [Google Scholar] [CrossRef]
- Lai, S.; Shizuri, Y.; Yamamura, S.; Kawai, K.; Furukawa, H. Three new phenolic metabolites from Penicillium species. Heterocycles 1991, 32, 297–305. [Google Scholar]
- Sun, K.; Li, Y.; Guo, L.; Wang, Y.; Liu, P.; Zhu, W. Indole diterpenoids and isocoumarin from the fungus, Aspergillus flavus, isolated from the Prawn, Penaeus vannamei. Mar. Drugs 2014, 12, 3970–3981. [Google Scholar] [CrossRef] [Green Version]
- Shaala, L.A.; Youssef, D.T.A.; Badr, J.M.; Harakeh, S.M.; Genta-Jouve, G. Bioactive diketopiperazines and nucleoside derivatives from a sponge-derived Streptomyces species. Mar. Drugs 2019, 17, 584. [Google Scholar] [CrossRef] [Green Version]
- Marfey, P. Determination of D-amino acids. II. Use of a bifunctional reagent, 1,5-difluoro-2,4-dinitrobenzene. Carlsberg Res. Commun. 1984, 49, 591–596. [Google Scholar] [CrossRef] [Green Version]
- Park, Y.C.; Gunasekera, S.P.; Lopez, J.V.; McCarthy, P.J.; Wright, A.E. Metabolites from the marine-derived fungus Chromocleista sp. isolated from a deep-water sediment sample collected in the Gulf of Mexico. J. Nat. Prod. 2006, 69, 580–584. [Google Scholar] [PubMed] [Green Version]
- Shaala, L.A.; Youssef, D.T.A. Identification and bioactivity of compounds from the fungus Penicillium sp. CYE-87 isolated from a marine tunicate. Mar. Drugs 2015, 13, 1698–1709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In PCR Protocols: A Guide to Methods and Application; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- National Center for Biotechnology Information. Available online: http://www.ncbi.nlm.nih.gov (accessed on 1 December 2019).
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The ClustalX windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [Green Version]
- Hall, T.A. BioEdit: A user friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [Green Version]
- Frisch, M.J.; Trucks, G.W.; Schlegel, H.B.; Scuseria, G.E.; Robb, M.A.; Cheeseman, J.R.; Scalmani, G.; Barone, V.; Mennucci, B.; Petersson, G.A.; et al. G09a: Gaussian 09; Revision A.02; Gaussian Inc.: Wallingford, CT, USA, 2009. [Google Scholar]
- Smith, S.G.; Goodman, J.M. Assigning stereochemistry to single diastereoisomers by GIAO NMR calculation:The DP4 Probability. J. Am. Chem. Soc. 2010, 132, 12946–12959. [Google Scholar] [CrossRef]
- Acar, J.F. The disc susceptibility test. In Antibiotics in Laboratory Medicine; Lorian, V., Ed.; Williams and Wilkins: Baltimore, MD, USA, 1980; pp. 24–54. [Google Scholar]
- Kiehlbauch, J.A.; Hannett, G.E.; Salfinger, M.; Archinal, W.; Monserrat, C.; Carlyn, C. Use of the National Committee for Clinical Laboratory Standards Guidelines for Disk Diffusion Susceptibility Testing in New York State Laboratories. J. Clin. Microbiol. 2000, 38, 3341–3348. [Google Scholar] [CrossRef] [Green Version]
- CLSI, Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Disk Susceptibility Tests, 9th ed.; CLSI Documents M07-A9; West Valley Road, Suite 2500: Wayne, PA, USA, 2007. [Google Scholar]
- Youssef, D.T.A.; Shaala, L.A.; Altyar, A.E. Cytotoxic phenylpropanoid derivatives and alkaloids from the flowers of Pancratium maritimum L. Plants 2022, 11, 476. [Google Scholar] [CrossRef]
- Shaala, L.A.; Youssef, D.T.A. Cytotoxic psammaplysin analogues from the Verongid Red Sea sponge Aplysinella species. Biomolecules 2019, 9, 841. [Google Scholar] [CrossRef] [Green Version]
- Youssef, D.T.A.; Mooberry, S.L. Hurghadolide A and swinholide I, potent actin-microfilament disrupters from the Red Sea sponge Theonella swinhoei. J. Nat. Prod. 2006, 69, 154–157. [Google Scholar] [CrossRef] [PubMed]
- Shaala, L.A.; Youssef, D.T.A. Hemimycalins, C.-E: Cytotoxic and antimicrobial alkaloids with hydantoin and 2-iminoimidazolidin4-one backbones from the Red Sea marine sponge Hemimycale sp. Mar. Drugs 2021, 19, 691. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Position | 1 (CD3OD) a | 2 (CDCl3) b | ||
|---|---|---|---|---|
| δC (mult.) c | δH (mult. J (Hz)) | δC (mult.) c | δH (mult. J (Hz)) | |
| 2 | 174.5, C | 166.1, C | ||
| 3 | 52.6, CH | 5.00 (t, 7.8) | 55.8, CH | 4.45 (dd, 11.0, 3.3) |
| 4 (NH) | - | 5.58 (brs) | ||
| 5 | 175.2, C | 167.9, C | ||
| 6 | 56.3, CH | 5.04 (dd, 9.6, 6.0) | 87.6, C | |
| 6-OH | 2.98 (brs) | |||
| 7 | 38.1, CH2 | 1.62 (m) | 36.8, CH2 | 2.23 (m), 2.17 (m) |
| 8 | 25.8, CH | 1.42 (m) | 19.9, CH2 | 2.17 (m), 2.00 (m) |
| 9 | 23.6, CH3 | 0.91 (d, 6.6) | 45.5, CH2 | 3.79 (m), 3.59 (m) |
| 10 | 22.0, CH3 | 0.85 (d, 6.6) | 36.9, CH2 | 3.63 (dd, 14.4, 4.2) 2.74 (dd, 14.4, 10.2) |
| 11 | 38.7, CH2 | 3.06 (dd, 13.2, 7.6) 2.93 (dd, 13.2, 7.2) | 135.7, C | |
| 12 | 138.0, C | 129.1, CH | 7.23 (d, 7.5) | |
| 13 | 129.6, CH | 7.25 (m) | 129.3, CH | 7.35 (t, 7.5) |
| 14 | 130.4, CH | 7.25 (m) | 127.6, CH | 7.29 (t, 7.5) |
| 15 | 128.1, CH | 7.21 (m) | 129.3, CH | 7.35 (t, 7.5) |
| 16 | 130.4, CH | 7.25 (m) | 129.1, CH | 7.23 (d, 7.5) |
| 17 | 129.6, CH | 7.25 (m) | ||
| 18 | 31.7, CH3 | 2.84 (s) | ||
| 19 | 172.8, C | |||
| 20 | 22.1, CH3 | 1.89 (s) | ||
| Carbon No. | Compound 2 | Cyclo(6-OH-d-Pro-l-Phe) [33] | ΔδC (ppm) |
|---|---|---|---|
| δC (CDCl3) | δC (CDCl3) | ||
| C-6 | 87.6 | 87.6 | 0.0 |
| C-7 | 36.8 | 36.8 | 0.0 |
| C-8 | 19.9 | 19.9 | 0.0 |
| C-9 | 45.5 | 45.4 | −0.1 |
| Compound No. | C. albicans | E. coli | S. aureus | |||
|---|---|---|---|---|---|---|
| Inhibition Zone (mm) | MIC (μM) | Inhibition Zone (mm) | MIC (μM) | Inhibition Zone (mm) | MIC (μM) | |
| 1 | 11 | 16 | 17 | 8 | 16 | 8 |
| 2 | 12 | 16 | 23 | 4 | 18 | 8 |
| 3 | 17 | 8 | 11 | 8 | 19 | 8 |
| 4 | 9 | 16 | 7 | 16 | 6 | 32 |
| Ciprofloxacin a | NT | 30 | 0.08 | 22 | ||
| Ketoconazole b | 30 | 0.26 | NT | NT | NT | |
| Compound | IC50 (μM) a | ||
|---|---|---|---|
| MDA-MB-231 | HeLa | HCT 116 | |
| 1 | 24.3 ± 0.2 | ≥50.0 | 15.1 ± 0.1 |
| 2 | 26.3 ± 0.3 | ≥50.0 | 16.2 ± 0.1 |
| 3 | 31.0 ± 0.2 | ≥50.0 | 19.3 ± 0.1 |
| 4 | 35.0 ± 0.2 | ≥50.0 | 17.7 ± 0.1 |
| 5-FU b | 13.0 ± 0.3 | 12.3 ± 0.2 | 4.6 ± 0.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Youssef, D.T.A.; Shaala, L.A.; Genta-Jouve, G. Asperopiperazines A and B: Antimicrobial and Cytotoxic Dipeptides from a Tunicate-Derived Fungus Aspergillus sp. DY001. Mar. Drugs 2022, 20, 451. https://doi.org/10.3390/md20070451
Youssef DTA, Shaala LA, Genta-Jouve G. Asperopiperazines A and B: Antimicrobial and Cytotoxic Dipeptides from a Tunicate-Derived Fungus Aspergillus sp. DY001. Marine Drugs. 2022; 20(7):451. https://doi.org/10.3390/md20070451
Chicago/Turabian StyleYoussef, Diaa T. A., Lamiaa A. Shaala, and Grégory Genta-Jouve. 2022. "Asperopiperazines A and B: Antimicrobial and Cytotoxic Dipeptides from a Tunicate-Derived Fungus Aspergillus sp. DY001" Marine Drugs 20, no. 7: 451. https://doi.org/10.3390/md20070451