
A Preliminary Comparison of Plasma Tryptophan Metabolites and Medium- and Long-Chain Fatty Acids in Adult Patients with Major Depressive Disorder and Schizophrenia

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects and Plasma Sampling
2.2. Detection of Tryptophan and Its Catabolites
2.3. Detection of Medium and Long-Chain Fatty Acids
2.4. Statistical Analyses
3. Results
3.1. Clinical Characteristics of the Recruited Participants
3.2. Alternation of Trp and Its Catabolites in SCH and MDD
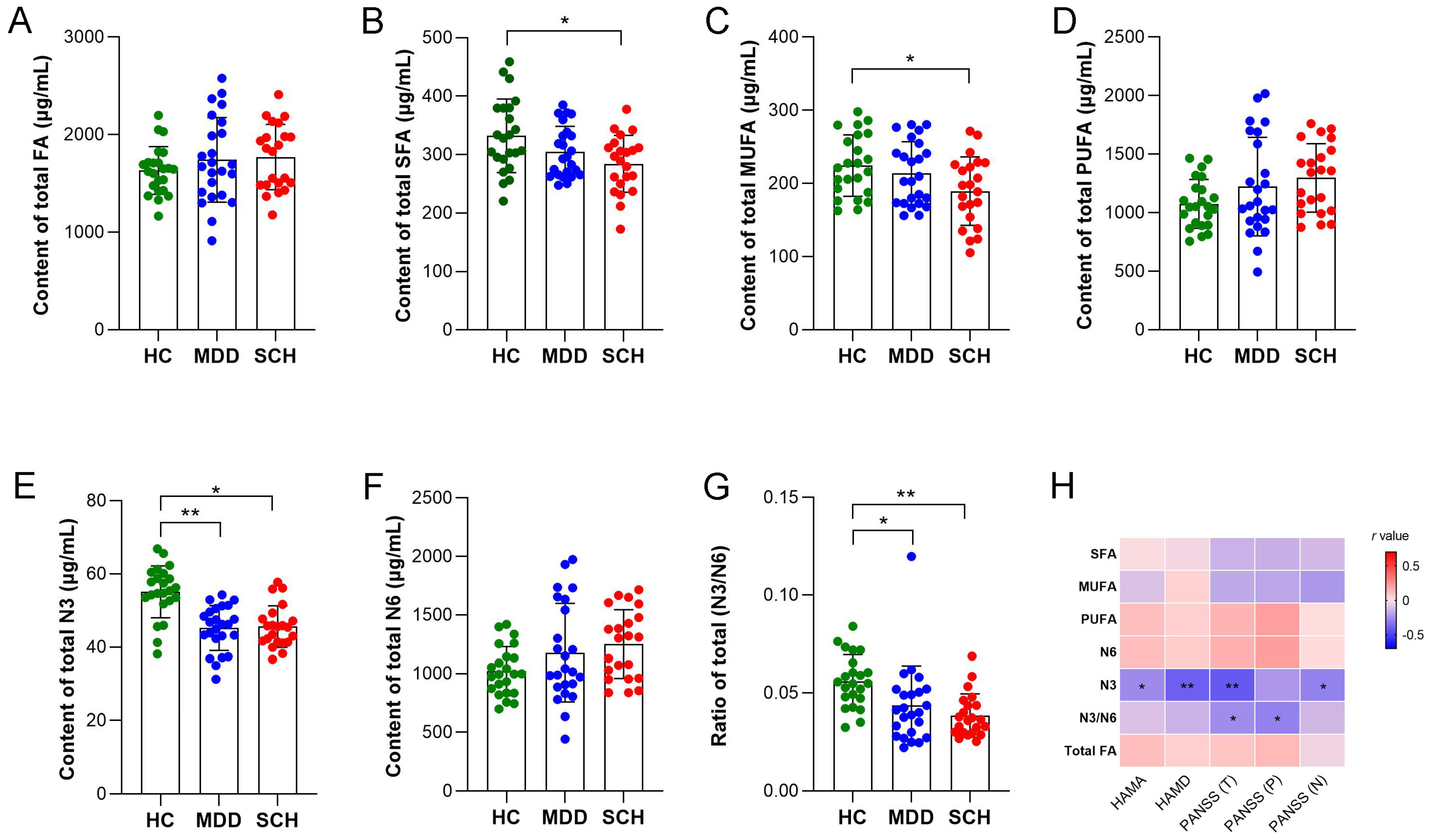
3.3. Alternation of MCFAs and LCFAs in SCH and MDD
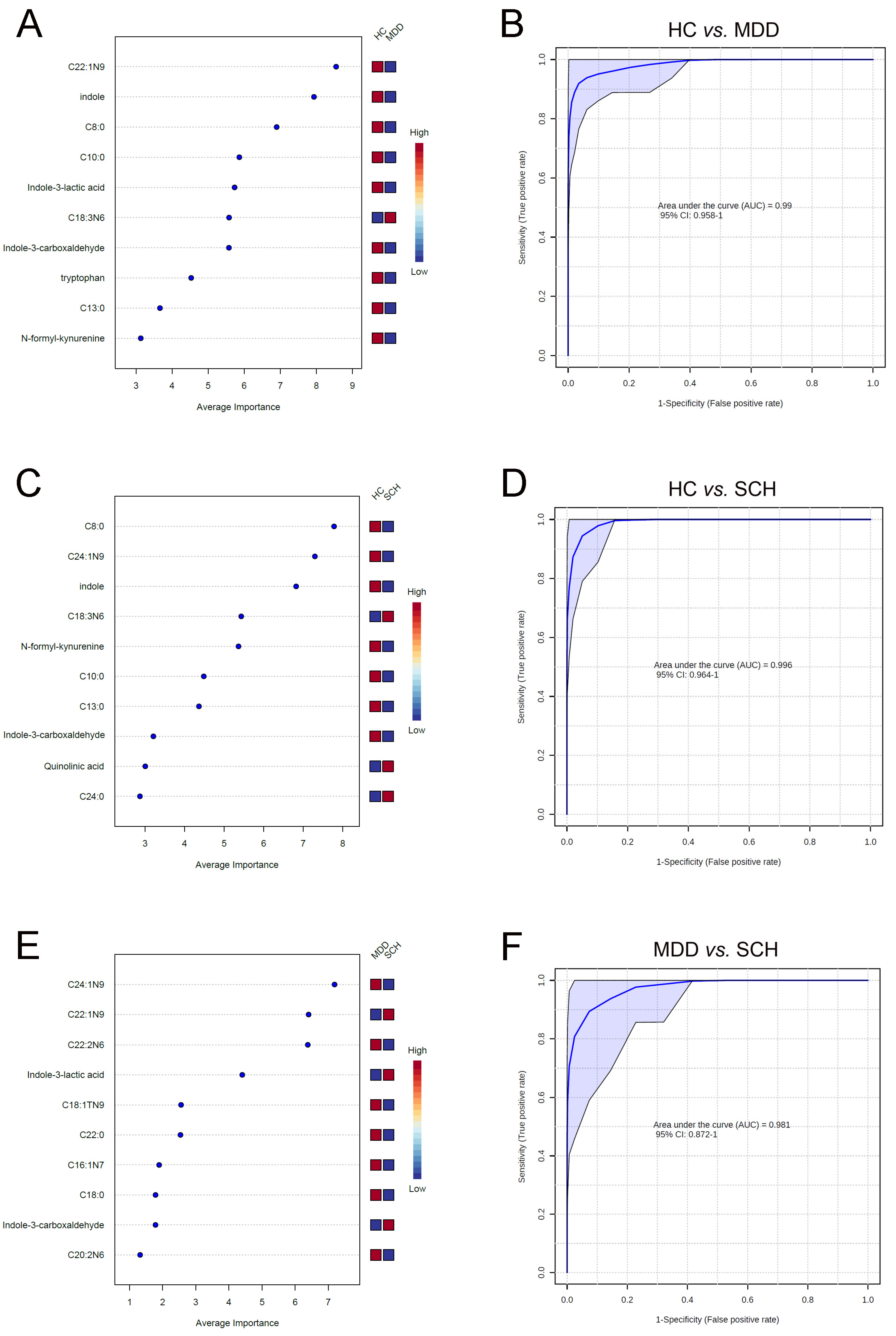
3.4. Characteristic Fatty Acids and Trp Catabolites in SCH and MDD
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Gur, S.; Weizman, S.; Stubbs, B.; Matalon, A.; Meyerovitch, J.; Hermesh, H.; Krivoy, A. Mortality, morbidity and medical resources utilization of patients with schizophrenia: A case-control community-based study. Psychiatry Res. 2018, 260, 177–181. [Google Scholar] [CrossRef] [PubMed]
- Rotenstein, L.S.; Ramos, M.A.; Torre, M.; Segal, J.B.; Peluso, M.J.; Guille, C.; Sen, S.; Mata, D.A. Prevalence of Depression, Depressive Symptoms, and Suicidal Ideation Among Medical Students: A Systematic Review and Meta-Analysis. JAMA 2016, 316, 2214–2236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosen, C.; Marvin, R.; Reilly, J.L.; Deleon, O.; Harris, M.S.; Keedy, S.K.; Solari, H.; Weiden, P.; Sweeney, J.A. Phenomenology of first-episode psychosis in schizophrenia, bipolar disorder, and unipolar depression: A comparative analysis. Clin. Schizophr. Relat. Psychoses 2012, 6, 145–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez-Aran, A.; Vieta, E. Cognition as a target in schizophrenia, bipolar disorder and depression. Eur. Neuropsychopharmacol. 2015, 25, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Li, D.; Chen, X. Characterizing the polygenic overlaps of bipolar disorder subtypes with schizophrenia and major depressive disorder. J. Affect. Disord. 2022, 309, 242–251. [Google Scholar] [CrossRef]
- Cannon, T.D.; Yu, C.; Addington, J.; Bearden, C.E.; Cadenhead, K.S.; Cornblatt, B.A.; Heinssen, R.; Jeffries, C.D.; Mathalon, D.H.; McGlashan, T.H.; et al. An Individualized Risk Calculator for Research in Prodromal Psychosis. Am. J. Psychiatry 2016, 173, 980–988. [Google Scholar] [CrossRef] [Green Version]
- Platten, M.; Nollen, E.A.A.; Rohrig, U.F.; Fallarino, F.; Opitz, C.A. Tryptophan metabolism as a common therapeutic target in cancer, neurodegeneration and beyond. Nat. Rev. Drug Discov. 2019, 18, 379–401. [Google Scholar] [CrossRef]
- Kanchanatawan, B.; Hemrungrojn, S.; Thika, S.; Sirivichayakul, S.; Ruxrungtham, K.; Carvalho, A.F.; Geffard, M.; Anderson, G.; Maes, M. Changes in Tryptophan Catabolite (TRYCAT) Pathway Patterning Are Associated with Mild Impairments in Declarative Memory in Schizophrenia and Deficits in Semantic and Episodic Memory Coupled with Increased False-Memory Creation in Deficit Schizophrenia. Mol. Neurobiol. 2018, 55, 5184–5201. [Google Scholar] [CrossRef]
- Milaneschi, Y.; Allers, K.A.; Beekman, A.T.F.; Giltay, E.J.; Keller, S.; Schoevers, R.A.; Sussmuth, S.D.; Niessen, H.G.; Penninx, B. The association between plasma tryptophan catabolites and depression: The role of symptom profiles and inflammation. Brain Behav. Immun. 2021, 97, 167–175. [Google Scholar] [CrossRef]
- Perica, M.M.; Delas, I. Essential fatty acids and psychiatric disorders. Nutr Clin. Pract 2011, 26, 409–425. [Google Scholar] [CrossRef]
- Weiser, M.J.; Butt, C.M.; Mohajeri, M.H. Docosahexaenoic Acid and Cognition throughout the Lifespan. Nutrients 2016, 8, 99. [Google Scholar] [CrossRef] [Green Version]
- Dimas, P.; Montani, L.; Pereira, J.A.; Moreno, D.; Trotzmuller, M.; Gerber, J.; Semenkovich, C.F.; Kofeler, H.C.; Suter, U. CNS myelination and remyelination depend on fatty acid synthesis by oligodendrocytes. eLife 2019, 8, e44702. [Google Scholar] [CrossRef]
- Goh, K.K.; Chen, C.Y.; Chen, C.H.; Lu, M.L. Effects of omega-3 polyunsaturated fatty acids supplements on psychopathology and metabolic parameters in schizophrenia: A meta-analysis of randomized controlled trials. J. Psychopharmacol. 2021, 35, 221–235. [Google Scholar] [CrossRef]
- Rapaport, M.H.; Nierenberg, A.A.; Schettler, P.J.; Kinkead, B.; Cardoos, A.; Walker, R.; Mischoulon, D. Inflammation as a predictive biomarker for response to omega-3 fatty acids in major depressive disorder: A proof-of-concept study. Mol. Psychiatry 2016, 21, 71–79. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Liu, L.; Lan, X.; Cohen, D.; Zhang, Y.; Ravindran, A.V.; Yuan, S.; Zheng, P.; Coghill, D.; Yang, L.; et al. Polyunsaturated fatty acids metabolism, purine metabolism and inosine as potential independent diagnostic biomarkers for major depressive disorder in children and adolescents. Mol. Psychiatry 2019, 24, 1478–1488. [Google Scholar] [CrossRef]
- Chiappelli, J.; Postolache, T.T.; Kochunov, P.; Rowland, L.M.; Wijtenburg, S.A.; Shukla, D.K.; Tagamets, M.; Du, X.; Savransky, A.; Lowry, C.A.; et al. Tryptophan Metabolism and White Matter Integrity in Schizophrenia. Neuropsychopharmacology 2016, 41, 2587–2595. [Google Scholar] [CrossRef] [Green Version]
- Davison, J.; O’Gorman, A.; Brennan, L.; Cotter, D.R. A systematic review of metabolite biomarkers of schizophrenia. Schizophr. Res. 2018, 195, 32–50. [Google Scholar] [CrossRef] [Green Version]
- Alkhalaf, L.M.; Ryan, K.S. Biosynthetic manipulation of tryptophan in bacteria: Pathways and mechanisms. Chem. Biol. 2015, 22, 317–328. [Google Scholar] [CrossRef] [Green Version]
- Stone, T.W.; Stoy, N.; Darlington, L.G. An expanding range of targets for kynurenine metabolites of tryptophan. Trends Pharmacol. Sci. 2013, 34, 136–143. [Google Scholar] [CrossRef] [Green Version]
- Yano, J.M.; Yu, K.; Donaldson, G.P.; Shastri, G.G.; Ann, P.; Ma, L.; Nagler, C.R.; Ismagilov, R.F.; Mazmanian, S.K.; Hsiao, E.Y. Indigenous bacteria from the gut microbiota regulate host serotonin biosynthesis. Cell 2015, 161, 264–276. [Google Scholar] [CrossRef] [Green Version]
- Beloborodova, N.V.; Chernevskaya, E.A.; Getsina, M.L. Indolic Structure Metabolites as Potential Biomarkers of Non-infectious Diseases. Curr. Pharm. Des. 2021, 27, 238–249. [Google Scholar] [CrossRef]
- Messaoud, A.; Mensi, R.; Douki, W.; Neffati, F.; Najjar, M.F.; Gobbi, G.; Valtorta, F.; Gaha, L.; Comai, S. Reduced peripheral availability of tryptophan and increased activation of the kynurenine pathway and cortisol correlate with major depression and suicide. World J. Biol. Psychiatry 2019, 20, 703–711. [Google Scholar] [CrossRef]
- Marx, W.; McGuinness, A.J.; Rocks, T.; Ruusunen, A.; Cleminson, J.; Walker, A.J.; Gomes-da-Costa, S.; Lane, M.; Sanches, M.; Diaz, A.P.; et al. The kynurenine pathway in major depressive disorder, bipolar disorder, and schizophrenia: A meta-analysis of 101 studies. Mol. Psychiatry 2021, 26, 4158–4178. [Google Scholar] [CrossRef]
- Zoga, M.; Oulis, P.; Chatzipanagiotou, S.; Masdrakis, V.G.; Pliatsika, P.; Boufidou, F.; Foteli, S.; Soldatos, C.R.; Nikolaou, C.; Papageorgiou, C. Indoleamine 2,3-dioxygenase and immune changes under antidepressive treatment in major depression in females. In Vivo 2014, 28, 633–638. [Google Scholar]
- Nemeth, H.; Toldi, J.; Vecsei, L. Role of kynurenines in the central and peripheral nervous systems. Curr. Neurovasc. Res. 2005, 2, 249–260. [Google Scholar] [CrossRef]
- Grant, R.S.; Coggan, S.E.; Smythe, G.A. The physiological action of picolinic Acid in the human brain. Int. J. Tryptophan Res. 2009, 2, 71–79. [Google Scholar] [CrossRef] [Green Version]
- Doolin, K.; Allers, K.A.; Pleiner, S.; Liesener, A.; Farrell, C.; Tozzi, L.; O’Hanlon, E.; Roddy, D.; Frodl, T.; Harkin, A.; et al. Altered tryptophan catabolite concentrations in major depressive disorder and associated changes in hippocampal subfield volumes. Psychoneuroendocrinology 2018, 95, 8–17. [Google Scholar] [CrossRef]
- Ozturk, M.; Yalin Sapmaz, S.; Kandemir, H.; Taneli, F.; Aydemir, O. The role of the kynurenine pathway and quinolinic acid in adolescent major depressive disorder. Int. J. Clin. Pract. 2021, 75, e13739. [Google Scholar] [CrossRef]
- Paul, E.R.; Schwieler, L.; Erhardt, S.; Boda, S.; Trepci, A.; Kampe, R.; Asratian, A.; Holm, L.; Yngve, A.; Dantzer, R.; et al. Peripheral and central kynurenine pathway abnormalities in major depression. Brain Behav. Immun. 2022, 101, 136–145. [Google Scholar] [CrossRef]
- Zhang, P.; Huang, J.; Gou, M.; Zhou, Y.; Tong, J.; Fan, F.; Cui, Y.; Luo, X.; Tan, S.; Wang, Z.; et al. Kynurenine metabolism and metabolic syndrome in patients with schizophrenia. J. Psychiatr. Res. 2021, 139, 54–61. [Google Scholar] [CrossRef]
- Almulla, A.F.; Vasupanrajit, A.; Tunvirachaisakul, C.; Al-Hakeim, H.K.; Solmi, M.; Verkerk, R.; Maes, M. The tryptophan catabolite or kynurenine pathway in schizophrenia: Meta-analysis reveals dissociations between central, serum, and plasma compartments. Mol. Psychiatry 2022, 27, 3679–3691. [Google Scholar] [CrossRef] [PubMed]
- Arnone, D.; Saraykar, S.; Salem, H.; Teixeira, A.L.; Dantzer, R.; Selvaraj, S. Role of Kynurenine pathway and its metabolites in mood disorders: A systematic review and meta-analysis of clinical studies. Neurosci. Biobehav. Rev. 2018, 92, 477–485. [Google Scholar] [CrossRef] [PubMed]
- Mitani, H.; Shirayama, Y.; Yamada, T.; Kawahara, R. Plasma levels of homovanillic acid, 5-hydroxyindoleacetic acid and cortisol, and serotonin turnover in depressed patients. Prog. Neuropsychopharmacol. Biol. Psychiatry 2006, 30, 531–534. [Google Scholar] [CrossRef] [PubMed]
- Moncrieff, J.; Cooper, R.E.; Stockmann, T.; Amendola, S.; Hengartner, M.P.; Horowitz, M.A. The serotonin theory of depression: A systematic umbrella review of the evidence. Mol. Psychiatry 2022. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, K.; Miura, I.; Kanno-Nozaki, K.; Horikoshi, S.; Mashiko, H.; Niwa, S.; Yabe, H. Associations between five-factor model of the Positive and Negative Syndrome Scale and plasma levels of monoamine metabolite in patients with schizophrenia. Psychiatry Res. 2015, 230, 419–423. [Google Scholar] [CrossRef]
- Roager, H.M.; Licht, T.R. Microbial tryptophan catabolites in health and disease. Nat. Commun. 2018, 9, 3294. [Google Scholar] [CrossRef] [Green Version]
- Zelante, T.; Puccetti, M.; Giovagnoli, S.; Romani, L. Regulation of host physiology and immunity by microbial indole-3-aldehyde. Curr. Opin. Immunol. 2021, 70, 27–32. [Google Scholar] [CrossRef]
- Delgado, I.; Cussotto, S.; Anesi, A.; Dexpert, S.; Aubert, A.; Aouizerate, B.; Beau, C.; Forestier, D.; Ledaguenel, P.; Magne, E.; et al. Association between the indole pathway of tryptophan metabolism and subclinical depressive symptoms in obesity: A preliminary study. Int. J. Obes. 2022, 46, 885–888. [Google Scholar] [CrossRef]
- Nimbkar, S.; Leena, M.M.; Moses, J.A.; Anandharamakrishnan, C. Medium chain triglycerides (MCT): State-of-the-art on chemistry, synthesis, health benefits and applications in food industry. Compr. Rev. Food Sci. Food Saf. 2022, 21, 843–867. [Google Scholar] [CrossRef]
- Schonfeld, P.; Wojtczak, L. Short- and medium-chain fatty acids in energy metabolism: The cellular perspective. J. Lipid Res. 2016, 57, 943–954. [Google Scholar] [CrossRef] [Green Version]
- Thevenet, J.; De Marchi, U.; Domingo, J.S.; Christinat, N.; Bultot, L.; Lefebvre, G.; Sakamoto, K.; Descombes, P.; Masoodi, M.; Wiederkehr, A. Medium-chain fatty acids inhibit mitochondrial metabolism in astrocytes promoting astrocyte-neuron lactate and ketone body shuttle systems. FASEB J. 2016, 30, 1913–1926. [Google Scholar] [CrossRef] [Green Version]
- Hollis, F.; Mitchell, E.S.; Canto, C.; Wang, D.; Sandi, C. Medium chain triglyceride diet reduces anxiety-like behaviors and enhances social competitiveness in rats. Neuropharmacology 2018, 138, 245–256. [Google Scholar] [CrossRef] [PubMed]
- Nafar, F.; Clarke, J.P.; Mearow, K.M. Coconut oil protects cortical neurons from amyloid beta toxicity by enhancing signaling of cell survival pathways. Neurochem. Int. 2017, 105, 64–79. [Google Scholar] [CrossRef]
- Andersen, J.V.; Westi, E.W.; Jakobsen, E.; Urruticoechea, N.; Borges, K.; Aldana, B.I. Astrocyte metabolism of the medium-chain fatty acids octanoic acid and decanoic acid promotes GABA synthesis in neurons via elevated glutamine supply. Mol. Brain 2021, 14, 132. [Google Scholar] [CrossRef]
- Hryhorczuk, C.; Florea, M.; Rodaros, D.; Poirier, I.; Daneault, C.; Des Rosiers, C.; Arvanitogiannis, A.; Alquier, T.; Fulton, S. Dampened Mesolimbic Dopamine Function and Signaling by Saturated but not Monounsaturated Dietary Lipids. Neuropsychopharmacology 2016, 41, 811–821. [Google Scholar] [CrossRef] [Green Version]
- Gupta, S.; Knight, A.G.; Gupta, S.; Keller, J.N.; Bruce-Keller, A.J. Saturated long-chain fatty acids activate inflammatory signaling in astrocytes. J. Neurochem. 2012, 120, 1060–1071. [Google Scholar] [CrossRef] [Green Version]
- Cone, J.J.; Chartoff, E.H.; Potter, D.N.; Ebner, S.R.; Roitman, M.F. Prolonged high fat diet reduces dopamine reuptake without altering DAT gene expression. PLoS ONE 2013, 8, e58251. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Fulton, S. Diet-induced obesity promotes depressive-like behaviour that is associated with neural adaptations in brain reward circuitry. Int. J. Obes. 2013, 37, 382–389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sartorius, T.; Ketterer, C.; Kullmann, S.; Balzer, M.; Rotermund, C.; Binder, S.; Hallschmid, M.; Machann, J.; Schick, F.; Somoza, V.; et al. Monounsaturated fatty acids prevent the aversive effects of obesity on locomotion, brain activity, and sleep behavior. Diabetes 2012, 61, 1669–1679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alemany, R.; Navarro, M.A.; Vogler, O.; Perona, J.S.; Osada, J.; Ruiz-Gutierrez, V. Olive oils modulate fatty acid content and signaling protein expression in apolipoprotein E knockout mice brain. Lipids 2010, 45, 53–61. [Google Scholar] [CrossRef]
- Wolfe, A.R.; Ogbonna, E.M.; Lim, S.; Li, Y.; Zhang, J. Dietary linoleic and oleic fatty acids in relation to severe depressed mood: 10 years follow-up of a national cohort. Prog. Neuropsychopharmacol. Biol. Psychiatry 2009, 33, 972–977. [Google Scholar] [CrossRef] [PubMed]
- Bazinet, R.P.; Laye, S. Polyunsaturated fatty acids and their metabolites in brain function and disease. Nat. Rev. Neurosci. 2014, 15, 771–785. [Google Scholar] [CrossRef] [PubMed]
- Lin, P.Y.; Huang, S.Y.; Su, K.P. A meta-analytic review of polyunsaturated fatty acid compositions in patients with depression. Biol. Psychiatry 2010, 68, 140–147. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, M.F.; Mutch, D.M.; Leri, F. The Relationship between Fatty Acids and Different Depression-Related Brain Regions, and Their Potential Role as Biomarkers of Response to Antidepressants. Nutrients 2017, 9, 298. [Google Scholar] [CrossRef] [Green Version]
- Hoen, W.P.; Lijmer, J.G.; Duran, M.; Wanders, R.J.; van Beveren, N.J.; de Haan, L. Red blood cell polyunsaturated fatty acids measured in red blood cells and schizophrenia: A meta-analysis. Psychiatry Res. 2013, 207, 1–12. [Google Scholar] [CrossRef]
- Jones, H.J.; Borges, M.C.; Carnegie, R.; Mongan, D.; Rogers, P.J.; Lewis, S.J.; Thompson, A.D.; Zammit, S. Associations between plasma fatty acid concentrations and schizophrenia: A two-sample Mendelian randomisation study. Lancet Psychiatry 2021, 8, 1062–1070. [Google Scholar] [CrossRef]
- Lin, P.Y.; Chang, C.H.; Chong, M.F.; Chen, H.; Su, K.P. Polyunsaturated Fatty Acids in Perinatal Depression: A Systematic Review and Meta-analysis. Biol. Psychiatry 2017, 82, 560–569. [Google Scholar] [CrossRef]
- Thesing, C.S.; Bot, M.; Milaneschi, Y.; Giltay, E.J.; Penninx, B. Bidirectional longitudinal associations of omega-3 polyunsaturated fatty acid plasma levels with depressive disorders. J. Psychiatr. Res. 2020, 124, 1–8. [Google Scholar] [CrossRef]
- Thesing, C.S.; Lok, A.; Milaneschi, Y.; Assies, J.; Bockting, C.L.H.; Figueroa, C.A.; Giltay, E.J.; Penninx, B.; Ruhe, H.G.; Schene, A.H.; et al. Fatty acids and recurrence of major depressive disorder: Combined analysis of two Dutch clinical cohorts. Acta Psychiatr. Scand. 2020, 141, 362–373. [Google Scholar] [CrossRef] [Green Version]
- Mongan, D.; Healy, C.; Jones, H.J.; Zammit, S.; Cannon, M.; Cotter, D.R. Plasma polyunsaturated fatty acids and mental disorders in adolescence and early adulthood: Cross-sectional and longitudinal associations in a general population cohort. Transl. Psychiatry 2021, 11, 321. [Google Scholar] [CrossRef]
- Robinson, D.G.; Gallego, J.A.; John, M.; Hanna, L.A.; Zhang, J.P.; Birnbaum, M.L.; Greenberg, J.; Naraine, M.; Peters, B.D.; McNamara, R.K.; et al. A potential role for adjunctive omega-3 polyunsaturated fatty acids for depression and anxiety symptoms in recent onset psychosis: Results from a 16 week randomized placebo-controlled trial for participants concurrently treated with risperidone. Schizophr. Res. 2019, 204, 295–303. [Google Scholar] [CrossRef]
- Hsu, M.C.; Tung, C.Y.; Chen, H.E. Omega-3 polyunsaturated fatty acid supplementation in prevention and treatment of maternal depression: Putative mechanism and recommendation. J. Affect. Disord. 2018, 238, 47–61. [Google Scholar] [CrossRef] [PubMed]
- Mocking, R.J.T.; Steijn, K.; Roos, C.; Assies, J.; Bergink, V.; Ruhe, H.G.; Schene, A.H. Omega-3 Fatty Acid Supplementation for Perinatal Depression: A Meta-Analysis. J. Clin. Psychiatry 2020, 81, 13281. [Google Scholar] [CrossRef] [PubMed]
- Chambergo-Michilot, D.; Branez-Condorena, A.; Falvy-Bockos, I.; Pacheco-Mendoza, J.; Benites-Zapata, V.A. Efficacy of omega-3 supplementation on sertraline continuous therapy to reduce depression or anxiety symptoms: A systematic review and meta-analysis. Psychiatry Res. 2021, 296, 113652. [Google Scholar] [CrossRef] [PubMed]
- Deane, K.H.O.; Jimoh, O.F.; Biswas, P.; O’Brien, A.; Hanson, S.; Abdelhamid, A.S.; Fox, C.; Hooper, L. Omega-3 and polyunsaturated fat for prevention of depression and anxiety symptoms: Systematic review and meta-analysis of randomised trials. Br. J. Psychiatry 2021, 218, 135–142. [Google Scholar] [CrossRef]
- Appleton, K.M.; Voyias, P.D.; Sallis, H.M.; Dawson, S.; Ness, A.R.; Churchill, R.; Perry, R. Omega-3 fatty acids for depression in adults. Cochrane Database Syst. Rev. 2021, 11, CD004692. [Google Scholar] [CrossRef]
- Chevalier, G.; Siopi, E.; Guenin-Mace, L.; Pascal, M.; Laval, T.; Rifflet, A.; Boneca, I.G.; Demangel, C.; Colsch, B.; Pruvost, A.; et al. Effect of gut microbiota on depressive-like behaviors in mice is mediated by the endocannabinoid system. Nat. Commun. 2020, 11, 6363. [Google Scholar] [CrossRef]
- Almulla, A.F.; Thipakorn, Y.; Vasupanrajit, A.; Abo Algon, A.A.; Tunvirachaisakul, C.; Hashim Aljanabi, A.A.; Oxenkrug, G.; Al-Hakeim, H.K.; Maes, M. The tryptophan catabolite or kynurenine pathway in major depressive and bipolar disorder: A systematic review and meta-analysis. Brain Behav. Immun. Health 2022, 26, 100537. [Google Scholar] [CrossRef]
- McNamara, R.K.; Able, J.A.; Rider, T.; Tso, P.; Jandacek, R. Effect of chronic fluoxetine treatment on male and female rat erythrocyte and prefrontal cortex fatty acid composition. Prog. Neuropsychopharmacol. Biol. Psychiatry 2010, 34, 1317–1321. [Google Scholar] [CrossRef] [Green Version]
- Thomas, J.; Khanam, R.; Vohora, D. Activation of indoleamine 2, 3-dioxygenase pathway by olanzapine augments antidepressant effects of venlafaxine in mice. Psychiatry Res. 2017, 258, 444–448. [Google Scholar] [CrossRef]
- Molina, J.D.; Avila, S.; Rubio, G.; Lopez-Munoz, F. Metabolomic Connections between Schizophrenia, Antipsychotic Drugs and Metabolic Syndrome: A Variety of Players. Curr. Pharm. Des. 2021, 27, 4049–4061. [Google Scholar] [CrossRef] [PubMed]
- Qureshi, W.; Santaren, I.D.; Hanley, A.J.; Watkins, S.M.; Lorenzo, C.; Wagenknecht, L.E. Risk of diabetes associated with fatty acids in the de novo lipogenesis pathway is independent of insulin sensitivity and response: The Insulin Resistance Atherosclerosis Study (IRAS). BMJ Open Diabetes Res. Care 2019, 7, e000691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiu, H.H.; Tsai, S.J.; Tseng, Y.J.; Wu, M.S.; Liao, W.C.; Huang, C.S.; Kuo, C.H. An efficient and robust fatty acid profiling method for plasma metabolomic studies by gas chromatography-mass spectrometry. Clin. Chim. Acta 2015, 451, 183–190. [Google Scholar] [CrossRef] [PubMed]
- Jackson, K.H.; Van Guilder, G.P.; Tintle, N.; Tate, B.; McFadden, J.; Perry, C.A. Plasma fatty acid responses to a calorie-restricted, DASH-style diet with lean beef. Prostaglandins Leukot. Essent. Fat. Acids 2022, 179, 102413. [Google Scholar] [CrossRef]
- Bogie, J.F.J.; Haidar, M.; Kooij, G.; Hendriks, J.J.A. Fatty acid metabolism in the progression and resolution of CNS disorders. Adv. Drug Deliv. Rev. 2020, 159, 198–213. [Google Scholar] [CrossRef]
- Machate, D.J.; Figueiredo, P.S.; Marcelino, G.; Guimaraes, R.C.A.; Hiane, P.A.; Bogo, D.; Pinheiro, V.A.Z.; Oliveira, L.C.S.; Pott, A. Fatty Acid Diets: Regulation of Gut Microbiota Composition and Obesity and Its Related Metabolic Dysbiosis. Int. J. Mol. Sci. 2020, 21, 4093. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Parameter | HC (n = 23) | MDD (n = 24) | SCH (n = 22) | F/H/ χ2 Value | p-Value |
|---|---|---|---|---|---|
| Sociodemographic | |||||
| Age [years, M (P25, P75)] a | 29 (26, 32) | 33 (27.25, 36) | 31 (27, 34.75) | H = 2.488 | 0.288 |
| Gender (male/female) c | 7/16 | 8/16 | 7/15 | χ2 = 0.045 | 0.978 |
| BMI [kg/m2, mean ± SD] b | 20.91 ± 2.78 | 21.01 ± 2.51 | 22.85 ± 3.65 | F = 2.967 | 0.058 |
| Marital status (single/married) c | 9/14 | 1/23 | 15/7 | χ2 = 20.484 | <0.001 |
| Smoking (almost none/intermittent) c | 16/7 | 13/11 | 14/8 | χ2 = 1.210 | 0.546 |
| Scale evaluation | |||||
| HAMD (mean ± SD) a | 3.61 ± 1.75 | 21.92 ± 7.62 * | 7.00 ± 2.94 *# | H = 50.719 | <0.001 |
| HAMA [M (P25, P75)] a | 5 (2, 6) | 24 (17, 29) * | 6 (4, 8) # | H = 44.128 | <0.001 |
| PANSS (T) [ M (P25, P75)] a | 34 (32, 39) | 58.50 (50.25, 65) * | 72.5 (59.25, 83.75) * | H = 44.576 | <0.001 |
| PANSS (P) [M (P25, P75)] a | 7 (7, 8) | 7 (7, 7) | 18 (15.5, 25) *# | H = 47.114 | <0.001 |
| PANSS (N) [M (P25, P75)] a | 8 (7, 9) | 11.00 (7.25, 13) * | 14.00 (12, 19.25) *# | H = 29.097 | <0.001 |
| PANSS (G) (mean ± SD) a | 19.39 ± 2.41 | 40.38 ± 8.35 * | 38.73 ± 10.57 * | H = 43.176 | <0.001 |
| Parameter (ng/mL) | HC | MDD | SCH | F Value | p-Value |
|---|---|---|---|---|---|
| l-kynurenine | 108.35 ± 14.12 | 117.55 ± 24.81 | 117.23 ± 19.26 | 1.570 | 0.216 |
| Picolinic acid | 6.62 ± 1.67 | 7.68 ± 2.98 | 7.39 ± 2.43 | 1.197 | 0.309 |
| Quinolinic acid | 41.19 ± 14.63 | 36.36 ± 3.81 | 38.44 ± 4.27 | 1.679 | 0.194 |
| 3-Indoxyl sulfate | 356.22 ± 185.53 | 317.24 ± 185.12 | 420.62 ± 200.26 | 1.856 | 0.164 |
| Parameter (μg/mL) | HC | MDD | SCH | F Value | p-Value |
|---|---|---|---|---|---|
| C8:0 | 0.183 ± 0.05 | 0.010 ± 0.003 ** | 0.010 ± 0.002 ** | 33.082 | <0.001 |
| C10:0 | 0.128 ± 0.001 | 0.009 ± 0.004 ** | 0.009 ± 0.004 ** | 6.664 | 0.002 |
| C12:0 | 0.166 ± 0.092 | 0.105 ± 0.052 ** | 0.119 ± 0.058 | 4.875 | 0.011 |
| C13:0 | 0.078 ± 0.071 | 0.011 ± 0.008 ** | 0.011 ± 0.008 ** | 16.166 | <0.001 |
| C14:0 | 3.859 ± 1.532 | 3.201 ± 1.300 | 3.310 ± 1.263 | 1.551 | 0.22 |
| C15:0 | 0.943 ± 0.279 | 0.941 ± 0.308 | 0.914 ± 0.316 | 0.071 | 0.931 |
| C16:0 | 223.736 ± 47.742 | 203.393 ± 35.148 | 197.029 ± 36.323 | 2.788 | 0.069 |
| C17:0 | 2.307 ± 0.548 | 2.020 ± 0.509 | 2.221 ± 0.552 | 1.71 | 0.189 |
| C18:0 | 96.214 ± 16.662 | 91.945 ± 11.846 # | 80.646 ± 17.381 ** | 6.127 | 0.004 |
| C20:0 | 0.755 ± 0.172 | 0.790 ± 0.289 ## | 1.381 ± 1.172 ** | 5.872 | 0.004 |
| C21:0 | 0.111 ± 0.027 | 0.113 ± 0.026 | 0.102 ± 0.024 | 1.031 | 0.362 |
| C22:0 | 0.234 ± 0.099 | 0.259 ± 0.056 | 0.209 ± 0.086 | 2.056 | 0.136 |
| C23:0 | 0.086 ± 0.033 | 0.091 ± 0.022 | 0.100 ± 0.032 | 1.194 | 0.309 |
| C24:0 | 0.843 ± 0.169 | 0.937 ± 0.319 | 1.015 ± 0.317 | 2.261 | 0.112 |
| C14:1N5 | 1.179 ± 0.428 | 1.573 ± 0.492 * | 1.677 ± 0.812 * | 4.551 | 0.014 |
| C16:1N7 | 11.553 ± 5.369 | 11.227 ± 2.719 | 9.270 ± 3.681 | 2.045 | 0.137 |
| C17:1N7 | 0.504 ± 0.200 | 0.576 ± 0.239 | 0.670 ± 0.308 | 2.521 | 0.088 |
| C18:1TN9 | 1.153 ± 0.280 | 1.455 ± 0.548 | 1.149 ± 0.834 | 2.031 | 0.139 |
| C18:1N9 | 182.544 ± 38.631 | 175.740 ± 39.452 | 165.288 ± 44.211 | 1.039 | 0.36 |
| C20:1N9 | 2.695 ± 0.928 | 2.592 ± 0.822 | 2.174 ± 0.685 | 2.554 | 0.085 |
| C22:1N9 | 6.020 ± 1.330 | 2.985 ± 2.111 ** | 6.104 ± 2.437 && | 18.178 | <0.001 |
| C24:1N9 | 14.558 ± 2.618 | 15.150 ± 3.552 ## | 10.549 ± 2.793 ** | 15.481 | <0.001 |
| C18:3N3 | 11.857 ± 4.518 | 10.793 ± 4.559 | 9.269 ± 3.454 | 1.948 | 0.151 |
| C20:3N3 | 0.587 ± 0.153 | 0.571 ± 0.285 | 0.511 ± 0.132 | 0.894 | 0.414 |
| C20:5N3 (EPA) | 11.573 ± 3.491 | 9.923 ± 2.362 | 11.056 ± 2.591 * | 2.060 | 0.136 |
| C22:6N3 (DHA) | 15.966 ± 4.189 | 11.276 ± 3.182 ** | 12.777 ± 2.447 ** | 11.865 | <0.001 |
| C22:5N3 (DPA) | 14.801 ± 3.149 | 12.519 ± 3.708 * | 12.464 ± 2.448 * | 4.083 | 0.021 |
| C18:2N6 | 928.338 ± 206.304 | 1085.928 ± 418.170 | 1170.837 ± 291.339 * | 3.354 | 0.041 |
| C18:3N6 | 1.290 ± 0.456 | 2.789 ± 1.344 ** | 2.561 ± 1.049 ** | 14.976 | <0.001 |
| C20:2N6 | 3.687 ± 0.671 | 3.594 ± 0.631 | 3.219 ± 0.719 | 3.059 | 0.054 |
| C20:3N6 | 14.085 ± 3.615 | 13.711 ± 4.045 | 12.383 ± 4.284 | 1.142 | 0.325 |
| C20:4N6 | 69.490 ± 14.571 | 65.819 ± 13.499 | 58.883 ± 14.304 * | 3.307 | 0.043 |
| C22:2N6 | 0.605 ± 0.143 | 0.788 ± 0.226 ** | 0.557 ± 0.199 && | 9.249 | <0.001 |
| C22:4N6 | 2.300 ± 0.805 | 2.840 ± 0.682 * | 2.638 ± 0.559 | 3.651 | 0.031 |
| C22:5N6 | 2.109 ± 0.664 | 2.397 ± 0.716 | 2.029 ± 0.615 | 1.916 | 0.155 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, J.-C.; Yu, H.; Li, R.; Zhou, C.-H.; Shi, Q.-Q.; Guo, L.; He, H. A Preliminary Comparison of Plasma Tryptophan Metabolites and Medium- and Long-Chain Fatty Acids in Adult Patients with Major Depressive Disorder and Schizophrenia. Medicina 2023, 59, 413. https://doi.org/10.3390/medicina59020413
Liu J-C, Yu H, Li R, Zhou C-H, Shi Q-Q, Guo L, He H. A Preliminary Comparison of Plasma Tryptophan Metabolites and Medium- and Long-Chain Fatty Acids in Adult Patients with Major Depressive Disorder and Schizophrenia. Medicina. 2023; 59(2):413. https://doi.org/10.3390/medicina59020413
Chicago/Turabian StyleLiu, Jun-Chang, Huan Yu, Rui Li, Cui-Hong Zhou, Qing-Qing Shi, Li Guo, and Hong He. 2023. "A Preliminary Comparison of Plasma Tryptophan Metabolites and Medium- and Long-Chain Fatty Acids in Adult Patients with Major Depressive Disorder and Schizophrenia" Medicina 59, no. 2: 413. https://doi.org/10.3390/medicina59020413