Mature miR-99a Upregulation in the Amniotic Fluid Samples from Female Fetus Down Syndrome Pregnancies: A Pilot Study

 , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Subjects
2.2. Sample Collection and Down Syndrome Diagnostic
2.3. Gene Expression Analysis
2.4. Statistical Analyses
2.5. Target Predictions and Bioinformatics Analysis
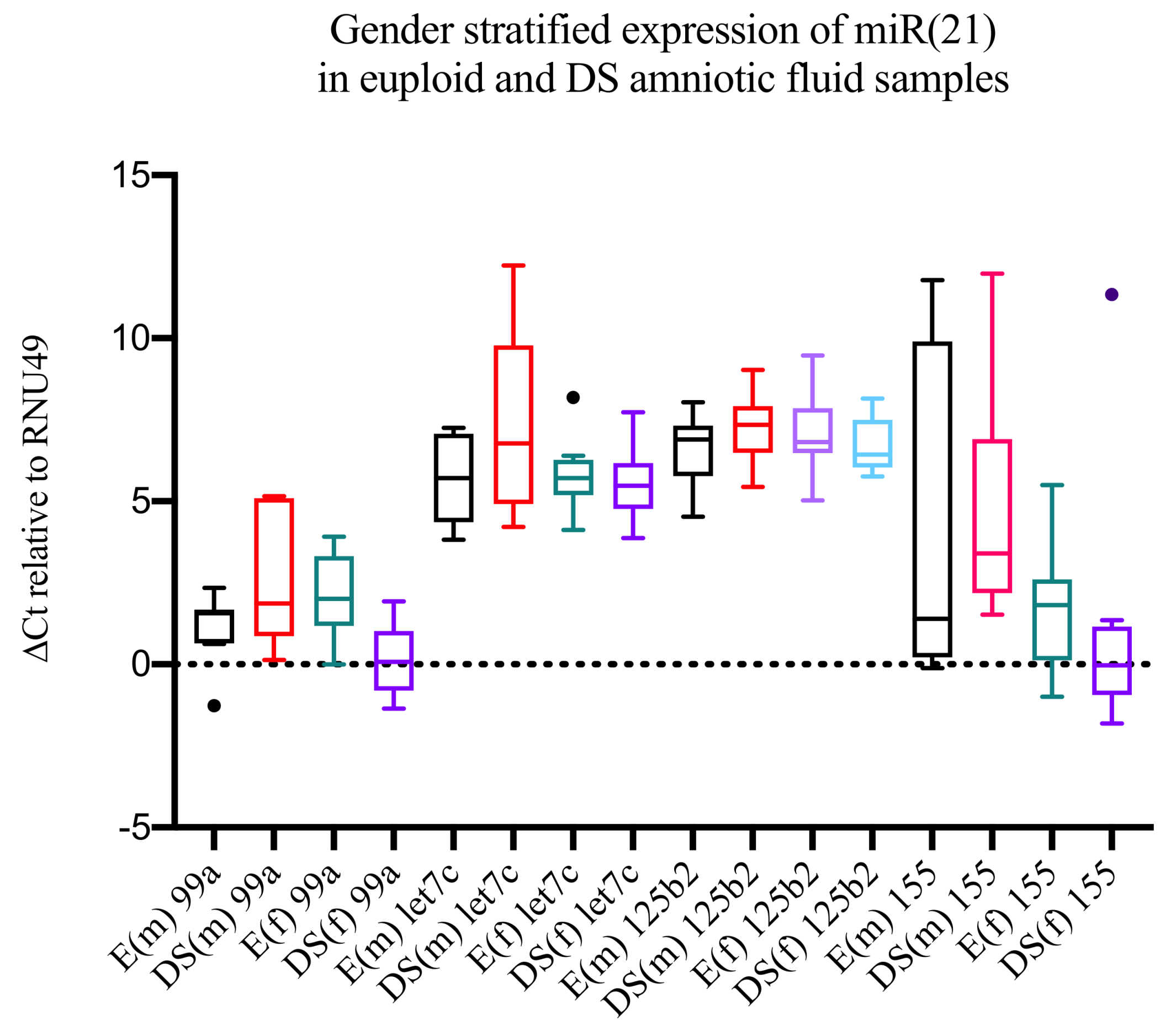
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Cartier, L.; Murphy-Kaulbeck, L. Counselling considerations for prenatal genetic screening. J. Obstet. Gynaecol. Can. 2012, 34, 489–493. [Google Scholar] [CrossRef]
- Hulten, M.A.; Patel, S.D.; Tankimanova, M.; Westgren, M.; Papadogiannakis, N.; Jonsson, A.M.; Iwarsson, E. On the origin of trisomy 21 Down syndrome. Mol. Cytogenet. 2008, 1, 21. [Google Scholar] [CrossRef] [PubMed]
- Antonarakis, S.E.; Lyle, R.; Dermitzakis, E.T.; Reymond, A.; Deutsch, S. Chromosome 21 and down syndrome: From genomics to pathophysiology. Nat. Rev. Genet. 2004, 5, 725–738. [Google Scholar] [CrossRef] [PubMed]
- Korenberg, J.R.; Chen, X.N.; Schipper, R.; Sun, Z.; Gonsky, R.; Gerwehr, S.; Carpenter, N.; Daumer, C.; Dignan, P.; Disteche, C.; et al. Down syndrome phenotypes: The consequences of chromosomal imbalance. Proc. Natl. Acad. Sci. USA 1994, 91, 4997–5001. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, M.A.; Kola, I. The “gene dosage effect” hypothesis versus the “amplified developmental instability” hypothesis in Down syndrome. J. Neural Transm. 1999, 57, 293–303. [Google Scholar]
- Mouillet, J.-F.; Ouyang, Y.; Coyne, C.; Sadovsky, Y. MicroRNAs in placental health and disease. Am. J. Obstet. Gynecol. 2015, 213, S163–S172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamhieh-Milz, J.; Moftah, R.F.H.; Bal, G.; Futschik, M.; Sterzer, V.; Khorramshahi, O.; Burow, M.; Thiel, G.; Stuke-Sontheimer, A.; Chaoui, R.; et al. Differentially expressed microRNAs in maternal plasma for the noninvasive prenatal diagnosis of Down syndrome (trisomy 21). Biomed. Res. Int. 2014, 2014, 402475. [Google Scholar] [CrossRef]
- Kotlabova, K.; Doucha, J.; Chudoba, D.; Calda, P.; Dlouha, K.; Hromadnikova, I. Extracellular chromosome 21-derived microRNAs in euploid & aneuploid pregnancies. Indian J. Med. Res. 2013, 138, 935–943. [Google Scholar] [PubMed]
- Xu, Y.; Li, W.; Liu, X.; Ma, H.; Tu, Z.; Dai, Y. Analysis of microRNA expression profile by small RNA sequencing in Down syndrome fetuses. Int. J. Mol. Med. 2013, 32, 1115–1125. [Google Scholar] [CrossRef]
- Lim, J.H.; Kim, D.J.; Lee, D.E.; Han, J.Y.; Chung, J.H.; Ahn, H.K.; Lee, S.W.; Lim, D.H.; Lee, Y.S.; Park, S.Y.; et al. Genome-wide microRNA expression profiling in placentas of fetuses with Down syndrome. Placenta 2015, 36, 322–328. [Google Scholar] [CrossRef]
- Modi, B.P.; Washington, S.; Walsh, S.W.; Jackson-Cook, C.; Archer, K.J.; Strauss, J.F., 3rd. Expression patterns of the chromosome 21 MicroRNA cluster (miR-99a, miR-125b and let-7c) in chorioamniotic membranes. Placenta 2017, 49, 1–9. [Google Scholar] [CrossRef]
- Erturk, B.; Karaca, E.; Aykut, A.; Durmaz, B.; Guler, A.; Buke, B.; Yeniel, A.O.; Ergenoglu, A.M.; Ozkinay, F.; Ozeren, M.; et al. Prenatal evaluation of microRNA expressions in pregnancies with Down syndrome. Biomed. Res. Int. 2016, 2016, 5312674. [Google Scholar] [CrossRef]
- Rozovski, U.; Jonish-Grossman, A.; Bar-Shira, A.; Ochshorn, Y.; Yaron, Y.; Goldstein, M. Genome-wide expression analysis of cultured trophoblast with trisomy 21 karyotype. Hum. Reprod. 2007, 22, 2538–2545. [Google Scholar] [CrossRef] [Green Version]
- Miura, K.; Miura, S.; Yamasaki, K.; Higashijima, A.; Kinoshita, A.; Yoshiura, K.-I.; Masuzaki, H. Identification of pregnancy-associated microRNAs in maternal plasma. Clin. Chem. 2010, 56, 1767–1771. [Google Scholar] [CrossRef]
- Luo, S.S.; Ishibashi, O.; Ishikawa, G.; Ishikawa, T.; Katayama, A.; Mishima, T.; Takizawa, T.; Shigihara, T.; Goto, T.; Izumi, A.; et al. Human villous trophoblasts express and secrete placenta-specific microRNAs into maternal circulation via exosomes. Biol. Reprod. 2009, 81, 717–729. [Google Scholar] [CrossRef] [PubMed]
- Kozomara, A.; Griffiths-Jones, S. miRBase: Annotating high confidence microRNAs using deep sequencing data. NAR 2014, 42, D68–D73. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Sui, W.; Li, W.; Tan, Q.; Chen, J.; Lin, X.; Guo, H.; Ou, M.; Xue, W.; Zhang, R.; et al. Integrated microRNA and protein expression analysis reveals novel microRNA regulation of targets in fetal down syndrome. Mol. Med. Rep. 2016, 14, 4109–4118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slonim, D.K.; Koide, K.; Johnson, K.L.; Tantravahi, U.; Cowan, J.M.; Jarrah, Z.; Bianchi, D.W. Functional genomic analysis of amniotic fluid cell-free mRNA suggests that oxidative stress is significant in Down syndrome fetuses. Proc. Natl. Acad. Sci. USA 2009, 106, 9425–9429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, R.; Wang, X.; Spitznagel, E.L., Jr.; Frelin, L.P.; Ting, J.C.; Ding, H.; Kim, J.-W.; Ruczinski, I.; Downey, T.J.; Pevsner, J. Primary and secondary transcriptional effects in the developing human Down syndrome brain and heart. Genome Biol. 2005, 6, R107. [Google Scholar] [CrossRef]
- Ding, J.; Li, X.; Hu, H. TarPmiR: A new approach for microRNA target site prediction. Bioinformatics 2016, 32, 2768–2775. [Google Scholar] [CrossRef]
- Chang, J.T.; Nevins, J.R. GATHER: A systems approach to interpreting genomic signatures. Bioinformatics 2006, 22, 2926–2933. [Google Scholar] [CrossRef] [PubMed]
- Varley, K.E.; Gertz, J.; Bowling, K.M.; Parker, S.L.; Reddy, T.E.; Pauli-Behn, F.; Cross, M.K.; Williams, B.A.; Stamatoyannopoulos, J.A.; Crawford, G.E.; et al. Dynamic DNA methylation across diverse human cell lines and tissues. Genome Res. 2013, 23, 555–567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grigoriu, A.; Ferreira, J.C.; Choufani, S.; Baczyk, D.; Kingdom, J.; Weksberg, R. Cell specific patterns of methylation in the human placenta. Epigenetics 2011, 6, 368–379. [Google Scholar] [CrossRef] [Green Version]
- Schroeder, D.I.; Blair, J.D.; Lott, P.; Yu, H.O.; Hong, D.; Crary, F.; Ashwood, P.; Walker, C.; Korf, I.; Robinson, W.P.; et al. The human placenta methylome. Proc. Natl. Acad. Sci. USA 2013, 110, 6037–6042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Izzo, A.; Manco, R.; de Cristofaro, T.; Bonfiglio, F.; Cicatiello, R.; Mollo, N.; De Martino, M.; Genesio, R.; Zannini, M.; Conti, A.; et al. Overexpression of chromosome 21 miRNAs may affect mitochondrial function in the hearts of Down syndrome fetuses. Int. J. Genom. 2017, 2017, 8737649. [Google Scholar] [CrossRef] [PubMed]
- Coppola, A.; Romito, A.; Borel, C.; Gehrig, C.; Gagnebin, M.; Falconnet, E.; Izzo, A.; Altucci, L.; Banfi, S.; Antonarakis, S.E.; et al. Cardiomyogenesis is controlled by the miR-99a/let-7c cluster and epigenetic modifications. Stem Cell Res. 2014, 12, 323–337. [Google Scholar] [CrossRef]
- Kehler, L.; Biro, O.; Lazar, L.; Rigo, J.; Nagy, B. Elevated hsa-miR-99a levels in maternal plasma may indicate congenital heart defects. Biomed. Rep. 2017, 3, 869–873. [Google Scholar] [CrossRef]
- Underwood, M.A.; Gilbert, W.M.; Sherman, M.P. Amniotic fluid: Not just fetal urine anymore. J. Perinatol. 2005, 25, 341–348. [Google Scholar] [CrossRef] [PubMed]
- Diogenes, T.C.P.; Mourato, F.A.; de Lima Filho, J.L.; Mattos, S.D.S. Gender differences in the prevalence of congenital heart disease in Down’s syndrome: A brief meta-analysis. BMC Med. Genet. 2017, 18, 111. [Google Scholar] [CrossRef]
- Santoro, M.; Coi, A.; Spadoni, I.; Bianchi, F.; Pierini, A. Sex differences for major congenital heart defects in Down syndrome: A population based study. Eur. J. Med. Genet. 2018, 61, 546–550. [Google Scholar] [CrossRef] [PubMed]
- Takano, T.; Akagi, M.; Takaki, H.; Inuzuka, R.; Nogimori, Y.; Ono, H.; Kaneko, M.; Hagiwara, N. Sex differences in congenital heart disease in Down syndrome: Study data from medical records and questionnaires in a region of Japan. BMJ Paediatr. Open 2019, 3, e000414. [Google Scholar] [CrossRef] [PubMed]
- Katz, M.G.; Fargnoli, A.S.; Kendle, A.P.; Hajjar, R.J.; Bridges, C.R. The role of microRNAs in cardiac development and regenerative capacity. Am. J. Physiol. Heart Circ. Physiol. 2016, 310, H528–H541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguirre, A.; Montserrat, N.; Zacchigna, S.; Nivet, E.; Hishida, T.; Krause, M.N.; Kurian, L.; Ocampo, A.; Vazquez-Ferrer, E.; Rodríguez-Esteban, C.; et al. In vivo activation of a conserved microRNA program induces mammalian heart regeneration. Cell Stem Cell 2014, 15, 589–604. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.C.; Chang, Y.M.; Pan, C.T.; Chen, C.C.; Ling, L.; Tsao, K.C.; Yang, R.B.; Li, W.H. Functional evolution of cardiac microRNAs in heart development and functions. Mol. Biol. Evol. 2014, 31, 2722–2734. [Google Scholar] [CrossRef]
- Li, Q.; Xie, J.; Wang, B.; Li, R.; Bai, J.; Ding, L.; Gu, R.; Xu, B. Overexpression of microRNA-99a attenuates cardiac hypertrophy. PLoS ONE 2016, 11, e0148480. [Google Scholar] [CrossRef]
- Li, Q.; Xie, J.; Li, R.; Shi, J.; Sun, J.; Gu, R.; Ding, L.; Wang, L.; Xu, B. Overexpression of microRNA-99a attenuates heart remodelling and improves cardiac performance after myocardial infarction. J. Cell. Mol. Med. 2014, 18, 919–928. [Google Scholar] [CrossRef] [PubMed]
- He, A.; Pu, W.T. Mature cardiomyocytes recall their progenitor experience via polycomb repressive complex 2. Circ. Res. 2012, 111, 162–164. [Google Scholar] [CrossRef] [PubMed]
- Mogra, R.; Zidere, V.; Allan, L.D. Prenatally detectable congenital heart defects in fetuses with Down syndrome. Ultrasound Obstet. Gynecol. 2011, 38, 320–324. [Google Scholar] [CrossRef] [PubMed]
- van der Bom, T.; Zomer, A.C.; Zwinderman, A.H.; Meijboom, F.J.; Bouma, B.J.; Mulder, B.J. The changing epidemiology of congenital heart disease. Nat. Rev. Cardiol. 2011, 8, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Smith, T.; Rajakaruna, C.; Caputo, M.; Emanueli, C. MicroRNAs in congenital heart disease. Ann. Transl. Med. 2015, 3, 333. [Google Scholar] [PubMed]
- Saxena, A.; Tabin, C.J. miRNA-processing enzyme Dicer is necessary for cardiac outflow tract alignment and chamber septation. Proc. Natl. Acad. Sci. USA 2010, 107, 87–91. [Google Scholar] [CrossRef]
- Ackerman, C.; Locke, A.E.; Feingold, E.; Reshey, B.; Espana, K.; Thusberg, J.; Mooney, S.; Bean, L.J.; Dooley, K.J.; Cua, C.L.; et al. An excess of deleterious variants in VEGF-A pathway genes in Down-syndrome-associated atrioventricular septal defects. Am. J. Hum. Genet. 2012, 91, 646–659. [Google Scholar] [CrossRef] [PubMed]
- Ripoll, C.; Rivals, I.; Yahya-Graison, E.A.; Dauphinot, L.; Paly, E.; Mircher, C.; Ravel, A.; Grattau, Y.; Blehaut, H.; Mégarbané, A.; et al. Molecular signatures of cardiac defects in Down syndrome lymphoblastoid cell lines suggest altered ciliome and Hedgehog pathways. PLoS ONE 2012, 7, e41616. [Google Scholar] [CrossRef] [PubMed]
- Locke, A.E.; Dooley, K.J.; Tinker, S.W.; Cheong, S.Y.; Feingold, E.; Allen, E.G.; Freeman, S.B.; Torfs, C.P.; Cua, C.L.; Epstein, M.P.; et al. Variation in folate pathway genes contributes to risk of congenital heart defects among individuals with Down syndrome. Genet. Epidemiol. 2010, 34, 613–623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, S.; Yi, H.; Wang, Z.; Dong, J. Screening key genes associated with congenital heart defects in Down syndrome based on differential expression network. Int. J. Clin. Exp. Pathol. 2015, 8, 8385–8393. [Google Scholar] [PubMed]
- Pinto, F.F.; Nunes, L.; Ferraz, F.; Sampayo, F. Down’s syndrome: Different distribution of congenital heart diseases between the sexes. Int. J. Cardiol. 1990, 27, 175–178. [Google Scholar] [CrossRef]
- Källén, B.; Mastroiacovo, P.; Robert, E. Major congenital malformations in Down syndrome. Am. J. Med. Genet. 1996, 65, 160–166. [Google Scholar] [CrossRef]
- Freeman, S.B.; Bean, L.H.; Allen, E.G.; Tinker, S.W.; E Locke, A.; Druschel, C.; A Hobbs, C.; A Romitti, P.; Royle, M.H.; Torfs, C.P.; et al. Ethnicity, sex, and the incidence of congenital heart defects: A report from the National Down Syndrome Project. Genet. Med. 2008, 10, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Weijerman, M.E.; van Furth, A.M.; Noordegraaf, A.V.; van Wouwe, J.P.; Broers, C.J.; Gemke, R.J. Prevalence, neonatal characteristics, and first-year mortality of Down syndrome: A national study. J. Pediatr. 2008, 152, 15–19. [Google Scholar] [CrossRef]
- Rankin, J.; Tennant, P.W.; Bythell, M.; Pearce, M.S. Predictors of survival in children born with Down syndrome: A registry-based study. Pediatrics 2012, 129, 1373–1381. [Google Scholar] [CrossRef] [PubMed]
- Bergström, S.; Carr, H.; Petersson, G.; Stephansson, O.; Bonamy, A.K.; Dahlström, A.; Halvorsen, C.P.; Johansson, S. Trends in congenital heart defects in infants with Down syndrome. Pediatrics 2016, 138, e20160123. [Google Scholar] [CrossRef]
- Mourato, F.A.; Villachan, L.R.; Sda, S.M. Prevalence and profile of congenital heart disease and pulmonary hypertension in Down syndrome in a pediatric cardiology service. Rev. Paul. Pediatr. 2014, 32, 159–163. [Google Scholar] [CrossRef] [Green Version]
- Morris, J.K.; Garne, E.; Wellesley, D.; Addor, M.C.; Arriola, L.; Barisic, I.; Beres, J.; Bianchi, F.; Budd, J.; Dias, C.M.; et al. Major congenital anomalies in babies born with Down syndrome: A EUROCAT population-based registry study. Am. J. Med. Genet. A 2014, 164, 2979–2986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elton, T.S.; Sansom, S.E.; Martin, M.M. Trisomy-21 gene dosage overexpression of miRNAs results in the haploinsufficiency of specific target proteins. RNA Biol. 2007, 7, 540–547. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.; Wu, X.; Druso, J.E.; Wei, H.; Park, A.Y.; Kraus, M.S.; Alcaraz, A.; Chen, J.; Chien, S.; Cerione, R.A.; et al. Cardiac developmental defects and eccentric right ventricular hypertrophy in cardiomyocyte focal adhesion kinase (FAK) conditional knockout mice. Proc. Natl. Acad. Sci. USA 2008, 105, 6638–6643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hakim, Z.S.; DiMichele, L.A.; Doherty, J.T.; Homeister, J.W.; Beggs, H.E.; Reichardt, L.F.; Schwartz, R.J.; Brackhan, J.; Smithies, O.; Mack, C.P.; et al. Conditional deletion of focal adhesion kinase leads to defects in ventricular septation and outflow tract alignment. Mol. Cell Biol. 2007, 27, 5352–5364. [Google Scholar] [CrossRef] [PubMed]
- Doherty, J.T.; Conlon, F.L.; Mack, C.P.; Taylor, J.M. Focal adhesion kinase is essential for cardiac looping and multichamber heart formation. Genesis 2010, 48, 492–504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conti, A.; Fabbrini, F.; D’Agostino, P.; Negri, R.; Greco, D.; Genesio, R.; D’Armiento, M.; Olla, C.; Paladini, D.; Zannini, M.; et al. Altered expression of mitochondrial and extracellular matrix genes in the heart of human fetuses with chromosome 21 trisomy. BMC Genom. 2007, 8, 268. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Zhang, Z.; Meng, Y. Systematic tracking of disrupted modules identifies altered pathways associated with congenital heart defects in Down syndrome. Med. Sci. Monit. 2015, 21, 3334–3342. [Google Scholar] [CrossRef]
- Li, C.; Jin, L.; Bai, Y.; Chen, Q.; Fu, L.; Yang, M.; Xiao, H.; Zhao, G.; Wang, S. Genome-wide expression analysis in Down syndrome: Insight into immunodeficiency. PLoS ONE 2012, 7, e49130. [Google Scholar] [CrossRef]
- Vis, J.C.; van Engelen, K.; Bouma, B.J.; Bilardo, C.M.; Blom, N.A.; Mulder, B. Cardiovascular disorders among persons with Down syndrome. In International Review of Research in Mental Retardation; Academic Press: Cambridge, MA, USA, 2010; Volume 39, pp. 65–194. [Google Scholar]
- Grossman, T.R.; Gamliel, A.; Wessells, R.J.; Taghli-Lamallem, O.; Jepsen, K.; Ocorr, K.; Korenberg, J.R.; Peterson, K.L.; Rosenfeld, M.G.; Bodmer, R.; et al. Over-expression of DSCAM and COL6A2 cooperatively generates congenital heart defects. PLoS Genet. 2011, 7, e1002344. [Google Scholar] [CrossRef] [PubMed]
- Qing, M.; Schumacher, K.; Heise, R.; Wöltje, M.; Vazquez-Jimenez, J.F.; Richter, T.; Arranda-Carrero, M.; Hess, J.; Von Bernuth, G.; Seghaye, M.C. Intramyocardial synthesis of pro-and anti-inflammatory cytokines in infants with congenital cardiac defects. J. Am. Coll. Cardiol. 2003, 41, 2266–2274. [Google Scholar] [CrossRef]
- Fan, S.; Li, K.; Zhang, D.; Liu, F. JNK and NF-κB signaling pathways are involved in cytokine changes in patients with congenital heart disease prior to and after transcatheter closure. Exp. Ther. Med. 2018, 15, 1525–1531. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Gutierrez, S.; García-Peláez, I.; Zentella-Dehesa, A.; Ramos-Kuri, M.; Hernández-Franco, P.; Hernández-Sánchez, F.; Rojas, E. NF-κB signaling blockade by Bay 11-7085 during early cardiac morphogenesis induces alterations of the outflow tract in chicken heart. Apoptosis 2006, 11, 1101–1109. [Google Scholar] [CrossRef]
- Neeb, Z.; Lajiness, J.D.; Bolanis, E.; Conway, S.J. Cardiac outflow tract anomalies. Wiley Interdiscip. Rev. Dev. Biol. 2013, 2, 499–530. [Google Scholar] [Green Version]
- Yeh, C.C.; Fan, Y.; Yang, Y.L.; Mann, M.J. Atrial ERK1/2 activation in the embryo leads to incomplete Septal closure: A novel mouse model of atrial Septal defect. J. Biomed. Sci. 2017, 24, 89. [Google Scholar] [CrossRef]
- Tian, Y.; Cohen, E.D.; Morrisey, E.E. The importance of Wnt signaling in cardiovascular development. Pediatr. Cardiol. 2010, 31, 342–348. [Google Scholar] [CrossRef]
- Marzi, M.J.; Ghini, F.; Cerruti, B.; de Pretis, S.; Bonetti, P.; Giacomelli, C.; Gorski, M.M.; Kress, T.; Pelizzola, M.; Muller, H.; et al. Degradation dynamics of microRNAs revealed by a novel pulse-chase approach. Genome Res. 2016, 26, 554–565. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| DS (n = 17) (Mean ± SD) | E (n = 22) (Mean ± SD) | p Value (t, df) | ||
|---|---|---|---|---|
| Maternal age | M + F | 35.65 ± 5.11 | 34.98 ± 4.98 | 0.684 (0.4105, 34.09) |
| M | 36.83 ± 5.31 | 34.88 ± 6.17 | 0.537 (0.6367, 11.70) | |
| F | 35.00 ± 5.14 | 35.04 ± 4.41 | 0.986 (0.0183, 19.83) | |
| Gestational age | M + F | 17.17 ± 1.85 | 18.44 ± 1.72 | 0.036 (2.184, 33,3) |
| M | 17.13 ± 2.42 | 17.96 ± 1.21 | 0.467 (0.77, 6.89) | |
| F | 17.19 ± 1.59 | 18.71 ± 1.95 | 0.043 (2.14, 22.94) | |
| % Males | 35.29 | 36.36 | 0.944 * | |
| Fold Changes (p Value/t, df) | |||
|---|---|---|---|
| M + F | M | F | |
| let7c | 0.61 (0.422/0.82, 23.49) | 0.12 (0.239/1.29, 6.69) | 1.16 (0.612/0.52, 20.37) |
| miR-125b-2 | 1.04 (0.998/0.002, 35.95) | 0.51 (0.280/1.14, 10.66) | 1.35 (0.357/0.94, 22) |
| miR-155 | 1.16 (0.504 *) | 0.81 (0.282 *) | 2.08 (0.072 *) |
| miR-99a | 2.07 (0.166/1.42, 26) | 0.61 (0.173/1.52, 7.08) | 3.84 (0.0005/4.13, 21.64) |
| Pri-miR-99a | 0.58 (0.731/0.34, 33.77) | 0.35 (0.7127/0.38, 11.04) | 0.8 (0.889/0.14, 17.6) |
| Males vs. Females | Euploid | DS |
|---|---|---|
| Fold Change (p Value/t, df) | Fold Change (p Value/t, df) | |
| miR-99a | 2.12 0.053 (2.093, 16.05) | 5.12 0.040 (2.541, 6.702) |
| let7c | 1.06 0.876 (0.1594, 11.22) | 0.29 0.199 (1.453, 5.752) |
| miR-125b2 | 1.40 0.361 (0.9408, 15,82) | 0.65 0.281 (1.157; 7.848) |
| miR-155 | 0.20 (0.6452 *) | 0.08 (0.0031 *) |
| primiR-99a | 1.00 0.998 (0.001839, 17.28) | 1.15 0.759 (0.3164, 7.901) |
| Amniocytes (Bayes Factor) | Fetal Heart (Bayes Factor) | |
|---|---|---|
| 1 | mmu04510:Focal adhesion (19.41) | hsa04060:Cytokine–cytokine receptor interaction (21.21) |
| 2 | mmu04060:Cytokine–cytokine receptor interaction (15.71) | mmu04510:Focal adhesion (14.84) |
| 3 | rno04010:MAPK signaling pathway (14.13) | mmu04060:Cytokine–cytokine receptor interaction (14.77) |
| 4 | mmu04010:MAPK signaling pathway (12.92) | hsa04630:Jak-STAT signaling pathway (11) |
| 5 | hsa04060:Cytokine–cytokine receptor interaction (12.28) | mmu05010:Alzheimer’s disease (10.89) |
| 6 | mmu04210:Apoptosis (10.07) | rno04060:Cytokine–cytokine receptor interaction (10.41) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vizitiu, A.-C.; Stambouli, D.; Pavel, A.-G.; Muresan, M.-C.; Anastasiu, D.M.; Bejinar, C.; Alexa, A.; Marian, C.; Sirbu, I.O.; Sima, L. Mature miR-99a Upregulation in the Amniotic Fluid Samples from Female Fetus Down Syndrome Pregnancies: A Pilot Study. Medicina 2019, 55, 728. https://doi.org/10.3390/medicina55110728
Vizitiu A-C, Stambouli D, Pavel A-G, Muresan M-C, Anastasiu DM, Bejinar C, Alexa A, Marian C, Sirbu IO, Sima L. Mature miR-99a Upregulation in the Amniotic Fluid Samples from Female Fetus Down Syndrome Pregnancies: A Pilot Study. Medicina. 2019; 55(11):728. https://doi.org/10.3390/medicina55110728
Chicago/Turabian StyleVizitiu, Anda-Cornelia, Danae Stambouli, Anca-Gabriela Pavel, Maria-Cezara Muresan, Diana Maria Anastasiu, Cristina Bejinar, Anda Alexa, Catalin Marian, Ioan Ovidiu Sirbu, and Laurentiu Sima. 2019. "Mature miR-99a Upregulation in the Amniotic Fluid Samples from Female Fetus Down Syndrome Pregnancies: A Pilot Study" Medicina 55, no. 11: 728. https://doi.org/10.3390/medicina55110728