
Increased Sulfation in Gracilaria fisheri Sulfated Galactans Enhances Antioxidant and Antiurolithiatic Activities and Protects HK-2 Cell Death Induced by Sodium Oxalate

 ,
,
Abstract
:1. Introduction
2. Results
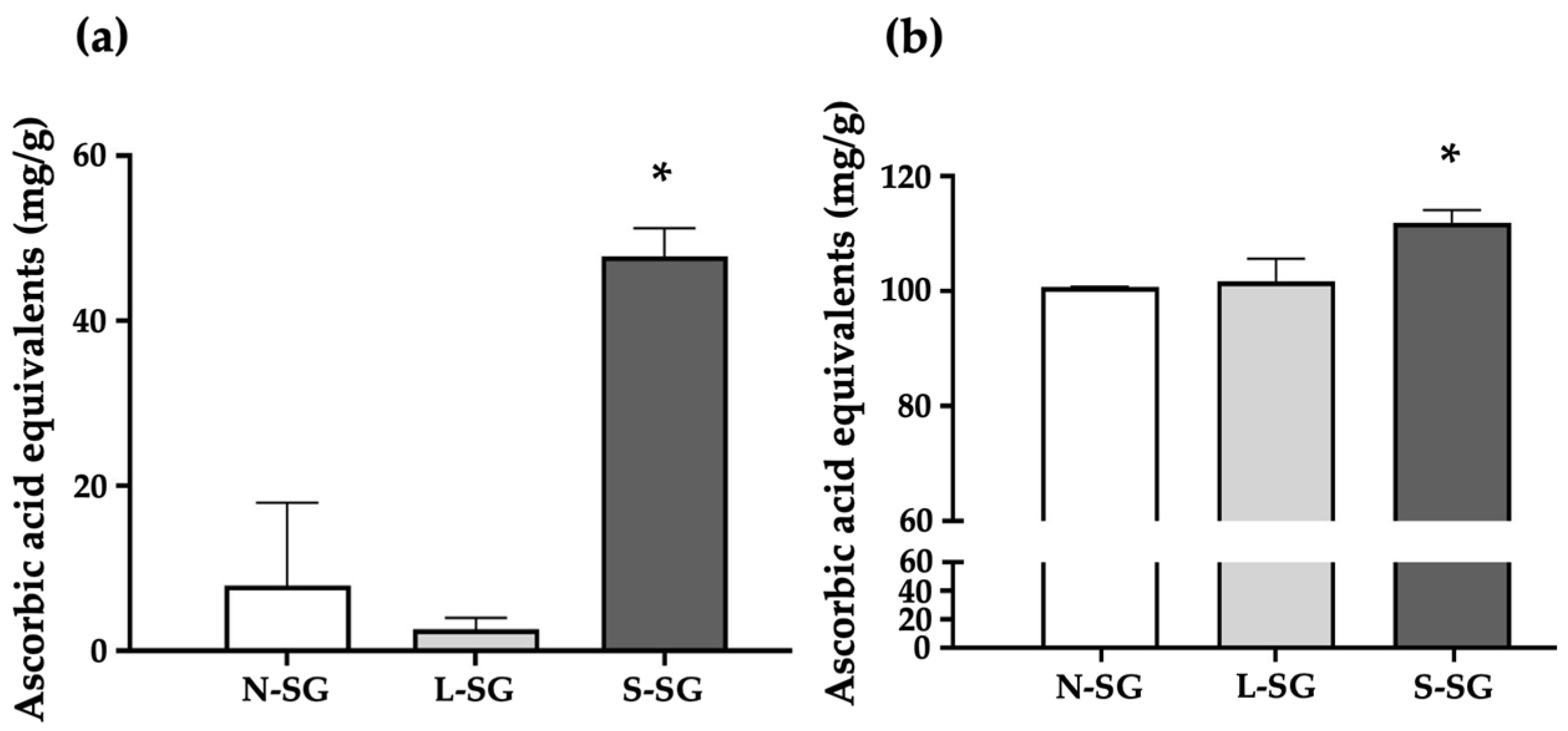
2.1. Sulfate Content and Average Molecular Weight of N-SG and Its Derivatives
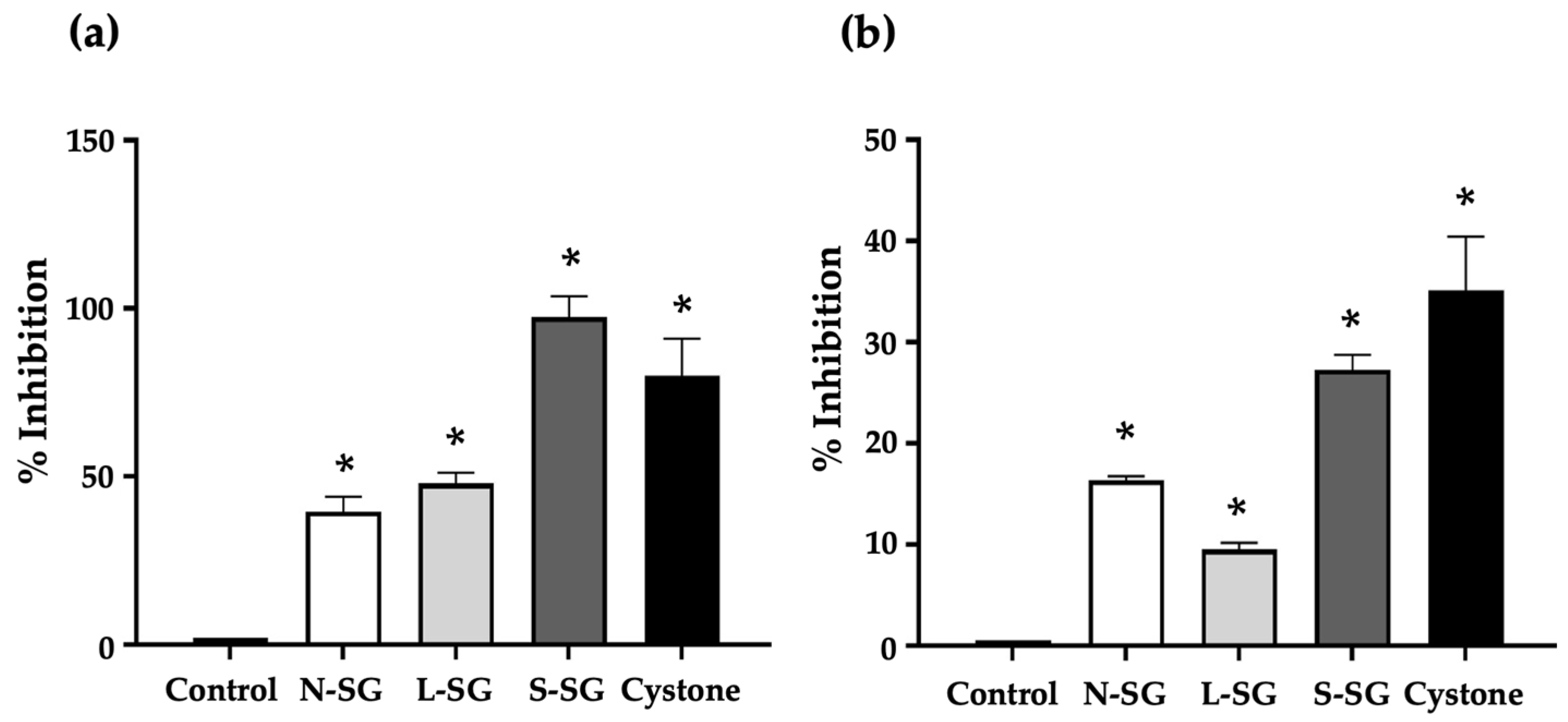
2.2. Antioxidant Activities of N-SG and Its Derivatives
2.3. Inhibitory Effect of N-SG and Its Derivatives on Calcium Oxalate Crystallization
2.4. Morphological Changes of Crystals after Treatment of N-SG and Its Derivatives
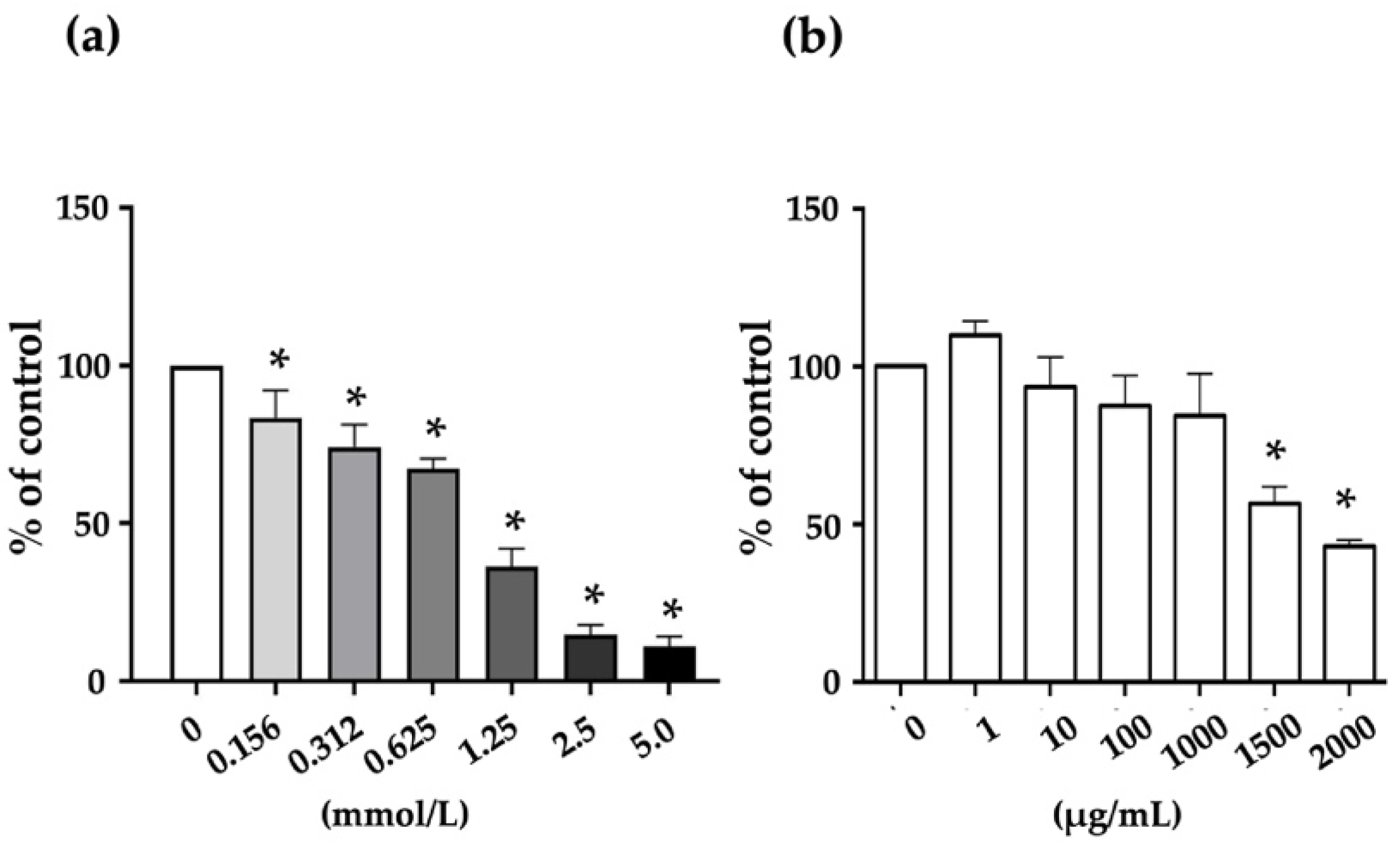
2.5. Cytotoxicity of Sodium Oxalate (NaOX) and S-SG on HK-2 Cells
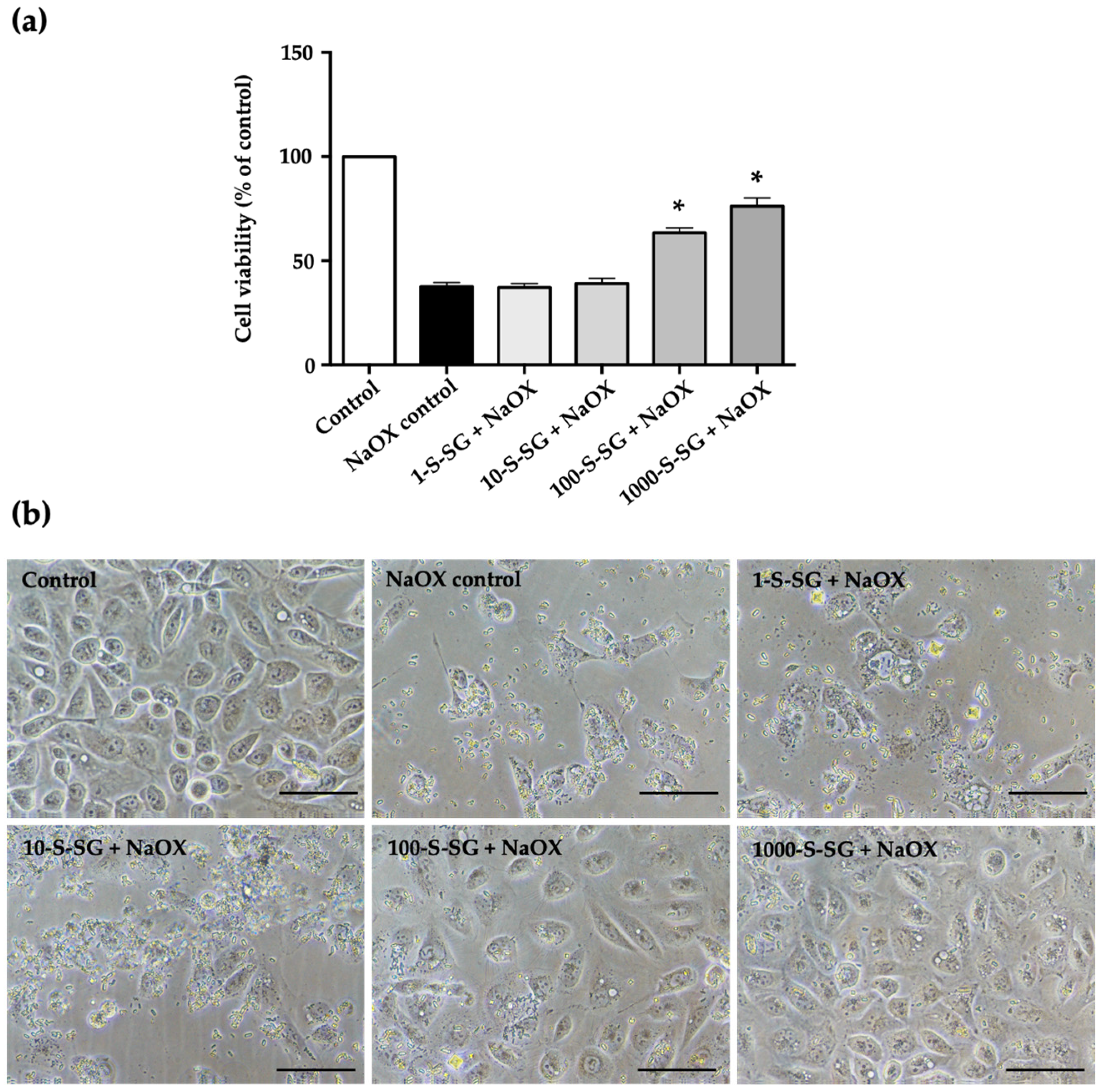
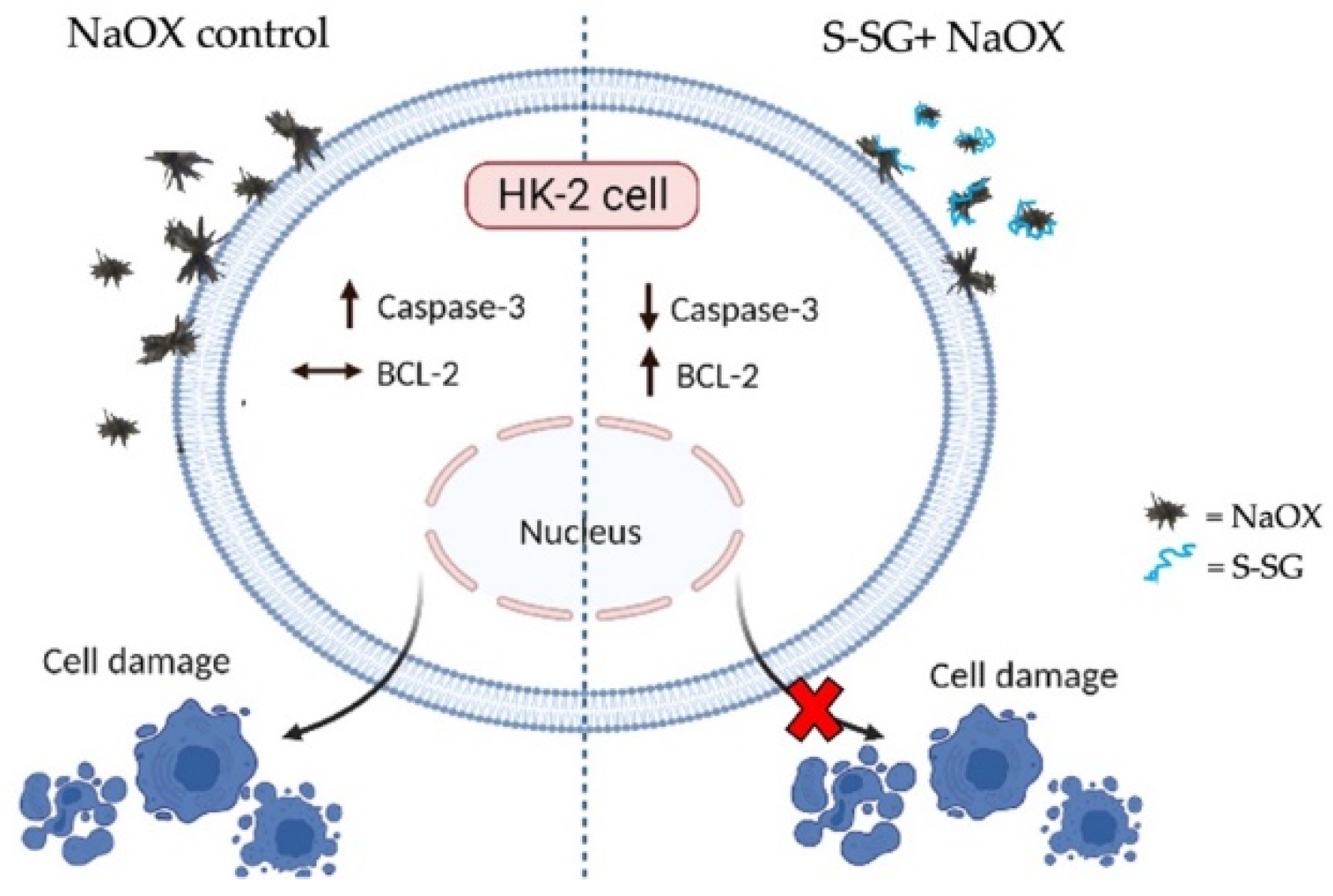
2.6. Protective Effect of S-SG against NaOX-Induced HK-2 Cell Damage
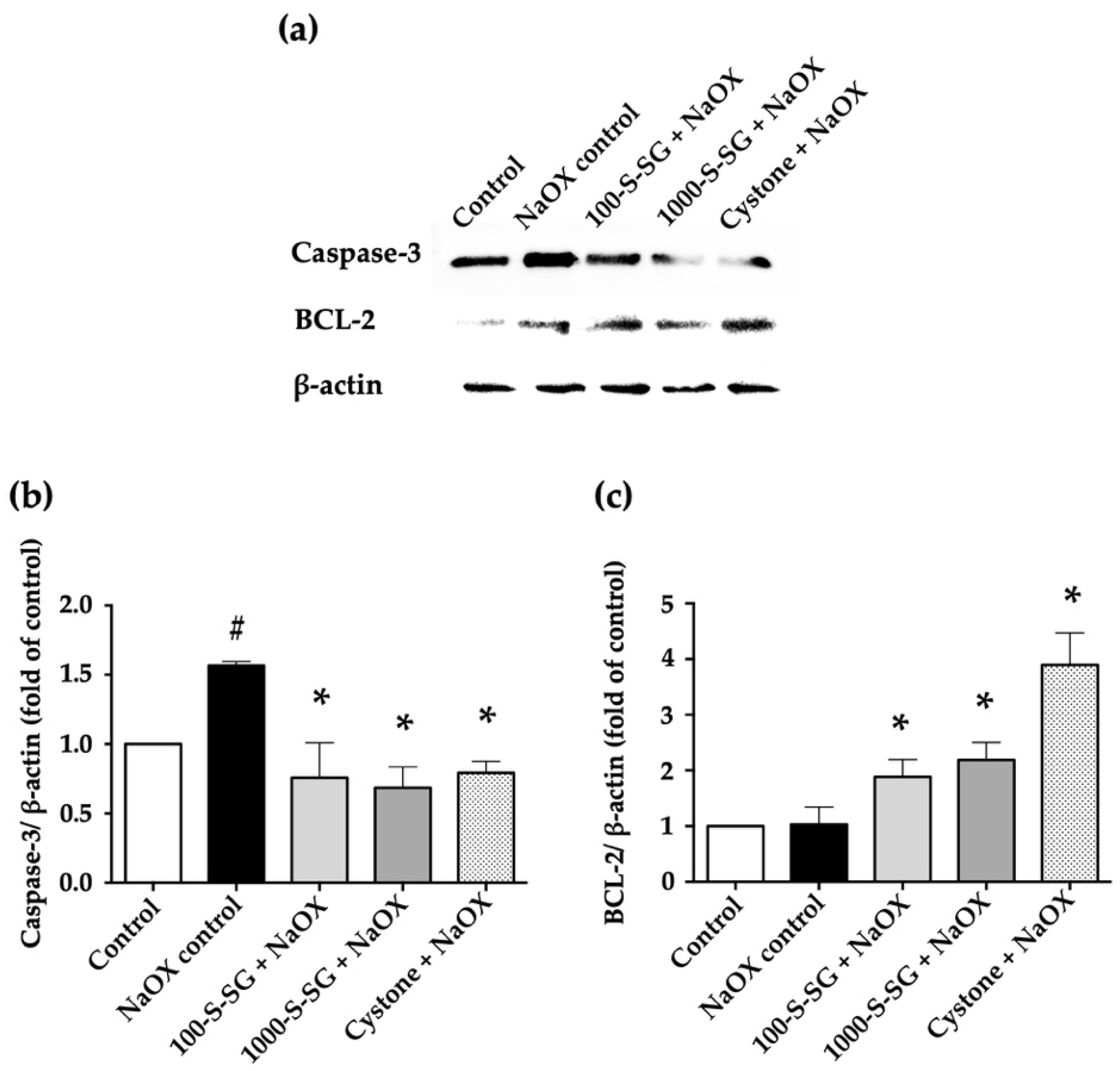
2.7. Expression of Apoptosis-Related Proteins
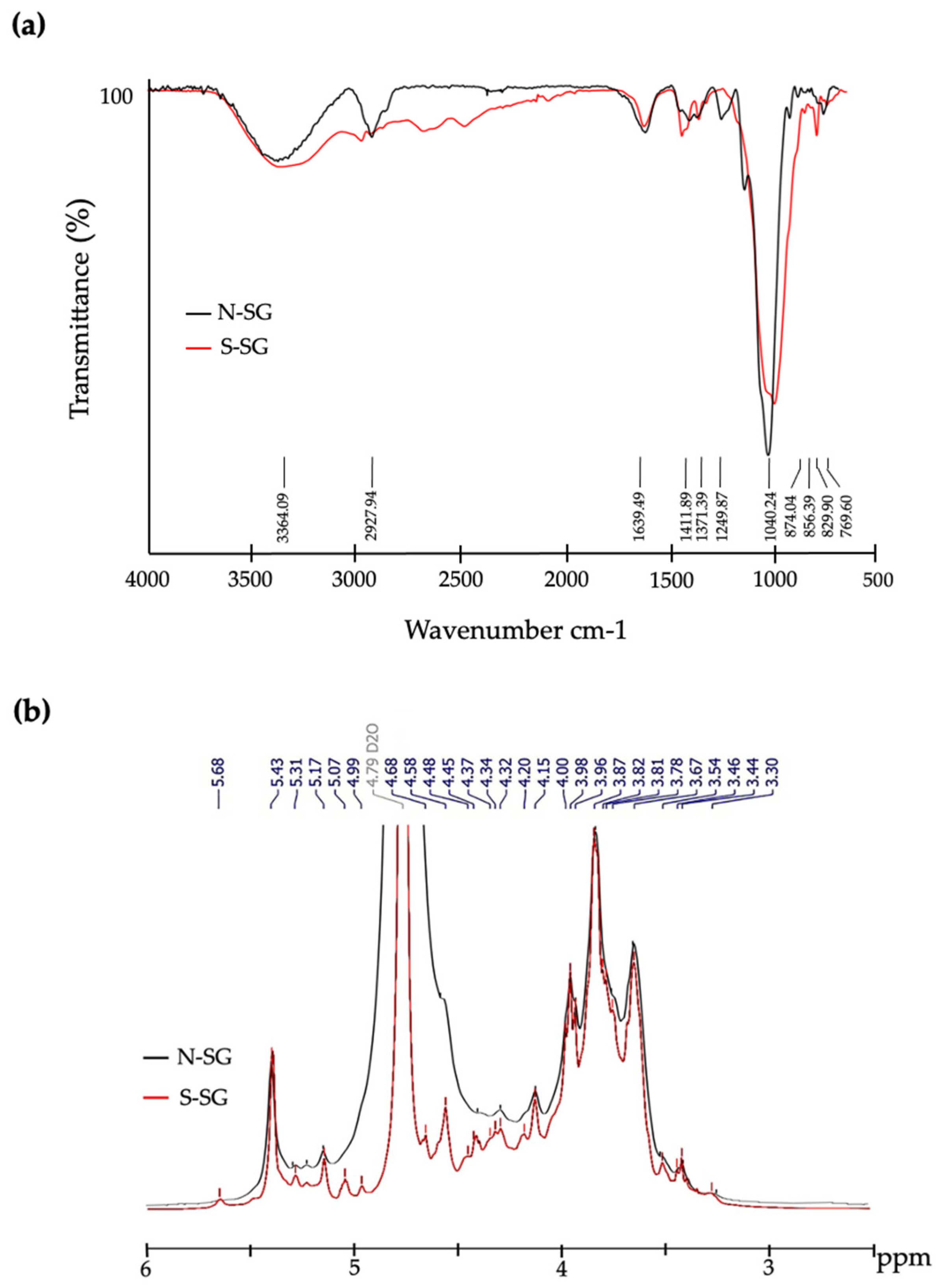
2.8. Structural Characterization of S-SG
3. Discussion
4. Materials and Methods
4.1. Materials and Chemicals
4.2. Preparation of Sulfated Galactans (N-SG) and Their Derivatives (L-SG and S-SG)
4.2.1. Native Sulfated Galactans (N-SG) from G. fisheri
4.2.2. Lower Molecular Weight SG (L-SG)
4.2.3. Sulfation SG (S-SG)
4.3. Evaluation of Antioxidant Activity
4.3.1. DPPH Radical Scavenging Activity Assay
4.3.2. Superoxide Radical Scavenging Activity Assay (O2−)
4.4. Investigation of Calcium Oxalate Crystallization
4.4.1. Nucleation Assay
4.4.2. Aggregation Assay
4.5. Crystal Morphology
4.6. Protective Effect of S-SG against HK-2 Cell Damage Induced by NaOX
4.6.1. Cytotoxicity Detection of Sodium Oxalate (NaOX) and S-SG on HK-2 Cells
4.6.2. Effect of S-SG on NaOX-Damaged HK-2 Cells
4.7. Determination of Apoptotosis-Related Protein Expression by Western Blot Analysis
4.8. Structural Characterization of S-SG
4.8.1. Sulfate Estimation Analysis
4.8.2. Average Molecular Weight Analysis
4.8.3. Fourier Transform Infrared (FTIR) Spectroscopy Analysis
4.8.4. Nuclear Magnetic Resonance (NMR) Spectroscopy Analysis
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gottlieb, M.; Long, B.; Koyfman, A. The evaluation and management of urolithiasis in the ED: A review of the literature. Am. J. Emerg. Med. 2018, 36, 699–706. [Google Scholar] [CrossRef] [PubMed]
- Arunkajohnsak, N.; Taweemonkongsap, T.; Leewansangtong, S.; Srinualnad, S.; Jongjitaree, K.; Chotikawanich, E. The correlation between demographic factors and upper urinary tract stone composition in the Thai population. Heliyon 2020, 6, e04649. [Google Scholar] [CrossRef] [PubMed]
- Worcester, E.M.; Coe, F.L. Clinical practice calcium kidney stones. NEJM 2010, 363, 954–963. [Google Scholar] [CrossRef]
- Ratkalkar, V.N.; Kleinman, J.G. Mechanisms of stone formation. Clin. Rev. Bone Miner. Metab. 2011, 9, 187–197. [Google Scholar] [CrossRef] [Green Version]
- De Araújo, L.; Costa-Pessoa, J.M.; de Ponte, M.C.; Oliveira-Souza, M. Sodium oxalate-induced acute kidney injury associated with glomerular and tubulointerstitial damage in rats. Front. Physiol. 2020, 11, 1076. [Google Scholar] [CrossRef] [PubMed]
- Bashir, S.; Gilani, A.H. Antiurolithic effect of Bergenia ligulata rhizome: An explanation of the underlying mechanisms. J. Ethnopharmacol. 2009, 122, 106–116. [Google Scholar] [CrossRef]
- De Bellis, R.; Piacentini, M.P.; Meli, M.A.; Mattioli, M.; Menotta, M.; Mari, M.; Valentini, L.; Palomba, L.; Desideri, D.; Chiarantini, L. In vitro effects on calcium oxalate crystallization kinetics and crystal morphology of an aqueous extract from Ceterach officinarum: Analysis of a potential antilithiatic mechanism. PLoS ONE 2019, 14, e0218734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, C.H.; Chen, Y.C.; Chen, L.D.; Pan, T.C.; Ho, C.Y.; Lai, M.T.; Tsai, F.J.; Chen, W.C. A traditional Chinese herbal antilithic formula, Wulingsan, effectively prevents the renal deposition of calcium oxalate crystal in ethylene glycol-fed rats. Urol. Res. 2008, 36, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Gomes, D.L.; Melo, K.R.T.; Queiroz, M.F.; Batista, L.A.N.C.; Santos, P.C.; Costa, M.S.S.P.; Almeida-Lima, J.; Gomes, R.B.C.; Costa, L.S.; Rocha, H.A.O. In vitro studies reveal antiurolithic effect of antioxidant sulfated polysaccharides from the green seaweed Caulerpa cupressoides var flabellata. Mar. Drugs 2019, 17, 326. [Google Scholar] [CrossRef] [Green Version]
- Veena, C.K.; Josephine, A.; Preetha, S.P.; Varalakshmi, P.; Sundarapandiyan, R. Renal peroxidative changes mediated by oxalate: The protective role of fucoidan. Life Sci. 2006, 79, 1789–1795. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.M.; Sun, X.Y.; Ouyang, J.M. Structural characterization, antioxidant activity, and biomedical application of astragalus polysaccharide degradation products. Int. J. Polym. Sci. 2018, 30, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Chen, R.Z.; Tan, L.; Jin, C.G.; Lu, J.; Tian, L.; Chang, Q.Q.; Wang, K. Extraction, isolation, characterization and antioxidant activity of polysaccharides from Astragalus membranaceus. Ind. Crops. Prod. 2015, 77, 434–443. [Google Scholar] [CrossRef]
- Sun, L.; Chu, J.; Sun, Z.; Chen, L. Physicochemical properties, immunomodulation and antitumor activities of polysaccharide from Pavlova viridis. Life Sci. 2016, 144, 156–161. [Google Scholar] [CrossRef] [PubMed]
- Qiong, W.; Cheng, Z.; Zheng-Xiang, N.; Bao, Y. Modification of low molecular weight polysaccharides from Tremella fuciformis and Their antioxidant activity in vitro. Int. J. Mol. Sci. 2007, 8, 670–679. [Google Scholar]
- Li, S.; Xiong, Q.; Lai, X.; Li, X.; Wan, M.; Zhang, J.; Yan, Y.; Cao, M.; Lu, L.; Guan, J.; et al. Molecular modification of polysaccharides and resulting bioactivities. Compr. Rev. Food Sci. Food Saf. 2016, 15, 237–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, H.; Fu, X.; Cao, C.; Li, C.; Chen, C.; Huang, Q. Sulfated modification, characterization, antioxidant and hypoglycemic activities of polysaccharides from Sargassum pallidum. Int. J. Biol. Macromol. 2019, 121, 407–414. [Google Scholar] [CrossRef] [PubMed]
- Wongprasert, K.; Rudtanatip, T.; Praiboon, J. Immunostimulatory activity of sulfated galactans isolated from the red seaweed Gracilaria fisheri and development of resistance against white spot syndrome virus (WSSV) in shrimp. Fish Shellfish. Immunol. 2014, 36, 52–60. [Google Scholar] [CrossRef]
- Rudtanatip, T.; Pariwatthanakun, C.; Somintara, S.; Sakaew, W.; Wongprasert, K. Structural characterization, antioxidant activity, and protective effect against hydrogen peroxide-induced oxidative stress of chemically degraded Gracilaria fisheri sulfated galactans. Int. J. Biol. Macromol. 2022, 206, 51–63. [Google Scholar] [CrossRef] [PubMed]
- Souza, B.W.S.; Cerqueira, M.A.; Bourbon, A.I.; Pinheiro, A.C.; Martins, J.T.; Teixeira, J.A.; Coimbra, M.A.; Vicente, A.A. Chemical characterization and antioxidant activity of sulfated polysaccharide from the red seaweed Gracilaria birdiae. Food Hydrocoll. 2012, 27, 287–292. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Ordóñez, E.; Rupérez, P. FTIR-ATR spectroscopy as a tool for polysaccharide identification in edible brown and red seaweeds. Food Hydrocoll. 2011, 25, 1514–1520. [Google Scholar] [CrossRef]
- Maciel, J.S.; Chaves, L.S.; Souza, B.W.S.; Teixeira, D.I.A.; Freitas, A.L.P.; Feitosa, J.P.A.; de Paula, R.C.M. Structural characterization of cold extracted fraction of soluble sulfated polysaccharide from red seaweed Gracilaria birdiae. Carbohyd. Polym. 2008, 71, 559–565. [Google Scholar] [CrossRef]
- Farias, W.R.L.; Valente, A.P.; Pereira, M.S.; Mourão, P.A.S. Structure and anticoagulant activity of galactans: Isolation of a unique sulfated galactan from the red algae Botryocladia occidentalis and comparison of its anticoagulant action with that of sulfated galactans from invertebrates. J. Biol. Chem. 2000, 275, 29299–29307. [Google Scholar] [CrossRef] [Green Version]
- Zhao, G.; Zhai, X.; Qu, M.; Tong, C.; Li, W. Sulfated modification of the polysaccharides from Crassostrea gigas and their antioxidant and hepatoprotective activities through metabolomics analysis. Int. J. Biol. Macromol. 2019, 129, 386–395. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Yu, P.; Li, Z.; Zhang, H.; Xu, Z. Antioxidant activities of sulfated polysaccharide fractions from Porphyra haitanesis. J. Appl. Phycol. 2003, 15, 305–310. [Google Scholar] [CrossRef]
- Gunasekaran, S.; Govindan, S.; Ramani, P. Sulfated modification, characterization and bioactivities of an acidic polysaccharide fraction from an edible mushroom Pleurotus eous (Berk.) Sacc. Heliyon 2021, 7, e05964. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.T.; Sun, X.Y.; Yu, K.; Gui, B.S.; Gui, Q.; Ouyang, J.M. Effect of content of sulfate groups in seaweed polysaccharides on antioxidant activity and repair effect of subcellular organelles in injured HK-2 cells. Oxid. Med. Cell. Longev. 2017, 2017, 2542950. [Google Scholar] [CrossRef]
- Oliveira, L.C.B.P.; Queiroz, M.F.; Fidelis, G.P.; Melo, K.R.T.; Câmara, R.B.G.; Alves, M.G.C.F.; Costa, L.S.; Teixeira, D.I.A.; Melo-Silveira, R.F.; Rocha, H.A.O. Antioxidant Sulfated Polysaccharide from Edible Red Seaweed Gracilaria birdiae is an Inhibitor of Calcium Oxalate Crystal Formation. Molecules 2020, 25, 2055. [Google Scholar] [CrossRef] [PubMed]
- Grohe, B.; O’Young, J.; Langdon, A.; Karttunen, M.; Goldberg, H.A.; Hunter, G.K. Citrate modulates calcium oxalate crystal growth by face-specific interactions. Cells Tissues Organs 2011, 194, 176–181. [Google Scholar] [CrossRef]
- Zhao, X.; Guo, F.; Hu, J.; Zhang, L.; Xue, C.; Zhang, Z.; Li, B. Antithrombotic activity of oral administered low molecular weight fucoidan from Laminaria Japonica. Thromb. Res. 2016, 144, 46–52. [Google Scholar] [CrossRef]
- Han, J.; Guo, D.; Sun, X.Y.; Wang, J.M.; Ouyang, J.M.; Gui, B.S. Comparison of the adhesion and endocytosis of calcium oxalate dihydrate to HK-2 cells before and after repair by Astragalus polysaccharide. Sci. Technol. Adv. Mater. 2019, 20, 1164–1177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ouyang, J.M.; Zheng, H.; Deng, S.P. Simultaneous formation of calcium oxalate (mono-, di-, and trihydrate) induced by potassium tartrate in gelatinous system. J. Cryst. Growth 2006, 293, 118–123. [Google Scholar] [CrossRef]
- Wesson, J.A.; Worcester, E.M.; Wiessner, J.H.; Mandel, N.S.; Kleinman, J.G. Control of calcium oxalate crystal structure and cell adherence by urinary macromolecules. Kidney Int. 1998, 53, 952–957. [Google Scholar] [CrossRef] [PubMed]
- He, J.Y.; Deng, S.P.; Ouyang, J.M. Morphology, particle size distribution, aggregation, and crystal phase of nanocrystallites in the urine of healthy persons and lithogenic patients. IEEE Trans. Nanobioscience 2010, 9, 156–163. [Google Scholar] [PubMed]
- Duan, C.Y.; Xia, Z.Y.; Zhang, G.N.; Gui, B.S.; Xue, J.F.; Ouyang, J.M. Changes in urinary nanocrystallites in calcium oxalate stone formers before and after potassium citrate intake. Int. J. Nanomed. 2013, 8, 909–918. [Google Scholar]
- Teodosio Melo, K.R.; Gomes Camara, R.B.; Queiroz, M.F.; Jacome Vidal, A.A.; Machado Lima, C.R.; Melo-Silveira, R.F.; Almeida-Lima, J.; Oliveira Rocha, H.A. Evaluation of sulfated polysaccharides from the brown seaweed Dictyopteris Justii as antioxidant agents and as inhibitors of the formation of calcium oxalate crystals. Molecules 2013, 18, 14543–14563. [Google Scholar] [CrossRef] [Green Version]
- Ouyang, J.M.; Wang, M.; Lu, P.; Tan, J. Degradation of sulfated polysaccharide extracted from algal Laminaria japonica and its modulation on calcium oxalate crystallization. Mater. Sci. Eng. C 2010, 30, 1022–1029. [Google Scholar] [CrossRef]
- Guo, D.; Yu, K.; Sun, X.Y.; Ouyang, J.M. Structural characterization and repair mechanism of Gracilaria lemaneiformis sulfated polysaccharides of different molecular weights on damaged renal epithelial cells. Oxidative Med. Cell. Longev. 2018, 2018, 7410389. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Wang, J.Q.; Nie, S.P.; Wang, Y.X.; Cui, S.W.; Xie, M.Y. Sulfated modification, characterization and property of a water-insoluble polysaccharide from Ganoderma atrum. Int. J. Biol. Macromol. 2015, 79, 248–255. [Google Scholar] [CrossRef]
- Wang, J.; Guo, H.; Zhang, J.; Wang, X.; Zhao, B.; Yao, J.; Wang, Y. Sulfated modification, characterization and structure–antioxidant relationships of Artemisia sphaerocephala polysaccharides. Carbohydr. Polym. 2010, 81, 897–905. [Google Scholar] [CrossRef]
- Lieske, J.C.; Leonard, R.; Toback, F.G. Adhesion of calcium oxalate monohydrate crystals to renal epithelial cells is inhibited by specific anions. Am. J. Physiol. Cell Physiol. 1995, 268, F604–F612. [Google Scholar] [CrossRef]
- Zhao, Y.W.; Guo, D.; Li, C.Y.; Ouyang, J.M. Comparison of the adhesion of calcium oxalate monohydrate to HK-2 cells before and after repair using tea polysaccharides. Int. J. Nanomed. 2019, 14, 4277–4292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mikita, P.; Vidhush, Y.; Oreoluwa, A.; Vikram, S.; Dean, G.A.; Ross, P.H.; Tanecia, M. Oxalate induces mitochondrial dysfunction and disrupts redox homeostasis in a human monocyte derived cell line. Redox Biol. 2018, 15, 207–215. [Google Scholar]
- Verzola, D.; Bertolotto, M.B.; Villaggio, B.; Ottonello, L.; Dallegri, F.; Frumento, G.; Berruti, V.; Gandolfo, M.T.; Garibotto, G.; Deferran, G. Taurine prevents apoptosis induced by high ambient glucose in human tubule renal cells. JIM 2002, 50, 443–451. [Google Scholar]
- Sarıca, K.; Aydin, H.; Yencilek, F.; Telci, D.; Yilmaz, B. Human umbilical vein endothelial cells accelerate oxalate-induced apoptosis of human renal proximal tubule epithelial cells in co-culture system which is prevented by pyrrolidine dithiocarbamate. Urol. Res. 2012, 40, 461–466. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.C.; Honeyman, W.T.; Cooney, R.; Kennington, L.; Scheid, R.C.; Jonassen, A.J. Mitochondrial dysfunction is a primary event in renal cell oxalate toxicity. Kidney Int. 2004, 66, 1890–1900. [Google Scholar] [CrossRef] [Green Version]
- Jeong, B.C.; Kwak, C.; Cho, K.S.; Kim, B.S.; Hong, S.K.; Kim, J.I.; Lee, C.; Kim, H.H. Apoptosis induced by oxalate in human renal tubular epithelial HK-2 cells. Urol. Res. 2005, 33, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Tandon, S.; Tanzeer Kaur, T.; Tandon, C. In vitro studies on calcium oxalate induced apoptosis attenuated by Didymocarpus pedicellata. Biointerface Res. Appl. Chem. 2022, 12, 7342–7355. [Google Scholar]
- Verkoelen, C.F.; Romijn, J.C.; Cao, C.; Schroder, F.H. Crystal-cell interaction inhibition by polysaccharides. Urol. J. 1996, 155, 749–752. [Google Scholar] [CrossRef] [Green Version]
- Qi, H.; Zhao, T.; Zhang, Q.; Li, Z.; Zhao, Z.; Xing, R. Antioxidant activity of different molecular weight sulfated polysaccharides from Ulva pertusa Kjellm (Chlorophyta). J. Appl. Phycol. 2005, 17, 527–534. [Google Scholar] [CrossRef]
- Patel, P.K.; Patel, M.A.; Vyas, B.A.; Shah, D.R.; Gandhi, T.R. Antiurolithiatic activity of saponin rich fraction from the fruits of Solanum xanthocarpum Schrad. & Wendl. (Solanaceae) against ethylene glycol induced urolithiasis in rats. J. Ethnopharmacol. 2012, 144, 160–170. [Google Scholar] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diameter of Crystals (µm) | Crystallizations (%) | ||
|---|---|---|---|
COM  | COD  | ||
| Control | 84.35 ± 3.16 | 99.10 ± 0.85 | 0.90 ± 0.84 |
| N-SG | 68.58 ± 6.70 * | 96.77 ± 1.46 | 3.23 ± 1.46 |
| L-SG | 79.93 ± 3.02 * | 95.00 ± 2.38 * | 5.00 ± 1.25 * |
| S-SG | 65.59 ± 6.71 * | 95.50 ± 0.21 * | 4.50 ± 0.22 * |
| Cystone | 26.50 ± 5.00 * | 87.92 ± 3.46 * | 12.08 ± 3.29 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sakaew, W.; Phanphak, J.; Somintara, S.; Hipkaeo, W.; Wongprasert, K.; Kovensky, J.; Pariwatthanakun, C.; Rudtanatip, T. Increased Sulfation in Gracilaria fisheri Sulfated Galactans Enhances Antioxidant and Antiurolithiatic Activities and Protects HK-2 Cell Death Induced by Sodium Oxalate. Mar. Drugs 2022, 20, 382. https://doi.org/10.3390/md20060382
Sakaew W, Phanphak J, Somintara S, Hipkaeo W, Wongprasert K, Kovensky J, Pariwatthanakun C, Rudtanatip T. Increased Sulfation in Gracilaria fisheri Sulfated Galactans Enhances Antioxidant and Antiurolithiatic Activities and Protects HK-2 Cell Death Induced by Sodium Oxalate. Marine Drugs. 2022; 20(6):382. https://doi.org/10.3390/md20060382
Chicago/Turabian StyleSakaew, Waraporn, Jenjiralai Phanphak, Somsuda Somintara, Wiphawi Hipkaeo, Kanokpan Wongprasert, José Kovensky, Choowadee Pariwatthanakun, and Tawut Rudtanatip. 2022. "Increased Sulfation in Gracilaria fisheri Sulfated Galactans Enhances Antioxidant and Antiurolithiatic Activities and Protects HK-2 Cell Death Induced by Sodium Oxalate" Marine Drugs 20, no. 6: 382. https://doi.org/10.3390/md20060382