
An Update of Lectins from Marine Organisms: Characterization, Extraction Methodology, and Potential Biofunctional Applications

 ,
,  ,
,  , , , , , , ,
, , , , , , ,  ,
,
Abstract
:1. Introduction
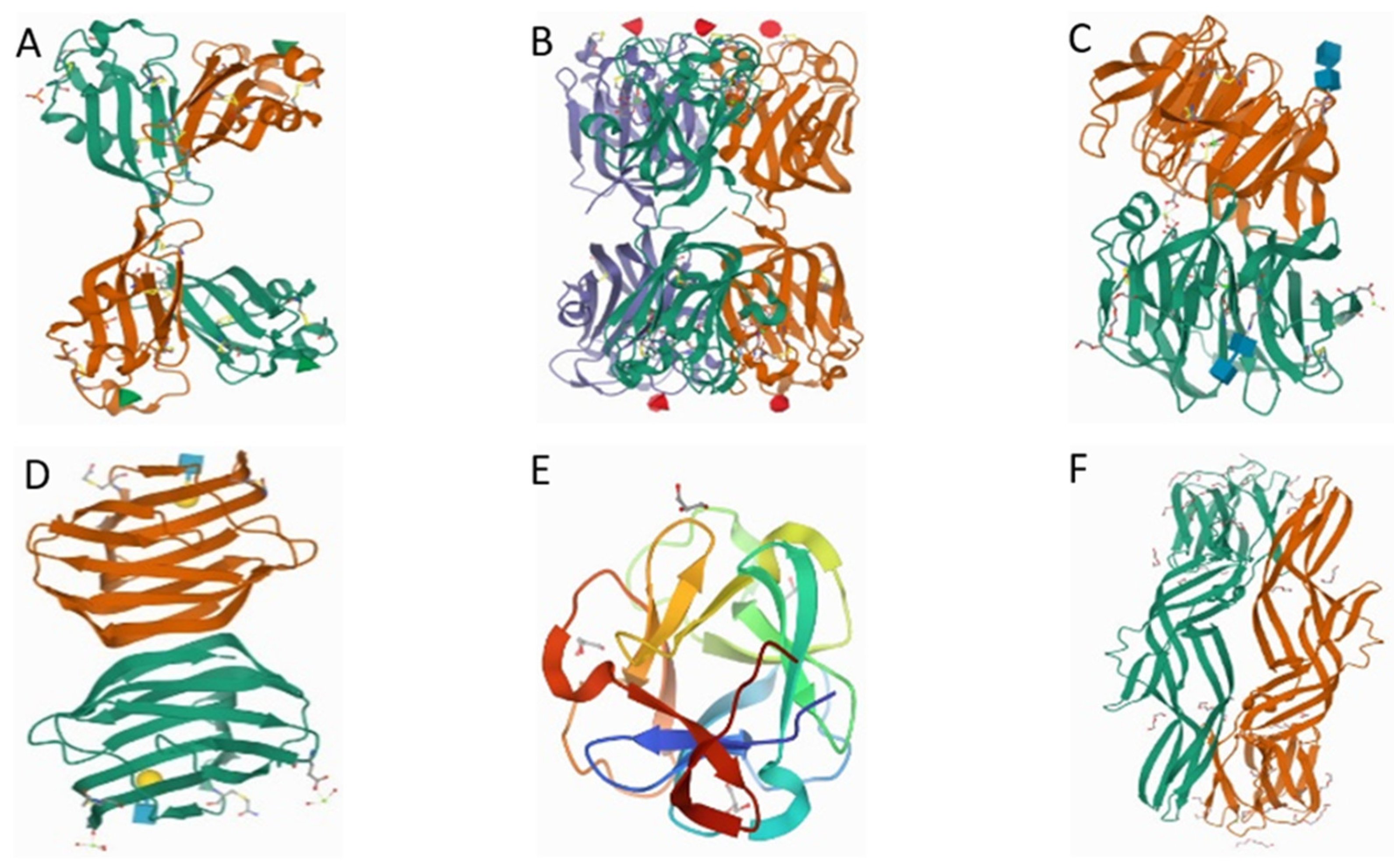
2. Marine Organism-Derived Lectins

2.1. Rhamnose Binding Lectins (RBLs)
2.2. Fucose-Binding Lectins (FTLs)
2.3. C-Type Lectins
2.4. Galectins
2.5. Galactose Binding Lectins
2.6. Mannose-Binding Lectins
2.7. Lactose-Binding Lectins
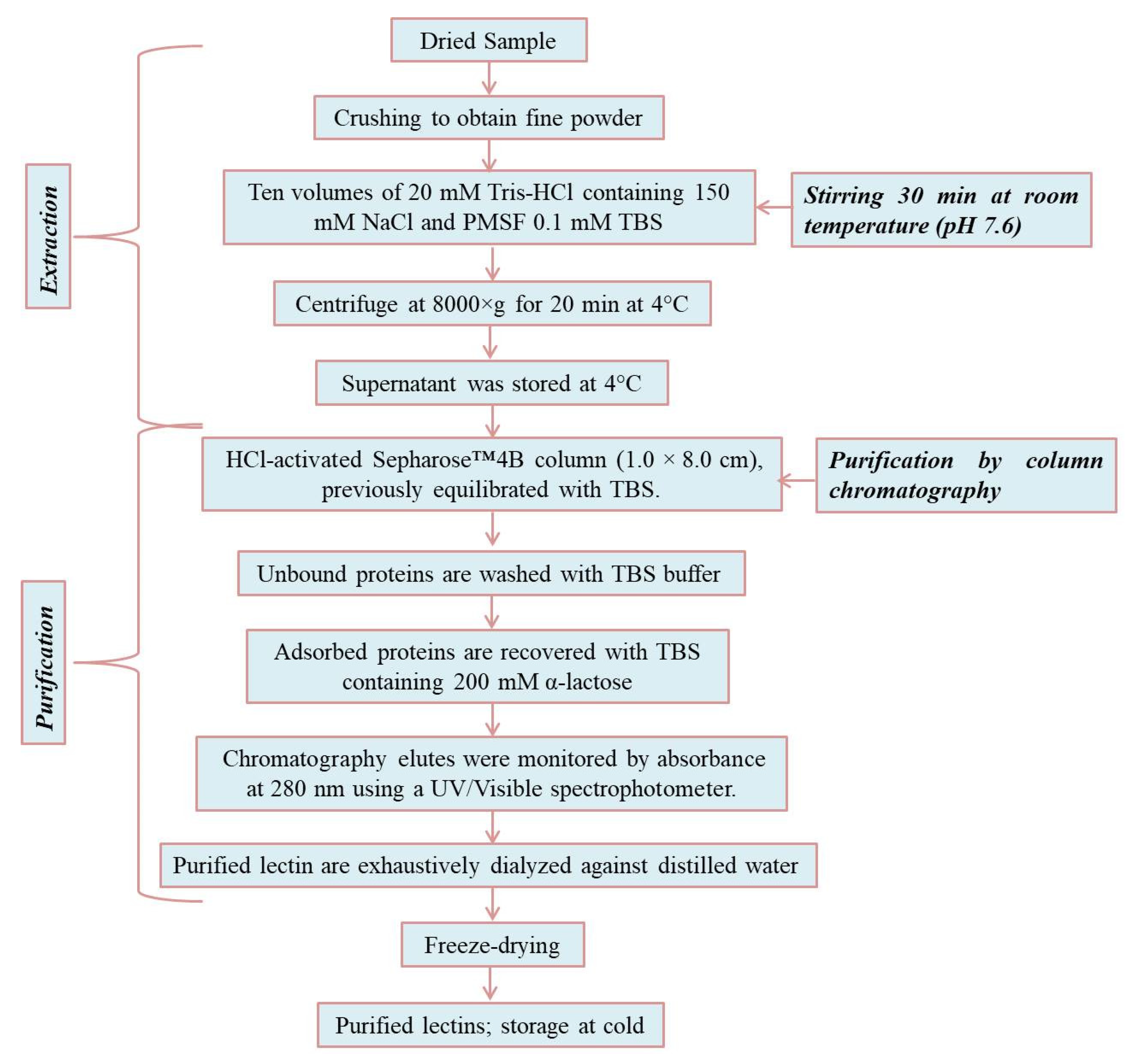
3. Extraction and Purification of Lectins
3.1. Extraction of Lectins from Fish Muscle
3.2. Extraction of Lectins from Fish Skin Mucus
3.3. Lectin Extraction and Purification from Crustaceans by Affinity Chromatography
3.4. Extraction of Lectins from Bivalves via Divinyl Sulphone Activation Method
3.5. Quantification of Lectins
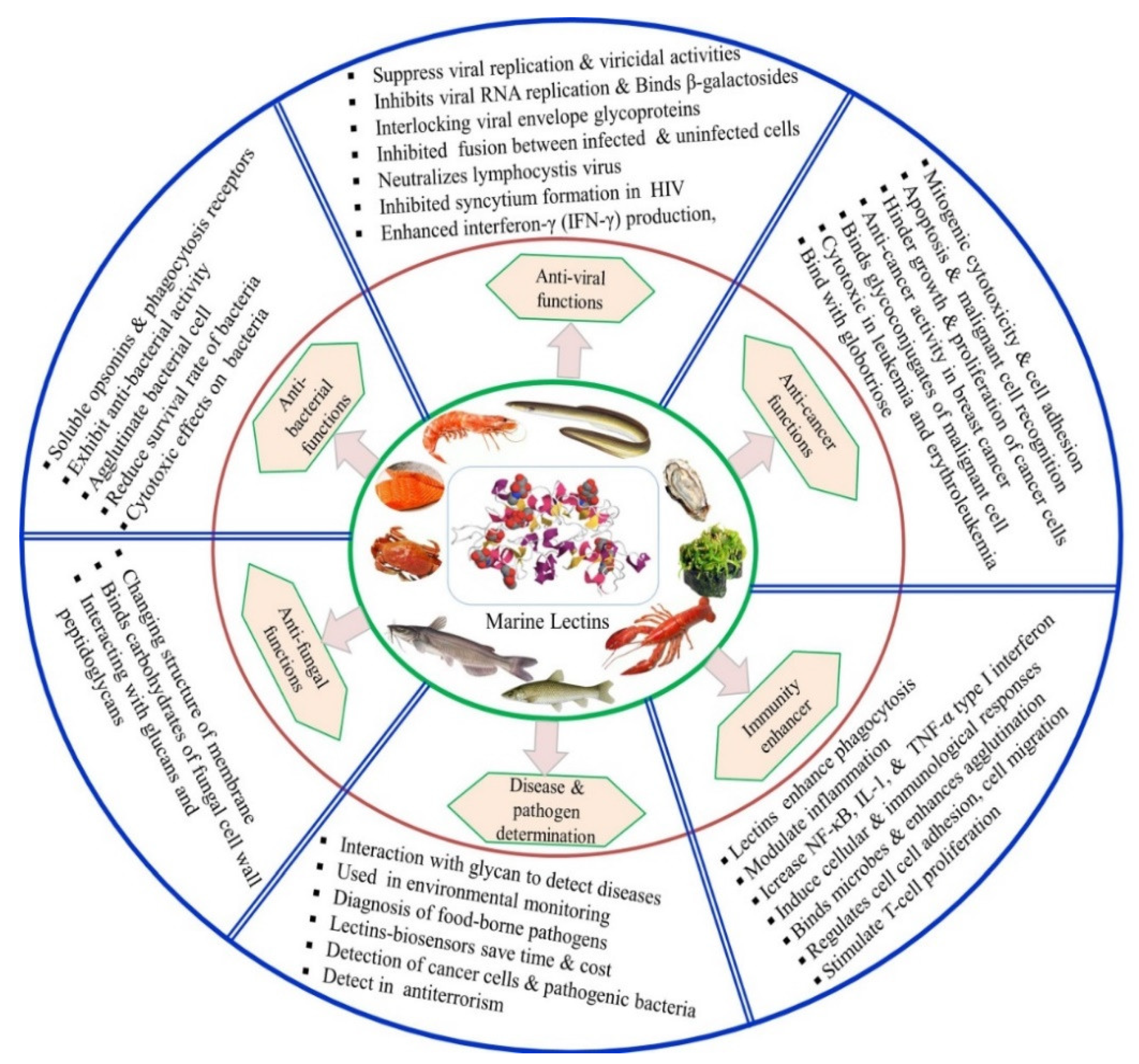
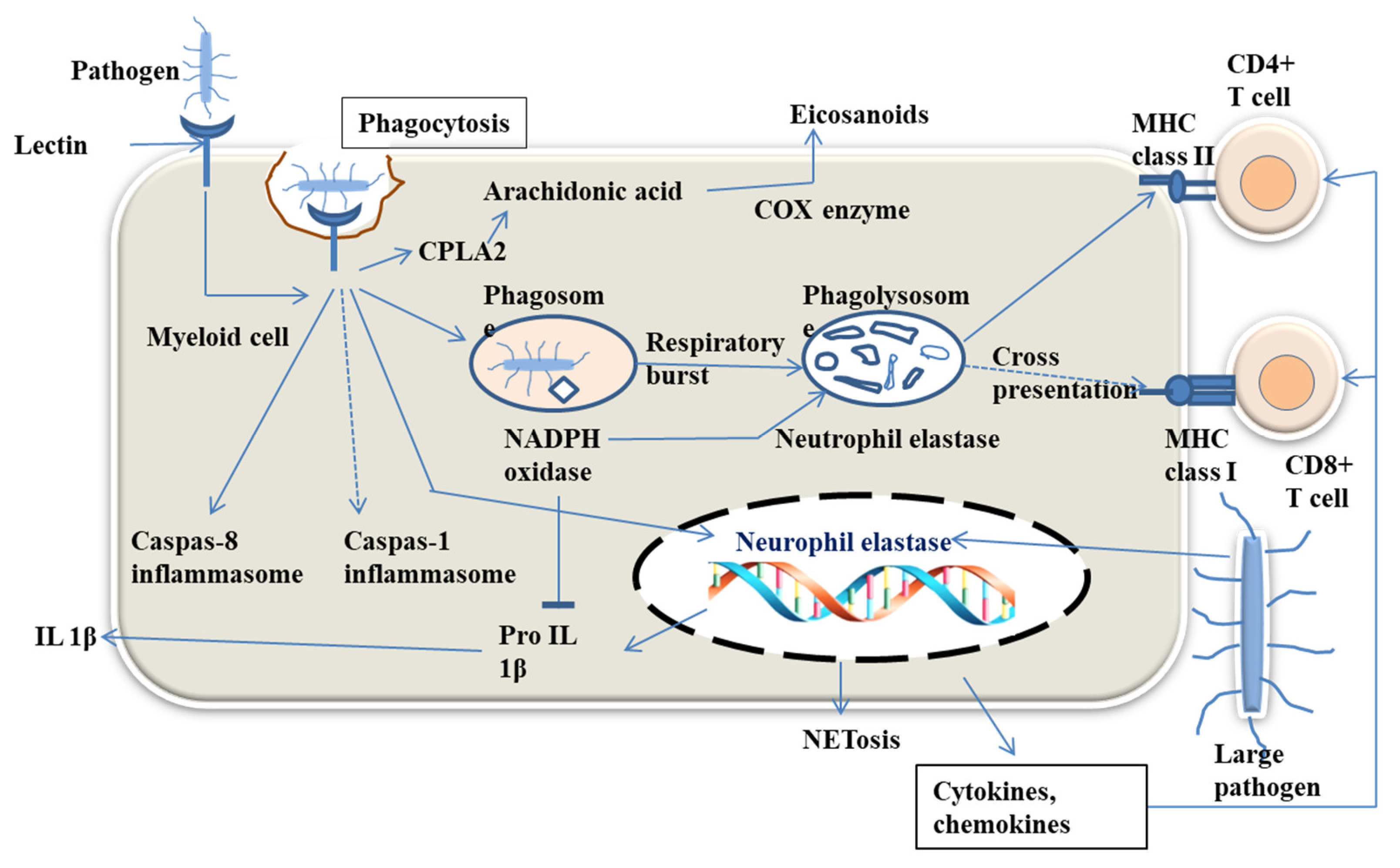
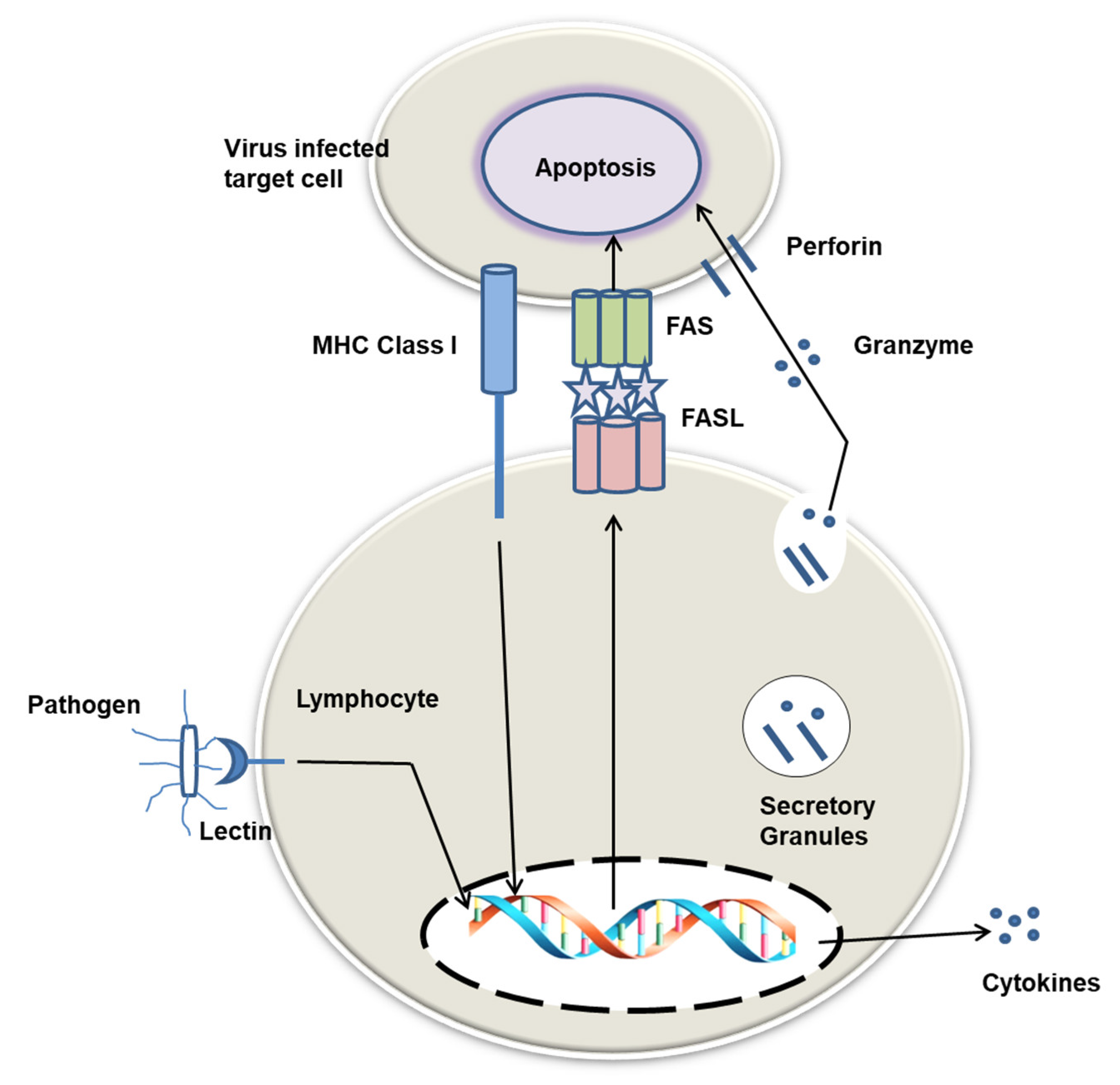
4. Physiological Functions of Lectins in Host Body
5. Other Biological Activities of Lectins
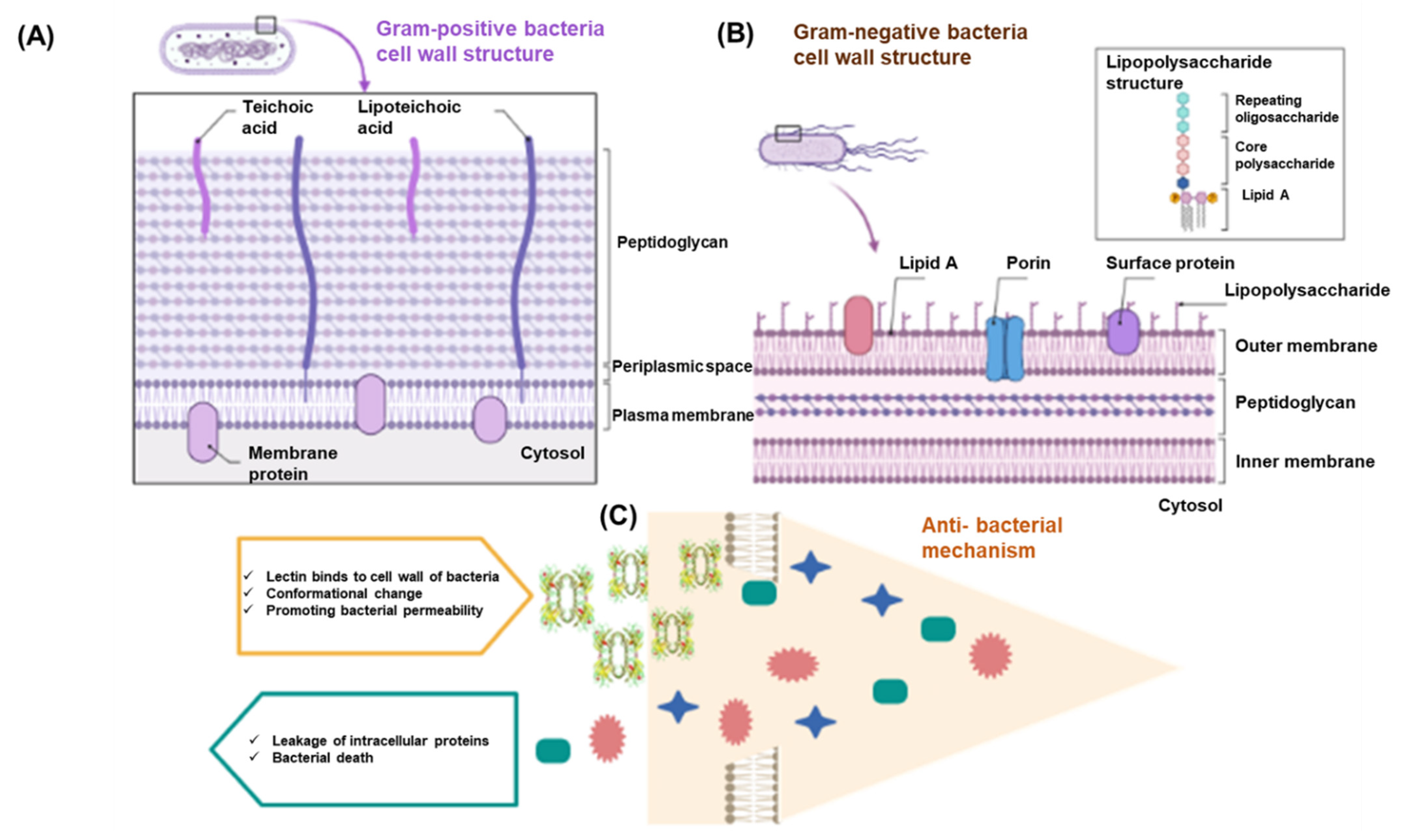
5.1. Antibacterial Activity

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biological Activity | Model System | Source of Lectin | Test Types | Applied Strain | Optimum Dose | Findings | References |
|---|---|---|---|---|---|---|---|
| Antibacterial | Microorganisms and bay scallops | Bay scallop (Argopecten irradians) C type lectin (Ai Lec) | RT-PCR |
| 50 μg/mL | Ai Lec was involved in the immune response to Gram-positive and Gram-negative microbial infection, especially Vibrio anguillarum and Micrococcus luteus in bay scallop. | [36] |
| Microorganism and demosponge | Demosponge (Suberites domuncula) Suberites lectin | PCR |
| 10 μg/mL | The lectin showed antibacterial activity against Gram-positive (Staphylococcus aureus) and Gram-negative bacteria (Escherichia coli). | [131] | |
| Microorganism and manila clam | Manila clam (Ruditapes philippinarum) Manila clam lectin (MCL-4) | Inverted microscope |
| 25 μg/mL | MCL-4 had bacteriostatic properties and may contribute to the host defense mechanisms against invading microorganisms in Manila clam | [151] | |
| Microorganism and rabbit erythrocyte | Cobia (Rachycentron canadum) Tetrameric lection | Ion chromatography |
| 250 μg/mL | The lectin showed antibacterial activity toward E. coli. | [10] | |
| Microorganism and human erythrocytes | Marine sponge (Cliona varians) CvL lectin | Affinity chromatography |
| 25 μg/mL | CvL lectin showed intense antibacterial activity against Bacillus subtilis. | [150] | |
| Antiviral | Cell line, virus, and fish | Flounder (Paralichthys olivaceus) Galectin-1 | qRt-PCR |
| 50 μg/mL | Galectin-1 from flounder was able to neutralize the lymphocystis disease virus (LCDV) and exhibited anti-inflammatory activity against LCDV. | [152] |
| Shrimp | Shrimp (Penaeus monodon) C-type lectin | PCR |
| - | A lectin domain containing PmAV protein isolated from shrimp was effective against white spot syndrome virus (WSSV) | [153] | |
| Virus and cell lines | Marine worm (Chaetopterus variopedatus) β-galactose-specific lectin (CVL) | qRt-PCR |
| 25–100 μg/mL | CVL blocked the cell–cell fusion process of the human immunodeficiency virus infected and uninfected cells with an EC50 of 0.07 μM and has the potential to be an anti-HIV-1 agent. | [154] | |
| Virus and cell lines | Sea worm (Serpula vermicularis) GlcNAc-specific lection (SVL) | ELISA |
| 30 μg/mL | SVL showed potential activity against human immunodeficiency virus (HIV-1) by producing viral p24 antigen, with EC50 values of 0.23 and 0.15 μg/mL. | [155] | |
| Antifungal | Microbial cells | Chinese amphioxus (Branchiostoma belcheri) C-type lectin (AmphiCTL1) | Q-PCR |
| 200 μg/mL | AmphiCTL-1 lectin showed potential activity against Saccharomyces cerevisiae. | [153] |
| Microbial cells | Orange-spotted grouper (Epinephelus coioides) C-type lectin (Ec-CTL) | Q-PCR |
| 10 μg | This lectin showed potent activity against S. cerevisiae. | [156] | |
| Microbial cells | Lamprey (Lampetra japonica) Serum lectin (NPGBP) | RT-PCR |
| 10 mg/mL | The lectin showed agglutinating activities against Candida albicans. | [157] | |
| Anticancer or antitumour | Virus and cell lines | Marine worm (Chaetopterus variopedatus) β-galactose-specific lectin (CVL) | qRt-PCR | - | 25–100 μg/mL | CVL blocked the cell–cell fusion process of the human immunodeficiency virus infected and uninfected cells with an EC50 of 0.07 μM and has the potential to be an anti-HIV-1 agent. | [154] |
| Virus and cell lines | Sea worm (Serpula vermicularis) GlcNAc-specific lection (SVL) | ELISA | - | 30 μg/mL | SVL showed potential activity against human immunodeficiency virus (HIV-1) by producing viral p24 antigen, with EC50 values of 0.23 and 0.15 μg/mL. | [155] | |
| Tumor cell line | Chinook salmon (Oncorhynchus tshawytscha) Roe lectin | Microplate reader | - | - | The lectin showed intense antiproliferative activity towards human breast cancer MCF-7 cells and hepatoma Hep G2 cells. | [158] | |
| Cancer cell lines | Marine sponge (Cliona varians) CvL lectin | Flow cytometry | - | 70–100 μg/mL | CvL lectin showed potential activity on K562 and Jurkat cancer cell lines. | [159] | |
| Cancer cells | Wheat germ (Triticum vulgaris) WGA lectin | Electrode array | - | 100 μg/mL | A label-free electrochemical impedance spectroscopy (EIS) biosensor could be promising for the label-free profiling of the glycan expression of cancer-related glycoproteins in the early stage of a cancer diagnosis. | [160] |
5.2. Anti-Viral Activity
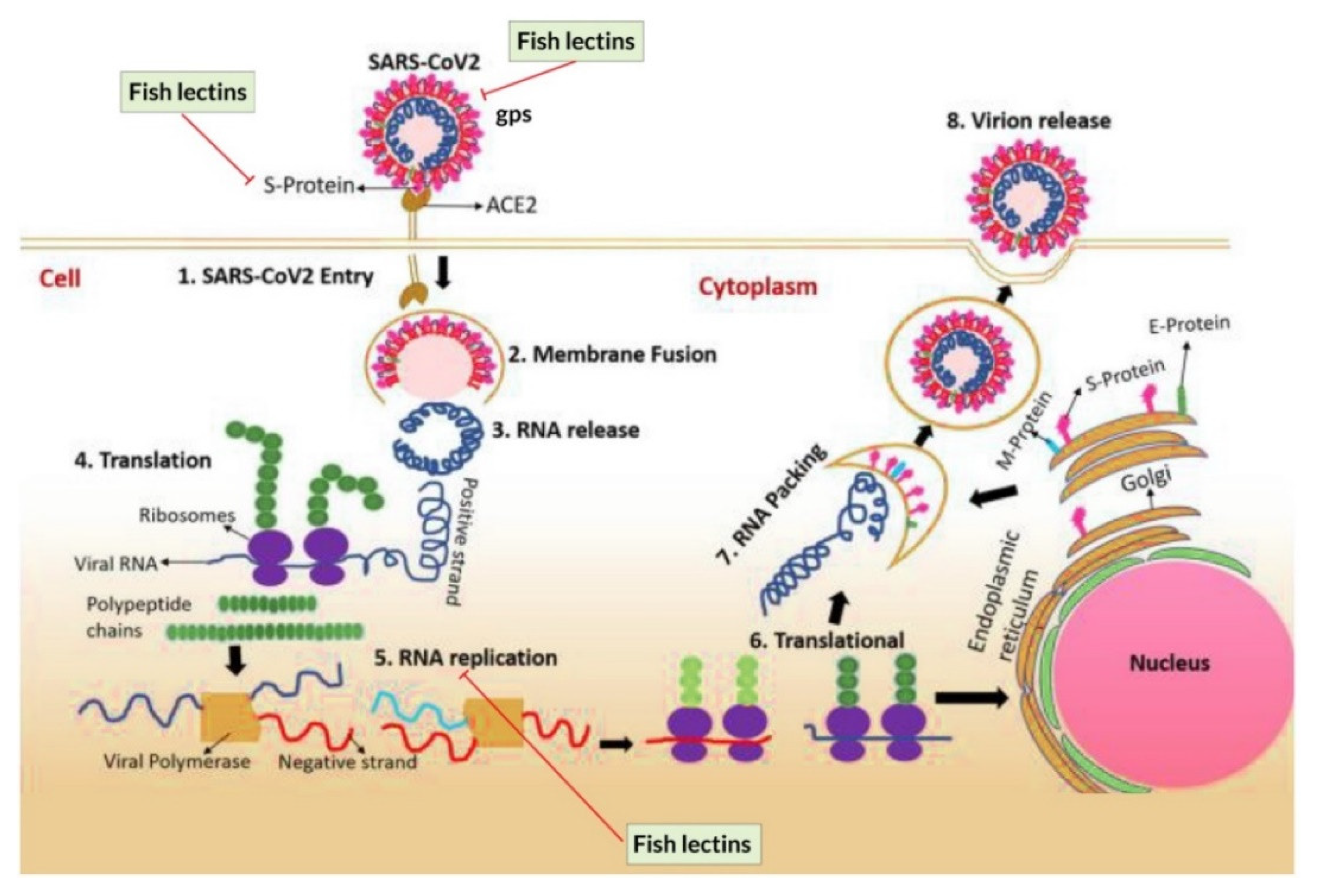
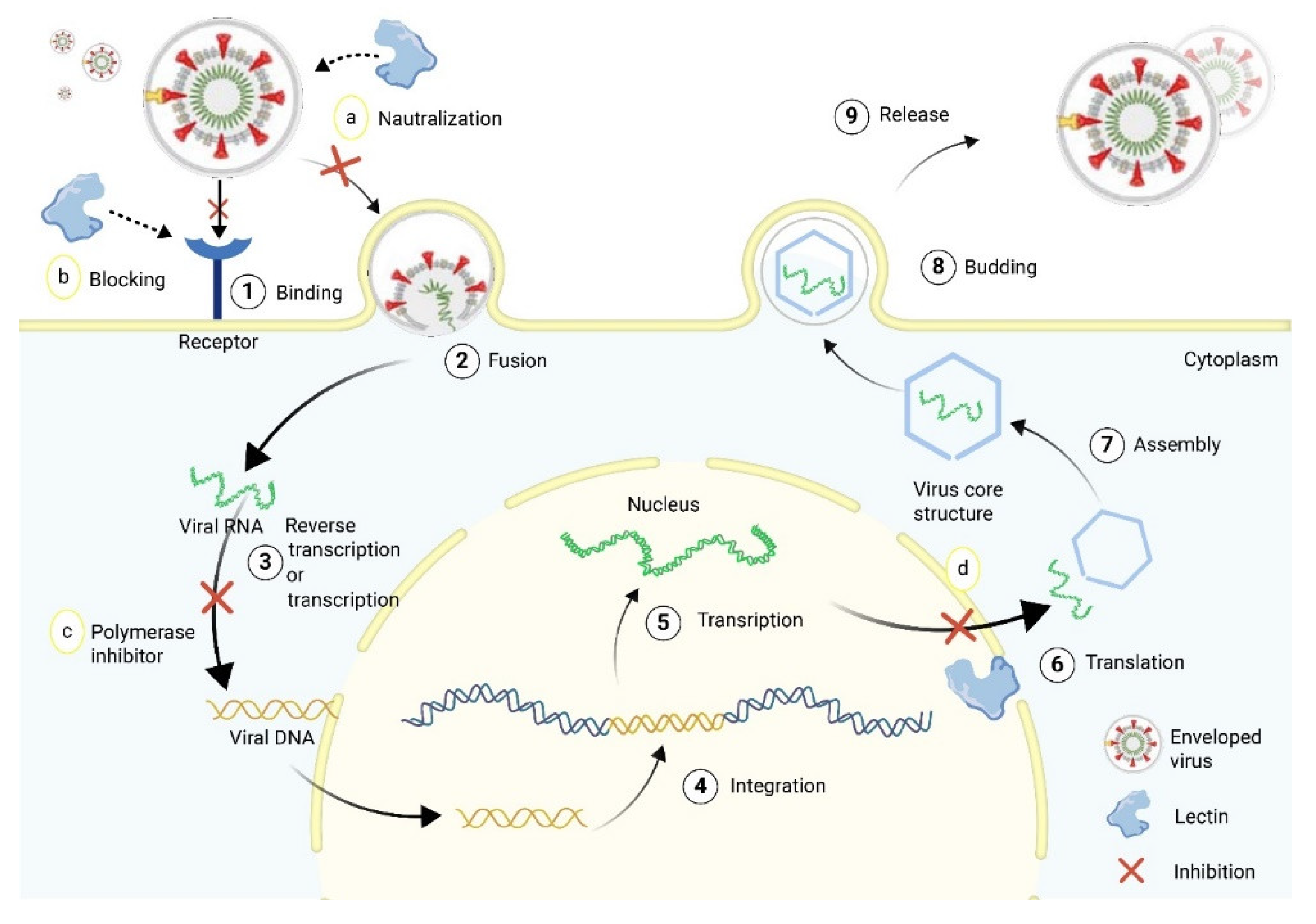
5.2.1. Coronavirus
5.2.2. Lectins as Potential Inhibitors of SARS-CoV-2
5.3. Anti-Fungal Activity
5.4. Activity Related to Homeostatic Maintenance of Intestinal Microbiota
5.5. Anti-Cancer or Anti-Tumor Activities
5.6. Lectins as an Immunity Enhancer
5.7. Other Potential Applications
6. Consumer Expectations in the Safety of Lectin-Based Food Products
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dias, R.D.O.; Machado, L.D.S.; Migliolo, L.; Franco, O.L. Insights into Animal and Plant Lectins with Antimicrobial Activities. Molecules 2015, 20, 519–541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manikandan, S.B.; Manikandan, R.; Arumugam, M.; Mullainadhan, P. An overview on human serum lectins. Heliyon 2020, 6, e04623. [Google Scholar] [CrossRef] [PubMed]
- Kotecha, H.; Poduval, P. Microbial lectins: Roles and applications. In Advances in Biological Science Research: A Practical Approach; Surya, M., Milind, N., Eds.; Elsevier: Amsterdam, The Netherlands, 2019. [Google Scholar]
- Muramoto, K. Lectins as Bioactive Proteins in Foods and Feeds. Food Sci. Technol. Res. 2017, 23, 487–494. [Google Scholar] [CrossRef] [Green Version]
- Elumalai, P.; Rubeena, A.S.; Arockiaraj, J.; Wongpanya, R.; Cammarata, M.; Ringø, E.; Vaseeharan, B. The role of lectins in finfish: A review. Rev. Fish. Sci. Aquac. 2019, 27, 152–169. [Google Scholar] [CrossRef]
- Cammarata, M.; Parisi, M.G.; Vasta, G.R. Chapter 18—Evolution and Immune Function of Fish Lectins. In Lessons in Immunity; Ballarin, L., Cammarata, M., Eds.; Academic Press: Cambridge, MA, USA, 2016; pp. 239–256. [Google Scholar]
- De Juan, L.L.; Recio, V.G.; López, P.J.; Juan, T.G.; Cordoba-Diaz, M.; Cordoba-Diaz, D. Pharmaceutical applications of lectins. J. Drug Deliv. Sci. Technol. 2017, 42, 126–133. [Google Scholar] [CrossRef]
- Liu, L.; Dang, Y. Antimicrobial activity of mannose binding lectin in grass carp (Ctenopharyngodon idella) in vivo and in vitro. Fish Shellfish Immunol. 2020, 98, 25–33. [Google Scholar] [CrossRef]
- Arasu, A.; Kumaresan, V.; Ganesh, M.-R.; Pasupuleti, M.; Arasu, M.V.; Al-Dhabi, N.A.; Arockiaraj, J. Bactericidal activity of fish galectin 4 derived membrane-binding peptide tagged with oligotryptophan. Dev. Comp. Immunol. 2017, 71, 37–48. [Google Scholar] [CrossRef]
- Ngai, P.H.; Ng, T. A mannose-specific tetrameric lectin with mitogenic and antibacterial activities from the ovary of a teleost, the cobia (Rachycentron canadum). Appl. Microbiol. Biotechnol. 2007, 74, 433–438. [Google Scholar] [CrossRef]
- Ewart, K.V.; Johnson, S.C.; Ross, N.W. Lectins of the innate immune system and their relevance to fish health. ICES J. Mar. Sci. 2001, 58, 380–385. [Google Scholar] [CrossRef]
- Bun Ng, T.; Chi Fai Cheung, R.; Cheuk Wing Ng, C.; Fei Fang, E.; Ho Wong, J. A review of fish lectins. Curr. Protein Pept. Sci. 2015, 16, 337–351. [Google Scholar]
- Da Silva Lino, M.A.; Bezerra, R.F.; da Silva, C.D.C.; Carvalho, E.V.M.M.; Coelho, L.C.B.B. Fish lectins: Abrief review. Adv. Zool. Res. 2014, 5, 95–114. [Google Scholar]
- Rubeena, A.S.; Divya, M.; Vaseeharan, B.; Karthikeyan, S.; Ringo, E.; Preetham, E. Antimicrobial and biochemical characterization of a C-type lectin isolated from pearl spot (Etroplus suratensis). Fish Shellfish Immunol. 2019, 87, 202–211. [Google Scholar] [CrossRef]
- Wang, K.; Liu, C.; Hou, Y.; Zhou, H.; Wang, X.; Mai, K.; He, G. Differential apoptotic and mitogenic effects of lectins in Zebrafish. Front. Endocrinol. 2019, 10, 356. [Google Scholar] [CrossRef] [Green Version]
- Da Silva, C.D.C.; Coriolano, M.C.; Da Silva Lino, M.A.; De Melo, C.M.L.; De Souza Bezerra, R.; De Carvalho, E.V.M.M.; Dos Santos, A.J.G.; Pereira, V.R.A.; Coelho, L.C.B.B. Purification and characterization of a mannose recognition lectin from Oreochromis niloticus (Tilapia Fish): Cytokine production in mice splenocytes. Appl. Biochem. Biotechnol. 2012, 166, 424–435. [Google Scholar] [CrossRef]
- Mu, L.; Yin, X.; Wu, H.; Han, K.; Guo, Z.; Ye, J. MAp34 Regulates the Non-specific Cell Immunity of Monocytes/Macrophages and Inhibits the Lectin Pathway of Complement Activation in a Teleost Fish. Front. Immunol. 2020, 11, 1706. [Google Scholar] [CrossRef]
- Russell, S.; Lumsden, J.S. Function and heterogeneity of fish lectins. Vet. Immunol. Immunopathol. 2005, 108, 111–120. [Google Scholar] [CrossRef]
- Marques, M.R.F.; Barracco, M.A. Lectins, as non-self-recognition factors, in crustaceans. Aquaculture 2000, 191, 23–44. [Google Scholar] [CrossRef]
- Yu, X.-Q.; Gan, H.R.; Kanost, M. Immulectin, an inducible C-type lectin from an insect, Manduca sexta, stimulates activation of plasma prophenol oxidase. Insect Biochem. Mol. Biol. 1999, 29, 585–597. [Google Scholar] [CrossRef]
- Shirai, T.; Watanabe, Y.; Lee, M.-S.; Ogawa, T.; Muramoto, K. Structure of rhamnose-binding lectin CSL3: Unique pseudo-tetrameric architecture of a pattern recognition protein. J. Mol. Biol. 2009, 391, 390–403. [Google Scholar] [CrossRef]
- Bianchet, M.A.; Odom, E.W.; Vasta, G.R.; Amzel, L.M. Structure and Specificity of a Binary Tandem Domain F-Lectin from Striped Bass (Morone saxatilis). J. Mol. Biol. 2010, 401, 239–252. [Google Scholar] [CrossRef] [Green Version]
- Capaldi, S.; Faggion, B.; Carrizo, M.E.; Destefanis, L.; Gonzalez, M.C.; Perduca, M.; Bovi, M.; Galliano, M.; Monaco, H.L. Three-dimensional structure and ligand-binding site of carp fishelectin (FEL). Acta Crystallogr. Sect. D Biol. Crystallogr. 2015, 71, 1123–1135. [Google Scholar] [CrossRef]
- Ghosh, A.; Banerjee, A.; Amzel, L.M.; Vasta, G.R.; Bianchet, M.A. Structure of the zebrafish galectin-1-L2 and model of its interaction with the infectious hematopoietic necrosis virus (IHNV) envelope glycoprotein. Glycobiology 2019, 29, 419–430. [Google Scholar] [CrossRef]
- García-Maldonado, E.; Cano-Sánchez, P.; Hernández-Santoyo, A. Molecular and functional characterization of a glycosylated Galactose-Binding lectin from Mytilus californianus. Fish Shellfish Immunol. 2017, 66, 564–574. [Google Scholar] [CrossRef]
- Jia, N.; Liu, N.; Cheng, W.; Jiang, Y.L.; Sun, H.; Chen, L.L.; Peng, J.; Zhang, Y.; Ding, Y.H.; Zhang, Z.H.; et al. Structural basis for receptor recognition and pore formation of a zebrafish aerolysin-like protein. EMBO Rep. 2016, 17, 235–248. [Google Scholar] [CrossRef] [Green Version]
- Gao, C.; Su, B.; Zhang, D.; Yang, N.; Song, L.; Fu, Q.; Zhou, S.; Tan, F.; Li, C. l-rhamnose-binding lectins (RBLs) in turbot (Scophthalmus maximus L.): Characterization and expression profiling in mucosal tissues. Fish Shellfish Immunol. 2018, 80, 264–273. [Google Scholar] [CrossRef]
- Fujii, Y.; Gerdol, M.; Kawsar, S.M.; Hasan, I.; Spazzali, F.; Yoshida, T.; Ogawa, Y.; Rajia, S.; Kamata, K.; Koide, Y.; et al. A GM1b/asialo-GM1 oligosaccharide-binding R-type lectin from purplish bifurcate mussels Mytilisepta virgata and its effect on MAP kinases. FEBS J. 2020, 287, 2612–2630. [Google Scholar] [CrossRef] [Green Version]
- Ozeki, Y.; Matsui, T.; Suzuki, M.; Titani, K. Amino acid sequence and molecular characterization of a D-galactoside-specific lectin purified from sea urchin (Anthocidaris crassispina) eggs. Biochemistry 1991, 30, 2391–2394. [Google Scholar] [CrossRef]
- Huang, Y.; Wang, M.; Zhao, X.; Shi, Q. Transcriptome sequencing of the gill and barbel of Southern catfish (Silurus meridionalis) revealed immune responses and novel rhamnose-binding lectins (RBLs). Genomics 2019, 111, 222–230. [Google Scholar] [CrossRef]
- Zhang, D.; Beck, B.H.; Mohammed, H.; Zhao, H.; Thongda, W.; Ye, Z.; Zeng, Q.; Shoemaker, C.A.; Fuller, S.A.; Peatman, E. l-rhamnose-binding lectins (RBLs) in Nile tilapia, Oreochromis niloticus: Characterization and expression profiling in mucosal tissues. Fish Shellfish Immunol. 2018, 72, 426–435. [Google Scholar] [CrossRef]
- Sugawara, S. Application of lectin from catfish eggs to cancer therapy: A fundamental study. Yakugaku Zasshi 2018, 138, 1451–1459. [Google Scholar] [CrossRef] [Green Version]
- Sugawara, S.; Im, C.; Kawano, T.; Tatsuta, T.; Koide, Y.; Yamamoto, D.; Ozeki, Y.; Nitta, K.; Hosono, M. Catfish rhamnose-binding lectin induces G0/1 cell cycle arrest in Burkitt’s lymphoma cells via membrane surface Gb3. Glycoconj. J. 2017, 34, 127–138. [Google Scholar] [CrossRef]
- Hosono, M.; Sugawara, S.; Matsuda, A.; Tatsuta, T.; Koide, Y.; Hasan, I.; Ozeki, Y.; Nitta, K. Binding profiles and cytokine-inducing effects of fish rhamnose-binding lectins on Burkitt’s lymphoma Raji cells. Fish Physiol. Biochem. 2014, 40, 1559–1572. [Google Scholar] [CrossRef]
- Nemoto, R.; Yamamoto, S.; Ogawa, T.; Naude, R.; Muramoto, K. Effect of chum salmon egg lectin on tight junctions in Caco-2 cell monolayers. Molecules 2015, 20, 8094–8106. [Google Scholar] [CrossRef] [Green Version]
- Zhu, L.; Song, L.; Xu, W.; Qian, P.-Y. Identification of a C-type lectin from the bay scallop Argopecten irradians. Mol. Biol. Rep. 2009, 36, 1167–1173. [Google Scholar] [CrossRef] [Green Version]
- Grinchenko, A.V.; Von Kriegsheim, A.; Shved, N.A.; Egorova, A.E.; Ilyaskina, D.V.; Karp, T.D.; Goncharov, N.V.; Petrova, I.Y.; Kumeiko, V.V. A novel C1q domain-containing protein isolated from the mollusk Modiolus kurilensis recognizing glycans enriched with acidic galactans and mannans. Mar. Drugs 2021, 19, 668. [Google Scholar] [CrossRef]
- Mizgina, T.O.; Chikalovets, I.V.; Molchanova, V.I.; Ziganshin, R.H.; Chernikov, O.V. Identification and Characterization of a Novel Lectin from the Clam Glycymeris yessoensis and Its Functional Characterization under Microbial Stimulation and Environmental Stress. Mar. Drugs 2021, 19, 474. [Google Scholar] [CrossRef]
- Rubeena, A.S.; Lakshmi, S.; George, D.; Subramaniyan, S.B.; Veerappan, A.; Preetham, E. Shrimp lectin (Md-Lec) conjugated copper sulfide nanoparticles enhance the elimination of aquatic pathogens in infected Nile tilapia (Oreochromis niloticus). RSC Adv. 2020, 10, 44216–44224. [Google Scholar] [CrossRef]
- Sivakamavalli, J.; Park, K.; Kwak, I.S.; Vaseeharan, B. Purification and partial characterization of carbohydrate-recognition protein C-type lectin from Hemifusus pugilinus. Carbohydr. Res. 2021, 499, 108224. [Google Scholar] [CrossRef]
- Xin, Z.; Yu, D.; Yang, B.; Chen, L.; Hayouka, Z.; Chen, X.; Gong, Y.; Dai, H.; Wang, L.; Zhao, Y.; et al. Molecular characterization, expression and immune functions of two C-type lectin from Venerupis philippinarum. Fish Shellfish Immunol. 2020, 107, 260–268. [Google Scholar] [CrossRef]
- Iiyama, C.; Yoneda, F.; Tsutsumi, M.; Tsutsui, S.; Nakamura, O. Mannose-binding C-type lectins as defense molecules on the body surface of the sea urchin Pseudocentrotus depressus. Dev. Comp. Immunol. 2021, 116, 103915. [Google Scholar] [CrossRef]
- Simão, M.; Moço, M.; Marques, L.; Santos, R. Characterization of the glycans involved in sea urchin Paracentrotus lividus reversible adhesion. Mar. Biol. 2020, 167, 125. [Google Scholar] [CrossRef]
- Qiu, W.; Chen, F.; Chen, R.; Li, S.; Zhu, X.; Xiong, M.; Wang, K.J. A New C-Type Lectin Homolog SpCTL6 Exerting Immunoprotective Effect and Regulatory Role in Mud Crab Scylla paramamosain. Front. Immunol. 2021, 12, 661823. [Google Scholar] [CrossRef]
- Imamichi, Y.; Hikosaka, K.; Kawai, N.; Koubaku, N.; Hosoi, M.; Mizuta, S.; Yokoyama, Y. Purification, characterization and cDNA cloning of a lectin from the brittle star Ophioplocus japonicus. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2022, 262, 110757. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, J.; Kong, X.; Zhao, X.; Pei, C.; Li, L. A C-type lectin, Nattectin-like protein (CaNTC) in Qihe crucian carp Carassius auratus: Binding ability with LPS, PGN and various bacteria, and agglutinating activity against bacteria. Fish Shellfish Immunol. 2017, 67, 382–392. [Google Scholar] [CrossRef]
- Huang, Z.; Ma, A.; Xia, D.; Wang, X.; Sun, Z.; Shang, X.; Yang, Z.; Qu, J. Immunological characterization and expression of lily-type lectin in response to environmental stress in turbot (Scophthalmus maximus). Fish Shellfish Immunol. 2016, 58, 323–331. [Google Scholar] [CrossRef]
- Xia, D.; Ma, A.; Huang, Z.; Shang, X.; Cui, W.; Yang, Z.; Qu, J. Molecular characterization and expression analysis of Lily-type lectin (SmLTL) in turbot Scophthalmus maximus, and its response to Vibrio anguillarum. J. Oceanol. Limnol. 2018, 36, 508–518. [Google Scholar] [CrossRef]
- Kugapreethan, R.; Wan, Q.; Nilojan, J.; Lee, J. Identification and characterization of a calcium-dependent lily-type lectin from black rockfish (Sebastes schlegelii): Molecular antennas are involved in host defense via pathogen recognition. Dev. Comp. Immunol. 2018, 81, 54–62. [Google Scholar] [CrossRef]
- Huang, X.; Han, K.; Li, T.; Wang, W.; Ren, Q. Novel L-type lectin from fresh water prawn, Macrobrachium rosenbergii participates in antibacterial and antiviral immune responses. Fish Shellfish Immunol. 2018, 77, 304–311. [Google Scholar] [CrossRef]
- Parisi, M.G.; Benenati, G.; Cammarata, M. Sea bass Dicentrarchus labrax (L.) bacterial infection and confinement stress acts on F-type lectin (DlFBL) serum modulation. J. Fish Dis. 2015, 38, 967–976. [Google Scholar] [CrossRef] [Green Version]
- Parisi, M.G.; Cammarata, M.; Benenati, G.; Salerno, G.; Mangano, V.; Vizzini, A.; Parrinello, N. A serum fucose-binding lectin (DlFBL) from adult Dicentrarchus labrax is expressed in larva and juvenile tissues and contained in eggs. Cell Tissue Res. 2010, 341, 279–288. [Google Scholar] [CrossRef] [Green Version]
- Cho, S.-Y.; Kwon, J.; Vaidya, B.; Kim, J.-O.; Lee, S.; Jeong, E.-H.; Baik, K.S.; Choi, J.-S.; Bae, H.-J.; Oh, M.-J.; et al. Modulation of proteome expression by F-type lectin during viral hemorrhagic septicemia virus infection in fathead minnow cells. Fish Shellfish Immunol. 2014, 39, 464–474. [Google Scholar] [CrossRef]
- Odom, E.W.; Vasta, G.R. Characterization of a binary tandem domain F-type lectin from striped bass (Morone saxatilis). J. Biol. Chem. 2006, 281, 1698–1713. [Google Scholar] [CrossRef] [Green Version]
- Cammarata, M.; Parisi, M.G.; Benenati, G.; Vasta, G.R.; Parrinello, N. A rhamnose-binding lectin from sea bass (Dicentrarchus labrax) plasma agglutinates and opsonizes pathogenic bacteria. Dev. Comp. Immunol. 2014, 44, 332–340. [Google Scholar] [CrossRef]
- Tateno, H.; Saneyoshi, A.; Ogawa, T.; Muramoto, K.; Kamiya, H.; Saneyoshi, M. Isolation and characterization of rhamnose-binding lectins from eggs of steelhead trout (Oncorhynchus mykiss) homologous to low density lipoprotein receptor superfamily. J. Biol. Chem. 1998, 273, 19190–19197. [Google Scholar] [CrossRef] [Green Version]
- Salerno, G.; Parisi, M.G.; Parrinello, D.; Benenati, G.; Vizzini, A.; Vazzana, M.; Vasta, G.R.; Cammarata, M. F-type lectin from the sea bass (Dicentrarchus labrax): Purification, cDNA cloning, tissue expression and localization, and opsonic activity. Fish Shellfish Immunol. 2009, 27, 143–153. [Google Scholar] [CrossRef] [Green Version]
- Lima, P.; Hartley-Tassell, L.; Cooper, O.; Wynne, J. Searching for the sweet spot of amoebic gill disease of farmed Atlantic salmon: The potential role of glycan-lectin interactions in the adhesion of Neoparamoeba perurans. Int. J. Parasitol. 2021, 51, 545–557. [Google Scholar] [CrossRef]
- Kong, H.J.; Kim, W.-J.; Kim, H.S.; Lee, Y.J.; Kim, C.H.; Nam, B.-H.; Kim, Y.-O.; Kim, D.-G.; Lee, S.-J.; Lim, S.-G.; et al. Molecular characterization of a tandem-repeat galectin-9 (RuGlec9) from Korean rose bitterling (Rhodeus uyekii). Fish Shellfish Immunol. 2012, 32, 939–944. [Google Scholar] [CrossRef]
- Tian, M.; Yang, N.; Zhang, L.; Fu, Q.; Tan, F.; Li, C. Expression profiling and functional characterization of galectin-3 of turbot (Scophthalmus maximus L.) in host mucosal immunity. Fish Shellfish Immunol. 2019, 84, 333–340. [Google Scholar] [CrossRef]
- Gerdol, M. First Insights into the Repertoire of Secretory Lectins in Rotifers. Mar. Drugs 2022, 20, 130. [Google Scholar] [CrossRef]
- Swarna, R.R.; Asaduzzaman, A.; Kabir, S.R.; Arfin, N.; Kawsar, S.M.; Rajia, S.; Fujii, Y.; Ogawa, Y.; Hirashima, K.; Kobayashi, N. Antiproliferative and Antimicrobial Potentials of a Lectin from Aplysia kurodai (Sea Hare) Eggs. Mar. Drugs 2021, 19, 394. [Google Scholar] [CrossRef]
- Bornhöfft, K.F.; Martorell Ribera, J.; Viergutz, T.; Venuto, M.T.; Gimsa, U.; Galuska, S.P.; Rebl, A. Characterization of sialic acid-binding immunoglobulin-type lectins in fish reveals teleost-specific structures and expression patterns. Cells 2020, 9, 836. [Google Scholar] [CrossRef] [Green Version]
- Carneiro, R.F.; Viana, J.T.; Torres, R.C.F.; Silva, L.T.D.; Andrade, A.L.; Vasconcelos, M.A.D.; Pinheiro, U.; Teixeira, E.H.; Nagano, C.S.; Sampaio, A.H. A new mucin-binding lectin from the marine sponge Aplysina fulva (AFL) exhibits antibiofilm effects. Arch. Biochem. Biophys. 2019, 662, 169–176. [Google Scholar] [CrossRef]
- Tateno, H. SUEL-related lectins, a lectin family widely distributed throughout organisms. Biosci. Biotechnol. Biochem. 2010, 74, 1141–1144. [Google Scholar] [CrossRef] [Green Version]
- Gerdol, M.; Greco, S.; Pallavicini, A. Extensive tandem duplication events drive the expansion of the C1q-domain-containing gene family in bivalves. Mar. Drugs 2019, 17, 583. [Google Scholar] [CrossRef] [Green Version]
- Ogawa, T.; Watanabe, M.; Naganuma, T.; Muramoto, K. Diversified carbohydrate-binding lectins from marine resources. J. Amino Acids 2011, 2011, 838914. [Google Scholar] [CrossRef] [Green Version]
- Sugawara, S.; Hosono, M.; Ogawa, Y.; Takayanagi, M.; Nitta, K. Molecular and sugar-binding heterogeneity of C-type lectins from Osmerus (Spirinchus) lanceolatus eggs. Biol. Pharm. Bull. 2005, 28, 791–796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stowell, S.R.; Arthur, C.M.; McBride, R.; Berger, O.; Razi, N.; Heimburg-Molinaro, J.; Rodrigues, L.C.; Gourdine, J.-P.; Noll, A.J.; Von Gunten, S. Microbial glycan microarrays define key features of host-microbial interactions. Nat. Chem. Biol. 2014, 10, 470. [Google Scholar] [CrossRef] [Green Version]
- Qiu, L.; Lin, L.; Yang, K.; Zhang, H.; Li, J.; Zou, F.; Jiang, S. Molecular cloning and expression analysis of a F-type lectin gene from Japanese sea perch (Lateolabrax japonicus). Mol. Biol. Rep. 2011, 38, 3751–3756. [Google Scholar] [CrossRef]
- Dara, M.; Giulianini, P.G.; Manfrin, C.; Parisi, M.G.; Parrinello, D.; La Corte, C.; Vasta, G.R.; Cammarata, M. F-type lectin from serum of the Antarctic teleost fish Trematomus bernacchii (Boulenger, 1902): Purification, structural characterization, and bacterial agglutinating activity. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2021, 256, 110633. [Google Scholar] [CrossRef]
- Xu, X.; Liao, K.; Shi, P.; Xu, J.; Ran, Z.; Zhou, C.; Zhang, L.; Cao, J.; Yan, X. Involvement of a novel Ca(2+)-independent C-type lectin from Sinonovacula constricta in food recognition and innate immunity. Fish Shellfish Immunol. 2020, 104, 374–382. [Google Scholar] [CrossRef]
- Okamoto, M.; Tsutsui, S.; Tasumi, S.; Suetake, H.; Kikuchi, K.; Suzuki, Y. Tandem repeat l-rhamnose-binding lectin from the skin mucus of ponyfish, Leiognathus nuchalis. Biochem. Biophys. Res. Commun. 2005, 333, 463–469. [Google Scholar] [CrossRef] [PubMed]
- Patel, D.M.; Kitani, Y.; Korsnes, K.; Iversen, M.H.; Brinchmann, M.F. A truncated galectin-3 isolated from skin mucus of atlantic salmon salmo salar binds to and modulates the proteome of the Gram-Negative Bacteria Moritella viscosa. Mar. Drugs 2020, 18, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zelensky, A.N.; Gready, J.E. C-type lectin-like domains in Fugu rubripes. BMC Genom. 2004, 5, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, G.; Lei, Y.; Kang, T.; Li, Z.; Fei, H.; Zeng, B.; Zhou, P.; Wang, C.; Lv, Z.; Huang, M.; et al. Two C-type lectins (ReCTL-1, ReCTL-2) from Rimicaris exoculata display broad nonself recognition spectrum with novel carbohydrate binding specificity. Fish Shellfish Immunol. 2020, 96, 152–160. [Google Scholar] [CrossRef]
- Zhang, H.; Robison, B.; Thorgaard, G.H.; Ristow, S.S. Cloning, mapping and genomic organization of a fish C-type lectin gene from homozygous clones of rainbow trout (Oncorhynchus mykiss). Biochim. Et Biophys. Acta (BBA)—Gene Struct. Expr. 2000, 1494, 14–22. [Google Scholar] [CrossRef]
- Ojeda, N.; Salazar, C.; Cárdenas, C.; Marshall, S. Expression of DC-SIGN-Like C-Type Lectin Receptors in Salmo salar. Dev. Comp. Immunol. 2020, 113, 103806. [Google Scholar] [CrossRef]
- Xue, D.; Guang-hua, W.; Yan-li, S.; Min, Z.; Yong-hua, H. Black rockfish C-type lectin, SsCTL4: A pattern recognition receptor that promotes bactericidal activity and virus escape from host immune defense. Fish Shellfish Immunol. 2018, 79, 340–350. [Google Scholar] [CrossRef]
- Yang, Q.; Wang, P.; Wang, S.; Wang, Y.; Feng, S.; Zhang, S.; Li, H. The hepatic lectin of zebrafish binds a wide range of bacteria and participates in immune defense. Fish Shellfish Immunol. 2018, 82, 267–278. [Google Scholar] [CrossRef]
- Yang, Q.; Wang, P.; Yang, S.; Li, X.; Zhang, X.; Ji, G.; Zhang, S.; Wang, S.; Li, H. A novel hepatic lectin of zebrafish Danio rerio is involved in innate immune defense. Fish Shellfish Immunol. 2020, 98, 670–680. [Google Scholar] [CrossRef]
- Ju, C.S.; He, L.B.; Pei, Y.Y.; Jiang, Y.; Huang, R.; Li, Y.M.; Liao, L.J.; Jang, S.H.; Wang, Y.P. Differential expression of two C-type lectins in grass carp Ctenopharyngodon idella and their response to grass carp reovirus. J. Fish Biol. 2016, 88, 787–793. [Google Scholar] [CrossRef]
- Li, D.; Nie, H.; Jahan, K.; Yan, X. Expression analyses of C-type lectins (CTLs) in Manila clam under cold stress provide insights for its potential function in cold resistance of Ruditapes philippinarum. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2020, 230, 108708. [Google Scholar] [CrossRef] [PubMed]
- Mohanty, J.; Sahoo, S.; Badhe, M.R.; Pillai, B.R.; Sahoo, P.K.; Suryawanshi, A.R.; Patnaik, B.B. Lectin-like activity of hemocyanin in freshwater prawn, Macrobrachium rosenbergii. Protein J. 2020, 39, 358–365. [Google Scholar] [CrossRef]
- Ragasa, L.R.P.; Dinglasan, J.L.N.; Felipe, I.R.E.; Basiao, Z.U.; Velarde, M.C. Exposure to Aeromonas hydrophila induces inflammation and increases expression of the gene encoding for a putative dual CTLD-containing lectin in milkfish liver. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2019, 230, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.W.; Yang, C.H.; Zhang, H.Q.; Pan, X.T.; Jin, Z.Y.; Zhang, H.W.; Xia, X.H. A C-type lectin with antibacterial activity in weather loach, Misgurnus anguillicaudatus. J. Fish Dis. 2020, 43, 1531–1539. [Google Scholar] [CrossRef]
- Cooper, D.N.W.; Barondes, S.H. God must love galectins; He made so many of them2. Glycobiology 1999, 9, 979–984. [Google Scholar] [CrossRef]
- Stancic, M.; Slijepcevic, D.; Nomden, A.; Vos, M.J.; De Jonge, J.C.; Sikkema, A.H.; Gabius, H.J.; Hoekstra, D.; Baron, W. Galectin-4, a novel neuronal regulator of myelination. Glia 2012, 60, 919–935. [Google Scholar] [CrossRef]
- McSharry, B.P.; Forbes, S.K.; Cao, J.Z.; Avdic, S.; Machala, E.A.; Gottlieb, D.J.; Abendroth, A.; Slobedman, B. Human cytomegalovirus upregulates expression of the lectin galectin 9 via induction of beta interferon. J. Virol. 2014, 88, 10990–10994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tandon, R.; Chew, G.M.; Byron, M.M.; Borrow, P.; Niki, T.; Hirashima, M.; Barbour, J.D.; Norris, P.J.; Lanteri, M.C.; Martin, J.N. Galectin-9 is rapidly released during acute HIV-1 infection and remains sustained at high levels despite viral suppression even in elite controllers. AIDS Res. Hum. Retrovir. 2014, 30, 654–664. [Google Scholar] [CrossRef] [Green Version]
- Niu, J.; Huang, Y.; Liu, X.; Luo, G.; Tang, J.; Wang, B.; Lu, Y.; Cai, J.; Jian, J. Functional characterization of galectin-3 from Nile tilapia (Oreochromis niloticus) and its regulatory role on monocytes/macrophages. Fish Shellfish Immunol. 2019, 95, 268–276. [Google Scholar] [CrossRef]
- Dutta, S.; Sinha, B.; Bhattacharya, B.; Chatterjee, B.; Mazumder, S. Characterization of a galactose binding serum lectin from the Indian catfish, Clarias batrachus: Possible involvement of fish lectins in differential recognition of pathogens. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2005, 141, 76–84. [Google Scholar] [CrossRef]
- Jackson, A.N.; McLure, C.A.; Dawkins, R.L.; Keating, P.J. Mannose binding lectin (MBL) copy number polymorphism in Zebrafish (D. rerio) and identification of haplotypes resistant to L. anguillarum. Immunogenetics 2007, 59, 861–872. [Google Scholar] [CrossRef] [PubMed]
- Marques, D.N.; De Almeida, A.S.; de Oliveira Sousa, A.R.; Pereira, R.; Andrade, A.L.; Chaves, R.P.; Carneiro, R.F.; de Vasconcelos, M.A.; do Nascimento-Neto, L.G.; Pinheiro, U. Antibacterial activity of a new lectin isolated from the marine sponge Chondrilla caribensis. Int. J. Biol. Macromol. 2018, 109, 1292–1301. [Google Scholar] [CrossRef]
- Ourth, D.D.; Rose, W.M.; Siefkes, M.J. Isolation of mannose-binding C-type lectin from sea lamprey (Petromyzon marinus) plasma and binding to Aeromonas salmonicida. Vet. Immunol. Immunopathol. 2008, 126, 407–412. [Google Scholar] [CrossRef] [PubMed]
- Van Der Vaart, M.; Spaink, H.P.; Meijer, A.H. Pathogen recognition and activation of the innate immune response in zebrafish. Adv. Hematol. 2012, 2012, 159807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubeena, A.S.; Preetham, E. Antimicrobial properties and phenoloxidase activation of the lectin isolated from kadal shrimp (Metapenaeus dobsoni). Fish Shellfish Immunol. 2019, 90, 118–125. [Google Scholar] [CrossRef] [PubMed]
- Endo, Y.; Matsushita, M.; Fujita, T. Role of ficolin in innate immunity and its molecular basis. Immunobiology 2007, 212, 371–379. [Google Scholar] [CrossRef]
- Mu, L.; Wu, H.; Han, K.; Wu, L.; Bian, X.; Li, B.; Guo, Z.; Yin, X.; Ye, J. Molecular and functional characterization of a mannose-binding lectin/ficolin-associated protein (MAp44) from Nile tilapia (Oreochromis niloticus) involved in the immune response to bacterial infection. Dev. Comp. Immunol. 2019, 101, 103438. [Google Scholar] [CrossRef]
- Magnadottir, B.; Gudmundsdottir, S.; Lange, S. A novel ladder-like lectin relates to sites of mucosal immunity in Atlantic halibut (Hippoglossus hippoglossus L.). Fish Shellfish Immunol. 2019, 87, 9–12. [Google Scholar] [CrossRef]
- Chernikov, O.; Kuzmich, A.; Chikalovets, I.; Molchanova, V.; Hua, K.-F. Lectin CGL from the sea mussel Crenomytilus grayanus induces Burkitt’s lymphoma cells death via interaction with surface glycan. Int. J. Biol. Macromol. 2017, 104, 508–514. [Google Scholar] [CrossRef]
- Sun, P.-P.; Ren, Y.-Y.; Zheng, J.; Hu, A.-J. Purification and characterization of a new lectin from loach skin mucus. J. Chem. 2019, 2019, 3853646. [Google Scholar] [CrossRef]
- Sanchez-Salgado, J.L.; Pereyra, M.A.; Vivanco-Rojas, O.; Sierra-Castillo, C.; Alpuche-Osorno, J.J.; Zenteno, E.; Agundis, C. Characterization of a lectin from the craysfish Cherax quadricarinatus hemolymph and its effect on hemocytes. Fish Shellfish Immunol. 2014, 39, 450–457. [Google Scholar] [CrossRef] [PubMed]
- Vazquez, L.; Massó, F.; Rosas, P.; Montaño, L.F.; Zenteno, E. Purification and characterization of a lectin from Macrobrachium rosenbergh (Crustacea, Decapoda) hemolymph. Comp. Biochem. Physiol. Part B Comp. Biochem. 1993, 105, 617–623. [Google Scholar] [CrossRef]
- Alpuche, J.; Pereyra, A.; Agundis, C.; Rosas, C.; Pascual, C.; Slomianny, M.-C.; Vázquez, L.; Zenteno, E. Purification and characterization of a lectin from the white shrimp Litopenaeus setiferus (Crustacea decapoda) hemolymph. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2005, 1724, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Porath, J.; Ersson, B. Biospecific methods for purification and characterization of pneumococcal polysaccharide PnS XIV. In Proceedings of the Symposium on New Approaches for Inducing Natural Immunity to Pyogenic Organisms, Winter Park, FL, USA, 1 January 1975; National Institutes of Health: Bethesda, MD, USA, 1975; pp. 101–108. [Google Scholar]
- Adhya, M.; Singha, B.; Chatterjee, B.P. Purification and characterization of an N-acetylglucosamine specific lectin from marine bivalve Macoma birmanica. Fish Shellfish Immunol. 2009, 27, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Zanetta, J.P.; Timmerman, P.; Leroy, Y. Gas-liquid chromatography of the heptafluorobutyrate derivatives of the O-methyl-glycosides on capillary columns: A method for the quantitative determination of the monosaccharide composition of glycoproteins and glycolipids. Glycobiology 1999, 9, 255–266. [Google Scholar] [CrossRef]
- He, F. Bradford Protein Assay. Bio-Protocol 2011, 1, e45. [Google Scholar] [CrossRef]
- Bidlingmeyer, B.A.; Cohen, S.A.; Tarvin, T.L. Rapid analysis of amino acids using pre-column derivatization. J. Chromatogr. B Biomed. Sci. Appl. 1984, 336, 93–104. [Google Scholar] [CrossRef]
- Viswambari Devi, R.; Basilrose, M.R.; Mercy, P.D. Prospect for lectins in arthropods. Ital. J. Zool. 2010, 77, 254–260. [Google Scholar] [CrossRef]
- Kim, Y.M.; Park, K.-I.; Choi, K.-S.; Alvarez, R.A.; Cummings, R.D.; Cho, M. Lectin from the Manila Clam Ruditapes philippinarum Is Induced upon Infection with the Protozoan Parasite Perkinsus olseni. J. Biol. Chem. 2006, 281, 26854–26864. [Google Scholar] [CrossRef] [Green Version]
- Arasu, A.; Kumaresan, V.; Sathyamoorthi, A.; Palanisamy, R.; Prabha, N.; Bhatt, P.; Roy, A.; Thirumalai, M.K.; Gnanam, A.J.; Pasupuleti, M.; et al. Fish lily type lectin-1 contains β-prism architecture: Immunological characterization. Mol. Immunol. 2013, 56, 497–506. [Google Scholar] [CrossRef]
- Arasu, A.; Kumaresan, V.; Palanisamy, R.; Arasu, M.V.; Al-Dhabi, N.A.; Ganesh, M.-R.; Arockiaraj, J. Bacterial membrane binding and pore formation abilities of carbohydrate recognition domain of fish lectin. Dev. Comp. Immunol. 2017, 67, 202–212. [Google Scholar] [CrossRef] [PubMed]
- Brinchmann, M.F.; Patel, D.M.; Pinto, N.; Iversen, M.H. Functional aspects of fish mucosal lectins—Interaction with non-self. Molecules 2018, 23, 1119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, D.M.; Brinchmann, M.F. Skin mucus proteins of lumpsucker (Cyclopterus lumpus). Biochem. Biophys. Rep. 2017, 9, 217–225. [Google Scholar] [CrossRef] [PubMed]
- Rajan, B.; Patel, D.M.; Kitani, Y.; Viswanath, K.; Brinchmann, M.F. Novel mannose binding natterin-like protein in the skin mucus of Atlantic cod (Gadus morhua). Fish Shellfish Immunol. 2017, 68, 452–457. [Google Scholar] [CrossRef] [PubMed]
- Magalhães, G.S.; Lopes-Ferreira, M.; Junqueira-de-Azevedo, I.L.; Spencer, P.J.; Araújo, M.; Portaro, F.; Ma, L.; Valente, R.; Juliano, L.; Fox, J. Natterins, a new class of proteins with kininogenase activity characterized from Thalassophryne nattereri fish venom. Biochimie 2005, 87, 687–699. [Google Scholar] [CrossRef]
- Wu, L.; Yang, X.; Duan, X.; Cui, L.; Li, G. Exogenous expression of marine lectins DlFBL and SpRBL induces cancer cell apoptosis possibly through PRMT5-E2F-1 pathway. Sci. Rep. 2014, 4, 4505. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Gao, Y.; Cui, L.; Wu, L.; Yang, X.; Chen, J. Anguilla japonica lectin 1 delivery through adenovirus vector induces apoptotic cancer cell death through interaction with PRMT5. J. Gene Med. 2016, 18, 65–74. [Google Scholar] [CrossRef]
- Rajan, B.; Fernandes, J.M.; Caipang, C.M.; Kiron, V.; Rombout, J.H.; Brinchmann, M.F. Proteome reference map of the skin mucus of Atlantic cod (Gadus morhua) revealing immune competent molecules. Fish Shellfish Immunol. 2011, 31, 224–231. [Google Scholar] [CrossRef]
- Rajan, B.; Kiron, V.; Fernandes, J.M.; Brinchmann, M.F. Localization and functional properties of two galectin-1 proteins in Atlantic cod (Gadus morhua) mucosal tissues. Dev. Comp. Immunol. 2013, 40, 83–93. [Google Scholar] [CrossRef]
- Cha, G.-H.; Liu, Y.; Peng, T.; Huang, M.-Z.; Xie, C.-Y.; Xiao, Y.-C.; Wang, W.-N. Molecular cloning, expression of a galectin gene in Pacific white shrimp Litopenaeus vannamei and the antibacterial activity of its recombinant protein. Mol. Immunol. 2015, 67 Pt B, 325–340. [Google Scholar] [CrossRef]
- Provan, F.; Jensen, L.; Uleberg, K.; Larssen, E.; Rajalahti, T.; Mullins, J.; Obach, A. Proteomic analysis of epidermal mucus from sea lice–infected A tlantic salmon, S almo salar L. J. Fish Dis. 2013, 36, 311–321. [Google Scholar] [CrossRef]
- Cordero, H.; Brinchmann, M.F.; Cuesta, A.; Meseguer, J.; Esteban, M.A. Skin mucus proteome map of European sea bass (Dicentrarchus labrax). Proteomics 2015, 15, 4007–4020. [Google Scholar] [CrossRef] [Green Version]
- Tunkijjanukij, S.; Mikkelsen, H.V.; Olafsen, J.A. A Heterogeneous Sialic Acid-Binding Lectin with Affinity for Bacterial LPS from Horse Mussel (Modiolus modiolus) Hemolymph. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 1997, 117, 273–286. [Google Scholar] [CrossRef]
- Chikalovets, I.V.; Kovalchuk, S.N.; Litovchenko, A.P.; Molchanova, V.I.; Pivkin, M.V.; Chernikov, O.V. A new Gal/GalNAc-specific lectin from the mussel Mytilus trossulus: Structure, tissue specificity, antimicrobial and antifungal activity. Fish Shellfish Immunol. 2016, 50, 27–33. [Google Scholar] [CrossRef]
- Bao, Y.; Shen, H.; Zhou, H.; Dong, Y.; Lin, Z. A tandem-repeat galectin from blood clam Tegillarca granosa and its induced mRNA expression response against bacterial challenge. Genes Genom. 2013, 35, 733–740. [Google Scholar] [CrossRef]
- He, J.; Shen, C.; Liang, H.; Fang, X.; Lu, J. Antimicrobial properties and immune-related gene expression of a C-type lectin isolated from Pinctada fucata martensii. Fish Shellfish Immunol. 2020, 105, 330–340. [Google Scholar] [CrossRef]
- Nair, S.V.; Burandt, M.; Hutchinson, A.; Raison, R.L.; Raftos, D.A. A C-type lectin from the Tunicate, Styela plicata, that modulates cellular activity. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2001, 129, 11–24. [Google Scholar] [CrossRef]
- Schröder, H.C.; Ushijima, H.; Krasko, A.; Gamulin, V.; Thakur, N.L.; Diehl-Seifert, B.; Müller, I.M.; Müller, W.E. Emergence and Disappearance of an Immune Molecule, an Antimicrobial Lectin, in Basal Metazoa: A Tachylectin-Related Protein in the Sponge Suberites Domuncula. J. Biol. Chem. 2003, 278, 32810–32817. [Google Scholar] [CrossRef] [Green Version]
- Hasan, I.; Ozeki, Y. Histochemical localization of N-acetylhexosamine-binding lectin HOL-18 in Halichondria okadai (Japanese black sponge), and its antimicrobial and cytotoxic anticancer effects. Int. J. Biol. Macromol. 2019, 124, 819–827. [Google Scholar] [CrossRef]
- Zhang, Q.; Wang, X.-Q.; Jiang, H.-S.; Jia, W.-M.; Zhao, X.-F.; Wang, J.-X. Calnexin functions in antibacterial immunity of Marsupenaeus japonicus. Dev. Comp. Immunol. 2014, 46, 356–363. [Google Scholar] [CrossRef]
- Jing, X.; Espinosa, E.P.; Perrigault, M.; Allam, B. Identification, molecular characterization and expression analysis of a mucosal C-type lectin in the eastern oyster, Crassostrea virginica. Fish Shellfish Immunol. 2011, 30, 851–858. [Google Scholar] [CrossRef]
- Guardiola, F.A.; Dioguardi, M.; Parisi, M.G.; Trapani, M.R.; Meseguer, J.; Cuesta, A.; Cammarata, M.; Esteban, M.A. Evaluation of waterborne exposure to heavy metals in innate immune defences present on skin mucus of gilthead seabream (Sparus aurata). Fish Shellfish Immunol. 2015, 45, 112–123. [Google Scholar] [CrossRef]
- Sinha, S.; Mandal, C.; Allen, A.K.; Mandal, C. Acute Phase Response of C-Reactive Protein of Labeo rohita to Aquatic Pollutants Is Accompanied by the Appearance of Distinct Molecular Forms. Arch. Biochem. Biophys. 2001, 396, 139–150. [Google Scholar] [CrossRef]
- Bretting, H.; Jacobs, G.; Donadey, C.; Vacelet, J. Immunohistochemical studies on the distribution and the function of the d-galactose-specific lectins in the sponge Axinella polypoides (Schmidt). Cell Tissue Res. 1983, 229, 551–571. [Google Scholar] [CrossRef]
- Ewart, K.V.; Li, Z.; Yang, D.S.C.; Fletcher, G.L.; Hew, C.L. The Ice-Binding Site of Atlantic Herring Antifreeze Protein Corresponds to the Carbohydrate-Binding Site of C-Type Lectins. Biochemistry 1998, 37, 4080–4085. [Google Scholar] [CrossRef]
- Achenbach, J.C.; Ewart, K.V. Structural and functional characterization of a C-type lectin-like antifreeze protein from rainbow smelt (Osmerus mordax). Eur. J. Biochem. 2002, 269, 1219–1226. [Google Scholar] [CrossRef]
- Yasumasu, S.; Wardrip, N.J.; Zenner, B.D.; Lee, Y.M.; Smith, A.J.; Hedrick, J.L. Fertilisation in fish: A cortical alveolar lectin and its potential role in the block to polyspermy. Zygote 1999, 8 (Suppl. 1), S66. [Google Scholar] [CrossRef]
- Dong, C.-H.; Yang, S.-T.; Yang, Z.-A.; Zhang, L.; Gui, J.-F. A C-type lectin associated and translocated with cortical granules during oocyte maturation and egg fertilization in fish. Dev. Biol. 2004, 265, 341–354. [Google Scholar] [CrossRef] [Green Version]
- Odintsova, N.A.; Belogortseva, N.I.; Ermak, A.V.; Molchanova, V.I.; Luk’yanov, P.A. Adhesive and growth properties of lectin from the ascidian Didemnum ternatanum on cultivated marine invertebrate cells. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 1999, 1448, 381–389. [Google Scholar] [CrossRef] [Green Version]
- Cheung, R.C.F.; Wong, J.H.; Pan, W.; Chan, Y.S.; Yin, C.; Dan, X.; Ng, T.B. Marine lectins and their medicinal applications. Appl. Microbiol. Biotechnol. 2015, 99, 3755–3773. [Google Scholar] [CrossRef]
- Silva, N.R.G.; Araújo, F.N.D. Antibacterial Activity of Plant Lectins: A Review. Braz. Arch. Biol. Technol. 2021, 64, e21200631. [Google Scholar] [CrossRef]
- Coelho, L.C.B.B.; Silva, P.M.D.S.; Lima, V.L.D.M.; Pontual, E.V.; Paiva, P.M.G.; Napoleao, T.H.; Correia, M.T.D.S. Lectins, interconnecting proteins with biotechnological/pharmacological and therapeutic applications. Evid.-Based Complement. Altern. Med. 2017, 2017, 1594074. [Google Scholar] [CrossRef]
- Sun, Y.-Y.; Liu, L.; Li, J.; Sun, L. Three novel B-type mannose-specific lectins of Cynoglossus semilaevis possess varied antibacterial activities against Gram-negative and Gram-positive bacteria. Dev. Comp. Immunol. 2016, 55, 194–202. [Google Scholar] [CrossRef]
- Tateno, H.; Yamaguchi, T.; Ogawa, T.; Muramoto, K.; Watanabe, T.; Kamiya, H.; Saneyoshi, M. Immunohistochemical localization of rhamnose-binding lectins in the steelhead trout (Oncorhynchus mykiss). Dev. Comp. Immunol. 2002, 26, 543–550. [Google Scholar] [CrossRef]
- Li, M.; Li, C.; Ma, C.; Li, H.; Zuo, H.; Weng, S.; Chen, X.; Zeng, D.; He, J.; Xu, X. Identification of a C-type lectin with antiviral and antibacterial activity from pacific white shrimp Litopenaeus vannamei. Dev. Comp. Immunol. 2014, 46, 231–240. [Google Scholar] [CrossRef]
- Kawabata, S.-i.; Tokunaga, F.; Kugi, Y.; Motoyama, S.; Miura, Y.; Hirata, M.; Iwanaga, S. Limulus factor D, a 43-kDa protein isolated from horseshoe crab hemocytes, is a serine protease homologue with antimicrobial activity. FEBS Lett. 1996, 398, 146–150. [Google Scholar] [CrossRef] [Green Version]
- Moura, R.M.; Queiroz, A.F.; Fook, J.M.; Dias, A.S.; Monteiro, N.K.; Ribeiro, J.K.; Moura, G.E.; Macedo, L.L.; Santos, E.A.; Sales, M.P. CvL, a lectin from the marine sponge Cliona varians: Isolation, characterization and its effects on pathogenic bacteria and Leishmania promastigotes. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2006, 145, 517–523. [Google Scholar] [CrossRef]
- Takahashi, K.G.; Kuroda, T.; Muroga, K. Purification and antibacterial characterization of a novel isoform of the Manila clam lectin (MCL-4) from the plasma of the Manila clam, Ruditapes philippinarum. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2008, 150, 45–52. [Google Scholar] [CrossRef]
- Liu, S.; Hu, G.; Sun, C.; Zhang, S. Anti-viral activity of galectin-1 from flounder Paralichthys olivaceus. Fish Shellfish Immunol. 2013, 34, 1463–1469. [Google Scholar] [CrossRef]
- Luo, T.; Zhang, X.; Shao, Z.; Xu, X. PmAV, a novel gene involved in virus resistance of shrimp Penaeus monodon. FEBS Lett. 2003, 551, 53–57. [Google Scholar] [CrossRef]
- Wang, J.-H.; Kong, J.; Li, W.; Molchanova, V.; Chikalovets, I.; Belogortseva, N.; Luk’yanov, P.; Zheng, Y.-T. A β-galactose-specific lectin isolated from the marine worm Chaetopterus variopedatus possesses anti-HIV-1 activity. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2006, 142, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Molchanova, V.; Chikalovets, I.; Chernikov, O.; Belogortseva, N.; Li, W.; Wang, J.-H.; Yang, D.-Y.O.; Zheng, Y.-T.; Lukyanov, P. A new lectin from the sea worm Serpula vermicularis: Isolation, characterization and anti-HIV activity. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2007, 145, 184–193. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y.; Morimoto, K.; Hirayama, M.; Hori, K. High mannose-specific lectin (KAA-2) from the red alga Kappaphycus alvarezii potently inhibits influenza virus infection in a strain-independent manner. Biochem. Biophys. Res. Commun. 2011, 405, 291–296. [Google Scholar] [CrossRef] [PubMed]
- Mori, T.; O’Keefe, B.R.; Sowder, R.C.; Bringans, S.; Gardella, R.; Berg, S.; Cochran, P.; Turpin, J.A.; Buckheit, R.W.; McMahon, J.B. Isolation and characterization of griffithsin, a novel HIV-inactivating protein, from the red alga Griffithsia sp. J. Biol. Chem. 2005, 280, 9345–9353. [Google Scholar] [CrossRef] [Green Version]
- Bah, C.S.F.; Fang, E.F.; Ng, T.B.; Mros, S.; McConnell, M.; Bekhit, A.E.-D.A. Purification and characterization of a rhamnose-binding chinook salmon roe lectin with antiproliferative activity toward tumor cells and nitric oxide-inducing activity toward murine macrophages. J. Agric. Food Chem. 2011, 59, 5720–5728. [Google Scholar] [CrossRef]
- Queiroz, A.F.; Silva, R.A.; Moura, R.M.; Dreyfuss, J.L.; Paredes-Gamero, E.J.; Souza, A.C.; Tersariol, I.L.; Santos, E.A.; Nader, H.B.; Justo, G.Z. Growth inhibitory activity of a novel lectin from Cliona varians against K562 human erythroleukemia cells. Cancer Chemother. Pharmacol. 2009, 63, 1023–1033. [Google Scholar] [CrossRef]
- Yang, H.; Li, Z.; Wei, X.; Huang, R.; Qi, H.; Gao, Q.; Li, C.; Zhang, C. Detection and discrimination of alpha-fetoprotein with a label-free electrochemical impedance spectroscopy biosensor array based on lectin functionalized carbon nanotubes. Talanta 2013, 111, 62–68. [Google Scholar] [CrossRef]
- Bi, J.; Ning, M.; Xie, X.; Fan, W.; Huang, Y.; Gu, W.; Wang, W.; Wang, L.; Meng, Q. A typical C-type lectin, perlucin-like protein, is involved in the innate immune defense of whiteleg shrimp Litopenaeus vannamei. Fish Shellfish Immunol. 2020, 103, 293–301. [Google Scholar] [CrossRef]
- Sun, J.; Gao, L.; Huang, S.; Wang, L.; Yang, W.; Zhang, T.; Jin, Y.; Song, L. CLec-TM1-ERK-GSK3b pathway regulates vibrio splendidus-induced IL-17 production in oyster. J. Immunol. 2021, 207, 640–650. [Google Scholar] [CrossRef]
- Vasta, G.R.; Wang, J.X. Galectin-mediated immune recognition: Opsonic roles with contrasting outcomes in selected shrimp and bivalve mollusk species. Dev. Comp. Immunol. 2020, 110, 103721. [Google Scholar] [CrossRef]
- Zhang, M.; Liu, Y.; Song, C.; Ning, J.; Cui, Z. Characterization and functional analysis of a novel mannose-binding lectin from the swimming crab Portunus trituberculatus. Fish Shellfish Immunol. 2019, 89, 448–457. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhang, W.; Mu, C.; Li, R.; Song, W.; Ye, Y.; Shi, C.; Liu, L.; Wang, H.; Wang, C. Identification and characterization of a novel galectin from the mud crab Scylla paramamosain. Fish Shellfish Immunol. 2020, 98, 699–709. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; Ren, Z.; Mu, C.; Li, R.; Ye, Y.; Song, W.; Shi, C.; Liu, L.; Wang, C. Characterisation and functional analysis of an L-type lectin from the swimming crab Portunus trituberculatus. Gene 2018, 664, 27–36. [Google Scholar] [CrossRef]
- Liu, Y.; Su, Y.; Zhang, A.; Cui, Z. A c-type lectin highly expressed in portunus trituberculatus intestine functions in amp regulation and prophenoloxidase activation. Antibiotics 2021, 10, 541. [Google Scholar] [CrossRef]
- Lan, T.; Li, Z.; Peng, M.; Niu, D.; Li, Y.; Li, J. A four-CRD C-type lectin from razor clam Sinonovacula constricta mediates agglutination and phagocytosis. Gene 2020, 728, 144287. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Huo, H.; Fu, Q.; Yang, N.; Xue, T.; Zhuang, C.; Liu, X.; Wang, B.; Su, B.; Li, C. Identification and characterization of a C-type lectin in turbot (Scophthalmus maximus) which functioning as a pattern recognition receptor that binds and agglutinates various bacteria. Fish Shellfish Immunol. 2021, 115, 104–111. [Google Scholar] [CrossRef]
- Chen, H.; Cai, X.; Qiu, H.; Fang, J.; Wu, X. A novel C-type lectin from Crassostrea gigas involved in the innate defense against Vibrio alginolyticus. Biochem. Biophys. Res. Commun. 2021, 566, 155–163. [Google Scholar] [CrossRef]
- Yan, J.; Chen, L.; Liu, Z.; Chen, Y.; Sun, Y.; Han, J.; Feng, L. The D5 region of the intelectin domain is a new type of carbohydrate recognition domain in the intelectin gene family. Dev. Comp. Immunol. 2018, 85, 150–160. [Google Scholar] [CrossRef]
- Ding, Z.; Zhao, X.; Wang, J.; Zhang, F.; Wang, W.; Liu, H. Intelectin mediated phagocytosis and killing activity of macrophages in blunt snout bream (Megalobrama amblycephala). Fish Shellfish Immunol. 2019, 87, 129–135. [Google Scholar] [CrossRef]
- Zhang, K.; Liu, X.; Li, X.; Liu, Y.; Yu, H.; Liu, J.; Zhang, Q. Antibacterial functions of a novel fish-egg lectin from spotted knifejaw (Oplegnathus punctatus) during host defense immune responses. Dev. Comp. Immunol. 2020, 111, 103758. [Google Scholar] [CrossRef]
- Madusanka, R.K.; Priyathilaka, T.T.; Janson, N.D.; Kasthuriarachchi, T.D.W.; Jung, S.; Tharuka, M.D.N.; Lee, J. Molecular, transcriptional and functional delineation of Galectin-8 from black rockfish (Sebastes schlegelii) and its potential immunological role. Fish Shellfish Immunol. 2019, 93, 449–462. [Google Scholar] [CrossRef] [PubMed]
- Niu, J.; Luo, G.; Liu, X.; Huang, Y.; Tang, J.; Wang, B.; Lu, Y.; Cai, J.; Jian, J. Characterization and functional analysis of a galectin-related protein B from Nile tilapia involved in the immune response to bacterial infection. J. Fish Dis. 2021, 44, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, Y.; Tasumi, S.; Tsutsui, S.; Okamoto, M.; Suetake, H. Molecular diversity of skin mucus lectins in fish. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2003, 136, 723–730. [Google Scholar] [CrossRef]
- Huang, Y.; Shi, Y.; Hu, S.; Wu, T.; Zhao, Z. Characterization and Functional Analysis of Two Transmembrane C-Type Lectins in Obscure Puffer (Takifugu obscurus). Front. Immunol. 2020, 11, 436. [Google Scholar] [CrossRef]
- Huang, Y.; Li, Y.F.; Wang, R.X.; Xie, M.F.; Shi, Y.; Zhao, Z. Calreticulin functions in antimicrobial immunity of obscure puffer Takifugu obscurus. Mol. Immunol. 2021, 140, 77–86. [Google Scholar] [CrossRef]
- Liang, Z.G.; Li, L.; Chen, S.N.; Mao, M.G.; Nie, P. Expression and antibacterial analysis of galectin-8 and -9 genes in mandarin fish, Siniperca chuatsi. Fish Shellfish Immunol. 2020, 107, 463–468. [Google Scholar] [CrossRef]
- Huang, L.; Bai, L.; Chen, Y.; Wang, Q.; Sha, Z. Identification, expression profile and analysis of the antimicrobial activity of collectin 11 (CL-11, CL-K1), a novel complement-associated pattern recognition molecule, in half-smooth tongue sole (Cynoglossus semilaevis). Fish Shellfish Immunol. 2019, 95, 679–687. [Google Scholar] [CrossRef]
- Zhang, T.; Jiang, S.; Sun, L. A Fish Galectin-8 Possesses Direct Bactericidal Activity. Int. J. Mol. Sci. 2021, 22, 376. [Google Scholar] [CrossRef]
- Yu, M.; Zhou, S.; Ding, Y.; Guo, H.; Li, Y.; Huang, Q.; Zheng, X.; Xiu, Y. Molecular characterization and functional study of a tandem-repeat Galectin-9 from Japanese flounder (Paralichthys olivaceus). Fish Shellfish Immunol. 2021, 112, 23–30. [Google Scholar] [CrossRef]
- Choi, K.M.; Joo, M.S.; Cho, D.H.; Woo, W.S.; Kang, G.; Heo, M.J.; Kim, D.H.; Park, C.I. Molecular identification and mrna expression profiles of galectin-9 gene in red sea bream (Pagrus major) infected with pathogens. Animals 2021, 11, 139. [Google Scholar] [CrossRef]
- Dong, Z.; Li, Y.; Liu, X.; Su, L.; Zhou, S.; Xiu, Y. Molecular characterization, expression analysis and immune effect of Galectin-8 from Japanese flounder (Paralichthys olivaceus). Fish Shellfish Immunol. 2021, 111, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Dang, Y.; Nie, L.; Chen, J. Molecular and functional characterisation of a mannose-binding lectin-like gene from Japanese sea bass (Lateolabrax japonicus). Dev. Comp. Immunol. 2020, 104, 103558. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, C.A.; Ramessar, K.; O’Keefe, B.R. Antiviral lectins: Selective inhibitors of viral entry. Antivir. Res. 2017, 142, 37–54. [Google Scholar] [CrossRef] [PubMed]
- Hirayama, M.; Ly, B.M.; Hori, K. Biological activity, cDNA cloning and primary structure of lectin KSA-2 from the cultivated red alga Kappaphycus striatum (Schmitz) Doty ex Silva. Phytochem. Lett. 2015, 14, 99–105. [Google Scholar]
- Sato, Y.; Hirayama, M.; Morimoto, K.; Yamamoto, N.; Okuyama, S.; Hori, K. High mannose-binding lectin with preference for the cluster of α1–2-mannose from the green alga Boodlea coacta is a potent entry inhibitor of HIV-1 and influenza viruses. J. Biol. Chem. 2011, 286, 19446–19458. [Google Scholar] [CrossRef] [Green Version]
- Barton, C.; Kouokam, J.C.; Lasnik, A.B.; Foreman, O.; Cambon, A.; Brock, G.; Montefiori, D.C.; Vojdani, F.; McCormick, A.A.; O’Keefe, B.R. Activity of and effect of subcutaneous treatment with the broad-spectrum antiviral lectin griffithsin in two laboratory rodent models. Antimicrob. Agents Chemother. 2014, 58, 120–127. [Google Scholar] [CrossRef] [Green Version]
- O’Keefe, B.R.; Vojdani, F.; Buffa, V.; Shattock, R.J.; Montefiori, D.C.; Bakke, J.; Mirsalis, J.; d’Andrea, A.-L.; Hume, S.D.; Bratcher, B. Scaleable manufacture of HIV-1 entry inhibitor griffithsin and validation of its safety and efficacy as a topical microbicide component. Proc. Natl. Acad. Sci. USA 2009, 106, 6099–6104. [Google Scholar] [CrossRef] [Green Version]
- Férir, G.; Huskens, D.; Palmer, K.E.; Boudreaux, D.M.; Swanson, M.D.; Markovitz, D.M.; Balzarini, J.; Schols, D. Combinations of griffithsin with other carbohydrate-binding agents demonstrate superior activity against HIV type 1, HIV type 2, and selected carbohydrate-binding agent-resistant HIV type 1 strains. AIDS Res. Hum. Retrovir. 2012, 28, 1513–1523. [Google Scholar] [CrossRef] [Green Version]
- Meuleman, P.; Albecka, A.; Belouzard, S.; Vercauteren, K.; Verhoye, L.; Wychowski, C.; Leroux-Roels, G.; Palmer, K.E.; Dubuisson, J. Griffithsin has antiviral activity against hepatitis C virus. Antimicrob. Agents Chemother. 2011, 55, 5159–5167. [Google Scholar] [CrossRef] [Green Version]
- Takebe, Y.; Saucedo, C.J.; Lund, G.; Uenishi, R.; Hase, S.; Tsuchiura, T.; Kneteman, N.; Ramessar, K.; Tyrrell, D.L.J.; Shirakura, M. Antiviral lectins from red and blue-green algae show potent in vitro and in vivo activity against hepatitis C virus. PLoS ONE 2013, 8, e64449. [Google Scholar]
- Palù, G.; Reale, A.; Bazan, N.G.; Panda, P.K.; Uversky, V.N.; Seyran, M.; Aljabali, A.A.; Sherchan, S.P.; Azad, G.K.; Baetas-da-Cruz, W.; et al. Biology of Coronaviruses and Predicted Origin of SARS-CoV-2. In COVID-19; Debmalya, B., Kenneth, L., Eds.; CRC Press: Boca Raton, FL, USA, 2022; pp. 19–26. [Google Scholar]
- Seyran, M.; Pizzol, D.; Adadi, P.; El-Aziz, T.M.A.; Hassan, S.S.; Soares, A.; Kandimalla, R.; Lundstrom, K.; Tambuwala, M.; Aljabali, A.A. Questions concerning the proximal origin of SARS-CoV-2. J. Med. Virol. 2020, 93, 1204. [Google Scholar] [CrossRef] [PubMed]
- Hassan, S.; Ghosh, S.; Attrish, D.; Choudhury, P.P.; Aljabali, A.A.; Uhal, B.D.; Lundstrom, K.; Rezaei, N.; Uversky, V.N.; Seyran, M. Possible transmission flow of SARS-CoV-2 based on ACE2 features. Molecules 2020, 25, 5906. [Google Scholar] [CrossRef] [PubMed]
- Boopathi, S.; Poma, A.B.; Kolandaivel, P. Novel 2019 coronavirus structure, mechanism of action, antiviral drug promises and rule out against its treatment. J. Biomol. Struct. Dyn. 2020, 39, 3409–3418. [Google Scholar] [CrossRef] [Green Version]
- Kang, S.; Peng, W.; Zhu, Y.; Lu, S.; Zhou, M.; Lin, W.; Wu, W.; Huang, S.; Jiang, L.; Luo, X. Recent Progress in understanding 2019 novel coronavirus associated with human respiratory disease: Detection, mechanism and treatment. Int. J. Antimicrob. Agents 2020, 55, 105950. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Král, P. Computational Design of ACE2-Based Peptide Inhibitors of SARS-CoV-2. ACS Nano 2020, 14, 5143–5147. [Google Scholar] [CrossRef] [Green Version]
- Bianchi, M.; Benvenuto, D.; Giovanetti, M.; Angeletti, S.; Ciccozzi, M.; Pascarella, S. SARS-CoV-2 Envelope and Membrane Proteins: Structural Differences Linked to Virus Characteristics? BioMed Res. Int. 2020, 2020, 4389089. [Google Scholar] [CrossRef]
- Hassan, S.S.; Lundstrom, K.; Barh, D.; Silva, R.J.S.; Andrade, B.S.; Azevedo, V.; Choudhury, P.P.; Palu, G.; Uhal, B.D.; Kandimalla, R. Implications derived from S-protein variants of SARS-CoV-2 from six continents. Int. J. Biol. Macromol. 2021, 191, 934–955. [Google Scholar] [CrossRef]
- El-Maradny, Y.A.; El-Fakharany, E.M.; Abu-Serie, M.M.; Hashish, M.H.; Selim, H.S. Lectins purified from medicinal and edible mushrooms: Insights into their antiviral activity against pathogenic viruses. Int. J. Biol. Macromol. 2021, 179, 239–258. [Google Scholar] [CrossRef]
- Kim, C.-H. SARS-CoV-2 Evolutionary Adaptation toward Host Entry and Recognition of Receptor O-Acetyl Sialylation in Virus–Host Interaction. Int. J. Mol. Sci. 2020, 21, 4549. [Google Scholar] [CrossRef]
- Rahimi, N. C-type lectin CD209L/L-SIGN and CD209/DC-SIGN: Cell adhesion molecules turned to pathogen recognition receptors. Biology 2021, 10, 1. [Google Scholar] [CrossRef]
- Singh, R.S.; Walia, A.K. Lectins from red algae and their biomedical potential. J. Appl. Phycol. 2018, 30, 1833–1858. [Google Scholar] [CrossRef] [Green Version]
- O’Keefe, B.R.; Giomarelli, B.; Barnard, D.L.; Shenoy, S.R.; Chan, P.K.; McMahon, J.B.; Palmer, K.E.; Barnett, B.W.; Meyerholz, D.K.; Wohlford-Lenane, C.L. Broad-spectrum in vitro activity and in vivo efficacy of the antiviral protein griffithsin against emerging viruses of the family Coronaviridae. J. Virol. 2010, 84, 2511–2521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, R.K.; Apte, G.R.; Lokhande, K.B.; Mishra, S.; Pal, J.K. Carbohydrate-binding agents: Potential of repurposing for COVID-19 therapy. Curr. Protein Pept. Sci. 2020, 21, 1085–1096. [Google Scholar] [CrossRef] [PubMed]
- Pereira, L.; Critchley, A.T. The COVID 19 novel coronavirus pandemic 2020: Seaweeds to the rescue? Why does substantial, supporting research about the antiviral properties of seaweed polysaccharides seem to go unrecognized by the pharmaceutical community in these desperate times? J. Appl. Phycol. 2020, 32, 1875–1877. [Google Scholar] [CrossRef]
- Lam, S.K.; Ng, T.B. Lectins: Production and practical applications. Appl. Microbiol. Biotechnol. 2011, 89, 45–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, P.; Wang, L.; Zhang, H.; Song, X.; Zhou, Z.; Yang, J.; Qiu, L.; Wang, L.; Song, L. A novel C-type lectin from bay scallop Argopecten irradians (AiCTL-7) agglutinating fungi with mannose specificity. Fish Shellfish Immunol. 2011, 30, 836–844. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.; Xu, D.; Zhou, J.; Cui, H.; Yan, Y.; Ouyang, Z.; Gong, J.; Huang, Y.; Huang, X.; Qin, Q. Molecular cloning, characterization and expression analysis of a C-type lectin (Ec-CTL) in orange-spotted grouper, Epinephelus coioides. Fish Shellfish Immunol. 2010, 28, 178–186. [Google Scholar] [CrossRef]
- Chikalovets, I.V.; Chernikov, O.V.; Pivkin, M.V.; Molchanova, V.I.; Litovchenko, A.P.; Li, W.; Lukyanov, P.A. A lectin with antifungal activity from the mussel Crenomytilus grayanus. Fish Shellfish Immunol. 2015, 42, 503–507. [Google Scholar] [CrossRef]
- Huang, C.-C.; Chen, C.-T.; Shiang, Y.-C.; Lin, Z.-H.; Chang, H.-T. Synthesis of fluorescent carbohydrate-protected Au nanodots for detection of Concanavalin A and Escherichia coli. Anal. Chem. 2009, 81, 875–882. [Google Scholar] [CrossRef]
- Zhou, Z.-j.; Sun, L. CsCTL1, a teleost C-type lectin that promotes antibacterial and antiviral immune defense in a manner that depends on the conserved EPN motif. Dev. Comp. Immunol. 2015, 50, 69–77. [Google Scholar] [CrossRef]
- Nabeta, H.W.; Kouokam, J.C.; Lasnik, A.B.; Fuqua, J.L.; Palmer, K.E. Novel antifungal activity of Q-Griffithsin, a broad-spectrum antiviral lectin. Microbiol. Spectr. 2021, 9, e00957-21. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.X.; Zhang, M.L.; Wang, X.W. C-Type Lectin maintains the homeostasis of intestinal microbiota and mediates biofilm formation by intestinal bacteria in shrimp. J. Immunol. 2021, 206, 1140–1150. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, Y.; Naganuma, T.; Ogawa, T.; Muramoto, K. Lectins of marine origin and their clinical applications. In Antitumor Potential and Other Emerging Medicinal Properties of Natural Compounds; Springer: Berlin/Heidelberg, Germany, 2013; pp. 33–54. [Google Scholar]
- Catanzaro, E.; Calcabrini, C.; Bishayee, A.; Fimognari, C. Antitumor potential of marine and freshwater lectins. Mar. Drugs 2020, 18, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, D.; Pan, S.; Zhou, M. Structural characterization and antitumor and mitogenic activity of a lectin from the gill of bighead carp (Aristichthys nobilis). Fish Physiol. Biochem. 2012, 38, 1815–1824. [Google Scholar] [CrossRef] [PubMed]
- Poisa-Beiro, L.; Dios, S.; Ahmed, H.; Vasta, G.R.; Martínez-López, A.; Estepa, A.; Alonso-Gutiérrez, J.; Figueras, A.; Novoa, B. Nodavirus infection of sea bass (Dicentrarchus labrax) induces up-regulation of galectin-1 expression with potential anti-inflammatory activity. J. Immunol. 2009, 183, 6600–6611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Chi, H.; Sun, B.-G.; Sun, L. The galectin-3-binding protein of Cynoglossus semilaevis is a secreted protein of the innate immune system that binds a wide range of bacteria and is involved in host phagocytosis. Dev. Comp. Immunol. 2013, 39, 399–408. [Google Scholar] [CrossRef]
- Kawano, T.; Sugawara, S.; Hosono, M.; Tatsuta, T.; Ogawa, Y.; Fujimura, T.; Taka, H.; Murayama, K.; Nitta, K. Globotriaosylceramide-expressing Burkitt’s lymphoma cells are committed to early apoptotic status by rhamnose-binding lectin from catfish eggs. Biol. Pharm. Bull. 2009, 32, 345–353. [Google Scholar] [CrossRef] [Green Version]
- Liao, J.-H.; Chien, C.-T.H.; Wu, H.-Y.; Huang, K.-F.; Wang, I.; Ho, M.-R.; Tu, I.-F.; Lee, I.-M.; Li, W.; Shih, Y.-L. A multivalent marine lectin from Crenomytilus grayanus possesses anti-cancer activity through recognizing globotriose Gb3. J. Am. Chem. Soc. 2016, 138, 4787–4795. [Google Scholar] [CrossRef]
- Matsumoto, R.; Fujii, Y.; Kawsar, S.; Kanaly, R.A.; Yasumitsu, H.; Koide, Y.; Hasan, I.; Iwahara, C.; Ogawa, Y.; Im, C.H. Cytotoxicity and glycan-binding properties of an 18 kDa lectin isolated from the marine sponge Halichondria okadai. Toxins 2012, 4, 323–338. [Google Scholar] [CrossRef] [Green Version]
- Fujii, Y.; Dohmae, N.; Takio, K.; Kawsar, S.M.; Matsumoto, R.; Hasan, I.; Koide, Y.; Kanaly, R.A.; Yasumitsu, H.; Ogawa, Y. A lectin from the mussel Mytilus galloprovincialis has a highly novel primary structure and induces glycan-mediated cytotoxicity of globotriaosylceramide-expressing lymphoma cells. J. Biol. Chem. 2012, 287, 44772–44783. [Google Scholar] [CrossRef] [Green Version]
- Hung, L.D.; Trinh, P.T.H. Structure and anticancer activity of a new lectin from the cultivated red alga, Kappaphycus striatus. J. Nat. Med. 2021, 75, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Ni, J.; Feng, H.; Xu, X.; Liu, T.; Ye, T.; Chen, K.; Li, G. Oncolytic vaccinia virus harboring aphrocallistes vastus lectin inhibits the growth of cervical cancer cells hela S3. Mar. Drugs 2021, 19, 532. [Google Scholar] [CrossRef] [PubMed]
- Brown, G.D.; Willment, J.A.; Whitehead, L. C-type lectins in immunity and homeostasis. Nat. Rev. Immunol. 2018, 18, 374–389. [Google Scholar] [CrossRef] [PubMed]
- Loh, S.H.; Park, J.-Y.; Cho, E.H.; Nah, S.-Y.; Kang, Y.-S. Animal lectins: Potential receptors for ginseng polysaccharides. J. Ginseng Res. 2017, 41, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Matsushita, M.; Kilpatrick, D.C.; Lee, B.L.; Wakamiya, N. Soluble host-defense lectins. J. Biomed. Biotechnol. 2012, 2012, 275970. [Google Scholar] [CrossRef] [Green Version]
- Drouin, M.; Saenz, J.; Chiffoleau, E. C-Type Lectin-Like Receptors: Head or Tail in Cell Death Immunity. Front. Immunol. 2020, 11, 251. [Google Scholar] [CrossRef] [Green Version]
- Vojdani, A.; Afar, D.; Vojdani, E. Reaction of Lectin-Specific Antibody with Human Tissue: Possible Contributions to Autoimmunity. J. Immunol. Res. 2020, 2020, 1438957. [Google Scholar] [CrossRef]
- Mayer, S.; Raulf, M.-K.; Lepenies, B. C-type lectins: Their network and roles in pathogen recognition and immunity. Histochem. Cell Biol. 2017, 147, 223–237. [Google Scholar] [CrossRef]
- Cordain, L.; Toohey, L.; Smith, M.; Hickey, M. Modulation of immune function by dietary lectins in rheumatoid arthritis. Br. J. Nutr. 2000, 83, 207–217. [Google Scholar] [CrossRef] [Green Version]
- Vasta, G.R.; Feng, C. F-Type Lectins: Structure, function, and evolution. In Methods in Molecular Biology; Humana Press Inc.: Totowa, NJ, USA, 2020; Volume 2132, pp. 225–239. [Google Scholar]
- Shao, Y.; Che, Z.; Xing, R.; Wang, Z.; Zhang, W.; Zhao, X.; Jin, C.; Li, C. Divergent immune roles of two fucolectin isoforms in Apostichopus japonicus. Dev. Comp. Immunol. 2018, 89, 1–6. [Google Scholar] [CrossRef]
- Vasta, G.R. Galectins in Host–Pathogen Interactions: Structural, Functional and Evolutionary Aspects. In Advances in Experimental Medicine and Biology; Springer: Berlin/Heidelberg, Germany, 2020; Volume 1204, pp. 169–196. [Google Scholar]
- Abreu, T.M.; Monteiro, V.S.; Martins, A.B.S.; Teles, F.B.; da Conceição Rivanor, R.L.; Mota, É.F.; Macedo, D.S.; de Vasconcelos, S.M.M.; Júnior, J.E.R.H.; Benevides, N.M.B. Involvement of the dopaminergic system in the antidepressant-like effect of the lectin isolated from the red marine alga Solieria filiformis in mice. Int. J. Biol. Macromol. 2018, 111, 534–541. [Google Scholar] [CrossRef] [PubMed]
- Espinosa, E.P.; Allam, B. Reverse genetics demonstrate the role of mucosal C-type lectins in food particle selection in the oyster Crassostrea virginica. J. Exp. Biol. 2018, 221, jeb174094. [Google Scholar] [CrossRef] [Green Version]
- Takeuchi, R.; Jimbo, M.; Tanimoto, F.; Iijima, M.; Yamashita, H.; Suzuki, G.; Harii, S.; Nakano, Y.; Yasumoto, K.; Watabe, S. N-acetyl-d-glucosamine-binding lectin in acropora tenuis attracts specific symbiodiniaceae cell culture strains. Mar. Drugs 2021, 19, 146. [Google Scholar] [CrossRef]
- Zhang, C.; Li, X.; Kim, S.-K. Application of marine biomaterials for nutraceuticals and functional foods. Food Sci. Biotechnol. 2012, 21, 625–631. [Google Scholar] [CrossRef]
- Axford, J. The impact of glycobiology on medicine. Trends Immunol. 2001, 22, 237–239. [Google Scholar] [CrossRef]
- Borman, S. Carbohydrate vaccines. Chem. Eng. News 2004, 82, 31–34. [Google Scholar] [CrossRef]
- Komath, S.S.; Kavitha, M.; Swamy, M.J. Beyond carbohydrate binding: New directions in plant lectin research. Org. Biomol. Chem. 2006, 4, 973–988. [Google Scholar] [CrossRef]
- He, S.; Simpson, B.K.; Sun, H.; Ngadi, M.O.; Ma, Y.; Huang, T. Phaseolus vulgaris lectins: A systematic review of characteristics and health implications. Crit. Rev. Food Sci. Nutr. 2018, 58, 70–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panacer, K.; Whorwell, P.J. Dietary Lectin exclusion: The next big food trend? World J. Gastroenterol. 2019, 25, 2973. [Google Scholar] [CrossRef]
- Burbano, C.; Muzquiz, M.; Ayet, G.; Cuadrado, C.; Pedrosa, M.M. Evaluation of antinutritional factors of selected varieties of Phaseolus vulgaris. J. Sci. Food Agric. 1999, 79, 1468–1472. [Google Scholar] [CrossRef]
- Vasconcelos, I.M.; Oliveira, J.T.A. Antinutritional properties of plant lectins. Toxicon 2004, 44, 385–403. [Google Scholar] [CrossRef] [PubMed]
- Gong, T.; Wang, X.; Yang, Y.; Yan, Y.; Yu, C.; Zhou, R.; Jiang, W. Plant lectins activate the NLRP3 inflammasome to promote inflammatory disorders. J. Immunol. 2017, 198, 2082–2092. [Google Scholar] [CrossRef] [PubMed] [Green Version]







| Types of Organisms | Lectin Family | Specificity | Tissue Expression | Features/Functions | References |
|---|---|---|---|---|---|
| Turbot (Scophthalmus maximus L.) | RBL | l-rhamnose or d-galactose | Egg cortex and ovary cells |
| [27] |
| Purplish bifurcate mussel (Mytilisepta virgata) | RBL | Sevil, a glycan binding lectin | Mantles and gills |
| [28] |
| Sea urchin (Anthocidaris crassispina) | RBL (SUEL-type) | d-Galactose | Eggs |
| [29] |
| Southern catfish (Silurus meridionalis) | RBL | l-rhamnose | Gill, barbel |
| [30] |
| Nile tilapia (Oreochromis niloticus) | RBL | l-rhamnose | Liver, gills, intestines |
| [31] |
| Catfish (Silurus asotus) | RBL | α-galactoside | Eggs |
| [32,33,34] |
| Turbot (Scophthalmus maximus L.) | RBL | l-rhamnose | Egg, ovaries |
| [27] |
| Chum salmon (Oncorhynchus keta) | RBL | l-rhamnose | Eggs |
| [35] |
| Shishamo smelt (Osmerus lanceolatus) | RBL | l-rhamnose | Eggs |
| [34] |
| Bay scallop (Argopecten irradians) | CTL | Galactose | Muscle, gonad, hepatopancreas, mantle margine, and gill |
| [36] |
| Horsemussels (Modiolus kurilensis) | CTL | Glycan | Hemolymph |
| [37] |
| Clam (Glycymeris yessoensis) | CTL | Peptidoglycan, LPS, β-1,3-glucan and mannan | Hemolymph |
| [38] |
| Kadal Shrimp (Metapenaeus dobsoni) | CTL | Glycan | Hemolymph |
| [39] |
| Molluscan snail (Hemifusus pugilinus) | CTL | Mannose | Haemolymph |
| [40] |
| Manila clam (Venerupis philippinarum) | CTL | Glucan | Gill tissues and hepatopancreas |
| [41] |
| Sea urchin (Pseudocentrotus depressus) | CTL | Mannose | Crushed body |
| [42] |
| Sea urchin (Pseudocentrotus depressus) | CTL | Glycan | Tube feet |
| [43] |
| Mud crab (Scylla paramamosain) | CTL | Glucan | Hemocytes, midgut, muscle, stomach, hapatopancreas, testis, ovaries, and heart |
| [44] |
| Brittle star (Ophioplocus japonicus) | CTL | Glucose/xylose | Whole body |
| [45] |
| Goldfish (Carassius auratus) | CTL | Mannose | Liver, spleen, kidney |
| [46] |
| Turbot (Scophthalmus maximus) Black rockfish (Sebastes schlegelii) (SsLTL) | LTL | d-mannose | Skin, gill, and intestine |
| [47,48,49] |
| Giant prawn (Macrobrachium rosenbergii) | LTL | Mannose | Hemocytes, intestine, and hepatopancreas |
| [50] |
| Sea bass (Dicentrarchus labrax L.) | FTL | Fucose-binding | Liver, larvae, eggs, intestine |
| [51,52] |
| Striped beakfish (rock bream) (Oplegnathus fasciatus) | FTL | Fucose-binding | Intestines |
| [53] |
| Striped Bass (Morone saxatilis) | FTL | Fucose-binding | Liver |
| [54] |
| sea bass (Dicentrarchus labrax) | FTL | l-rhamnose | Liver and intestine |
| [55] |
| Steelhead trout (Oncorhynchus mykiss) | FTL | l-rhamnose | Eggs |
| [56] |
| Sea bass (Dicentrarchus labrax) | FTL | l-fucose-binding | Intestine, liver |
| [57] |
| Atlantic Salmon (Salmo salar) | Galectin | Glycans | Gill epithelial cell |
| [58] |
| Striped snakehead (Channa striatus) | Galectin | Galactosidase | Liver |
| [9] |
| Korean rose bitterling (Rhodeus uyekii) | Galectin | β-galactoside | Liver, brain, kidney, ovary, gills, spleen |
| [59] |
| Turbot (Scophthalmus maximus L.) | Galectin | β-galactoside | Skin and brain |
| [60] |
| Euryhaline rotifers (Brachionus Plicatilis and Proales similis) | Galectins | Carbohydrate-binding domains with long N-terminal region (i.e., ~100 amino acids) β-galactosyl binding lectins |
| [61] | |
| Sea Hare (Aplysia kurodai) | GBL | d-galacturonic acid and d-galactose | Eggs |
| [62] |
| Pikeperch (Sander lucioperca), rainbow trout (Oncorhynchus mykiss), and maraena whitefish (Coregonus maraena) | Siglecs (Siglec1, Siglec15, CD22, and myelin-associated glycoprotein (MAG)) | Sialic-acid-binding | Head kidney, liver, gills, spleen, heart, and muscle |
| [63] |
| Marine sponge (Aplysina fulva) | AFL | Galactose | Crude extract |
| [64] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmmed, M.K.; Bhowmik, S.; Giteru, S.G.; Zilani, M.N.H.; Adadi, P.; Islam, S.S.; Kanwugu, O.N.; Haq, M.; Ahmmed, F.; Ng, C.C.W.; et al. An Update of Lectins from Marine Organisms: Characterization, Extraction Methodology, and Potential Biofunctional Applications. Mar. Drugs 2022, 20, 430. https://doi.org/10.3390/md20070430
Ahmmed MK, Bhowmik S, Giteru SG, Zilani MNH, Adadi P, Islam SS, Kanwugu ON, Haq M, Ahmmed F, Ng CCW, et al. An Update of Lectins from Marine Organisms: Characterization, Extraction Methodology, and Potential Biofunctional Applications. Marine Drugs. 2022; 20(7):430. https://doi.org/10.3390/md20070430
Chicago/Turabian StyleAhmmed, Mirja Kaizer, Shuva Bhowmik, Stephen G. Giteru, Md. Nazmul Hasan Zilani, Parise Adadi, Shikder Saiful Islam, Osman N. Kanwugu, Monjurul Haq, Fatema Ahmmed, Charlene Cheuk Wing Ng, and et al. 2022. "An Update of Lectins from Marine Organisms: Characterization, Extraction Methodology, and Potential Biofunctional Applications" Marine Drugs 20, no. 7: 430. https://doi.org/10.3390/md20070430