
Oxytocin’s Regulation of Thermogenesis May Be the Link to Prader–Willi Syndrome
Abstract
:1. Introduction
2. An Update on Prader–Willi Syndrome
2.1. Phenotype and Features of Prader–Willi Syndrome
2.2. Pharmacological Intervention with Oxytocin in Prader–Willi Syndrome
2.3. The Special Case of the Thermosensory and Pain Perception System in Prader–Willi Syndrome
3. Oxytocin’s Regulation of Thermogenesis
3.1. Oxytocin’s Regulation of Thermogenesis in Genetic Studies
3.1.1. Oxytocin in Bone, Brain and Brown Adipose Tissue after Cold Stress
3.1.2. Oxytocin in Myosin after Cold Stress
3.1.3. Oxytocin and Transient-Receptor-Potential-Vanilloid-1 (TRPV1) and Pain Perception after Cold Stress
3.2. Oxytocin’s Regulation of Thermogenesis In Ex Vivo Studies
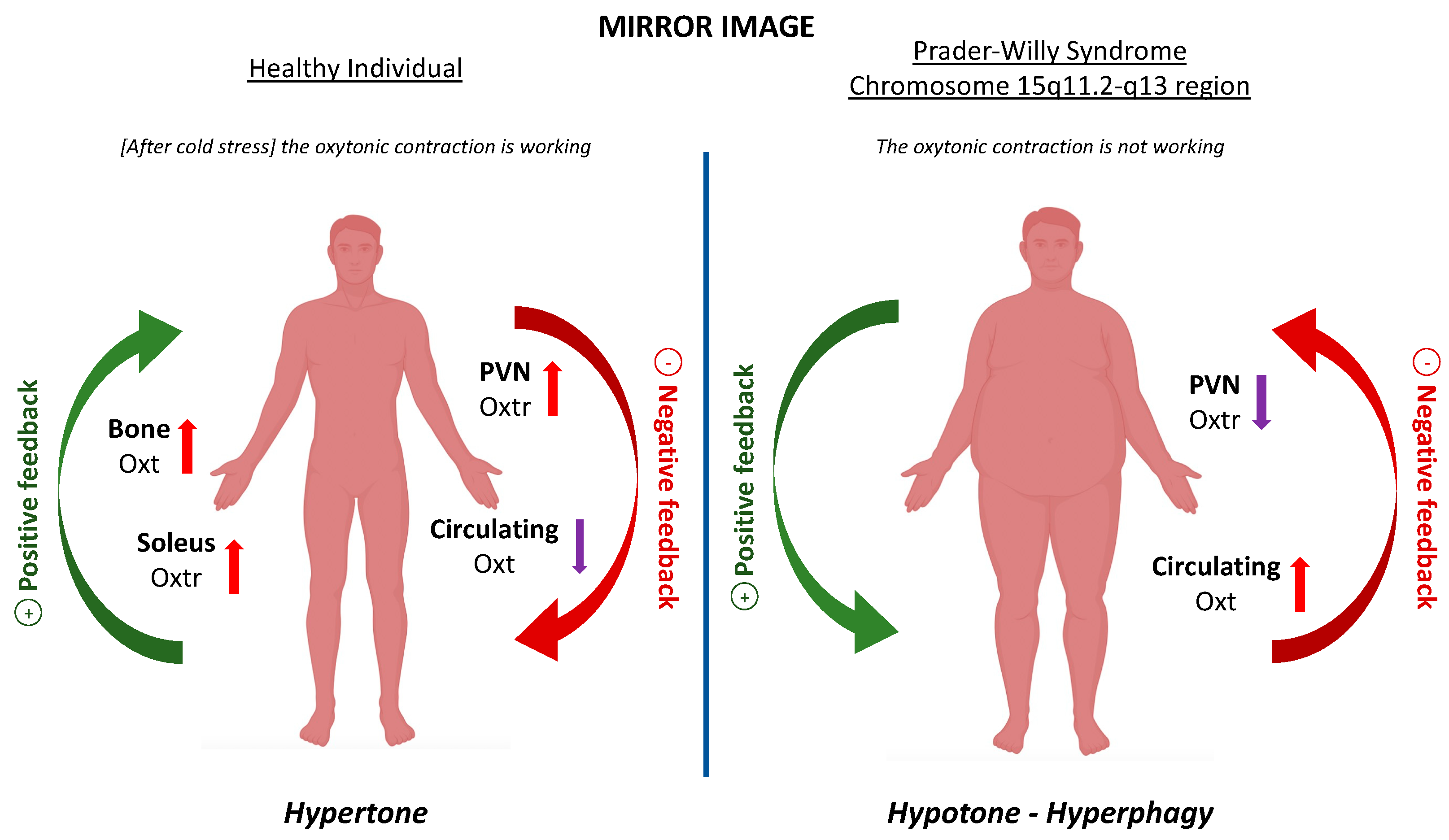
4. The Link with Prader–Willi Syndrome
5. Future Directions
6. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Heseding, H.M.; Jahn, K.; Eberlein, C.K.; Wieting, J.; Maier, H.B.; Proskynitopoulos, P.J.; Glahn, A.; Bleich, S.; Frieling, H.; Deest, M. Distinct promoter regions of the oxytocin receptor gene are hypomethylated in Prader-Willi syndrome and in Prader-Willi syndrome associated psychosis. Transl. Psychiatry 2022, 12, 246. [Google Scholar] [CrossRef]
- Cassidy, S.B.; Schwartz, S.; Miller, J.L.; Driscoll, D.J. Prader-Willi syndrome. Genet. Med. 2012, 14, 10–26. [Google Scholar] [CrossRef] [Green Version]
- Donaldson, M.D.; Chu, C.E.; Cooke, A.; Wilson, A.; Greene, S.A.; Stephenson, J.B. The Prader-Willi syndrome. Arch. Dis. Child. 1994, 70, 58–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oztan, O.; Zyga, O.; Stafford, D.E.J.; Parker, K.J. Linking oxytocin and arginine vasopressin signaling abnormalities to social behavior impairments in Prader-Willi syndrome. Neurosci. Biobehav. Rev. 2022, 142, 104870. [Google Scholar] [CrossRef] [PubMed]
- Camerino, C. Low sympathetic tone and obese phenotype in oxytocin-deficient mice. Obesity 2009, 17, 980–984. [Google Scholar] [CrossRef]
- Takayanagi, Y.; Kasahara, Y.; Onaka, T.; Takahashi, N.; Kawada, T.; Nishimori, K. Oxytocin receptor-deficient mice developed late-onset obesity. NeuroReport 2008, 19, 951–955. [Google Scholar] [CrossRef] [PubMed]
- Camerino, C. The Long Way of Oxytocin from the Uterus to the Heart in 70 Years from Its Discovery. Int. J. Mol. Sci. 2023, 24, 2556. [Google Scholar] [CrossRef]
- Camerino, C.; Conte, E.; Cannone, M.; Caloiero, R.; Fonzino, A.; Tricarico, D. Nerve Growth Factor, Brain-Derived Neurotrophic Factor and Osteocalcin Gene Relationship in Energy Regulation, Bone Homeostasis and Reproductive Organs Analyzed by mRNA Quantitative Evaluation and Linear Correlation Analysis. Front. Physiol. 2016, 7, 456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camerino, C.; Conte, E.; Caloiero, R.; Fonzino, A.; Carratù, M.; Lograno, M.D.; Tricarico, D. Evaluation of Short and Long Term Cold Stress Challenge of Nerve Grow Factor, Brain-Derived Neurotrophic Factor, Osteocalcin and Oxytocin mRNA Expression in BAT, Brain, Bone and Reproductive Tissue of Male Mice Using Real-Time PCR and Linear Correlation Analysis. Front. Physiol. 2018, 8, 1101. [Google Scholar] [CrossRef] [Green Version]
- Camerino, C.; Conte, E.; Carratù, M.R.; Fonzino, A.; Lograno, M.D.; Tricarico, D. Oxytocin/Osteocalcin/IL-6 and NGF/BDNF mRNA Levels in Response to Cold Stress Challenge in Mice: Possible Oxytonic Brain-Bone-Muscle-Interaction. Front. Physiol. 2019, 10, 1437. [Google Scholar] [CrossRef]
- Conte, E.; Romano, A.; De Bellis, M.; de Ceglia, M.; Rosaria Carratù, M.; Gaetani, S.; Maqoud, F.; Tricarico, D.; Camerino, C. Oxtr/TRPV1 expression and acclimation of skeletal muscle to cold-stress in male mice. J. Endocrinol. 2021, 249, 135–148. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Zhang, R.; Wu, R.; Gu, Y.; Lu, Y. The effects of oxytocin to rectify metabolic dysfunction in obese mice are associated with increased thermogenesis. Mol. Cell. Endocrinol. 2020, 514, 110903. [Google Scholar] [CrossRef] [PubMed]
- Bittel, D.C.; Kibiryeva, N.; Sell, S.M.; Strong, T.V.; Butler, M.G. Whole genome microarray analysis of gene expression in Prader-Willi syndrome. Am. J. Med. Genet. A 2007, 143A, 430–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zayan, U.; Caccialupi Da Prato, L.; Muscatelli, F.; Matarazzo, V. Modulation of the thermosensory system by oxytocin. Front. Mol. Neurosci. 2022, 15, 1075305. [Google Scholar] [CrossRef] [PubMed]
- Duerden, E.G.; Taylor, M.J.; Lee, M.; McGrath, P.A.; Davis, K.D.; Roberts, S.W. Decreased sensitivity to thermal stimuli in adolescents with autism spectrum disorder: Relation to symptomatology and cognitive ability. J. Pain 2015, 16, 463–471. [Google Scholar] [CrossRef] [PubMed]
- Fründt, O.; Grashorn, W.; Schöttle, D.; Peiker, I.; David, N.; Engel, A.K.; Forkmann, K.; Wrobel, N.; Münchau, A.; Bingel, U. Quantitative Sensory Testing in adults with Autism Spectrum Disorders. J. Autism Dev. Disord. 2017, 47, 1183–1192. [Google Scholar] [CrossRef]
- Chien, Y.-L.; Chao, C.-C.; Wu, S.-W.; Hsueh, H.-W.; Chiu, Y.-N.; Tsai, W.-C.; Gau, S.S.-F.; Hsieh, S.-T. Small fiber pathology in autism and clinical implications. Neurology 2020, 95, e2697–e2706. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, T.; Iwabuchi, H.; Oishi, M. Accidental hypothermia in an infant with Prader-Willi syndrome. Eur. J. Pediatr. 2003, 162, 550–551. [Google Scholar] [CrossRef] [PubMed]
- McVea, S.; Thompson, A.J.; Abid, N.; Richardson, J. Thermal dysregulation in Prader-Willi syndrome: A potentially fatal complication in adolescence, not just in infancy. BMJ Case Rep. 2016, 2016, bcr2016215344. [Google Scholar] [CrossRef]
- Althammer, F.; Muscatelli, F.; Grinevich, V.; Schaaf, C.P. Oxytocin-based therapies for treatment of Prader-Willi and Schaaf-Yang syndromes: Evidence, disappointments, and future research strategies. Transl. Psychiatry 2022, 12, 318. [Google Scholar] [CrossRef]
- Da Prato, L.C.; Zayan, U.; Abdallah, D.; Point, V.; Schaller, F.; Pallesi-Pocachard, E.; Montheil, A.; Canaan, S.; Gaiarsa, J.-L.; Muscatelli, F.; et al. Early life oxytocin treatment improves thermo-sensory reactivity and maternal behavior in neonates lacking the autism-associated gene Magel2. Neuropsychopharmacology 2022, 47, 1901–1912. [Google Scholar] [CrossRef] [PubMed]
- MacIver, N.J. Oxytocin Treatment May Improve Infant Feeding and Social Skills in Prader-Willi Syndrome. Pediatrics 2017, 139, e20163833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaaf, C.P.; Gonzalez-Garay, M.L.; Xia, F.; Potocki, L.; Gripp, K.W.; Zhang, B.; Peters, B.A.; McElwain, M.A.; Drmanac, R.; Beaudet, A.L.; et al. Truncating mutations of MAGEL2 cause Prader-Willi phenotypes and autism. Nat. Genet. 2013, 45, 1405–1408. [Google Scholar] [CrossRef] [Green Version]
- Rice, L.J.; Einfeld, S.L.; Hu, N.; Carter, C.S. A review of clinical trials of oxytocin in Prader-Willi syndrome. Curr. Opin. Psychiatry 2018, 31, 123–127. [Google Scholar] [CrossRef]
- Schaller, F.; Watrin, F.; Sturny, R.; Massacrier, A.; Szepetowski, P.; Muscatelli, F. A single postnatal injection of oxytocin rescues the lethal feeding behaviour in mouse newborns deficient for the imprinted Magel2 gene. Hum. Mol. Genet. 2010, 19, 4895–4905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, J.L.; Lynn, C.H.; Driscoll, D.C.; Goldstone, A.P.; Gold, J.-A.; Kimonis, V.; Dykens, E.; Butler, M.G.; Shuster, J.J.; Driscoll, D.J. Nutritional phases in Prader-Willi syndrome. Am. J. Med. Genet. A 2011, 155A, 1040–1049. [Google Scholar] [CrossRef] [Green Version]
- Miller, J.L.; Tamura, R.; Butler, M.G.; Kimonis, V.; Sulsona, C.; Gold, J.-A.; Driscoll, D.J. Oxytocin treatment in children with Prader-Willi syndrome: A double-blind, placebo-controlled, crossover study. Am. J. Med. Genet. A 2017, 173, 1243–1250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butler, M.G.; Hossain, W.; Sulsona, C.; Driscoll, D.J.; Manzardo, A.M. Increased plasma chemokine levels in children with Prader-Willi syndrome. Am. J. Med. Genet. A 2015, 167A, 563–571. [Google Scholar] [CrossRef] [PubMed]
- Johnson, L.; Manzardo, A.M.; Miller, J.L.; Driscoll, D.J.; Butler, M.G. Elevated plasma oxytocin levels in children with Prader-Willi syndrome compared with healthy unrelated siblings. Am. J. Med. Genet. A 2016, 170, 594–601. [Google Scholar] [CrossRef] [PubMed]
- Qian, Y.; Xia, F.; Zuo, Y.; Zhong, M.; Yang, L.; Jiang, Y.; Zou, C. Do patients with Prader-Willi syndrome have favorable glucose metabolism? Orphanet J. Rare Dis. 2022, 17, 187. [Google Scholar] [CrossRef]
- Shapira, N.A.; Lessig, M.C.; He, A.G.; James, G.A.; Driscoll, D.J.; Liu, Y. Satiety dysfunction in Prader-Willi syndrome demonstrated by fMRI. J. Neurol. Neurosurg. Psychiatry 2005, 76, 260–262. [Google Scholar] [CrossRef]
- Einfeld, S.L.; Smith, E.; McGregor, I.S.; Steinbeck, K.; Taffe, J.; Rice, L.J.; Horstead, S.K.; Rogers, N.; Hodge, M.A.; Guastella, A.J. A double-blind randomized controlled trial of oxytocin nasal spray in Prader Willi syndrome. Am. J. Med. Genet. A 2014, 164A, 2232–2239. [Google Scholar] [CrossRef] [PubMed]
- Damen, L.; Grootjen, L.N.; Donze, S.H.; Juriaans, A.F.; de Graaff, L.C.G.; van der Velden, J.A.E.M.; Hokken-Koelega, A.C.S. Three years of growth hormone treatment in young adults with Prader-Willi Syndrome previously treated with growth hormone in childhood: Effects on glucose homeostasis and metabolic syndrome. Clin. Endocrinol. 2020, 93, 439–448. [Google Scholar] [CrossRef] [PubMed]
- Hollander, E.; Levine, K.G.; Ferretti, C.J.; Freeman, K.; Doernberg, E.; Desilva, N.; Taylor, B.P. Intranasal oxytocin versus placebo for hyperphagia and repetitive behaviors in children with Prader-Willi Syndrome: A randomized controlled pilot trial. J. Psychiatr. Res. 2021, 137, 643–651. [Google Scholar] [CrossRef]
- Dykens, E.M.; Miller, J.; Angulo, M.; Roof, E.; Reidy, M.; Hatoum, H.T.; Willey, R.; Bolton, G.; Korner, P. Intranasal carbetocin reduces hyperphagia in individuals with Prader-Willi syndrome. JCI Insight 2018, 3, e98333. [Google Scholar] [CrossRef]
- Diene, G.; Angulo, M.; Hale, P.M.; Jepsen, C.H.; Hofman, P.L.; Hokken-Koelega, A.; Ramesh, C.; Turan, S.; Tauber, M. Liraglutide for Weight Management in Children and Adolescents With Prader-Willi Syndrome and Obesity. J. Clin. Endocrinol. Metab. 2022, 108, 4–12. [Google Scholar] [CrossRef] [PubMed]
- Tauber, M.; Boulanouar, K.; Diene, G.; Çabal-Berthoumieu, S.; Ehlinger, V.; Fichaux-Bourin, P.; Molinas, C.; Faye, S.; Valette, M.; Pourrinet, J.; et al. The Use of Oxytocin to Improve Feeding and Social Skills in Infants With Prader-Willi Syndrome. Pediatrics 2017, 139, e20162976. [Google Scholar] [CrossRef] [Green Version]
- Grinevich, V.; Stoop, R. Interplay between Oxytocin and Sensory Systems in the Orchestration of Socio-Emotional Behaviors. Neuron 2018, 99, 887–904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuppens, R.J.; Delhanty, P.J.D.; Huisman, T.M.; van der Lely, A.J.; Hokken-Koelega, A.C.S. Acylated and unacylated ghrelin during OGTT in Prader-Willi syndrome: Support for normal response to food intake. Clin. Endocrinol. 2016, 85, 488–494. [Google Scholar] [CrossRef] [PubMed]
- Tauber, M.; Mantoulan, C.; Copet, P.; Jauregui, J.; Demeer, G.; Diene, G.; Rogé, B.; Laurier, V.; Ehlinger, V.; Arnaud, C.; et al. Oxytocin may be useful to increase trust in others and decrease disruptive behaviours in patients with Prader-Willi syndrome: A randomised placebo-controlled trial in 24 patients. Orphanet J. Rare Dis. 2011, 6, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ince, E.; Ciftçi, E.; Tekin, M.; Kendirli, T.; Tutar, E.; Dalgiç, N.; Oncel, S.; Dogru, U. Characteristics of hyperthermia and its complications in patients with Prader Willi syndrome. Pediatr. Int. 2005, 47, 550–553. [Google Scholar] [CrossRef]
- Priano, L.; Miscio, G.; Grugni, G.; Milano, E.; Baudo, S.; Sellitti, L.; Picconi, R.; Mauro, A. On the origin of sensory impairment and altered pain perception in Prader-Willi syndrome: A neurophysiological study. Eur. J. Pain 2009, 13, 829–835. [Google Scholar] [CrossRef] [PubMed]
- Trayhurn, P. Origins and early development of the concept that brown adipose tissue thermogenesis is linked to energy balance and obesity. Biochimie 2017, 134, 62–70. [Google Scholar] [CrossRef] [PubMed]
- Kasahara, Y.; Sato, K.; Takayanagi, Y.; Mizukami, H.; Ozawa, K.; Hidema, S.; So, K.-H.; Kawada, T.; Inoue, N.; Ikeda, I.; et al. Oxytocin receptor in the hypothalamus is sufficient to rescue normal thermoregulatory function in male oxytocin receptor knockout mice. Endocrinology 2013, 154, 4305–4315. [Google Scholar] [CrossRef] [Green Version]
- Mosialou, I.; Shikhel, S.; Liu, J.-M.; Maurizi, A.; Luo, N.; He, Z.; Huang, Y.; Zong, H.; Friedman, R.A.; Barasch, J.; et al. MC4R-dependent suppression of appetite by bone-derived lipocalin 2. Nature 2017, 543, 385–390. [Google Scholar] [CrossRef]
- Oumi, T.; Ukena, K.; Matsushima, O.; Ikeda, T.; Fujita, T.; Minakata, H.; Nomoto, K. Annetocin: An oxytocin-related peptide isolated from the earthworm, Eisenia foetida. Biochem. Biophys. Res. Commun. 1994, 198, 393–399. [Google Scholar] [CrossRef] [PubMed]
- Mizunoya, W.; Iwamoto, Y.; Sato, Y.; Tatsumi, R.; Ikeuchi, Y. Cold exposure increases slow-type myosin heavy chain 1 (MyHC1) composition of soleus muscle in rats. Anim. Sci. J. 2014, 85, 293–304. [Google Scholar] [CrossRef]
- Palmer, B.F.; Clegg, D.J. Non-shivering thermogenesis as a mechanism to facilitate sustainable weight loss. Obes. Rev. 2017, 18, 819–831. [Google Scholar] [CrossRef] [PubMed]
- Blondin, D.P.; Haman, F. Shivering and nonshivering thermogenesis in skeletal muscles. Handb. Clin. Neurol. 2018, 156, 153–173. [Google Scholar] [CrossRef]
- Camerino, C. The New Frontier in Oxytocin Physiology: The Oxytonic Contraction. Int. J. Mol. Sci. 2020, 21, 5144. [Google Scholar] [CrossRef] [PubMed]
- Kasahara, Y.; Takayanagi, Y.; Kawada, T.; Itoi, K.; Nishimori, K. Impaired thermoregulatory ability of oxytocin-deficient mice during cold-exposure. Biosci. Biotechnol. Biochem. 2007, 71, 3122–3126. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, Y.; Ueda, T.; Irahara, M.; Aono, T. Releases of oxytocin and prolactin during breast massage and suckling in puerperal women. Eur. J. Obstet. Gynecol. Reprod. Biol. 1994, 53, 17–20. [Google Scholar] [CrossRef] [PubMed]
- Pette, D.; Staron, R.S. Cellular and molecular diversities of mammalian skeletal muscle fibers. In Reviews of Physiology, Biochemistry and Pharmacology; Springer: Berlin/Heidelberg, Germany, 1990; Volume 116, pp. 1–76. [Google Scholar] [CrossRef]
- Termin, A.; Staron, R.S.; Pette, D. Myosin heavy chain isoforms in histochemically defined fiber types of rat muscle. Histochemistry 1989, 92, 453–457. [Google Scholar] [CrossRef]
- Rivero, J.L.; Talmadge, R.J.; Edgerton, V.R. Fibre size and metabolic properties of myosin heavy chain-based fibre types in rat skeletal muscle. J. Muscle Res. Cell Motil. 1998, 19, 733–742. [Google Scholar] [CrossRef] [PubMed]
- Rivero, J.L.; Talmadge, R.J.; Edgerton, V.R. Interrelationships of myofibrillar ATPase activity and metabolic properties of myosin heavy chain-based fibre types in rat skeletal muscle. Histochem. Cell Biol. 1999, 111, 277–287. [Google Scholar] [CrossRef] [PubMed]
- De Jager, N.; Hudson, N.J.; Reverter, A.; Wang, Y.-H.; Nagaraj, S.H.; Cafe, L.M.; Greenwood, P.L.; Barnard, R.T.; Kongsuwan, K.P.; Dalrymple, B.P. Chronic exposure to anabolic steroids induces the muscle expression of oxytocin and a more than fiftyfold increase in circulating oxytocin in cattle. Physiol. Genom. 2011, 43, 467–478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elabd, C.; Basillais, A.; Beaupied, H.; Breuil, V.; Wagner, N.; Scheideler, M.; Zaragosi, L.-E.; Massiéra, F.; Lemichez, E.; Trajanoski, Z.; et al. Oxytocin controls differentiation of human mesenchymal stem cells and reverses osteoporosis. Stem Cells 2008, 26, 2399–2407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arrowsmith, S.; Quenby, S.; Weeks, A.; Burdyga, T.; Wray, S. Poor spontaneous and oxytocin-stimulated contractility in human myometrium from postdates pregnancies. PLoS ONE 2012, 7, e36787. [Google Scholar] [CrossRef] [PubMed]
- Nersesyan, Y.; Demirkhanyan, L.; Cabezas-Bratesco, D.; Oakes, V.; Kusuda, R.; Dawson, T.; Sun, X.; Cao, C.; Cohen, A.M.; Chelluboina, B.; et al. Oxytocin Modulates Nociception as an Agonist of Pain-Sensing TRPV1. Cell Rep. 2017, 21, 1681–1691. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Hernandez, A.; Charlet, A. Oxytocin, GABA, and TRPV1, the Analgesic Triad? Front. Mol. Neurosci. 2018, 11, 398. [Google Scholar] [CrossRef] [Green Version]
- Fukushima, A.; Kataoka, N.; Nakamura, K. An oxytocinergic neural pathway that stimulates thermogenic and cardiac sympathetic outflow. Cell Rep. 2022, 40, 111380. [Google Scholar] [CrossRef]
- Bittel, D.C.; Butler, M.G. Prader-Willi syndrome: Clinical genetics, cytogenetics and molecular biology. Expert Rev. Mol. Med. 2005, 7, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Swaab, D.F. Prader-Willi syndrome and the hypothalamus. Acta Paediatr. Suppl. 1997, 423, 50–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butler, M.G.; Sturich, J.; Lee, J.; Myers, S.E.; Whitman, B.Y.; Gold, J.-A.; Kimonis, V.; Scheimann, A.; Terrazas, N.; Driscoll, D.J. Growth standards of infants with Prader-Willi syndrome. Pediatrics 2011, 127, 687–695. [Google Scholar] [CrossRef] [Green Version]
- Butler, M.G. Prader-Willi syndrome: Current understanding of cause and diagnosis. Am. J. Med. Genet. 1990, 35, 319–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carrel, A.L.; Moerchen, V.; Myers, S.E.; Bekx, M.T.; Whitman, B.Y.; Allen, D.B. Growth hormone improves mobility and body composition in infants and toddlers with Prader-Willi syndrome. J. Pediatr. 2004, 145, 744–749. [Google Scholar] [CrossRef]
- Martin, A.; State, M.; Anderson, G.M.; Kaye, W.M.; Hanchett, J.M.; McConaha, C.W.; North, W.G.; Leckman, J.F. Cerebrospinal fluid levels of oxytocin in Prader-Willi syndrome: A preliminary report. Biol. Psychiatry 1998, 44, 1349–1352. [Google Scholar] [CrossRef] [PubMed]
- Passoni, I.; Leonzino, M.; Gigliucci, V.; Chini, B.; Busnelli, M. Carbetocin is a Functional Selective Gq Agonist That Does Not Promote Oxytocin Receptor Recycling After Inducing β-Arrestin-Independent Internalisation. J. Neuroendocrinol. 2016, 28. [Google Scholar] [CrossRef] [Green Version]
- Green, L.; Fein, D.; Modahl, C.; Feinstein, C.; Waterhouse, L.; Morris, M. Oxytocin and autistic disorder: Alterations in peptide forms. Biol. Psychiatry 2001, 50, 609–613. [Google Scholar] [CrossRef]
- Kabasakalian, A.; Ferretti, C.J.; Hollander, E. Oxytocin and Prader-Willi Syndrome. Curr. Top. Behav. Neurosci. 2018, 35, 529–557. [Google Scholar] [CrossRef]
- Elefteriou, F.; Ahn, J.D.; Takeda, S.; Starbuck, M.; Yang, X.; Liu, X.; Kondo, H.; Richards, W.G.; Bannon, T.W.; Noda, M.; et al. Leptin regulation of bone resorption by the sympathetic nervous system and CART. Nature 2005, 434, 514–520. [Google Scholar] [CrossRef] [PubMed]
- Gainer, H.; Lively, M.O.; Morris, M. Immunological and related techniques for studying neurohypophyseal peptide-processing pathways. In Methods in Neurosciences; Smith, A.I., Ed.; Academic Press: Cambridge, MA, USA, 1995; pp. 195–207. [Google Scholar]


{kind=link}
| Nutritional Phase 1a: infants | Severe neonatal hypotonia with poor appetite and problems sucking necessitating assisted feeding. |
| Nutritional Phase 1b: around age 2 years | Improved appetite. |
| Nutritional Phase 2°: age 2 to 3 years | Weight gain without a change in appetite. |
| Nutritional Phase 2b: age 4–5 years | Increased interest in food with increased anxiety and behavioral problems. |
| Nutritional Phase 3: age 8 | Insatiable appetite with worsening behavioral problems combined with other endocrine problems due to hypothalamic dysfunction. These include episodes of hyperpyrexia with no infection and/or hypothermia with no causes, abnormal detection of cold and hot, abnormal perception of pain, cognitive and growth delay and autistic spectrum disorder. Post-mortem analysis indicated a decrease in Oxytocin receptor level in brain of Prader–Willi Syndrome patients and an increase in plasmatic Oxytocin. |
| First study [32] | 24 children aged 5–11 years with or without PWS diagnosis. 5 days IN-Oxt or placebo/4 weeks washout and again 5 days IN-Oxt or placebo. Positive but not significative trends of improvement in behavioral and appetite parameters. |
| Second study [33] | 26 children aged 3–11 years. 8 weeks placebo double blind IN-Oxt randomized trial. Positive trends were found in boys but not in girls, and patients with a deletion had a significant positive effect but not patients with maternal uniparental disomy associated with more severe symptoms. |
| Third study [34] | Children aged 5–18 years. Placebo was associated with improvement in hyperphagia and behavior but not the treatment with Oxytocin. |
| Fourth study [37] | 18 Infants under 6 months old. Escalating doses of Oxytocin administered over a 7 day period. Normalization of suckling, improved swallowing, improvement in muscle tone and motor coordination. Improvement in gross motor skills also in long-term effects since treated patients were able to crawl as toddlers. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Camerino, C. Oxytocin’s Regulation of Thermogenesis May Be the Link to Prader–Willi Syndrome. Curr. Issues Mol. Biol. 2023, 45, 4923-4935. https://doi.org/10.3390/cimb45060313
Camerino C. Oxytocin’s Regulation of Thermogenesis May Be the Link to Prader–Willi Syndrome. Current Issues in Molecular Biology. 2023; 45(6):4923-4935. https://doi.org/10.3390/cimb45060313
Chicago/Turabian StyleCamerino, Claudia. 2023. "Oxytocin’s Regulation of Thermogenesis May Be the Link to Prader–Willi Syndrome" Current Issues in Molecular Biology 45, no. 6: 4923-4935. https://doi.org/10.3390/cimb45060313