
A Comparative Study of the Anti-Obesity Effects of Dietary Sea Cucumber Saponins and Energy Restriction in Response to Weight Loss and Weight Regain in Mice
 and
and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Effects on Food Intake, Body Weight, and Visceral Weight Ratio
2.2. Effects on Adipose Mass, Morphology, and Mrna Expression of Fsp27 and PLIN1
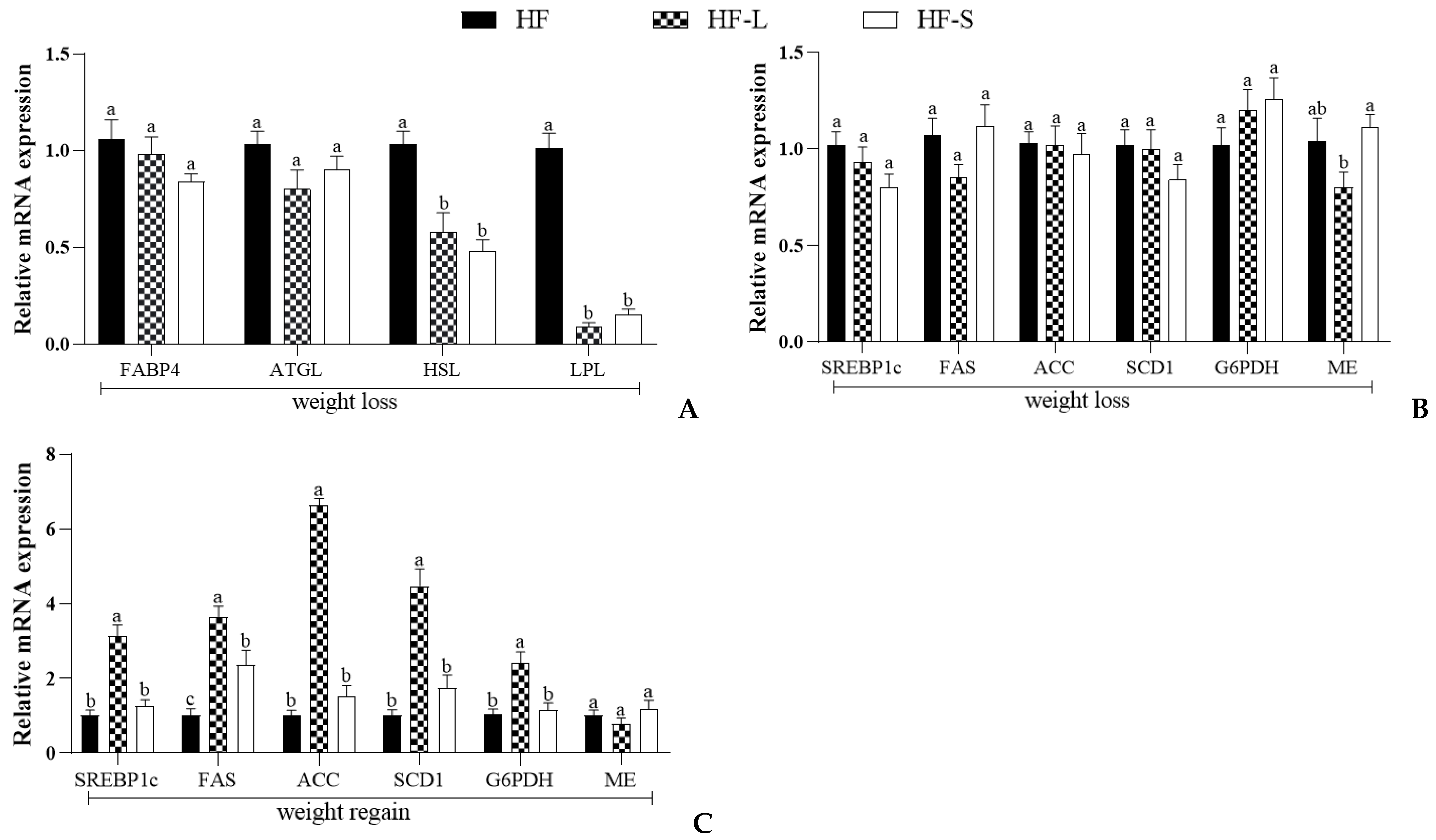
2.3. Effects on the Expression of Epididymal White Adipose Tissue Genes Involved in Lipid Metabolism
2.4. Effects on the Serum and Liver Lipid
2.5. Effects on Fatty Acid Composition of Liver Lipids
2.6. Effects on the Expression of Hepatic and Muscular Genes Involved in Lipid Metabolism
2.7. Effects on the OGTT and Serum Parameters
3. Materials and Methods
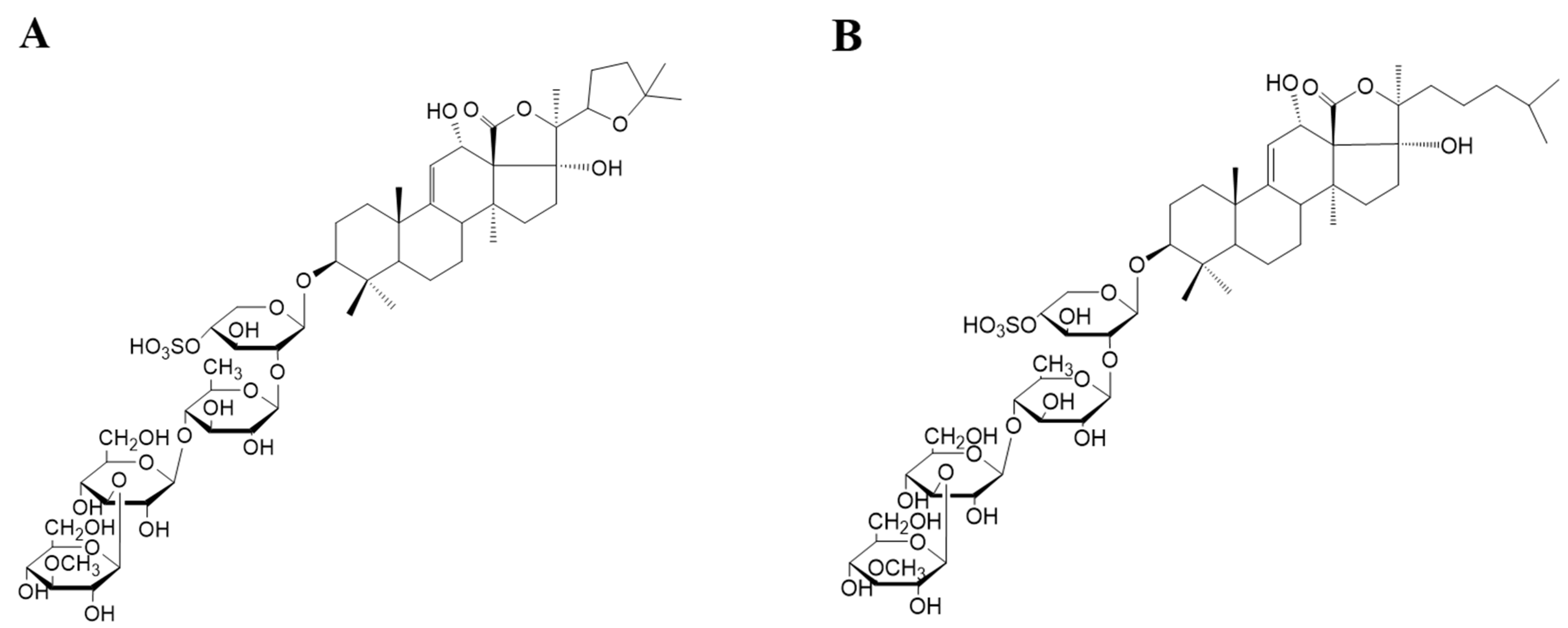
3.1. Preparation and Analysis of Sea Cucumber Saponins
3.2. Animal and Experimental Design
3.3. Oral Glucose Tolerance Test
3.4. Histological Analysis of Liver and Epididymal White Adiposes
3.5. Biochemical Analysis of Serum
3.6. Hepatic Lipid and Fatty Acid Composition Analysis
3.7. RNA Extraction and Real Time (RT)-PCR
3.8. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Conway, B.; Rene, A. Obesity as a disease: No lightweight matter. Obes. Rev. 2004, 5, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Martin-Rodriguez, E.; Guillen-Grima, F.; Marti, A.; Brugos-Larurnbe, A. Comorbidity associated with obesity in a large population: The APNA study. Obes. Res. Clin. Pract. 2015, 9, 435–447. [Google Scholar] [CrossRef] [PubMed]
- Swift, D.L.; McGee, J.E.; Earnest, C.P.; Carlisle, E.; Nygard, M.; Johannsen, N.M. The Effects of Exercise and Physical Activity on Weight Loss and Maintenance. Prog. Cardiovasc. Dis. 2018, 61, 206–213. [Google Scholar] [CrossRef] [PubMed]
- Guthold, R.; Stevens, G.A.; Riley, L.M.; Bull, F.C. Worldwide trends in insufficient physical activity from 2001 to 2016: A pooled analysis of 358 population-based surveys with 1.9 million participants. Lancet Glob. Health 2018, 6, E1077–E1086. [Google Scholar] [CrossRef] [Green Version]
- Guthold, R.; Stevens, G.A.; Riley, L.M.; Bull, F.C. Global trends in insufficient physical activity among adolescents: A pooled analysis of 298 population-based surveys with 1.6 million participants. Lancet Child. Adolesc. Health 2020, 4, 23–35. [Google Scholar] [CrossRef]
- MacLean, P.S.; Higgins, J.A.; Johnson, G.C.; Fleming-Elder, B.K.; Donahoo, W.T.; Melanson, E.L.; Hill, J.O. Enhanced metabolic efficiency contributes to weight regain after weight loss in obesity-prone rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2004, 287, R1306–R1315. [Google Scholar] [CrossRef]
- Yu, Y.H.; Deng, C.; Huang, X.F. Obese reversal by a chronic energy restricted diet leaves an increased Arc NPY/AgRP, but no alteration in POMC/CART, mRNA expression in diet-induced obese mice. Behav. Brain Res. 2009, 205, 50–56. [Google Scholar] [CrossRef]
- Zhao, Y.C.; Xue, C.H.; Zhang, T.T.; Wang, Y.M. Saponins from Sea Cucumber and Their Biological Activities. J. Agric. Food Chem. 2018, 66, 7222–7237. [Google Scholar] [CrossRef]
- Wang, Y.M.; Wang, J.H.; Yanagita, R.C.; Liu, C.H.; Hu, X.Q.; Dong, P.; Xue, C.H.; Xue, Y. Effects of two sulfated triterpene saponins echinoside A and holothurin A on the inhibition of dietary fat absorption and obesity reduction. Biosci. Biotechnol. Biochem. 2014, 78, 139–146. [Google Scholar] [CrossRef]
- Hu, X.Q.; Wang, Y.M.; Wang, J.F.; Xue, Y.; Li, Z.J.; Nagao, K.; Yanagita, T.; Xue, C.H. Dietary saponins of sea cucumber alleviate orotic acid-induced fatty liver in rats via PPAR alpha and SREBP-1c signaling. Lipids Health Dis. 2010, 9, 25. [Google Scholar] [CrossRef] [Green Version]
- Meng, J.; Hu, X.Q.; Zhang, T.T.; Dong, P.; Li, Z.J.; Xue, C.H.; Chang, Y.G.; Wang, Y.M. Saponin from sea cucumber exhibited more significant effects than ginsenoside on ameliorating high fat diet-induced obesity in C57BL/6 mice. Medchemcomm 2018, 9, 725–734. [Google Scholar] [CrossRef] [PubMed]
- Barbosa-da-Silva, S.; Fraulob-Aquino, J.C.; Lopes, J.R.; Mandarim-de-Lacerda, C.A.; Aguila, M.B. Weight Cycling Enhances Adipose Tissue Inflammatory Responses in Male Mice. PLoS ONE 2012, 7, e39837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puri, V.; Konda, S.; Ranjit, S.; Aouadi, M.; Chawla, A.; Chouinard, M.; Chakladar, A.; Czech, M.P. Fat-specific protein 27, a novel lipid droplet protein that enhances triglyceride storage. J. Biol. Chem. 2007, 282, 34213–34218. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Yu, S.; Chen, L.L.; Hu, X.; Zheng, J.; Deng, X.L. Involvement of PPAR/FSP27 in the pathogenic mechanism underlying insulin resistance: Tipping the balance between lipogenesis and fat storage in adult catch-up growth rats. Nutr. Metab. 2019, 16, 11. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.Q.; Gong, J.Y.; Wu, H.; Xu, W.Y.; Wu, L.Z.; Xu, D.J.; Gao, J.L.; Wu, J.W.; Yang, H.Y.; Yang, M.J.; et al. Perilipin1 promotes unilocular lipid droplet formation through the activation of Fsp27 in adipocytes. Nat. Commun. 2013, 4, 1594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.J.; Raza, S.H.A.; Zhao, C.P.; Cheng, G.; Zan, L.S. Overexpression of PLIN1 Promotes Lipid Metabolism in Bovine Adipocytes. Animals 2020, 10, 1944. [Google Scholar] [CrossRef]
- Chung, J.Y.; Hong, J.; Kim, H.J.; Song, Y.; Yong, S.B.; Lee, J.; Kim, Y.H. White adipocyte-targeted dual gene silencing of FABP4/5 for anti-obesity, anti-inflammation and reversal of insulin resistance: Efficacy and comparison of administration routes. Biomaterials 2021, 279, 121209. [Google Scholar] [CrossRef]
- Tian, J.; Wen, H.; Zeng, L.B.; Jiang, M.; Wu, F.; Liu, W.; Yang, C.G. Changes in the activities and mRNA expression levels of lipoprotein lipase (LPL), hormone-sensitive lipase (HSL) and fatty acid synthetase (FAS) of Nile tilapia (Oreochromis niloticus) during fasting and re-feeding. Aquaculture 2013, 400, 29–35. [Google Scholar] [CrossRef]
- Zimmermann, R.; Lass, A.; Haemmerle, G.; Zechner, R. Fate of fat: The role of adipose triglyceride lipase in lipolysis. Biochim. Biophys. Acta. Mol. Cell Biol. Lipids 2009, 1791, 494–500. [Google Scholar] [CrossRef]
- Fruhbeck, G.; Mendez-Gimenez, L.; Fernandez-Formoso, J.A.; Fernandez, S.; Rodriguez, A. Regulation of adipocyte lipolysis. Nutr. Res. Rev. 2014, 27, 63–93. [Google Scholar] [CrossRef] [Green Version]
- Taskinen, M.R.; Nikkila, E.A. Basal and postprandial lipoprotein lipase activity in adipose tissue during caloric restriction and refeeding. Metabolism 1987, 36, 625–630. [Google Scholar] [CrossRef]
- Li, Z.P.; Kim, H.J.; Park, M.S.; Ji, G.E. Effects of fermented ginseng root and ginseng berry on obesity and lipid metabolism in mice fed a high-fat diet. J. Ginseng. Res. 2018, 42, 312–319. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Meng, J.; Shi, H.H.; Wang, C.C.; Li, Z.J.; Xue, C.H.; Wang, Y.M.; Zhang, T.T. Dietary Supplementation with Sea Cucumber Saponins and Exercise Can Significantly Suppress Adipose Accumulation in Mice Fed with High-Fat Diet. J. Ocean Univ. China 2021, 20, 629–640. [Google Scholar] [CrossRef]
- Xu, J.; Han, J.; Long, Y.S.; Lock, J.; Weir, G.C.; Epstein, P.N.; Liu, Y.Q. Malic enzyme is present in mouse islets and modulates insulin secretion. Diabetologia 2008, 51, 2281–2289. [Google Scholar] [CrossRef] [Green Version]
- Mailloux, R.J.; Harper, M.E. Glucose regulates enzymatic sources of mitochondrial NADPH in skeletal muscle cells; a novel role for glucose-6-phosphate dehydrogenase. FASEB J. 2010, 24, 2495–2506. [Google Scholar] [CrossRef] [PubMed]
- Korczynska, J.; Stelmanska, E.; Swierczynski, J. Differential effect of long-term food restriction on fatty acid synthase and leptin gene expression in rat white adipose tissue. Horm. Metab. Res. 2003, 35, 593–597. [Google Scholar] [CrossRef] [PubMed]
- Stelmanska, E.; Korczynska, J.; Swierezynski, J. Tissue-specific effect of refeeding after short- and long-term caloric restriction on malic enzyme gene expression in rat tissues. Acta. Biochim. Pol. 2004, 51, 805–814. Available online: https://pubmed.ncbi.nlm.nih.gov/15448740/ (accessed on 8 December 2019). [CrossRef] [Green Version]
- Sun, Q.Q.; Nie, S.S.; Wang, L.X.; Yang, F.; Meng, Z.M.; Xiao, H.Y.; Xiang, B.; Li, X.J.; Fu, X.H.; Wang, S. Factors that Affect Pancreatic Islet Cell Autophagy in Adult Rats: Evaluation of a Calorie-Restricted Diet and a High-Fat Diet. PLoS ONE 2016, 11, e0151104. [Google Scholar] [CrossRef] [Green Version]
- Dallaire, M.P.; Taga, H.; Ma, L.; Corl, B.A.; Gervais, R.; Lebeuf, Y.; Richard, F.J.; Chouinard, P.Y. Effects of abomasal infusion of conjugated linoleic acids, Sterculia foetida oil, and fish oil on production performance and the extent of fatty acid Δ⁹-desaturation in dairy cows. J. Dairy. Sci. 2014, 97, 6411–6425. [Google Scholar] [CrossRef] [Green Version]
- Kasher-Meron, M.; Youn, D.Y.; Zong, H.H.; Pessin, J.E. Lipolysis defect in white adipose tissue and rapid weight regain. Am. J. Physiol. Endocrinol. Metab. 2019, 317, E185–E193. [Google Scholar] [CrossRef]
- Issemann, I.; Green, S. Activation of a member of the steroid-hormone receptor superfamily by peroxisome proliferators. Nature 1990, 347, 645–650. [Google Scholar] [CrossRef] [PubMed]
- Fan, C.Y.; Pan, J.; Chu, R.Y.; Lee, D.; Kluckman, K.D.; Usuda, N.; Singh, I.; Yeldandi, A.V.; Rao, M.S.; Maeda, N.; et al. Hepatocellular and hepatic peroxisomal alterations in mice with a disrupted peroxisomal fatty acyl-coenzyme A oxidase gene. J. Biol. Chem. 1996, 271, 24698–24710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonnefont, J.P.; Demaugre, F.; Prip-Buus, C.; Saudubray, J.M.; Brivet, M.; Abadi, N.; Thuillier, L. Carnitine palmitoyltransferase deficiencies. Mol. Genet. Metab. 1999, 68, 424–440. [Google Scholar] [CrossRef] [PubMed]
- Ding, S.B.; Jiang, J.J.; Zhang, G.F.; Bu, Y.J.; Zhang, G.H.; Zhao, X.M. Resveratrol and caloric restriction prevent hepatic steatosis by regulating SIRT1-autophagy pathway and alleviating endoplasmic reticulum stress in high-fat diet-fed rats. PLoS ONE 2017, 12, e0183541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sumiyoshi, M.; Kimura, Y. Effects of Eleutherococcus senticosus Cortex on Recovery from the Forced Swimming Test and Fatty Acid beta-Oxidation in the Liver and Skeletal Muscle of mice. Nat. Prod. J. 2016, 6, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Huh, J.Y.; Park, Y.J.; Ham, M.; Kim, J.B. Crosstalk between Adipocytes and Immune Cells in Adipose Tissue Inflammation and Metabolic Dysregulation in Obesity. Mol. Cells 2014, 37, 365–371. [Google Scholar] [CrossRef] [PubMed]
- Lumeng, C.N.; Saltiel, A.R. Inflammatory links between obesity and metabolic disease. J. Clin. Investig. 2011, 121, 2111–2117. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Kitade, H.; Ni, Y.; Ota, T. Roles of Chemokines and Chemokine Receptors in Obesity-Associated Insulin Resistance and Nonalcoholic Fatty Liver Disease. Biomolecules 2015, 5, 1563–1579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, X.Q.; Li, Z.J.; Xue, Y.; Xu, J.; Xue, C.H.; Wang, J.F.; Wang, Y.M. Dietary Saponins of Sea Cucumber Ameliorate Obesity, Hepatic Steatosis, and Glucose Intolerance in High-Fat Diet-Fed Mice. J. Med. Food 2012, 15, 909–916. [Google Scholar] [CrossRef] [PubMed]
- Anson, R.M.; Guo, Z.H.; de Cabo, R.; Iyun, T.; Rios, M.; Hagepanos, A.; Ingram, D.K.; Lane, M.A.; Mattson, M.P. Intermittent fasting dissociates beneficial effects of dietary restriction on glucose metabolism and neuronal resistance to injury from calorie intake. Proc. Natl. Acad. Sci. USA 2003, 100, 6216–6220. [Google Scholar] [CrossRef] [Green Version]
- Lee, A.H.; Dixit, V.D. Dietary Regulation of Immunity. Immunity 2020, 53, 510–523. [Google Scholar] [CrossRef]
- Kong, L.C.; Wuillemin, P.H.; Bastard, J.P.; Sokolovska, N.; Gougis, S.; Fellahi, S.; Darakhshan, F.; Bonnefont-Rousselot, D.; Bittar, R.; Dore, J.; et al. Insulin resistance and inflammation predict kinetic body weight changes in response to dietary weight loss and maintenance in overweight and obese subjects by using a Bayesian network approach. Am. J. Clin. Nutr. 2013, 98, 1385–1394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. Available online: https://pubmed.ncbi.nlm.nih.gov/13428781/ (accessed on 29 December 2019). [CrossRef]
- Liu, X.F.; Cui, J.; Leng, K.L.; Xue, C.H.; Li, Z.J.; Xue, Y.; Wang, Y.M. Docosahexaenoic acid-enriched phospholipids exhibit superior effects on obesity-related metabolic disorders to egg yolk phospholipids and soybean phospholipids in mice. Eur. J. Lipid Sci. Technol. 2016, 118, 1712–1721. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Weight Loss | Weight Regain | ||||
|---|---|---|---|---|---|---|
| HF | HF-L | HF-S | HF | HF-L | HF-S | |
| Food intake (g d−1) | 4.60 ± 0.12 a | 2.90 ± 0.00 b | 4.55 ± 0.15 a | 4.45 ± 0.18 a’ | 4.76 ± 0.22 a’ | 4.43 ± 0.18 a’ |
| Initial body weight (g) | 43.46 ± 0.94 a | 41.99 ± 0.83 a | 42.35 ± 0.84 a | 48.71 ± 2.18 a’ | 38.33 ± 1.05 b’ | 44.55 ± 0.84 a’ |
| Final body weight (g) | 46.58 ± 1.23 a | 38.49 ± 0.73 b | 43.86 ± 0.78 a | 49.76 ± 2.32 a’ | 45.80 ± 1.15 a’ | 47.72 ± 1.30 a’ |
| Body weight gain (g) | 3.14 ± 0.38 a | −3.24 ± 0.29 c | 1.29 ± 0.52 b | 1.38 ± 0.43 c’ | 8.37 ± 0.49 a’ | 3.60 ± 0.72 b’ |
| Liver (g per 100 g BW) | 3.34 ± 0.15 a | 3.48 ± 0.06 a | 3.32 ± 0.06 a | 3.46 ± 0.21 a’ | 3.44 ± 0.03 a’ | 3.45 ± 0.10 a’ |
| Heart (g per 100 g BW) | 0.40 ± 0.02 a | 0.46 ± 0.01 a | 0.44 ± 0.02 a | 0.41 ± 0.02 a’ | 0.44 ± 0.01 a’ | 0.43 ± 0.02 a’ |
| Kidneys (g per 100 g BW) | 1.36 ± 0.04 a | 1.37 ± 0.06 a | 1.38 ± 0.07 a | 1.49 ± 0.04 a’ | 1.44 ± 0.04 a’ | 1.47 ± 0.05 a’ |
| Spleen (g per 100 g BW) | 0.21 ± 0.02 a | 0.18 ± 0.01 a | 0.20 ± 0.01 a | 0.25 ± 0.04 a’ | 0.23 ± 0.02 a’ | 0.27 ± 0.04 a’ |
| Fatty Acid Composition% | Weight Loss | Weight Regain | ||||
|---|---|---|---|---|---|---|
| HF | HF-L | HF-S | HF | HF-L | HF-S | |
| C16:0 | 23.21 ± 1.8 a | 23.41 ± 3.28 a | 24.02 ± 2.64 a | 22.92 ± 2.52 a’ | 22.95 ± 2.98 a’ | 24.48 ± 2.69 a’ |
| C16:1 | 1.76 ± 0.11 a | 1.56 ± 0.06 a | 1.46 ± 0.06 a | 1.81 ± 0.13 a’ | 0.87 ± 0.08 b’ | 1.60 ± 0.10 ab’ |
| C18:0 | 13.42 ± 1.48 a | 12.62 ± 1.51 a | 13.53 ± 1.62 a | 11.26 ± 1.35 b’ | 16.02 ± 1.28 a’ | 13.15 ± 1.58 ab’ |
| C18:1 | 19.85 ± 0.79 a | 21.05 ± 1.89 a | 18.51 ± 2.22 a | 22.55 ± 2.93 a’ | 17.59 ± 1.23 b’ | 19.67 ± 2.36 ab’ |
| C18:2 | 19.14 ± 2.10 a | 20.51 ± 1.64 a | 18.61 ± 1.30 a | 21.37 ± 1.92 a’ | 17.62 ± 2.64 b’ | 18.68 ± 1.49 ab’ |
| C18:3 | 0.60 ± 0.02 a | 0.61 ± 0.04 a | 0.52 ± 0.02 a | 0.90 ± 0.05 a’ | 0.77 ± 0.05 a’ | 0.86 ± 0.11 a’ |
| C20:4 | 14.80 ± 1.33 a | 13.70 ± 1.78 a | 15.76 ± 1.42 a | 12.78 ± 1.79 b’ | 17.05 ± 1.88 a’ | 14.45 ± 1.73 ab’ |
| C22:2 | 1.67 ± 0.05 a | 1.13 ± 0.00 b | 1.46 ± 0.07 a | 1.32 ± 0.11 a’ | 1.29 ± 0.17 a’ | 1.19 ± 0.10 a’ |
| C22:6 | 5.55 ± 0.39 a | 5.41 ± 0.32 a | 6.13 ± 0.31 a | 5.10 ± 0.71 a’ | 5.84 ± 0.82 a’ | 5.92 ± 0.71 a’ |
| C16:1/C16:0 | 0.08 ± 0.00 a | 0.07 ± 0.00 a | 0.06 ± 0.00 b | 0.08 ± 0.00 a’ | 0.04 ± 0.00 b’ | 0.07 ± 0.01 a’ |
| C18:1/C18:0 | 1.48 ± 0.13 a | 1.67 ± 0.22 a | 1.37 ± 0.16 a | 2.00 ± 0.14 a’ | 1.10 ± 0.09 b’ | 1.50 ± 0.15 b’ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wen, L.; Li, R.; Zhao, Y.-C.; Yang, J.-Y.; Li, X.-Y.; Xue, C.-H.; Zhang, T.-T.; Wang, Y.-M. A Comparative Study of the Anti-Obesity Effects of Dietary Sea Cucumber Saponins and Energy Restriction in Response to Weight Loss and Weight Regain in Mice. Mar. Drugs 2022, 20, 629. https://doi.org/10.3390/md20100629
Wen L, Li R, Zhao Y-C, Yang J-Y, Li X-Y, Xue C-H, Zhang T-T, Wang Y-M. A Comparative Study of the Anti-Obesity Effects of Dietary Sea Cucumber Saponins and Energy Restriction in Response to Weight Loss and Weight Regain in Mice. Marine Drugs. 2022; 20(10):629. https://doi.org/10.3390/md20100629
Chicago/Turabian StyleWen, Lu, Rong Li, Ying-Cai Zhao, Jin-Yue Yang, Xiao-Yue Li, Chang-Hu Xue, Tian-Tian Zhang, and Yu-Ming Wang. 2022. "A Comparative Study of the Anti-Obesity Effects of Dietary Sea Cucumber Saponins and Energy Restriction in Response to Weight Loss and Weight Regain in Mice" Marine Drugs 20, no. 10: 629. https://doi.org/10.3390/md20100629