
Design, Synthesis and Evaluation of Novel Phorbazole C Derivatives as MNK Inhibitors through Virtual High-Throughput Screening


Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
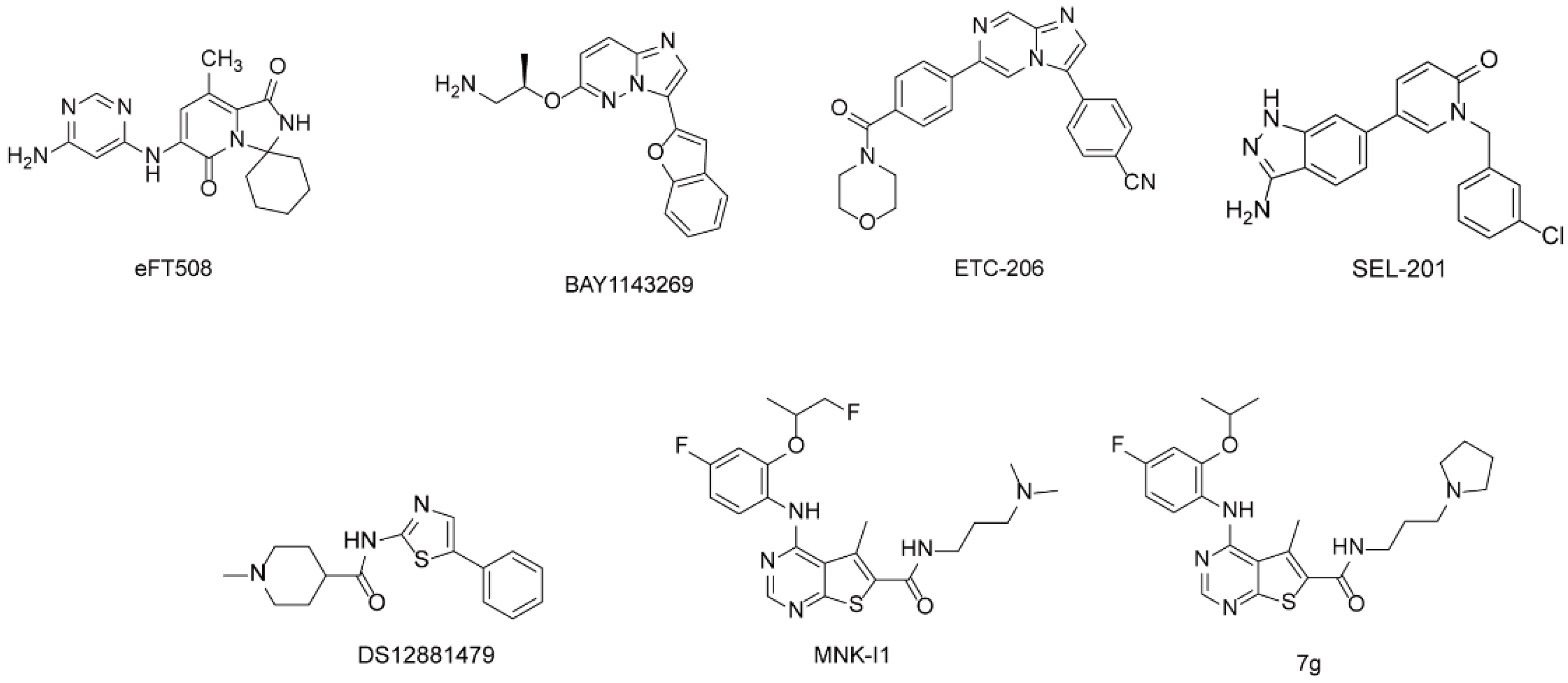
2.1. The Discovery of Hit Compounds
2.2. Design and Synthesis of Derivatives
2.3. The Structure–Activity Relationship (SAR) of Derivatives
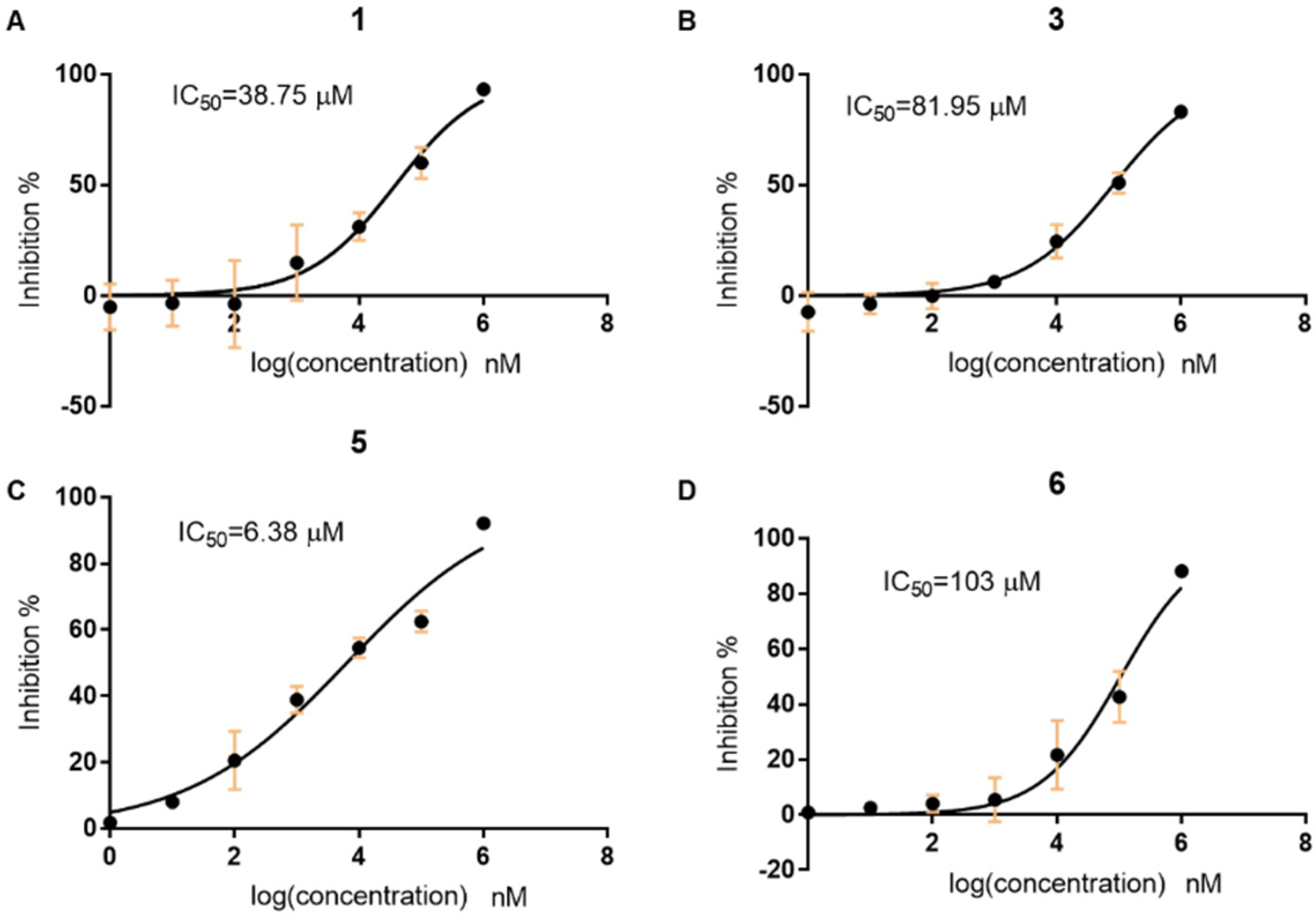
2.4. Evaluation the Activity of the Compounds
3. Discussion
4. Materials and Methods
4.1. Reagents and Tumor Cell Lines
4.2. SDS-PAGE and Western Blot
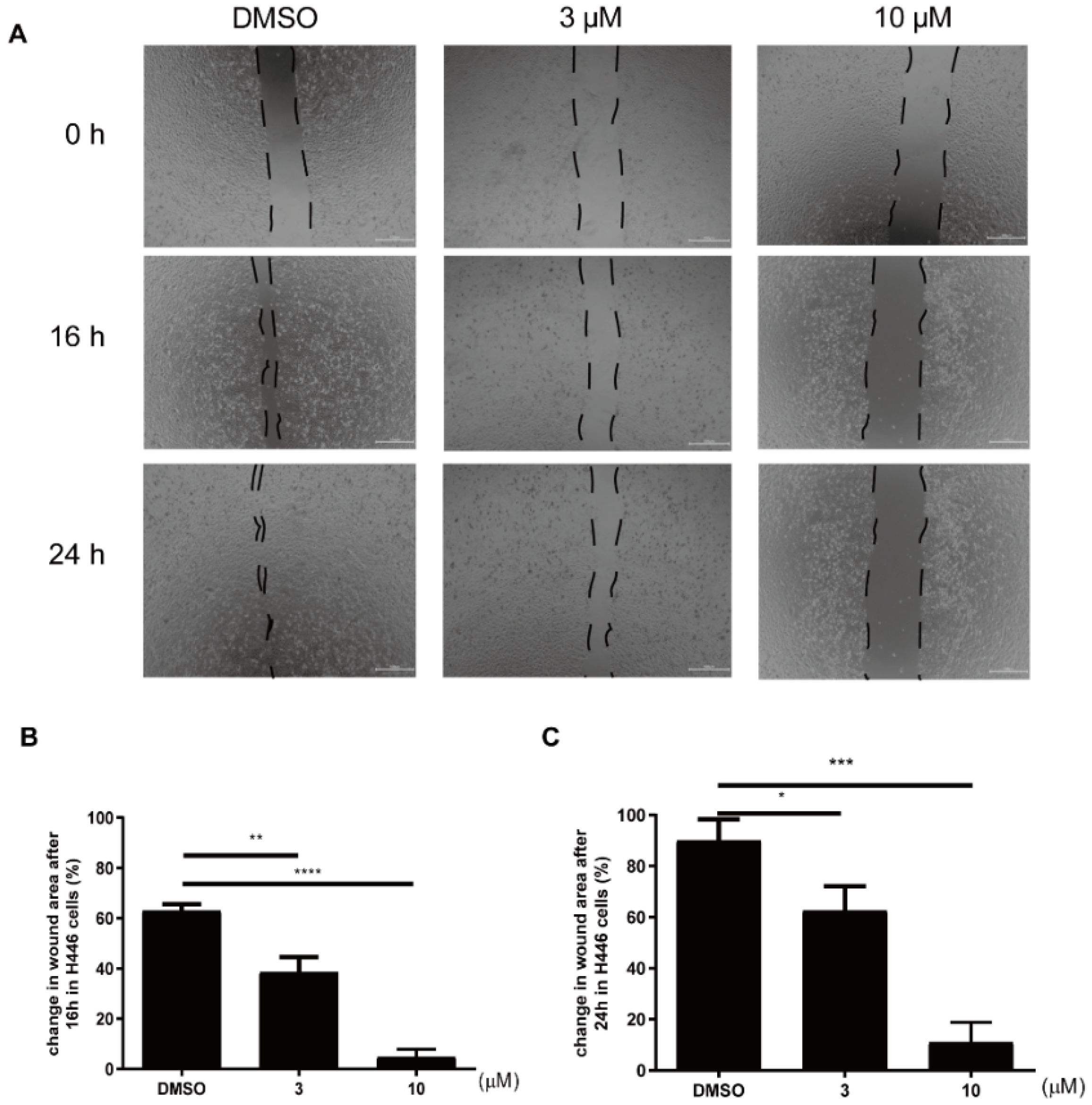
4.3. Scratch Wound-Healing Assay
4.4. MTT Assay
4.5. Molecular Docking
4.6. General Methods for Synthetic Chemistry
4.6.1. General Synthesis Method for Compounds 2A and 9A–9E
4.6.2. General Synthesis Method for 3A–3F and 10A–10E
4.6.3. General Synthesis Method for 4A–4F and 11A–11E
4.6.4. General Synthesis Method for 5A–5V and 12A–12G
4.6.5. General Synthesis Method for 6A–6V and 13A–13G
4.6.6. General Synthesis Method for 7A–7C, 10, 12–29 and 14A–14G
4.6.7. General Synthesis Method for 1–9 and 11
4.7. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hay, N. Mnk Earmarks Eif4e for Cancer Therapy. Proc. Natl. Acad. Sci. USA 2010, 107, 13975–13976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bramham, C.R.; Jensen, K.B.; Proud, C.G. Tuning Specific Translation in Cancer Metastasis and Synaptic Memory: Control at the Mnk-Eif4e Axis. Trends Biochem. Sci. 2016, 41, 847–858. [Google Scholar] [CrossRef]
- Jin, X.; Yu, R.; Wang, X.; Proud, C.G.; Jiang, T. Progress in Developing Mnk Inhibitors. Eur. J. Med. Chem. 2021, 219, 113420. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Kannan, S.; Verma, C.S.; Nacro, K. Update on the Development of Mnk Inhibitors as Therapeutic Agents. J. Med. Chem. 2022, 65, 983–1007. [Google Scholar] [CrossRef] [PubMed]
- Joshi, S.; Platanias, L.C. Mnk Kinase Pathway: Cellular Functions and Biological Outcomes. World J. Biol. Chem. 2014, 5, 321–333. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Merrett, J.E.; Jensen, K.B.; Proud, C.G. The Map Kinase-Interacting Kinases (Mnks) as Targets in Oncology. Expert Opin. Ther. Targets 2019, 23, 187–199. [Google Scholar] [CrossRef]
- Chu, J.; Pelletier, J. Therapeutic Opportunities in Eukaryotic Translation. Cold Spring Harb. Perspect. Biol. 2018, 10, a032995. [Google Scholar] [CrossRef]
- Topisirovic, I.; Ruiz-Gutierrez, M.; Borden, K.L. Phosphorylation of the Eukaryotic Translation Initiation Factor Eif4e Contributes to Its Transformation and Mrna Transport Activities. Cancer Res. 2004, 64, 8639–8642. [Google Scholar] [CrossRef] [Green Version]
- Robichaud, N.; del Rincon, S.V.; Huor, B.; Alain, T.; Petruccelli, L.A.; Hearnden, J.; Goncalves, C.; Grotegut, S.; Spruck, C.H.; Furic, L.; et al. Phosphorylation of Eif4e Promotes Emt and Metastasis Via Translational Control of Snail and Mmp-3. Oncogene 2015, 34, 2032–2042. [Google Scholar] [CrossRef] [Green Version]
- Carter, J.H.; Deddens, J.A.; Spaulding, I.V.; Lucas, D.N.R.; Colligan, B.M.; Lewis, T.G.; Hawkins, E.; Jones, J.; Pemberton, J.O.; Douglass, L.E.; et al. Phosphorylation of Eif4e Serine 209 Is Associated with Tumour Progression and Reduced Survival in Malignant Melanoma. Br. J. Cancer 2016, 114, 444–453. [Google Scholar] [CrossRef] [Green Version]
- Bianchini, A.; Loiarro, M.; Bielli, P.; Busa, R.; Paronetto, M.P.; Loreni, F.; Geremia, R.; Sette, C. Phosphorylation of Eif4e by Mnks Supports Protein Synthesis, Cell Cycle Progression and Proliferation in Prostate Cancer Cells. Carcinogenesis 2008, 29, 2279–2288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, S.; Ramalingam, S.S.; Kauh, J.; Xu, Z.; Khuri, F.R.; Sun, S.Y. Phosphorylated Eukaryotic Translation Initiation Factor 4 (Eif4e) Is Elevated in Human Cancer Tissues. Cancer Biol. Ther. 2009, 8, 1463–1469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, J.; Li, J.; Xu, L.; Xie, G.; Wen, Q.; Luo, J.; Li, D.; Huang, D.; Fan, S. Phosphorylated Mnk1 and Eif4e Are Associated with Lymph Node Metastasis and Poor Prognosis of Nasopharyngeal Carcinoma. PLoS ONE 2014, 9, e89220. [Google Scholar] [CrossRef] [PubMed]
- Robichaud, N.; Hsu, B.E.; Istomine, R.; Alvarez, F.; Blagih, J.; Ma, E.H.; Morales, S.V.; Dai, D.L.; Li, G.; Souleimanova, M.; et al. Translational Control in the Tumor Microenvironment Promotes Lung Metastasis: Phosphorylation of Eif4e in Neutrophils. Proc. Natl. Acad. Sci. USA 2018, 115, E2202–E2209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rowlett, R.M.; Chrestensen, C.A.; Nyce, M.; Harp, M.G.; Pelo, J.W.; Cominelli, F.; Ernst, P.B.; Pizarro, T.T.; Sturgill, T.W.; Worthington, M.T. Mnk Kinases Regulate Multiple Tlr Pathways and Innate Proinflammatory Cytokines in Macrophages. Am. J. Physiol. Gastrointest Liver Physiol. 2008, 294, G452–G459. [Google Scholar] [CrossRef] [Green Version]
- Moore, C.E.; Pickford, J.; Cagampang, F.R.; Stead, R.L.; Tian, S.; Zhao, X.; Tang, X.; Byrne, C.D.; Proud, C.G. Mnk1 and Mnk2 Mediate Adverse Effects of High-Fat Feeding in Distinct Ways. Sci. Rep. 2016, 6, 23476. [Google Scholar] [CrossRef] [Green Version]
- Sandeman, L.Y.; Kang, W.X.; Wang, X.; Jensen, K.B.; Wong, D.; Bo, T.; Gao, L.; Zhao, J.; Byrne, C.D.; Page, A.J.; et al. Disabling Mnk Protein Kinases Promotes Oxidative Metabolism and Protects against Diet-Induced Obesity. Mol. Metab. 2020, 42, 101054. [Google Scholar] [CrossRef]
- Conn, C.S.; Yang, H.; Tom, H.J.; Ikeda, K.; Oses-Prieto, J.A.; Vu, H.; Oguri, Y.; Nair, S.; Gill, R.M.; Kajimura, S.; et al. The Major Cap-Binding Protein Eif4e Regulates Lipid Homeostasis and Diet-Induced Obesity. Nat. Metab. 2021, 3, 244–257. [Google Scholar] [CrossRef] [PubMed]
- Joshi, S.; Kaur, S.; Redig, A.J.; Goldsborough, K.; David, K.; Ueda, T.; Watanabe-Fukunaga, R.; Baker, D.P.; Fish, E.N.; Fukunaga, R.; et al. Type I Interferon (Ifn)-Dependent Activation of Mnk1 and Its Role in the Generation of Growth Inhibitory Responses. Proc. Natl. Acad. Sci. USA 2009, 106, 12097–12102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joshi, S.; Sharma, B.; Kaur, S.; Majchrzak, B.; Ueda, T.; Fukunaga, R.; Verma, A.K.; Fish, E.N.; Platanias, L.C. Essential Role for Mnk Kinases in Type Ii Interferon (Ifngamma) Signaling and Its Suppressive Effects on Normal Hematopoiesis. J. Biol. Chem. 2011, 286, 6017–6026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fortin, C.F.; Mayer, T.Z.; Cloutier, A.; McDonald, P.P. Translational Control of Human Neutrophil Responses by Mnk1. J. Leukoc. Biol. 2013, 94, 693–703. [Google Scholar] [CrossRef]
- Geter, P.A.; Ernlund, A.W.; Bakogianni, S.; Alard, A.; Arju, R.; Giashuddin, S.; Gadi, A.; Bromberg, J.; Schneider, R.J. Hyperactive Mtor and Mnk1 Phosphorylation of Eif4e Confer Tamoxifen Resistance and Estrogen Independence through Selective Mrna Translation Reprogramming. Genes Dev. 2017, 31, 2235–2249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knight, J.R.; Alexandrou, C.; Skalka, G.L.; Vlahov, N.; Pennel, K.; Officer, L.; Teodosio, A.; Kanellos, G.; Gay, D.M.; May-Wilson, S.; et al. Mnk Inhibition Sensitizes Kras-Mutant Colorectal Cancer to Mtorc1 Inhibition by Reducing Eif4e Phosphorylation and C-Myc Expression. Cancer Discov. 2021, 11, 1228–1247. [Google Scholar] [CrossRef] [PubMed]
- Ueda, T.; Sasaki, M.; Elia, A.J.; Chio, I.; Hamada, K.; Fukunaga, R.; Mak, T.W. Combined Deficiency for Map Kinase-Interacting Kinase 1 and 2 (Mnk1 and Mnk2) Delays Tumor Development. Proc. Natl. Acad. Sci. USA 2010, 107, 13984–13990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furic, L.; Rong, L.; Larsson, O.; Koumakpayi, I.H.; Yoshida, K.; Brueschke, A.; Petroulakis, E.; Robichaud, N.; Pollak, M.; Gaboury, L.A.; et al. Eif4e Phosphorylation Promotes Tumorigenesis and Is Associated with Prostate Cancer Progression. Proc. Natl. Acad. Sci. USA 2010, 107, 14134–14139. [Google Scholar] [CrossRef] [Green Version]
- Knauf, U.; Tschopp, C.; Gram, H. Negative Regulation of Protein Translation by Mitogen-Activated Protein Kinase-Interacting Kinases 1 and 2. Mol. Cell. Biol. 2001, 21, 5500–5511. [Google Scholar] [CrossRef] [Green Version]
- Tamaoki, T.; Nomoto, H.; Takahashi, I.; Kato, Y.; Morimoto, M.; Tomita, F. Staurosporine, a Potent Inhibitor of Phospholipid/Ca++Dependent Protein Kinase. Biochem. Biophys. Res. Commun. 1986, 135, 397–402. [Google Scholar] [CrossRef]
- Jin, X.; Merrett, J.; Tong, S.; Flower, B.; Xie, J.; Yu, R.; Tian, S.; Gao, L.; Zhao, J.; Wang, X.; et al. Design, Synthesis and Activity of Mnk1 and Mnk2 Selective Inhibitors Containing Thieno[2,3-D]Pyrimidine Scaffold. Eur. J. Med. Chem. 2019, 162, 735–751. [Google Scholar] [CrossRef]
- Zhan, Y.; Guo, J.; Yang, W.; Goncalves, C.; Rzymski, T.; Dreas, A.; Żyłkiewicz, E.; Mikulski, M.; Brzózka, K.; Golas, A.; et al. Mnk1/2 Inhibition Limits Oncogenicity and Metastasis of Kit-Mutant Melanoma. J. Clin. Investig. 2017, 127, 4179–4192. [Google Scholar] [CrossRef]
- Yang, H.; Chennamaneni, L.R.; Ho, M.W.T.; Ang, S.H.; Tan, E.S.W.; Jeyaraj, D.A.; Yeap, Y.S.; Liu, B.; Ong, E.H.; Joy, J.K.; et al. Optimization of Selective Mitogen-Activated Protein Kinase Interacting Kinases 1 and 2 Inhibitors for the Treatment of Blast Crisis Leukemia. J. Med. Chem. 2018, 61, 4348–4369. [Google Scholar] [CrossRef]
- Matsui, Y.; Yasumatsu, I.; Yoshida, K.I.; Iimura, S.; Ikeno, Y.; Nawano, T.; Fukano, H.; Ubukata, O.; Hanzawa, H.; Tanzawa, F.; et al. A Novel Inhibitor Stabilizes the Inactive Conformation of Mapk-Interacting Kinase 1. Acta Crystallogr. F Struct. Biol. Commun. 2018, 74, 156–160. [Google Scholar] [CrossRef] [PubMed]
- Reich, S.H.; Sprengeler, P.A.; Chiang, G.G.; Appleman, J.R.; Chen, J.; Clarine, J.; Eam, B.; Ernst, J.T.; Han, Q.; Goel, V.K.; et al. Structure-Based Design of Pyridone-Aminal Eft508 Targeting Dysregulated Translation by Selective Mitogen-Activated Protein Kinase Interacting Kinases 1 and 2 (Mnk1/2) Inhibition. J. Med. Chem. 2018, 61, 3516–3540. [Google Scholar] [CrossRef] [PubMed]
- Santag, S.; Siegel, F.; Wengner, A.M.; Lange, C.; Bömer, U.; Eis, K.; Pühler, F.; Lienau, P.; Bergemann, L.; Michels, M.; et al. Bay 1143269, a Novel Mnk1 Inhibitor, Targets Oncogenic Protein Expression and Shows Potent Anti-Tumor Activity. Cancer Lett. 2017, 390, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Nuzzo, G.; Ciavatta, M.L.; Kiss, R.; Mathieu, V.; Leclercqz, H.; Manzo, E.; Villani, G.; Mollo, E.; Lefranc, F.; D’Souza, L.; et al. Chemistry of the Nudibranch Aldisa Andersoni: Structure and Biological Activity of Phorbazole Metabolites. Mar. Drugs 2012, 10, 1799–1811. [Google Scholar] [CrossRef]
- Radspieler, A.; Liebscher, J. Total Synthesis of Phorbazole C. Tetrahedron 2001, 57, 4867–4871. [Google Scholar] [CrossRef]
- Proud, C.G. Mnks, Eif4e Phosphorylation and Cancer. Biochim. Biophys. Acta 2015, 1849, 766–773. [Google Scholar] [CrossRef]
- Jauch, R.; Cho, M.K.; Jakel, S.; Netter, C.; Schreiter, K.; Aicher, B.; Zweckstetter, M.; Jackle, H.; Wahl, M.C. Mitogen-Activated Protein Kinases Interacting Kinases Are Autoinhibited by a Reprogrammed Activation Segment. EMBO J. 2006, 25, 4020–4032. [Google Scholar] [CrossRef]
- Pinto-Diez, C.; Ferreras-Martin, R.; Carrion-Marchante, R.; Gonzalez, V.M.; Martin, M.E. Deeping in the Role of the Map-Kinases Interacting Kinases (Mnks) in Cancer. Int. J. Mol. Sci. 2020, 21, 2967. [Google Scholar] [CrossRef] [Green Version]
- Diab, S.; Kumarasiri, M.; Yu, M.; Teo, T.; Proud, C.; Milne, R.; Wang, S. Map Kinase-Interacting Kinases—Emerging Targets against Cancer. Chem. Biol. 2014, 21, 441–452. [Google Scholar] [CrossRef]
- Guo, Z.; Peng, G.; Li, E.; Xi, S.; Zhang, Y.; Li, Y.; Lin, X.; Li, G.; Wu, Q.; He, J. Map Kinase-Interacting Serine/Threonine Kinase 2 Promotes Proliferation, Metastasis, and Predicts Poor Prognosis in Non-Small Cell Lung Cancer. Sci. Rep. 2017, 7, 10612. [Google Scholar] [CrossRef] [Green Version]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jin, X.; Li, M.; Qiu, T.; Yu, R.; Jiang, T. Design, Synthesis and Evaluation of Novel Phorbazole C Derivatives as MNK Inhibitors through Virtual High-Throughput Screening. Mar. Drugs 2022, 20, 429. https://doi.org/10.3390/md20070429
Jin X, Li M, Qiu T, Yu R, Jiang T. Design, Synthesis and Evaluation of Novel Phorbazole C Derivatives as MNK Inhibitors through Virtual High-Throughput Screening. Marine Drugs. 2022; 20(7):429. https://doi.org/10.3390/md20070429
Chicago/Turabian StyleJin, Xin, Maowei Li, Tingting Qiu, Rilei Yu, and Tao Jiang. 2022. "Design, Synthesis and Evaluation of Novel Phorbazole C Derivatives as MNK Inhibitors through Virtual High-Throughput Screening" Marine Drugs 20, no. 7: 429. https://doi.org/10.3390/md20070429