
Gene Expression Profiles of Methyltransferases and Demethylases Associated with Metastasis, Tumor Invasion, CpG73 Methylation, and HPV Status in Head and Neck Squamous Cell Carcinoma
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients and Sample Collection
2.2. RNA Extraction and Quantitative Reverse Transcription PCR (RT-qPCR)
2.3. Data Collection and Analysis
2.4. Statistical Analysis
3. Results
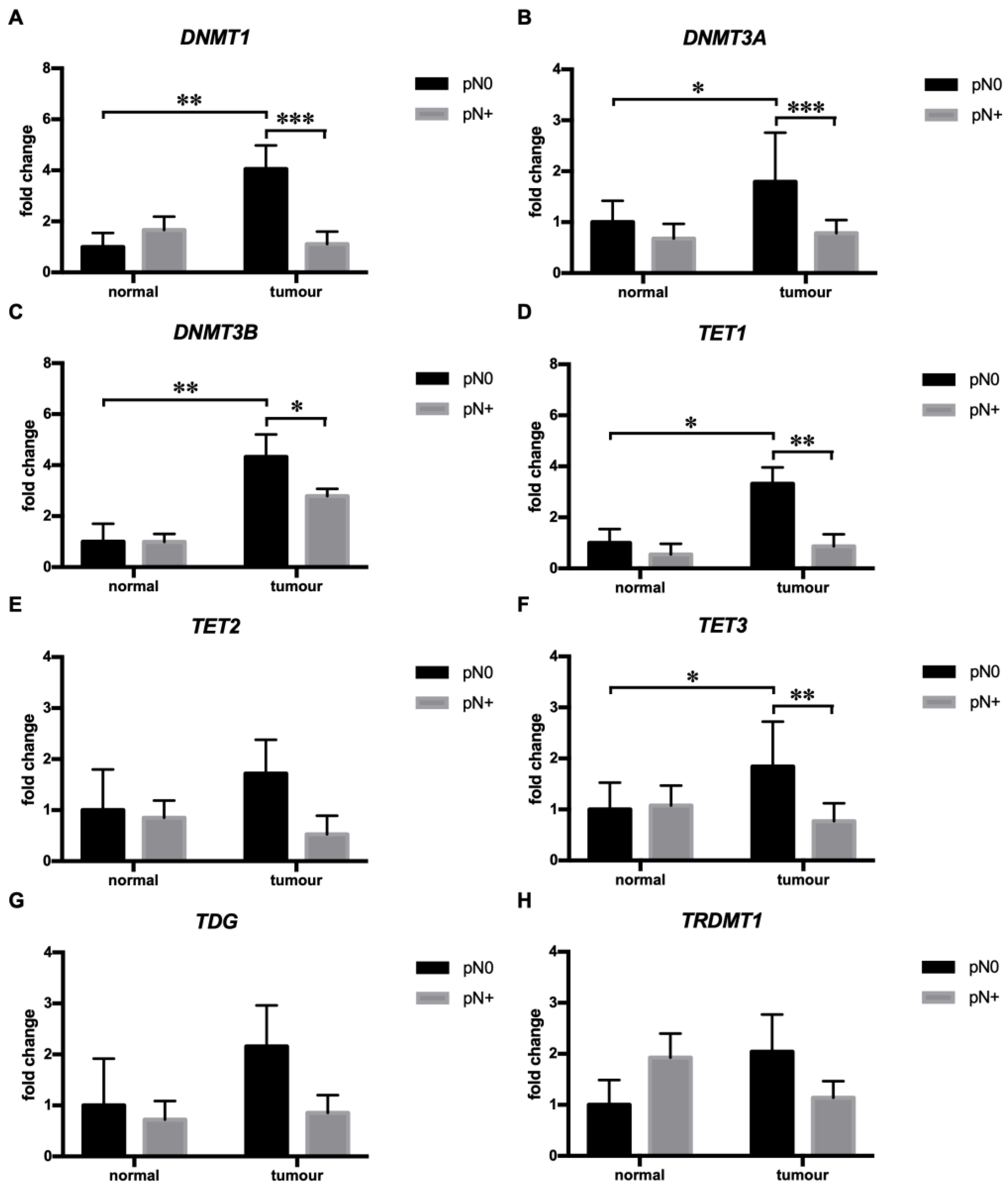
3.1. Tumors with Regional Lymph Node Metastases (pN+ Tumors) of HNSCC Showed Diminished mRNA Expression of DNMT1, DNMT3A, DNMT3B, TET1, and TET2
3.2. Perivascular Invasion (PVI) Was Associated with Diminished DNMT3B mRNA Expression in HNSCC
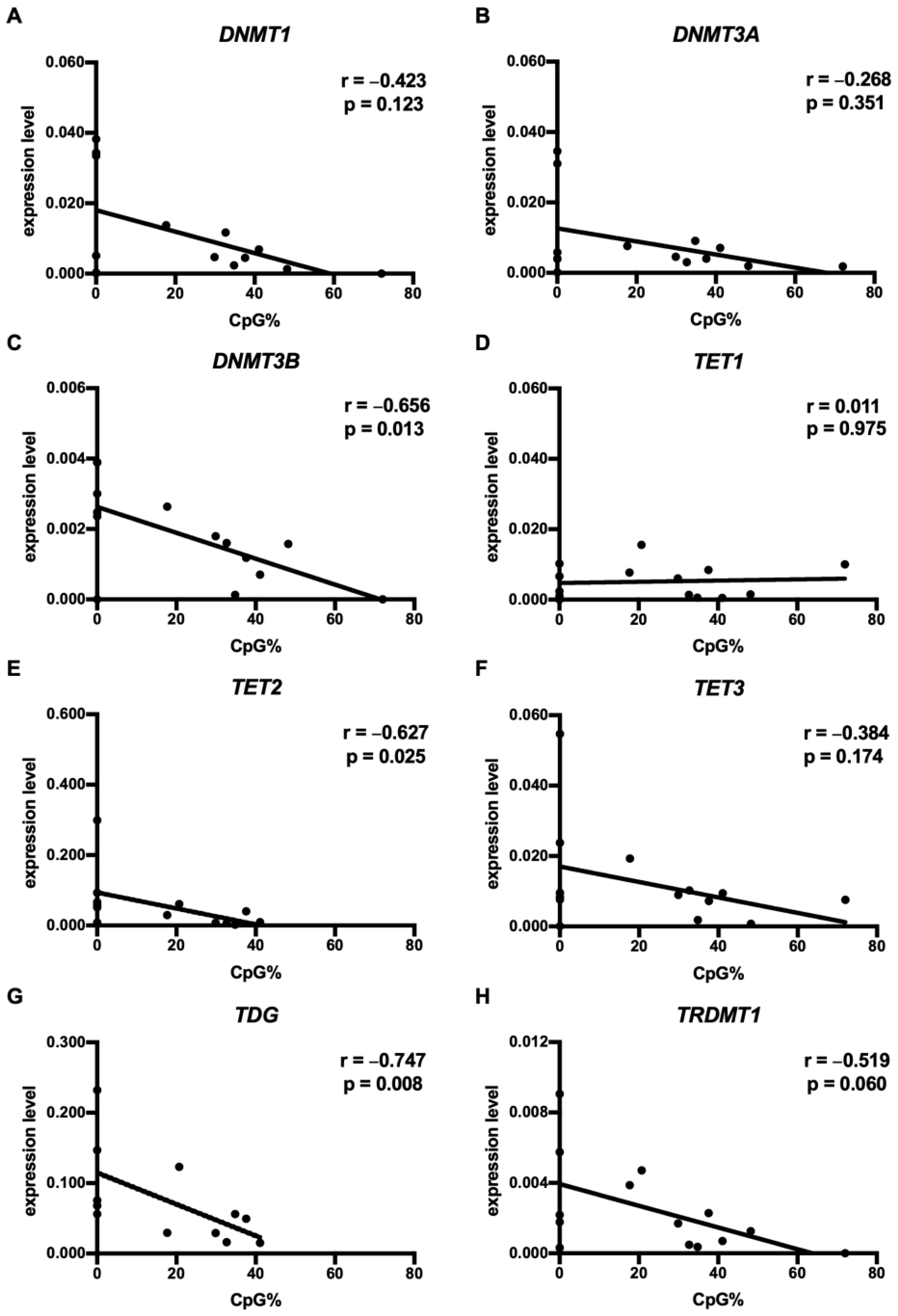
3.3. CpG73 Methylation Was Associated with the Diminished mRNA Expression of DNMT3B, TET2, and TDG in HNSCC
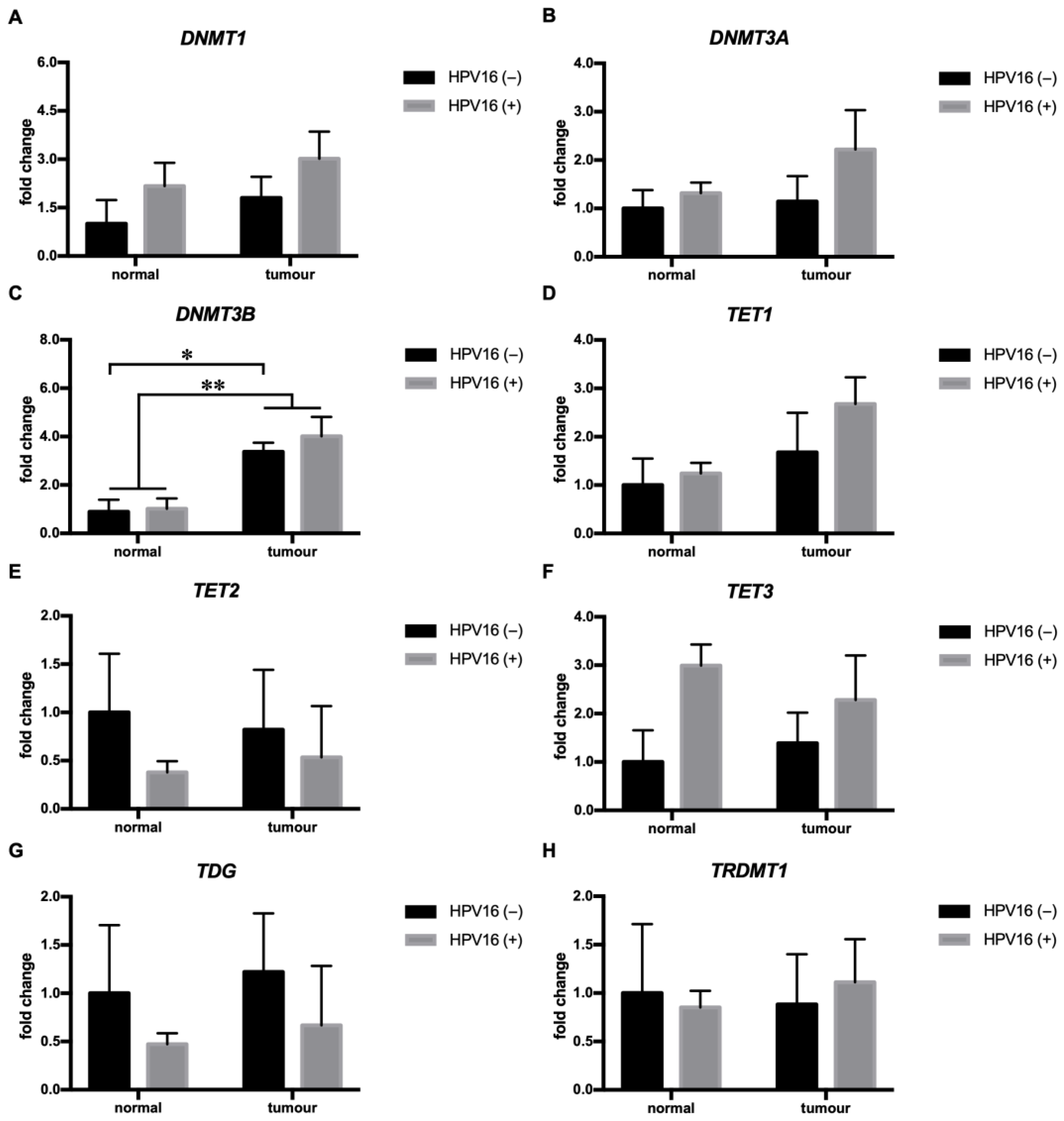
3.4. HPV16 Was Associated with Increased DNMT3B mRNA Expression in HNSCC
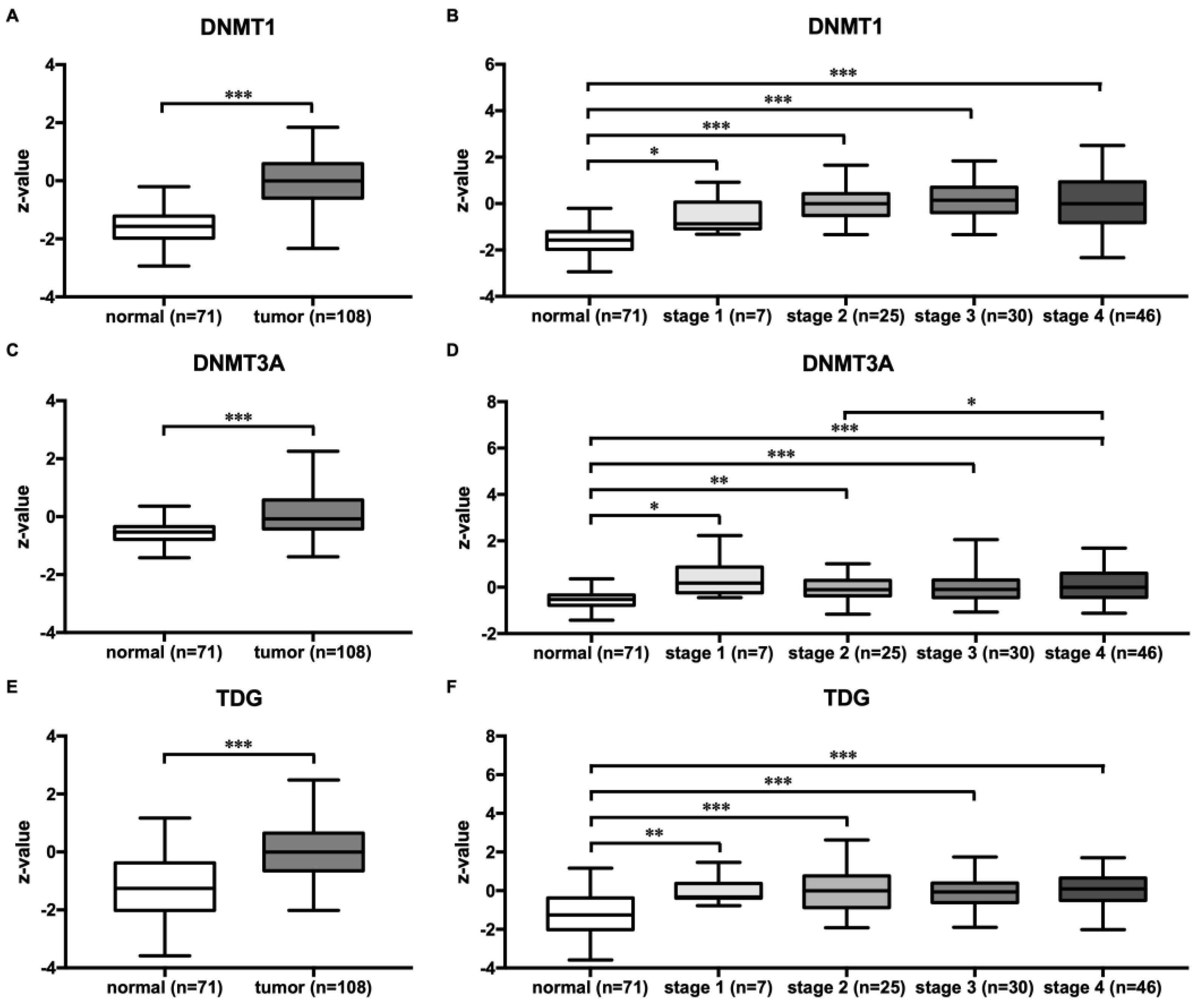
3.5. HNSCC Tumors Exhibited Higher DNMT1, 3A and TDG Protein Expression
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Johnson, D.E.; Burtness, B.; Leemans, C.R.; Lui, V.W.Y.; Bauman, J.E.; Grandis, J.R. Head and neck squamous cell carcinoma. Nat. Rev. Dis. Primers 2020, 6, 92. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Luce, D.; Gérin, M.; Leclerc, A.; Morcet, J.F.; Brugère, J.; Goldberg, M. Sinonasal cancer and occupational exposure to formaldehyde and other substances. Int. J. Cancer 1993, 53, 224–231. [Google Scholar] [CrossRef]
- Zhang, Z.F.; Morgenstern, H.; Spitz, M.R.; Tashkin, D.P.; Yu, G.P.; Hsu, T.C.; Schantz, S.P. Environmental tobacco smoking, mutagen sensitivity, and head and neck squamous cell carcinoma. Cancer Epidemiol. Biomark. Prev. 2000, 9, 1043–1049. [Google Scholar] [PubMed]
- Leemans, C.R.; Braakhuis, B.J.; Brakenhoff, R.H. The molecular biology of head and neck cancer. Nat. Rev. Cancer 2011, 11, 9–22. [Google Scholar] [CrossRef] [PubMed]
- Samuels, S.E.; Eisbruch, A.; Beitler, J.J.; Corry, J.; Bradford, C.R.; Saba, N.F.; van den Brekel, M.W.; Smee, R.; Strojan, P.; Suárez, C.; et al. Management of locally advanced HPV-related oropharyngeal squamous cell carcinoma: Where are we? Eur. Arch. Otorhinolaryngol. 2016, 273, 2877–2894. [Google Scholar] [CrossRef]
- Gillison, M.L.; Chaturvedi, A.K.; Anderson, W.F.; Fakhry, C. Epidemiology of Human Papillomavirus-Positive Head and Neck Squamous Cell Carcinoma. J. Clin. Oncol. 2015, 33, 3235–3242. [Google Scholar] [CrossRef]
- Dawson, M.A.; Kouzarides, T. Cancer epigenetics: From mechanism to therapy. Cell 2012, 150, 12–27. [Google Scholar] [CrossRef]
- Portela, A.; Esteller, M. Epigenetic modifications and human disease. Nat. Biotechnol. 2010, 28, 1057–1068. [Google Scholar] [CrossRef]
- Goelz, S.E.; Vogelstein, B.; Hamilton, S.R.; Feinberg, A.P. Hypomethylation of DNA from benign and malignant human colon neoplasms. Science 1985, 228, 187–190. [Google Scholar] [CrossRef]
- Jones, P.A. Functions of DNA methylation: Islands, start sites, gene bodies and beyond. Nat. Rev. Genet. 2012, 13, 484–492. [Google Scholar] [CrossRef] [PubMed]
- Jithesh, P.V.; Risk, J.M.; Schache, A.G.; Dhanda, J.; Lane, B.; Liloglou, T.; Shaw, R.J. The epigenetic landscape of oral squamous cell carcinoma. Br. J. Cancer 2013, 108, 370–379. [Google Scholar] [CrossRef] [PubMed]
- Lechner, M.; Frampton, G.M.; Fenton, T.; Feber, A.; Palmer, G.; Jay, A.; Pillay, N.; Forster, M.; Cronin, M.T.; Lipson, D.; et al. Targeted next-generation sequencing of head and neck squamous cell carcinoma identifies novel genetic alterations in HPV+ and HPV- tumors. Genome Med. 2013, 5, 49. [Google Scholar] [CrossRef] [PubMed]
- Smith, I.M.; Mydlarz, W.K.; Mithani, S.K.; Califano, J.A. DNA global hypomethylation in squamous cell head and neck cancer associated with smoking, alcohol consumption and stage. Int. J. Cancer 2007, 121, 1724–1728. [Google Scholar] [CrossRef]
- Viet, C.T.; Yu, G.; Asam, K.; Thomas, C.M.; Yoon, A.J.; Wongworawat, Y.C.; Haghighiabyaneh, M.; Kilkuts, C.A.; McGue, C.M.; Couey, M.A.; et al. The REASON score: An epigenetic and clinicopathologic score to predict risk of poor survival in patients with early stage oral squamous cell carcinoma. Biomark. Res. 2021, 9, 42. [Google Scholar] [CrossRef] [PubMed]
- Supic, G.; Kozomara, R.; Zeljic, K.; Jovic, N.; Magic, Z. Prognostic value of the DNMTs mRNA expression and genetic polymorphisms on the clinical outcome in oral cancer patients. Clin. Oral. Investig. 2017, 21, 173–182. [Google Scholar] [CrossRef]
- Misawa, K.; Imai, A.; Mochizuki, D.; Mima, M.; Endo, S.; Misawa, Y.; Kanazawa, T.; Mineta, H. Association of TET3 epigenetic inactivation with head and neck cancer. Oncotarget 2018, 9, 24480–24493. [Google Scholar] [CrossRef]
- Wang, Y.; Hu, H.; Wang, Q.; Li, Z.; Zhu, Y.; Zhang, W.; Jiang, H.; Cheng, J. The level and clinical significance of 5-hydroxymethylcytosine in oral squamous cell carcinoma: An immunohistochemical study in 95 patients. Pathol. Res. Pract. 2017, 213, 969–974. [Google Scholar] [CrossRef]
- Su, C.W.; Chang, Y.C.; Chien, M.H.; Hsieh, Y.H.; Chen, M.K.; Lin, C.W.; Yang, S.F. Loss of TIMP3 by promoter methylation of Sp1 binding site promotes oral cancer metastasis. Cell Death Dis. 2019, 10, 793. [Google Scholar] [CrossRef]
- Sun, R.; Juan, Y.C.; Su, Y.F.; Zhang, W.B.; Yu, Y.; Yang, H.Y.; Yu, G.Y.; Peng, X. Hypermethylated PAX1 and ZNF582 genes in the tissue sample are associated with aggressive progression of oral squamous cell carcinoma. J. Oral. Pathol. Med. 2020, 49, 751–760. [Google Scholar] [CrossRef]
- Liu, Y.; Chang, K.; Fu, K.; Dong, X.; Chen, X.; Liu, J.; Cui, N.; Ni, J. DNA demethylation of claudin-4 suppresses migration and invasion in laryngeal squamous carcinoma cells. Hum. Pathol. 2018, 75, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Bo, H.; Zhu, F.; Li, Q.; Chen, T.; Lei, S.; Fan, L. Hypomethylated SPANXA1/A2 promotes the metastasis of head and neck squamous cell carcinoma. Med. Oncol. 2020, 37, 112. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Qu, L.; He, G.; Tian, L.; Li, L.; Zhou, H.; Jin, Q.; Ren, J.; Wang, Y.; Wang, J.; et al. Regulation of laryngeal squamous cell cancer progression by the lncRNA H19/miR-148a-3p/DNMT1 axis. Oncotarget 2016, 7, 11553–11566. [Google Scholar] [CrossRef]
- Chen, L.H.; Hsu, W.L.; Tseng, Y.J.; Liu, D.W.; Weng, C.F. Involvement of DNMT 3B promotes epithelial-mesenchymal transition and gene expression profile of invasive head and neck squamous cell carcinomas cell lines. BMC Cancer 2016, 16, 431. [Google Scholar] [CrossRef]
- Hafkamp, H.C.; Speel, E.J.; Haesevoets, A.; Bot, F.J.; Dinjens, W.N.; Ramaekers, F.C.; Hopman, A.H.; Manni, J.J. A subset of head and neck squamous cell carcinomas exhibits integration of HPV 16/18 DNA and overexpression of p16INK4A and p53 in the absence of mutations in p53 exons 5-8. Int. J. Cancer 2003, 107, 394–400. [Google Scholar] [CrossRef]
- Werness, B.A.; Levine, A.J.; Howley, P.M. Association of human papillomavirus types 16 and 18 E6 proteins with p53. Science 1990, 248, 76–79. [Google Scholar] [CrossRef]
- Boyer, S.N.; Wazer, D.E.; Band, V. E7 protein of human papilloma virus-16 induces degradation of retinoblastoma protein through the ubiquitin-proteasome pathway. Cancer Res. 1996, 56, 4620–4624. [Google Scholar]
- Ghittoni, R.; Accardi, R.; Hasan, U.; Gheit, T.; Sylla, B.; Tommasino, M. The biological properties of E6 and E7 oncoproteins from human papillomaviruses. Virus Genes 2010, 40, 1–13. [Google Scholar] [CrossRef]
- Gheit, T. Mucosal and Cutaneous Human Papillomavirus Infections and Cancer Biology. Front. Oncol. 2019, 9, 355. [Google Scholar] [CrossRef]
- Burgers, W.A.; Blanchon, L.; Pradhan, S.; de Launoit, Y.; Kouzarides, T.; Fuks, F. Viral oncoproteins target the DNA methyltransferases. Oncogene 2007, 26, 1650–1655. [Google Scholar] [CrossRef]
- McCabe, M.T.; Davis, J.N.; Day, M.L. Regulation of DNA methyltransferase 1 by the pRb/E2F1 pathway. Cancer Res. 2005, 65, 3624–3632. [Google Scholar] [CrossRef] [PubMed]
- Yeung, C.L.; Tsang, T.Y.; Yau, P.L.; Kwok, T.T. Human papillomavirus type 16 E6 suppresses microRNA-23b expression in human cervical cancer cells through DNA methylation of the host gene C9orf3. Oncotarget 2017, 8, 12158–12173. [Google Scholar] [CrossRef] [PubMed]
- Sartor, M.A.; Dolinoy, D.C.; Jones, T.R.; Colacino, J.A.; Prince, M.E.; Carey, T.E.; Rozek, L.S. Genome-wide methylation and expression differences in HPV(+) and HPV(-) squamous cell carcinoma cell lines are consistent with divergent mechanisms of carcinogenesis. Epigenetics 2011, 6, 777–787. [Google Scholar] [CrossRef] [PubMed]
- Leonard, S.M.; Wei, W.; Collins, S.I.; Pereira, M.; Diyaf, A.; Constandinou-Williams, C.; Young, L.S.; Roberts, S.; Woodman, C.B. Oncogenic human papillomavirus imposes an instructive pattern of DNA methylation changes which parallel the natural history of cervical HPV infection in young women. Carcinogenesis 2012, 33, 1286–1293. [Google Scholar] [CrossRef]
- Richards, K.L.; Zhang, B.; Baggerly, K.A.; Colella, S.; Lang, J.C.; Schuller, D.E.; Krahe, R. Genome-wide hypomethylation in head and neck cancer is more pronounced in HPV-negative tumors and is associated with genomic instability. PLoS ONE 2009, 4, e4941. [Google Scholar] [CrossRef]
- Colacino, J.A.; Dolinoy, D.C.; Duffy, S.A.; Sartor, M.A.; Chepeha, D.B.; Bradford, C.R.; McHugh, J.B.; Patel, D.A.; Virani, S.; Walline, H.M.; et al. Comprehensive analysis of DNA methylation in head and neck squamous cell carcinoma indicates differences by survival and clinicopathologic characteristics. PLoS ONE 2013, 8, e54742. [Google Scholar] [CrossRef] [PubMed]
- Kostareli, E.; Holzinger, D.; Bogatyrova, O.; Hielscher, T.; Wichmann, G.; Keck, M.; Lahrmann, B.; Grabe, N.; Flechtenmacher, C.; Schmidt, C.R.; et al. HPV-related methylation signature predicts survival in oropharyngeal squamous cell carcinomas. J. Clin. Investig. 2013, 123, 2488–2501. [Google Scholar] [CrossRef]
- Degli Esposti, D.; Sklias, A.; Lima, S.C.; Beghelli-de la Forest Divonne, S.; Cahais, V.; Fernandez-Jimenez, N.; Cros, M.P.; Ecsedi, S.; Cuenin, C.; Bouaoun, L.; et al. Unique DNA methylation signature in HPV-positive head and neck squamous cell carcinomas. Genome Med. 2017, 9, 33. [Google Scholar] [CrossRef]
- Misawa, K.; Imai, A.; Matsui, H.; Kanai, A.; Misawa, Y.; Mochizuki, D.; Mima, M.; Yamada, S.; Kurokawa, T.; Nakagawa, T.; et al. Identification of novel methylation markers in HPV-associated oropharyngeal cancer: Genome-wide discovery, tissue verification and validation testing in ctDNA. Oncogene 2020, 39, 4741–4755. [Google Scholar] [CrossRef]
- Fakhry, C.; Westra, W.H.; Li, S.; Cmelak, A.; Ridge, J.A.; Pinto, H.; Forastiere, A.; Gillison, M.L. Improved survival of patients with human papillomavirus-positive head and neck squamous cell carcinoma in a prospective clinical trial. J. Natl. Cancer Inst. 2008, 100, 261–269. [Google Scholar] [CrossRef]
- Dayyani, F.; Etzel, C.J.; Liu, M.; Ho, C.H.; Lippman, S.M.; Tsao, A.S. Meta-analysis of the impact of human papillomavirus (HPV) on cancer risk and overall survival in head and neck squamous cell carcinomas (HNSCC). Head Neck Oncol. 2010, 2, 15. [Google Scholar] [CrossRef] [PubMed]
- Menezes, F.D.S.; Fernandes, G.A.; Antunes, J.L.F.; Villa, L.L.; Toporcov, T.N. Global incidence trends in head and neck cancer for HPV-related and -unrelated subsites: A systematic review of population-based studies. Oral. Oncol. 2021, 115, 105177. [Google Scholar] [CrossRef] [PubMed]
- Čelešnik, H.; Büdefeld, T.; Čizmarević, B.; Švagan, M.; Potočnik, U. MIR137/MIR2682 locus is associated with perineural invasiveness in head and neck cancer. J. Oral. Pathol. Med. 2021, 50, 874–881. [Google Scholar] [CrossRef]
- Chen, F.; Chandrashekar, D.S.; Varambally, S.; Creighton, C.J. Pan-cancer molecular subtypes revealed by mass-spectrometry-based proteomic characterization of more than 500 human cancers. Nat. Commun. 2019, 10, 5679. [Google Scholar] [CrossRef] [PubMed]
- Chandrashekar, D.S.; Karthikeyan, S.K.; Korla, P.K.; Patel, H.; Shovon, A.R.; Athar, M.; Netto, G.J.; Qin, Z.S.; Kumar, S.; Manne, U.; et al. UALCAN: An update to the integrated cancer data analysis platform. Neoplasia 2022, 25, 18–27. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, F.; Chandrashekar, D.S.; Varambally, S.; Creighton, C.J. Proteogenomic characterization of 2002 human cancers reveals pan-cancer molecular subtypes and associated pathways. Nat. Commun. 2022, 13, 2669. [Google Scholar] [CrossRef]
- Han, Z.; Yang, B.; Wang, Y.; Zeng, X.; Tian, Z. Identification of Expression Patterns and Potential Prognostic Significance of m. Front. Oncol. 2021, 11, 592107. [Google Scholar] [CrossRef]
- Zhang, W.; Xu, J. DNA methyltransferases and their roles in tumorigenesis. Biomark. Res. 2017, 5, 1. [Google Scholar] [CrossRef]
- Dong, J.; Li, J.; Li, Y.; Ma, Z.; Yu, Y.; Wang, C.Y. Transcriptional super-enhancers control cancer stemness and metastasis genes in squamous cell carcinoma. Nat. Commun. 2021, 12, 3974. [Google Scholar] [CrossRef]
- Xue, M.; Shi, Q.; Zheng, L.; Li, Q.; Yang, L.; Zhang, Y. Gene signatures of m5C regulators may predict prognoses of patients with head and neck squamous cell carcinoma. Am. J. Transl. Res. 2020, 12, 6841–6852. [Google Scholar]
- Liu, S.; Zhao, Y.; Xu, Y.; Sang, M.; Zhao, R.; Gu, L.; Shan, B. The clinical significance of methylation of MAGE-A1 and-A3 promoters and expression of DNA methyltransferase in patients with laryngeal squamous cell carcinoma. Am. J. Otolaryngol. 2020, 41, 102318. [Google Scholar] [CrossRef]
- Li, M.; Wang, Y.; Song, Y.; Bu, R.; Yin, B.; Fei, X.; Guo, Q.; Wu, B. Expression profiling and clinicopathological significance of DNA methyltransferase 1, 3A and 3B in sporadic human renal cell carcinoma. Int. J. Clin. Exp. Pathol. 2014, 7, 7597–7609. [Google Scholar] [PubMed]
- Zhang, F.; Zhu, Y.; Fan, G.; Hu, S. MicroRNA-2682-3p inhibits osteosarcoma cell proliferation by targeting CCND2, MMP8 and Myd88. Oncol. Lett. 2018, 16, 3359–3364. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, Y.; Zhang, L.; You, G.; Li, C.; Meng, B.; Zhou, M.; Zhang, M. LINC01006 promotes cell proliferation and metastasis in pancreatic cancer via miR-2682-5p/HOXB8 axis. Cancer Cell. Int. 2019, 19, 320. [Google Scholar] [CrossRef]
- Williams, K.; Christensen, J.; Helin, K. DNA methylation: TET proteins-guardians of CpG islands? EMBO Rep. 2011, 13, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Heawchaiyaphum, C.; Ekalaksananan, T.; Patarapadungkit, N.; Worawichawong, S.; Pientong, C. Epstein-Barr Virus Infection Alone or Jointly with Human Papillomavirus Associates with Down-Regulation of miR-145 in Oral Squamous-Cell Carcinoma. Microorganisms 2021, 9, 2496. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clinicopathological Features | Men (n = 30) Number (%) | Women (n = 4) Number (%) |
|---|---|---|
| Age at diagnosis | 44–79 yrs. (mean 61.5 yrs.) | 50–65 yrs. (mean 57.8 yrs.) |
| Site | ||
| Lip and oral cavity | 4 (13.3%) | 1 (25.0%) |
| Pharynx | 20 (66.7%) | 2 (50.0%) |
| Larynx | 6 (20.0%) | 1 (25.0%) |
| Invasion 1 | ||
| Perivascular | ||
| Yes | 9 (30.0%) | 0 (0.0%) |
| No | 20 (66.7%) | 4 (100.0%) |
| Missing | 1 (3.3%) | / |
| Lymphovascular | ||
| Yes | 7 (23.3%) | 1 (25.0%) |
| No | 18 (60.0%) | 1 (25.0%) |
| Missing | 5 (16.7%) | 2 (50.0%) |
| Perineural | ||
| Yes | 8 (26.7%) | 0 (0.0%) |
| No | 21 (70.0%) | 4 (100.0%) |
| Missing | 1 (3.3%) | 0 (0.0%) |
| p16 status | ||
| Yes | 7 (23.3%) | / |
| No | 12 (40.0%) | 2 (50.0%) |
| Missing | 11 (36.7%) | 2 (50.0%) |
| Nicotine | ||
| Yes | 26 (86.6%) | 4 (100.0%) |
| No | 2 (6.7%) | / |
| Missing | 2 (6.7%) | / |
| Alcohol | ||
| Yes | 26 (86.6%) | 4 (100%) |
| No | 2 (6.7%) | / |
| Missing | 2 (6.7%) | / |
| Stage | ||
| pT (1–2) | 8 (26.6%) | 2 (50%) |
| pT (3–4) | 20 (66.7%) | 2 (50%) |
| Missing | 2 (6.7%) | / |
| pN (0) | 6 (20%) | 1 (25.0%) |
| pN (+) | 22 (73.3%) | 3 (75.0%) |
| Missing | 2 (6.7%) | / |
| Gene | Primer Sequence | |
|---|---|---|
| DNMT1 | F: R: | 5′-AGCCGAGCGAGCCAGAGATA-3′ 5′-CGTGTCAGAGATGCCTGCTT-3′ |
| DNMT3A | F: R: | 5′-ATGGAATCGCTACAGGGCTC-3′ 5′-CTTCTGTGTGACGCTGCG-3′ |
| DNMT3B | F: R: | 5′-ATGGCAAGTTCTCCGAGGTC-3′ 5′-CGATAGGAGACGAGCTTATTGA-3′ |
| TET1 | F: R: | 5′-AGCTGTCTTGATCGAGTTATACA-3′ 5′-CCCTTCTTTACCGGTGTACACTA-3′ |
| TET2 | F: R: | 5′-CTGGCAAACATTCAGCAGCA-3′ 5′-TTGAATTCAGCAGCTCAGTCC-3′ |
| TET3 | F: R: | 5′-GGAACTCATGGAGGAGCGGTAT-3′ 5′-GATCACAGCGTTCTGGCAGT-3′ |
| TRDMT1 | F: R: | 5′-TCTCCAACCTCTCTTGGCATTC-3′ 5′-GGAACTCCATCAGTACCTGACCA-3′ |
| TDG | F: R: | 5′-TGGACGTTCAAGAGGTGCAA-3′ 5′-CTTAACTCCACGCTCTCAATTAGC-3′ |
| ERBB3 | F: R: | 5′-TGAGGCGATACTTGGAACGG-3′ 5′-TGGCCAGCATATGATCTGTC-3′ |
| ACTB | F: R: | 5′-CATCGAGCACGGCATCGTCA-3′ 5′-TAGCACAGCCTGGATAGCAAC-3′ |
| GAPDH | F: R: | 5′-TGAGAACGGGAAGCTTGTCA-3′ 5′-CCCTGCAAATGAGCCCCA-3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Goričan, L.; Büdefeld, T.; Čelešnik, H.; Švagan, M.; Lanišnik, B.; Potočnik, U. Gene Expression Profiles of Methyltransferases and Demethylases Associated with Metastasis, Tumor Invasion, CpG73 Methylation, and HPV Status in Head and Neck Squamous Cell Carcinoma. Curr. Issues Mol. Biol. 2023, 45, 4632-4646. https://doi.org/10.3390/cimb45060294
Goričan L, Büdefeld T, Čelešnik H, Švagan M, Lanišnik B, Potočnik U. Gene Expression Profiles of Methyltransferases and Demethylases Associated with Metastasis, Tumor Invasion, CpG73 Methylation, and HPV Status in Head and Neck Squamous Cell Carcinoma. Current Issues in Molecular Biology. 2023; 45(6):4632-4646. https://doi.org/10.3390/cimb45060294
Chicago/Turabian StyleGoričan, Larisa, Tomaž Büdefeld, Helena Čelešnik, Matija Švagan, Boštjan Lanišnik, and Uroš Potočnik. 2023. "Gene Expression Profiles of Methyltransferases and Demethylases Associated with Metastasis, Tumor Invasion, CpG73 Methylation, and HPV Status in Head and Neck Squamous Cell Carcinoma" Current Issues in Molecular Biology 45, no. 6: 4632-4646. https://doi.org/10.3390/cimb45060294