
A Survey on the Distribution of Ovothiol and ovoA Gene Expression in Different Tissues and Cells: A Comparative Analysis in Sea Urchins and Mussels

 ,
,  ,
,  , , , and
, , , and
Abstract
:1. Introduction
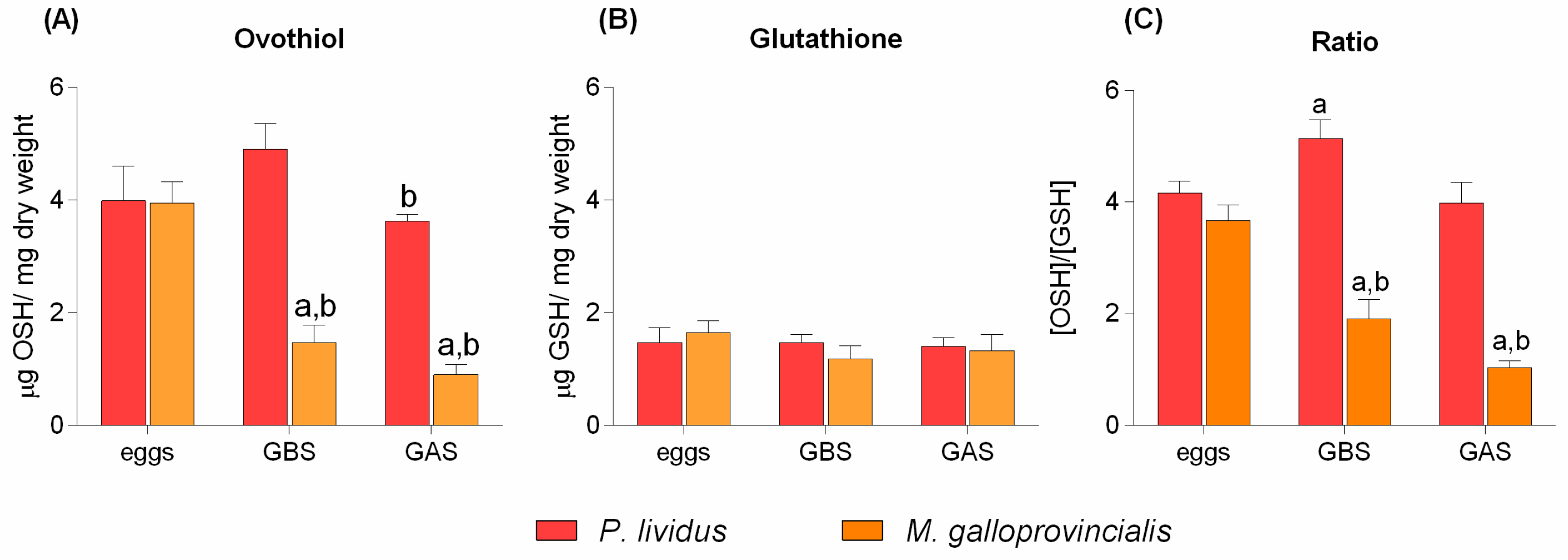
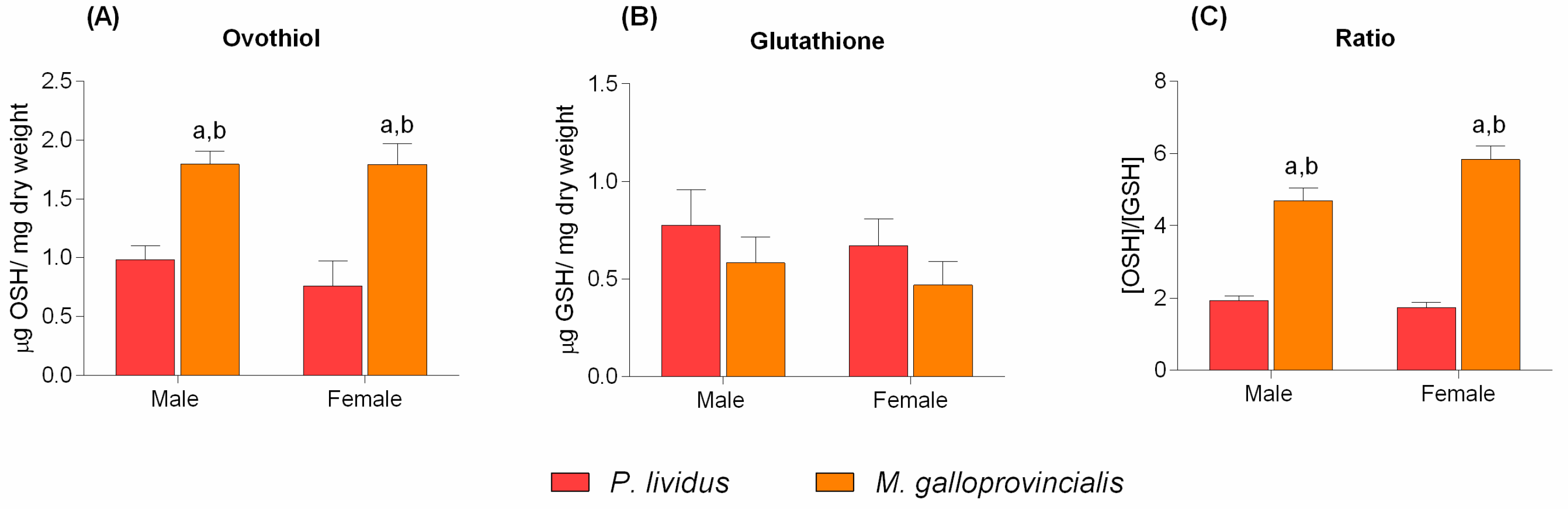
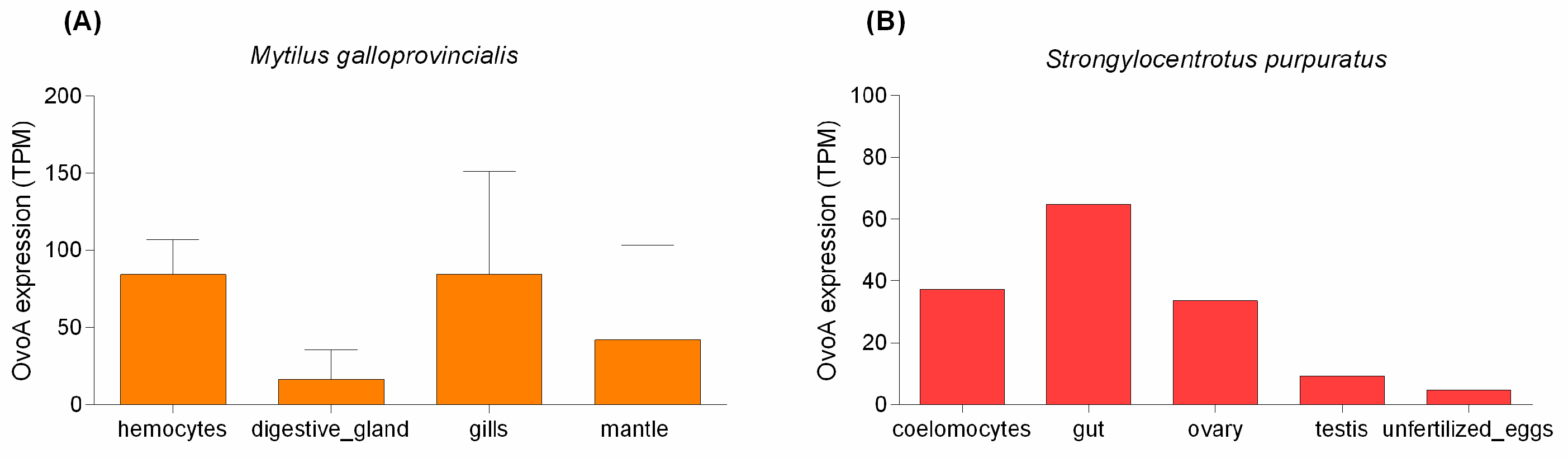
2. Results
3. Discussion
4. Materials and Method
4.1. Animal Collection
4.2. Tissue and Cells Sampling
4.3. Ovothiol Determination
4.4. Histological Analysis
4.5. In Silico Analysis of ovoA Expression in Sea Urchin and Mussel Tissues and Cells
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Blunt, J.W.; Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine Natural Products. Nat. Prod. Rep. 2018, 35, 8–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine Natural Products. Nat. Prod. Rep. 2020, 37, 175–223. [Google Scholar] [CrossRef] [PubMed]
- Castellano, I.; Seebeck, F.P. On Ovothiol Biosynthesis and Biological Roles: From Life in the Ocean to Therapeutic Potential. Nat. Prod. Rep. 2018, 35, 1241–1250. [Google Scholar] [CrossRef] [Green Version]
- Palumbo, A.; Castellano, I.; Napolitano, A. Ovothiol: A Potent Natural Antioxidant from Marine Organisms. In Blue Biotechnology; Wiley-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2018; pp. 583–610. ISBN 978-3-527-80171-8. [Google Scholar]
- Osik, N.A.; Zelentsova, E.A.; Tsentalovich, Y.P. Kinetic Studies of Antioxidant Properties of Ovothiol A. Antioxidants 2021, 10, 1470. [Google Scholar] [CrossRef] [PubMed]
- Marjanovic, B.; Simic, M.G.; Jovanovic, S.V. Heterocyclic Thiols as Antioxidants: Why Ovothiol C Is a Better Antioxidant than Ergothioneine. Free Radic. Biol. Med. 1995, 18, 679–685. [Google Scholar] [CrossRef]
- Shapiro, B.M. The Control of Oxidant Stress at Fertilization. Science 1991, 252, 533–536. [Google Scholar] [CrossRef]
- Castellano, I.; Migliaccio, O.; D’Aniello, S.; Merlino, A.; Napolitano, A.; Palumbo, A. Shedding Light on Ovothiol Biosynthesis in Marine Metazoans. Sci. Rep. 2016, 6, 21506. [Google Scholar] [CrossRef] [Green Version]
- Yanshole, V.V.; Yanshole, L.V.; Zelentsova, E.A.; Tsentalovich, Y.P. Ovothiol A Is the Main Antioxidant in Fish Lens. Metabolites 2019, 9, 95. [Google Scholar] [CrossRef] [Green Version]
- Diaz de Cerio, O.; Reina, L.; Squatrito, V.; Etxebarria, N.; Gonzalez-Gaya, B.; Cancio, I. Gametogenesis-Related Fluctuations in Ovothiol Levels in the Mantle of Mussels from Different Estuaries: Fighting Oxidative Stress for Spawning in Polluted Waters. Biomolecules 2020, 10, 373. [Google Scholar] [CrossRef] [Green Version]
- Milito, A.; Castellano, I.; Burn, R.; Seebeck, F.P.; Brunet, C.; Palumbo, A. First Evidence of Ovothiol Biosynthesis in Marine Diatoms. Free. Radic. Biol. Med. 2020, 152, 680–688. [Google Scholar] [CrossRef]
- Milito, A.; Orefice, I.; Smerilli, A.; Castellano, I.; Napolitano, A.; Brunet, C.; Palumbo, A. Insights into the Light Response of Skeletonema Marinoi: Involvement of Ovothiol. Marine Drugs 2020, 18, 477. [Google Scholar] [CrossRef] [PubMed]
- Ariyanayagam, M.R.; Fairlamb, A.H. Ovothiol and Trypanothione as Antioxidants in Trypanosomatids. Mol. Biochem. Parasitol. 2001, 115, 189–198. [Google Scholar] [CrossRef]
- Röhl, I.; Schneider, B.; Schmidt, B.; Zeeck, E. ʟ-Ovothiol A: The Egg Release Pheromone of the Marine Polychaete Platynereis Dumerilii: Annelida: Polychaeta. Z. Für Nat. C 1999, 54, 1145–1174. [Google Scholar] [CrossRef]
- Breithaupt, T.; Hardege, J.D. Pheromones mediating sex and dominance in aquatic animals in Chemical ecology in aquatic systems. In Chemical Ecology in Aquatic Systems; Brönmark, C., Hansson, L.-A., Eds.; Oxford University Press: London, UK, 2012; ISBN 978-0-19-958309-6. [Google Scholar]
- Torres, J.P.; Lin, Z.; Watkins, M.; Salcedo, P.F.; Baskin, R.P.; Elhabian, S.; Safavi-Hemami, H.; Taylor, D.; Tun, J.; Concepcion, G.P.; et al. Small-Molecule Mimicry Hunting Strategy in the Imperial Cone Snail Conus Imperialis. Sci. Adv. 2021, 7, eabf2704. [Google Scholar] [CrossRef]
- Milito, A.; Cocurullo, M.; Columbro, A.; Nonnis, S.; Tedeschi, G.; Castellano, I.; Arnone, M.I.; Palumbo, A. Ovothiol Ensures the Correct Developmental Programme of the Sea Urchin Paracentrotus Lividus Embryo. Open Biol. 2022, 12, 210262. [Google Scholar] [CrossRef] [PubMed]
- Russo, G.; Russo, M.; Castellano, I.; Napolitano, A.; Palumbo, A. Ovothiol Isolated from Sea Urchin Oocytes Induces Autophagy in the Hep-G2 Cell Line. Mar. Drugs 2014, 12, 4069–4085. [Google Scholar] [CrossRef] [Green Version]
- Brancaccio, M.; Russo, M.; Masullo, M.; Palumbo, A.; Russo, G.L.; Castellano, I. Sulfur-Containing Histidine Compounds Inhibit γ-Glutamyl Transpeptidase Activity in Human Cancer Cells. J. Biol. Chem. 2019, 294, 14603–14614. [Google Scholar] [CrossRef]
- Milito, A.; Brancaccio, M.; Lisurek, M.; Masullo, M.; Palumbo, A.; Castellano, I. Probing the Interactions of Sulfur-Containing Histidine Compounds with Human Gamma-Glutamyl Transpeptidase. Mar. Drugs 2019, 17, 650. [Google Scholar] [CrossRef] [Green Version]
- Brancaccio, M.; D’Argenio, G.; Lembo, V.; Palumbo, A.; Castellano, I. Antifibrotic Effect of Marine Ovothiol in an In Vivo Model of Liver Fibrosis. Oxidative Med. Cell. Longev. 2018, 2018, 5045734. [Google Scholar] [CrossRef] [Green Version]
- Castellano, I.; Di Tomo, P.; Di Pietro, N.; Mandatori, D.; Pipino, C.; Formoso, G.; Napolitano, A.; Palumbo, A.; Pandolfi, A. Anti-Inflammatory Activity of Marine Ovothiol A in an In Vitro Model of Endothelial Dysfunction Induced by Hyperglycemia. Oxidative Med. Cell. Longev. 2018, 2018, 2087373. [Google Scholar] [CrossRef] [Green Version]
- Milito, A.; Brancaccio, M.; D’Argenio, G.; Castellano, I. Natural Sulfur-Containing Compounds: An Alternative Therapeutic Strategy against Liver Fibrosis. Cells 2019, 8, 1356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braunshausen, A.; Seebeck, F.P. Identification and Characterization of the First Ovothiol Biosynthetic Enzyme. J. Am. Chem. Soc. 2011, 133, 1757–1759. [Google Scholar] [CrossRef] [PubMed]
- Naowarojna, N.; Huang, P.; Cai, Y.; Song, H.; Wu, L.; Cheng, R.; Li, Y.; Wang, S.; Lyu, H.; Zhang, L.; et al. In Vitro Reconstitution of the Remaining Steps in Ovothiol A Biosynthesis: C–S Lyase and Methyltransferase Reactions. Org. Lett. 2018, 20, 5427–5430. [Google Scholar] [CrossRef] [PubMed]
- Gerdol, M.; Sollitto, M.; Pallavicini, A.; Castellano, I. The Complex Evolutionary History of Sulfoxide Synthase in Ovothiol Biosynthesis. Proc. R. Soc. B. 2019, 286, 20191812. [Google Scholar] [CrossRef] [PubMed]
- Brancaccio, M.; Tangherlini, M.; Danovaro, R.; Castellano, I. Metabolic Adaptations to Marine Environments: Molecular Diversity and Evolution of Ovothiol Biosynthesis in Bacteria. Genome Biol. Evol. 2021, 13, evab169. [Google Scholar] [CrossRef] [PubMed]
- Palumbo, A.; Misuraca, G.; d’Ischia, M.; Donaudy, F.; Prota, G. Isolation and Distribution of 1-Methyl-5-Thiol-l-Histidine Disulphide and a Related Metabolite in Eggs from Echinoderms. Comp. Biochem. Physiol. Part B Comp. Biochem. 1984, 78, 81–83. [Google Scholar] [CrossRef]
- Turner, E.; Klevit, R.; Hopkins, P.B.; Shapiro, B.M. Ovothiol: A Novel Thiohistidine Compound from Sea Urchin Eggs That Confers NAD(P)H-O2 Oxidoreductase Activity on Ovoperoxidase. J. Biol. Chem. 1986, 261, 13056–13063. [Google Scholar] [CrossRef]
- Byrne, M. Annual Reproductive Cycles of the Commercial Sea Urchin Paracentrotus Lividus from an Exposed Intertidal and a Sheltered Subtidal Habitat on the West Coast of Ireland. Mar. Biol. 1990, 104, 275–289. [Google Scholar] [CrossRef]
- Santos, P.M.; Albano, P.; Raposo, A.; Ferreira, S.M.F.; Costa, J.L.; Pombo, A. The Effect of Temperature on Somatic and Gonadal Development of the Sea Urchin Paracentrotus Lividus (Lamarck, 1816). Aquaculture 2020, 528, 735487. [Google Scholar] [CrossRef]
- Milito, A.; Murano, C.; Castellano, I.; Romano, G.; Palumbo, A. Antioxidant and Immune Response of the Sea Urchin Paracentrotus Lividus to Different Re-Suspension Patterns of Highly Polluted Marine Sediments. Mar. Environ. Res. 2020, 160, 104978. [Google Scholar] [CrossRef]
- Grundemann, D.; Harlfinger, S.; Golz, S.; Geerts, A.; Lazar, A.; Berkels, R.; Jung, N.; Rubbert, A.; Schomig, E. Discovery of the Ergothioneine Transporter. Proc. Natl. Acad. Sci. USA 2005, 102, 5256–5261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halliwell, B.; Cheach, I.K.; Tang, R.M.Y. Ergothioneine—A diet-derived antioxidant with therapeuticpotential. Febs Lett. 2018, 592, 3357–3366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shapiro, B.M.; Turner, E. Oxidative Stress and the Role of Novel Thiol Compounds at Fertilization. Biofactors 1988, 1, 85–88. [Google Scholar]
- Riley, J.C.M.; Behrman, H.R. Oxygen Radicals and Reactive Oxygen Species in Reproduction. Proc. Soc. Exp. Biol. Med. 1991, 198, 781–791. [Google Scholar] [CrossRef] [PubMed]
- Cianelli, D.; Uttieri, M.; Buonocore, B.; Falco, P.; Zambardino, G.; Zambianchi, E. Dynamics of a very special Mediterranean coastal area: The Gulf of Naples. In Mediterranean Ecosystems: Dynamics, Management & Conservation; William, G.S., Ed.; Nova Science Publishers: New York, NY, USA, 2012; ISBN 978-1-61209-146-4. [Google Scholar]
- Montuori, P.; Lama, P.; Aurino, S.; Naviglio, D.; Triassi, M. Metals Loads into the Mediterranean Sea: Estimate of Sarno River Inputs and Ecological Risk. Ecotoxicology 2013, 22, 295–307. [Google Scholar] [CrossRef] [Green Version]
- Tornero, V.; Ribera d’Alcalà, M. Contamination by Hazardous Substances in the Gulf of Naples and Nearby Coastal Areas: A Review of Sources, Environmental Levels and Potential Impacts in the MSFD Perspective. Sci. Total Environ. 2014, 466–467, 820–840. [Google Scholar] [CrossRef]
- Fasano, E.; Arnese, A.; Esposito, F.; Albano, L.; Masucci, A.; Capelli, C.; Cirillo, T.; Nardone, A. Evaluation of the impact of anthropogenic activities on arsenic, cadmium, chromium, mercury, lead, and polycyclic aromatic hydrocarbon levels in seafood from the Gulf of Naples, Italy. J. Environ. Sci. Health Part A 2018, 53, 786–792. [Google Scholar] [CrossRef]
- Perugini, M.; Visciano, P.; Manera, M.; Turno, G.; Lucisano, A.; Amorena, M. Polycyclic Aromatic Hydrocarbons in Marine Organisms from the Gulf of Naples. Tyrrhenian Sea. J. Agric. Food Chem. 2007, 55, 2049–2054. [Google Scholar] [CrossRef]
- Mercogliano, R.; Santonicola, S.; De Felice, A.; Anastasio, A.; Murru, N.; Ferrante, M.C.; Cortesi, M.L. Occurrence and Distribution of Polycyclic Aromatic Hydrocarbons in Mussels from the Gulf of Naples, Tyrrhenian Sea, Italy. Mar. Pollut. Bull. 2016, 104, 386–390. [Google Scholar] [CrossRef]
- Murano, C.; Vaccari, L.; Casotti, R.; Corsi, I.; Palumbo, A. Occurrence of microfibres in wild specimens of adult sea urchin Paracentrotus lividus (Lamarck, 1816) from a coastal area of the central Mediterranean Sea. Mar. Pollut. Bull. 2022, 176, 113448. [Google Scholar] [CrossRef]
- Mitta, G.; Vandenbulcke, F.; Noel, T.; Romestand, B.; Beauvillain, J.C.; Salzet, M.; Roch, P. Differential Distribution and Defence Involvement of Antimicrobial Peptides in Mussel. J. Cell Sci. 2000, 113, 2759–2769. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Mendikute, A.; Elizondo, M.; Venier, P.; Cajaraville, M.P. Characterization of Mussel Gill Cells in Vivo and in Vitro. Cell Tissue Res. 2005, 321, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Smith, L.C.; Arizza, V.; Barela Hudgell, M.A.; Barone, G.; Bodnar, A.G.; Buckley, K.M.; Cunsolo, V.; Dheilly, N.M.; Franchi, N.; Fugmann, S.D.; et al. Echinodermata: The Complex Immune System in Echinoderms. In Advances in Comparative Immunology; Cooper, E.L., Ed.; Springer International Publishing: Cham, Switzerland, 2018; pp. 409–501. ISBN 978-3-319-76767-3. [Google Scholar]
- Canesi, L.; Ciacci, C.; Balbi, T. Invertebrate Models for Investigating the Impact of Nanomaterials on Innate Immunity: The Example of the Marine Mussel Mytilus spp. Curr. Bionanotechnol. 2016, 2, 77–83. [Google Scholar] [CrossRef]
- Buckley, K.M.; Rast, J.P. Immune activity at the gut epithelium in the larval sea urchin. Cell Tissue Res. 2019, 377, 469–474. [Google Scholar] [CrossRef] [PubMed]
- Murano, C.; Bergami, E.; Liberatori, G.; Palumbo, A.; Corsi, I. Interplay Between Nanoplastics and the Immune System of the Mediterranean Sea Urchin Paracentrotus Lividus. Front. Mar. Sci. 2021, 8, 647394. [Google Scholar] [CrossRef]
- Liberatori, G.; Grassi, G.; Guidi, P.; Bernardeschi, M.; Fiorati, A.; Scarcelli, V.; Genovese, M.; Faleri, C.; Protano, G.; Frenzilli, G.; et al. Effect-Based Approach to Assess Nanostructured Cellulose Sponge Removal Efficacy of Zinc Ions from Seawater to Prevent Ecological Risks. Nanomaterials 2020, 10, 1283. [Google Scholar] [CrossRef]
- Machado, I.; Moura, P.; Pereira, F.; Vasconcelos, P.; Gaspar, M.B. Reproductive Cycle of the Commercially Harvested Sea Urchin (Paracentrotus Lividus) along the Western Coast of Portugal. Invertebr. Biol. 2019, 138, 40–54. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An Ultra-Fast All-in-One FASTQ Preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast Gapped-Read Alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [Green Version]
- Kopylova, E.; Noé, L.; Touzet, H. SortMeRNA: Fast and Accurate Filtering of Ribosomal RNAs in Metatranscriptomic Data. Bioinformatics 2012, 28, 3211–3217. [Google Scholar] [CrossRef]
- MacManes, M.D. The Oyster River Protocol: A Multi-Assembler and Kmer Approach for de Novo Transcriptome Assembly. PeerJ 2018, 6, e5428. [Google Scholar] [CrossRef] [PubMed]
- Patro, R.; Duggal, G.; Love, M.I.; Irizarry, R.A.; Kingsford, C. Salmon Provides Fast and Bias-Aware Quantification of Transcript Expression. Nat. Methods 2017, 14, 417–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, P.; Binns, D.; Chang, H.-Y.; Fraser, M.; Li, W.; McAnulla, C.; McWilliam, H.; Maslen, J.; Mitchell, A.; Nuka, G.; et al. InterProScan 5: Genome-Scale Protein Function Classification. Bioinformatics 2014, 30, 1236–1240. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| μg Ovothiol/mg Dry Weight | ||
|---|---|---|
| Tissue | Female | Male |
| Digestive gland | 1.4 ± 0.04 | 1.0 ± 0.02 |
| Gills | 2.0 ± 0.05 | 2.5 ± 0.10 |
| Mantle | 2.3 ± 0.22 | 0.29 ± 0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Murano, C.; Zuccarotto, A.; Leone, S.; Sollitto, M.; Gerdol, M.; Castellano, I.; Palumbo, A. A Survey on the Distribution of Ovothiol and ovoA Gene Expression in Different Tissues and Cells: A Comparative Analysis in Sea Urchins and Mussels. Mar. Drugs 2022, 20, 268. https://doi.org/10.3390/md20040268
Murano C, Zuccarotto A, Leone S, Sollitto M, Gerdol M, Castellano I, Palumbo A. A Survey on the Distribution of Ovothiol and ovoA Gene Expression in Different Tissues and Cells: A Comparative Analysis in Sea Urchins and Mussels. Marine Drugs. 2022; 20(4):268. https://doi.org/10.3390/md20040268
Chicago/Turabian StyleMurano, Carola, Annalisa Zuccarotto, Serena Leone, Marco Sollitto, Marco Gerdol, Immacolata Castellano, and Anna Palumbo. 2022. "A Survey on the Distribution of Ovothiol and ovoA Gene Expression in Different Tissues and Cells: A Comparative Analysis in Sea Urchins and Mussels" Marine Drugs 20, no. 4: 268. https://doi.org/10.3390/md20040268