
Pathophysiological Implications of Interstitial Cajal-like Cells (ICC-like) in Uterus: A Comparative Study with Gastrointestinal ICCs
 , ,
, ,
Abstract
:1. Introduction
1.1. Historical Background of ICC
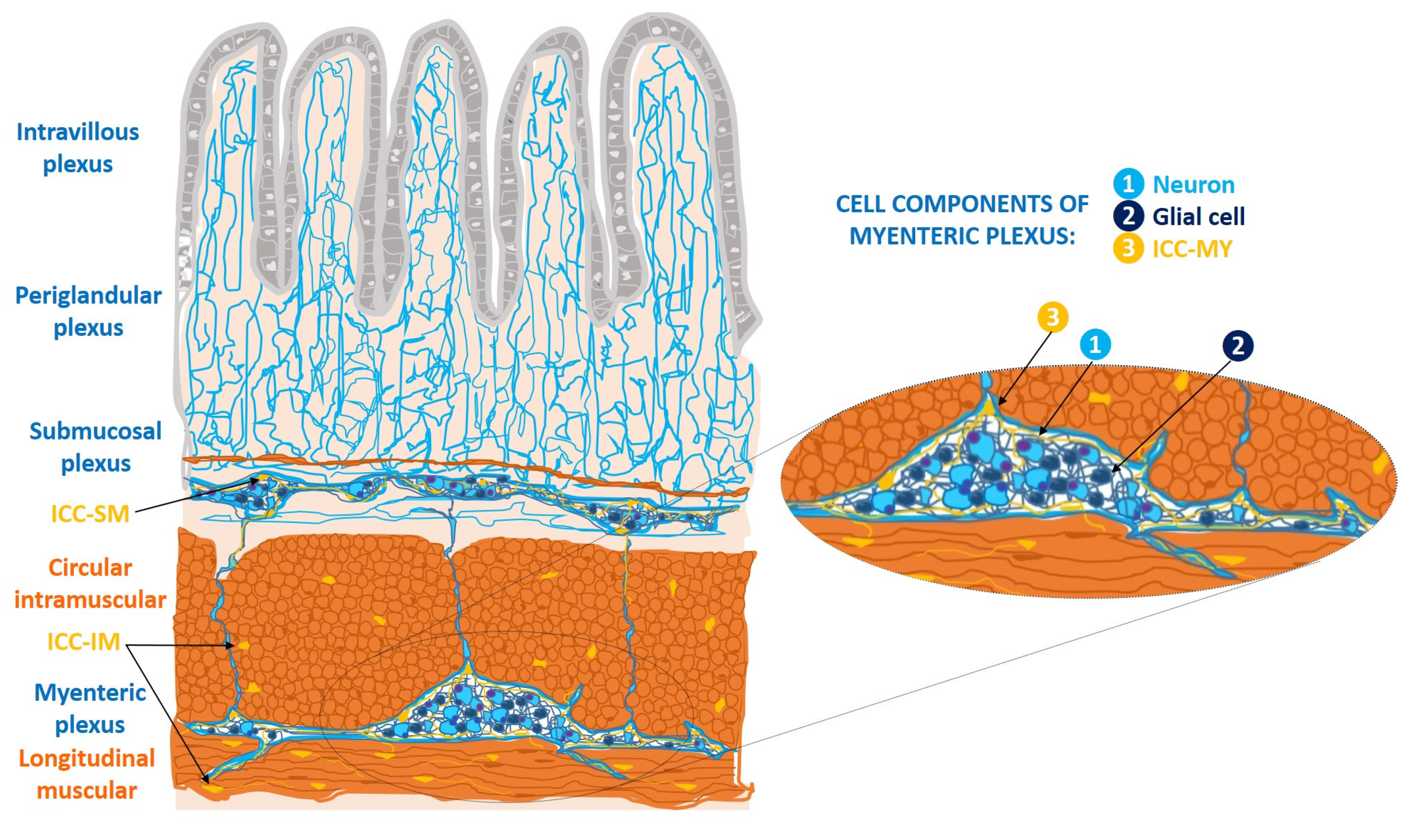
1.2. ICC at the Gastrointestinal Tract Level
1.2.1. Ultrastructural and Immunohistochemical Characteristics
1.2.2. Functions
1.2.3. Disfunctions
2. ICC-like Cells Outside the Digestive System
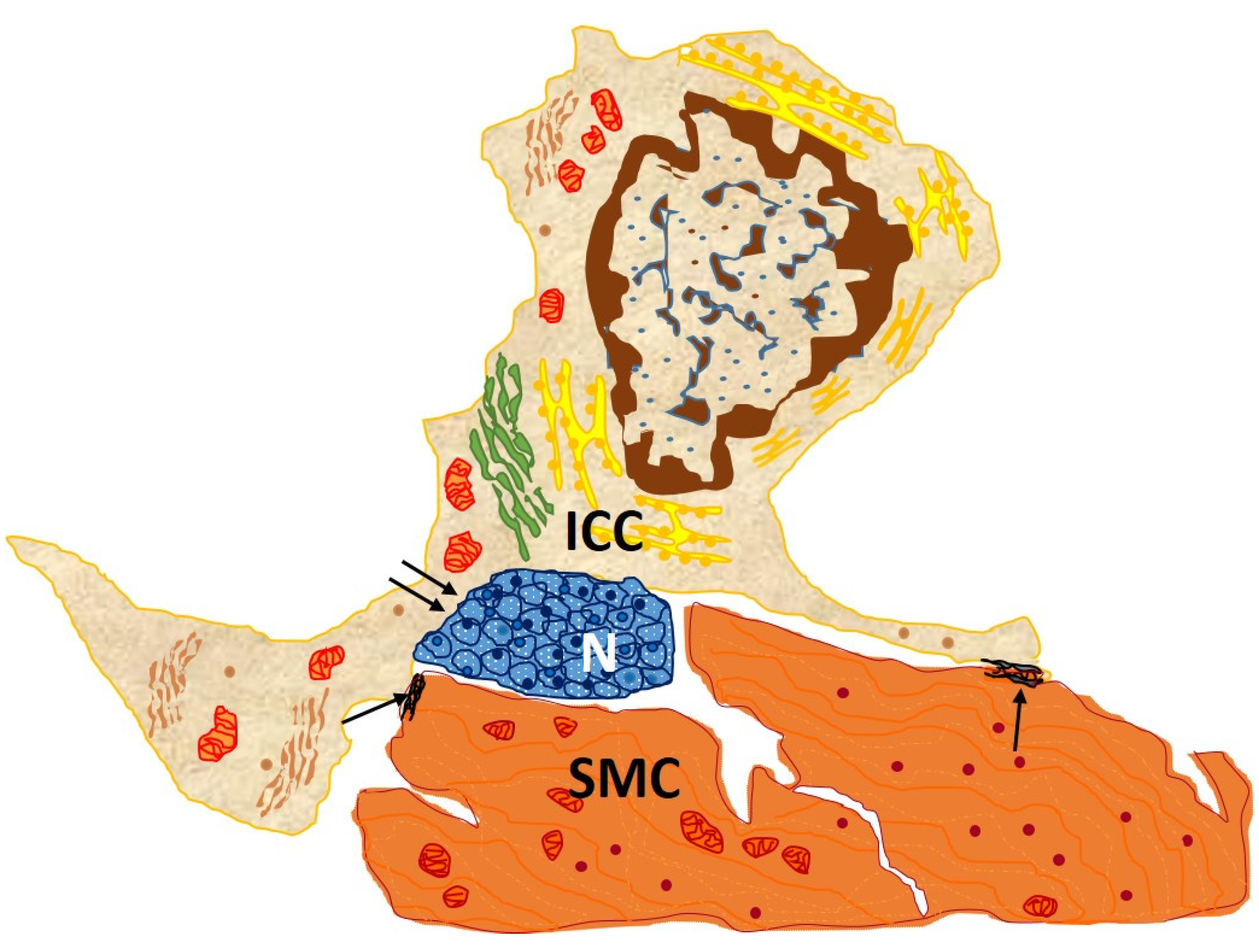
2.1. Ultraestructural Characteristics
- -
- They are of great length, tens to hundreds of μm, and considered one of the longest structures in the human body [27];
- -
- They vary in number from one to five, there frequently being two or three per cell. Thus, the three-dimensional appearance of ICC-like cells is that of a polyhedron with a variable number of vertices according to the number of extensions;
- -
- They are thin, <0.2 μm, and moniliform in appearance, with dilations along their length harboring Ca2+ channels and cytoplasmic organelles [27].
2.2. Immunohistochemical Characteristics
2.3. Functions of ICC-like Cells
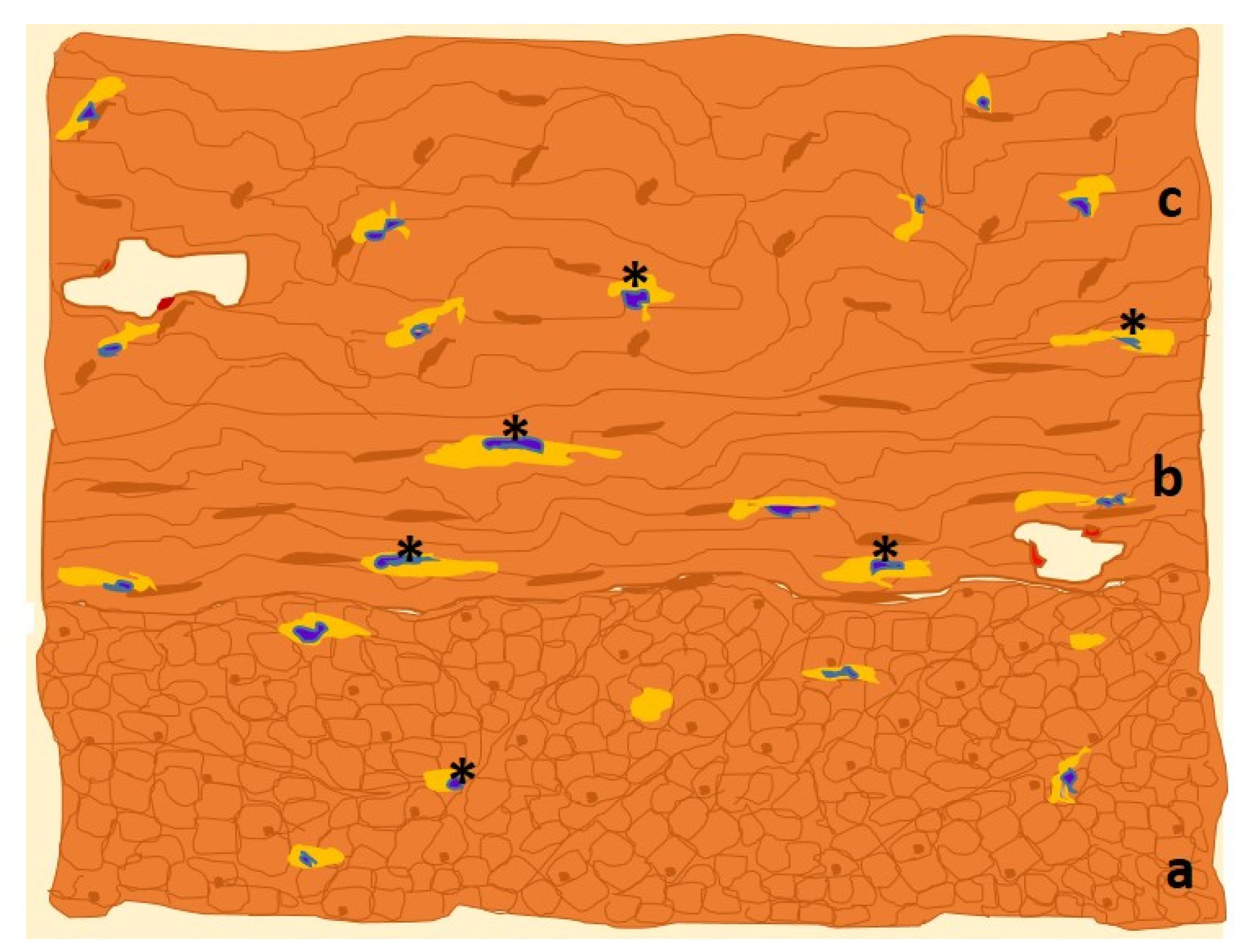
3. ICC-like Cells in Female Genital Tract
3.1. Ultraestructural and Immunohistochemical Features
3.2. Functions of Uterine ICC-like Cells
3.2.1. ICC-like Cells Implications in the Physiology of Pregnancy and Labor
3.2.2. Physiopathological Implications
- Gynecological disorders
- Obstetric disorders
4. Conclusions
5. Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cajal, S.R. Textura del Sistema Nervioso del Hombre y de los Vertebrados, 1st ed.; Imprenta y Librería de Nicolás Moya: Madrid, Spain, 1899–1904. [Google Scholar]
- Cajal, S.R.; Sala, C. Terminación de los nervios y tubos glandulares del páncreas de los vertebrados. Trab. Lab. Histol. Faculd. Med. Barcelona 1891, 28, 1–5. [Google Scholar]
- Cajal, S.R. Los Ganglios y Plexos Nerviosos del Intestino de los Mamíferos; Imprenta y Librería de Nicolás Moya: Madrid, Spain, 1893; pp. 5–37. [Google Scholar]
- Thuneberg, L. Interstitial cells of Cajal: Intestinal pacemaker cells? Adv. Anat. Embryol. Cell Biol. 1982, 71, 1–130. [Google Scholar]
- Garcia-Lopez, P.; Garcia-Marin, V.; Martínez-Murillo, R.; Freire, M. Updating old ideas and recent advances regarding the Interstitial Cells of Cajal. Brain Res. Rev. 2009, 61, 154–169. [Google Scholar] [CrossRef]
- Faussone-Pellegrini, M.S.; Thuneberg, L. Guide to the identification of interstitial cells of Cajal. Microsc. Res. Tech. 1999, 47, 248–266. [Google Scholar] [CrossRef]
- Gomez-Pinilla, P.J.; Gibbons, S.J.; Bardsley, M.R.; Lorincz, A.; Pozo, M.J.; Pasricha, P.J.; Van de Rijn, M.; West, R.B.; Sarr, M.G.; Kendrick, M.L.; et al. Ano1 is a selective marker of interstitial cells of cajal in the human and mouse gastrointestinal tract. Am. J. Physiol. 2009, 296, 1370–1381. [Google Scholar] [CrossRef]
- Varga, I.; Klein, M.; Urban, L.; Danihel, L., Jr.; Polak, S.; Danihel, L., Sr. Recently discovered interstitial cells “telocytes” as players in the pathogenesis of uterine leiomyomas. Med. Hypotheses 2018, 110, 64–67. [Google Scholar] [CrossRef]
- Hirst, G.; Edwards, F. Role of interstitial cells of Cajal in the control of gastric motility. J. Pharmacol. Sci. 2004, 96, 1–10. [Google Scholar] [CrossRef]
- Hirst, G.D.; Ward, S.M. Interstitial cells: Involvement in rhythmicity and neural control of gut smooth muscle. J. Physiol. 2003, 550, 337–346. [Google Scholar] [CrossRef]
- Sanders, K.M.; Ward, S.M.; Koh, S.D. Interstitial cells: Regulators of smooth muscle function. Physiol. Rev. 2014, 94, 859–907. [Google Scholar] [CrossRef]
- Takahashi, T. Pathophysiological significance of neuronal nitric oxide synthase in the gastrointestinal tract. J. Gastroenterol. 2003, 38, 421–430. [Google Scholar] [CrossRef]
- Popescu, L.M.; Faussone-Pellegrini, M.S. Telocytes—A case of serendipity: The winding way from Interstitial Cells of Cajal (ICC), via Interstitial Cajal-Like Cells (ICLC) to telocytes. J. Cell Mol. Med. 2010, 14, 729–740. [Google Scholar] [CrossRef] [PubMed]
- López-Pingarrón, L.; Almeida, H.; Soria-Aznar, M.; Reyes-Gonzales, M.C.; Rodríguez-Moratinos, A.B.; Muñoz-Hoyos, A.; García, J.J. Interstitial Cells of Cajal and Enteric Nervous System in Gastrointestinal and Neurological Pathology, Relation to Oxidative Stress. Curr. Issues Mol. Biol. 2023, 45, 3552–3572. [Google Scholar] [CrossRef] [PubMed]
- Gfroerer, S.; Rolle, U. Interstitial cells of Cajal in the normal human gut and in Hirschsprung disease. Pediatr. Surg. Int. 2013, 29, 889–897. [Google Scholar] [CrossRef] [PubMed]
- Knowles, C.H.; Farrugia, G. Gastrointestinal neuromuscular pathology in chronic constipation. Best Pract. Res. Clin. Gastroenterol. 2011, 25, 43–57. [Google Scholar] [CrossRef] [PubMed]
- Jabari, S.; da Silveira, A.B.M.; de Oliveira, E.C.; Quint, K.; Wirries, A.; Neuhuber, W.; Brehmer, A. Interstitial cells of Cajal: Crucial for the development of megacolon in human Chagas’ disease? Colorectal. Dis. 2013, 15, 592–598. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, I.B.; Guerini, J.C.; Gramática, L.; Piccinni, D.J.; Avila, R.E.; Spitale, L.S. Inmunohistochemistry Expression of Vimentin, CD 34 and c-Kit (CD117) in Gastrointestinal Stromal Tumors. Int. J. Morphol. 2010, 28, 453–459. [Google Scholar] [CrossRef]
- Eizaguirre-Zarza, B.; Burgos-Bretones, J.J.; Tumores, G.I.S.T. Revisión de la literature. Rev. Esp. Patol. 2006, 39, 209–218. [Google Scholar] [CrossRef]
- Chang, I.Y.; Glasgow, N.J.; Takayama, I.; Horiguchi, K.; Sanders, K.M.; Ward, S.M. Loss of interstitial cells of Cajal and development of electrical dysfunction in murine small bowel obstruction. J. Physiol. 2001, 536, 555–568. [Google Scholar] [CrossRef]
- Popescu, L.M.; Ciontea, S.M.; Cretoiu, D. Interstitial Cajal-like cells in human uterus and fallopian tube. Ann. N. Y Acad. Sci. 2007, 1101, 139–165. [Google Scholar] [CrossRef]
- Janas, P.; Kucybala, I.; Radon-Pokracka, M.; Huras, H. Telocytes in the female reproductive system: An overview of up-to-date knowledge. Adv. Clin. Exp. Med. 2018, 27, 559–565. [Google Scholar] [CrossRef]
- Wang, J.P.; Ding, G.F.; Wang, Q.Z. Interstitial cells of Cajal mediate excitatory sympathetic neurotransmission in guinea pig prostate. Cell Tissue Res. 2013, 352, 479–486. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Cao, Y.; Song, Y.; Huang, Q.; Wang, F.; Yang, W.; Yang, C. Telocytes in liver. Curr. Stem Cell Res. Ther. 2016, 11, 415–419. [Google Scholar] [CrossRef]
- Song, D.; Cretoiu, D.; Cretoiu, S.M.; Wang, X. Telocytes and lung disease. Histol. Histopathol. 2016, 31, 1303–1314. [Google Scholar] [CrossRef] [PubMed]
- Rusu, M.C.; Mirancea, N.; Mănoiu, V.S.; Vâlcu, M.; Nicolescu, M.I.; Păduraru, D. Skin telocytes. Ann. Anat. 2012, 194, 359–367. [Google Scholar] [CrossRef]
- Aleksandrovych, V.; Walocha, J.A.; Gil, K. Telocytes in female reproductive system (human and animal). J. Cell Mol. Med. 2016, 20, 994–1000. [Google Scholar] [CrossRef] [PubMed]
- Roatesi, I.; Radu, B.M.; Cretoiu, D.; Cretoiu, S.M. Uterine telocytes: A review of current knowledge. Reprod. Biol. 2015, 93, 1–13. [Google Scholar] [CrossRef]
- Cretoiu, D.; Cretoiu, S.M. Telocytes in the reproductive organs: Current understanding and future challenges. Semin. Cell Dev. Biol. 2016, 55, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Hutchings, G.; Williams, O.; Cretoiu, D.; Ciontea, S.M. Myometrial interstitial cells and the coordination of myometrial contractility. J. Cell Mol. Med. 2009, 13, 4268–4282. [Google Scholar] [CrossRef]
- Shmygol, A.; Blanks, A.M.; Bru-Mercier, G.; Gullam, J.E.; Thornton, S. Control of uterine Ca2+ by membrane voltage: Toward understanding the excitation-contraction coupling in human myometrium. Ann. N. Y. Acad. Sci. 2007, 1101, 97–109. [Google Scholar] [CrossRef]
- Hashitani, H.; Lang, R.J. Functions of ICC-like cells in the urinary tract and male genital organs. J. Cell Mol. Med. 2010, 14, 1199–1211. [Google Scholar] [CrossRef]
- Dixon, R.E.; Ramsey, K.H.; Schripsema, J.H.; Sanders, K.M.; Ward, S.M. Time-dependent disruption of oviduct pacemaker cells by chlamydia infection in mice. Biol. Reproduct. 2010, 83, 244–253. [Google Scholar] [CrossRef] [PubMed]
- Gartner, L.; Hiatt, J. Texto Atlas de Histología, 3rd ed.; McGraw Hill: México City, Mexico, 2008. [Google Scholar]
- Hatta, K.; Huang, M.L.; Weisel, R.D.; Li, R.K. Culture of rat endometrial telocytes. J. Cell Mol. Med. 2012, 16, 1392–1396. [Google Scholar] [CrossRef]
- Varga, I.; Urban, L.; Kajanová, M.; Polák, Š. Functional histology and possible clinical significance of recently discovered telocytes inside the female reproductive system. Arch. Gynecol. Obstet. 2016, 294, 417–422. [Google Scholar] [CrossRef] [PubMed]
- Aleksandrovych, V.; Pasternak, A.; Gil, K. Telocytes in the architecture of uterine fibroids. Folia Med. Cracov. 2019, 59, 33–44. [Google Scholar] [CrossRef]
- Young, R.C.; Zhang, P. Inhibition of in vitro contractions of human myometrium by mibefradil, a T-type calcium channel blocker: Support for a model using excitation-contraction coupling, and autocrine and paracrine signaling mechanisms. J. Soc. Gynecol. Investig. 2005, 12, 7–12. [Google Scholar] [CrossRef]
- Drife, J.; Magowan, B.A. Clinical Obstetrics and Gynaecology, 1st ed.; Elsevier: Amsterdam, The Netherlands, 2004. [Google Scholar]
- Pasqualini, J.R. Enzymes involved in the formation and transformation of steroid hormones in the fetal and placental compartments. J. Steroid Biochem. Mol. Biol. 2005, 97, 401–415. [Google Scholar] [CrossRef] [PubMed]
- Casanova, R.; Chuang, A.; Goepfert, A.R.; Hueppchen, N.A.; Weiss, P.M. American College of Obstetricians and Gynecologists. In Beckmann and Ling's Obstetrics and Gynecology, 8th ed.; Wolters Kluwer: Philadelphia, PA, USA, 2019. [Google Scholar]
- Young, R.C.; Zhang, P. Tissue-level bioelectrical signals as the trigger for uterine contractions in human pregnancy. J. Soc. Gynecol. Investig. 2004, 11, 478–482. [Google Scholar] [CrossRef]
- Salama, N. Immunohistochemical characterization of telocytes in rat uterus in different reproductive states. Egypt. J. Histol. 2013, 36, 185–194. [Google Scholar] [CrossRef]
- Cretoiu, S.M.; Radu, B.M.; Banciu, A.; Banciu, D.D.; Cretoiu, D.; Ceafalan, L.C.; Popescu, L.M. Isolated human uterine telocytes: Immunocytochemistry and electrophysiology of T-type calcium channels. Histochem. Cell Biol. 2015, 143, 83–94. [Google Scholar] [CrossRef]
- Rosenbaum, S.T.; Svalø, J.; Nielsen, K.; Larsen, T.; Jørgensen, J.C.; Bouchelouche, P. Immunolocalization and expression of small-conductance calcium-activated potassium channels in human myometrium. J. Cell Mol. Med. 2012, 16, 3001–3008. [Google Scholar] [CrossRef]
- Cretoiu, S.M.; Simionescu, A.A.; Caravia, L.; Curici, A.; Cretoiu, D.; Popescu, L.M. Complex effects of imatinib on spontaneous and oxytocin-induced contractions in human non-pregnant myometrium. Acta Physiol. Hung. 2011, 98, 329–338. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Flores, L.; Gutiérrez, R.; García, M.P.; Sáez, F.J.; Aparicio, F.; Díaz Flores, L., Jr.; Madrid, J.F. Uptake and intracytoplasmic storage of pigmented particles by human CD34+ stromal cells/ telocytes: Endocytic property of telocytes. J. Cell Mol. Med. 2014, 18, 2478–2487. [Google Scholar] [CrossRef] [PubMed]
- Grinnell, F.; Head, J.R.; Hoffpauir, J. Fibronectin and cell shape in vivo: Studies on the endometrium during pregnancy. J. Cell Biol. 1982, 94, 597–606. [Google Scholar] [CrossRef]
- Walocha, J.; Miodoński, A.J.; Szczepański, W.; Skrzat, J.; Stachura, J. Two types of vascularisation of intramural uterine leiomyomata revealed by corrosion casting and immunohistochemical study. Folia Morphol. 2004, 63, 37–41. [Google Scholar]
- Aleksandrovych, V.; Gil, A.; Wrona, A. Sex steroid hormone receptors of telocytes-potential key role in leiomyoma development. Folia Med. Cracov. 2020, 60, 81–95. [Google Scholar] [CrossRef]
- Aleksandrovych, V.; Kurnik-Łucka, M.; Bereza, T.; Bialas, M.; Pasternak, A.; Cretoiu, D.; Walocha, J.A.; Gil, K. The Autonomic innervation and uterine telocyte interplay in leiomyoma formation. Cell Transplant. 2019, 28, 619–629. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Bai, C.; Wang, X. Telocyte morphologies and potential roles in diseases. J. Cell Physiol. 2012, 227, 2311–2317. [Google Scholar] [CrossRef]
- Chi, C.; Jiang, X.J.; Su, L.; Shen, Z.J.; Yang, X.J. In vitro morphology, viability and cytokine secretion of uterine telocyte-activated mouse peritoneal macrophages. J. Cell Mol. Med. 2015, 19, 2741–2750. [Google Scholar] [CrossRef]
- Pérez-Martínez, S.; Viggiano, M.; Franchi, A.; Herrero, M.; Ortiz, M.; Gimeno, M.; Villalón, M. Effect of nitric oxide synthase inhibitors on ovum transport and oviductal smooth muscle activity in the rat oviduct. J. Reprod. Fertil. 2000, 118, 111–117. [Google Scholar] [CrossRef]
- Vercellini, P.; Viganò, P.; Somigliana, E.; Fedele, L. Endometriosis: Pathogenesis and treatment. Nat. Rev. Endocrinol. 2014, 10, 261–275. [Google Scholar] [CrossRef]
- Martins, A.F.; Neto, A.C.; Rodrigues, A.R.; Oliveira, S.M.; Sousa-Mendes, C.; Leite-Moreira, A.; Gouveia, A.M.; Almeida, H.; Neves, D. Metformin Prevents Endothelial Dysfunction in Endometriosis through Downregulation of ET-1 and Upregulation of eNOS. Biomedicines 2022, 10, 2782. [Google Scholar] [CrossRef] [PubMed]
- Neto, A.C.; Santos-Pereira, M.; Abreu-Mendes, P.; Neves, D.; Almeida, H.; Cruz, F.; Charrua, A. The Unmet Needs for Studying Chronic Pelvic/Visceral Pain Using Animal Models. Biomedicines 2023, 11, 696. [Google Scholar] [CrossRef]
- Garfield, R.E.; Sims, S.; Daniel, E.E. Gap junctions: Their presence and necessity in myometrium during parturition. Science 1977, 198, 958–960. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Felix, J.C.; Lee, J.L.; Tan, P.Y.; Tourgeman, D.E.; O’Meara, A.T.; Amezcua, C.A. The proto-oncogene c-kit is expressed in leiomyosarcomas of the uterus. Gynecol. Oncol. 2003, 90, 402–406. [Google Scholar] [CrossRef] [PubMed]
- Popescu, L.M.; Vidulescu, C.; Curici, A.; Caravia, L.; Simionescu, A.A.; Ciontea, S.M.; Simion, S. Imatinib inhibits spontaneous rhythmic contractions of human uterus and intestine. Eur. J. Pharmacol. 2006, 546, 177–181. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Cytological Issues | Enteric ICC | Myometrial ICC-like Cells |
|---|---|---|
| Shape: | ||
| cell | Oval or spindle | Spindle or stellate |
| nucleus | Oval, mostly euchromatic | Oval, heterochromatic |
| Mitochondria | +++ | ++ |
| ER 1: | ||
| smooth | ++ | + |
| rough | + | + |
| Golgi complex | + | + |
| Filaments: | ||
| thin | + | + |
| intermediate | ++ | + |
| Microtubules | + | + |
| Calcium releasing units | + | |
| Caveolae | + | + |
| Basal lamina | +- | |
| Gap junctions | + | + |
| Intercellular contacts: | ||
| nerve endings | ++ | + |
| blood vessels | + | |
| immune cells | +++ | |
| muscular cells | + | + |
| other interstitial cells | + | + |
| Immuno-histochemical markers | c-Kit, Ano-1 | c-Kit, CD34, PDGFR-α, smooth muscle actin (α-SMA), CD44, vimentin, Sca-1, connexin 43 |
| Non-Pregnant Uterus | Pregnant Uterus | Postpartum Uterus | |
|---|---|---|---|
| Endometrial ICC-like cells | + | ++ | + |
| Myometrial ICC-like cells | ++ | + | +++ |
| Cav3.1 | ++ | ++ | |
| Cav3.2 | + | ++ | |
| SK3 | ++ | + |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
López-Pingarrón, L.; Almeida, H.; Pereboom-Maicas, D.; García, J.J. Pathophysiological Implications of Interstitial Cajal-like Cells (ICC-like) in Uterus: A Comparative Study with Gastrointestinal ICCs. Curr. Issues Mol. Biol. 2023, 45, 7557-7571. https://doi.org/10.3390/cimb45090476
López-Pingarrón L, Almeida H, Pereboom-Maicas D, García JJ. Pathophysiological Implications of Interstitial Cajal-like Cells (ICC-like) in Uterus: A Comparative Study with Gastrointestinal ICCs. Current Issues in Molecular Biology. 2023; 45(9):7557-7571. https://doi.org/10.3390/cimb45090476
Chicago/Turabian StyleLópez-Pingarrón, Laura, Henrique Almeida, Desirée Pereboom-Maicas, and Joaquín J. García. 2023. "Pathophysiological Implications of Interstitial Cajal-like Cells (ICC-like) in Uterus: A Comparative Study with Gastrointestinal ICCs" Current Issues in Molecular Biology 45, no. 9: 7557-7571. https://doi.org/10.3390/cimb45090476