
TMEM211 Promotes Tumor Progression and Metastasis in Colon Cancer
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Transient Transfection
2.3. Real-Time PCR (RT-PCR)
2.4. Cell Invasion and Migration
2.5. Western Blotting
2.6. Statistical Analysis
3. Results
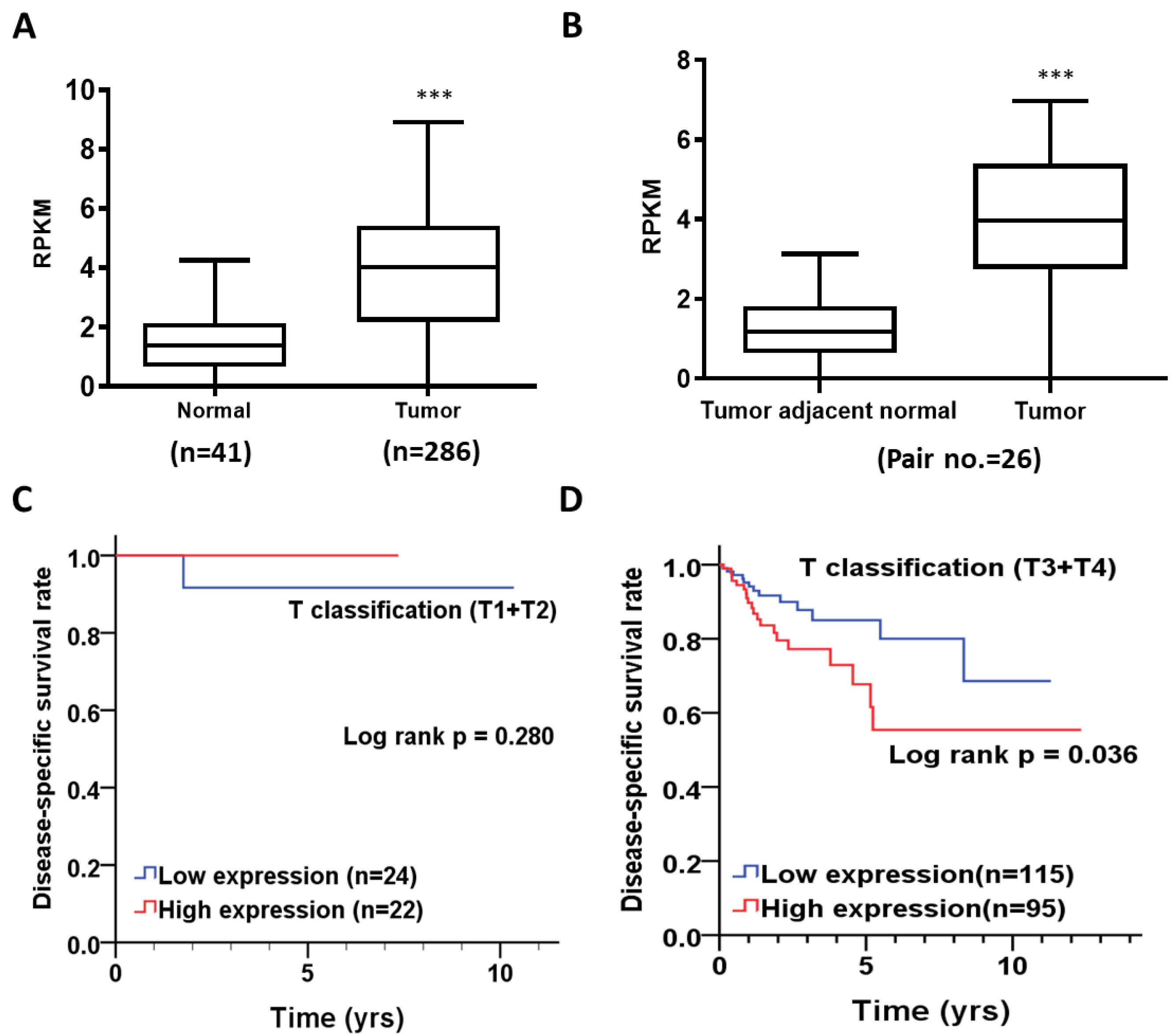
3.1. Differential Expression of TMEM211 of Normal and Tumor Tissues of Colon Cancer Patients
3.2. The Association of TMEM211 Expression with Prognosis in Colon Cancer Patients Stratified by Clinicopathological Outcomes
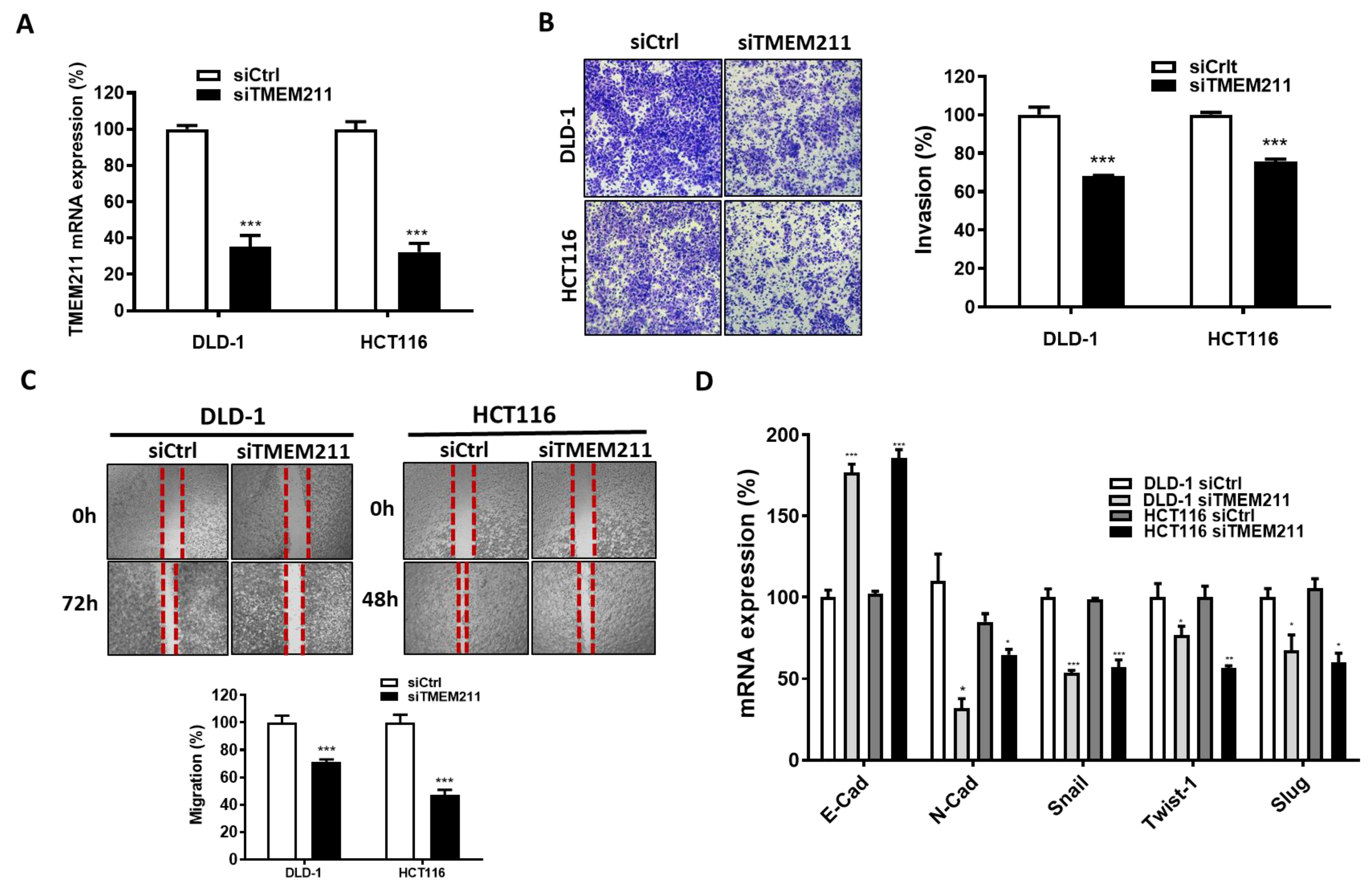
3.3. The Involvement of TMEM211 in Migration/Invasion and EMT Marker Expression of Colon Cancer Cells
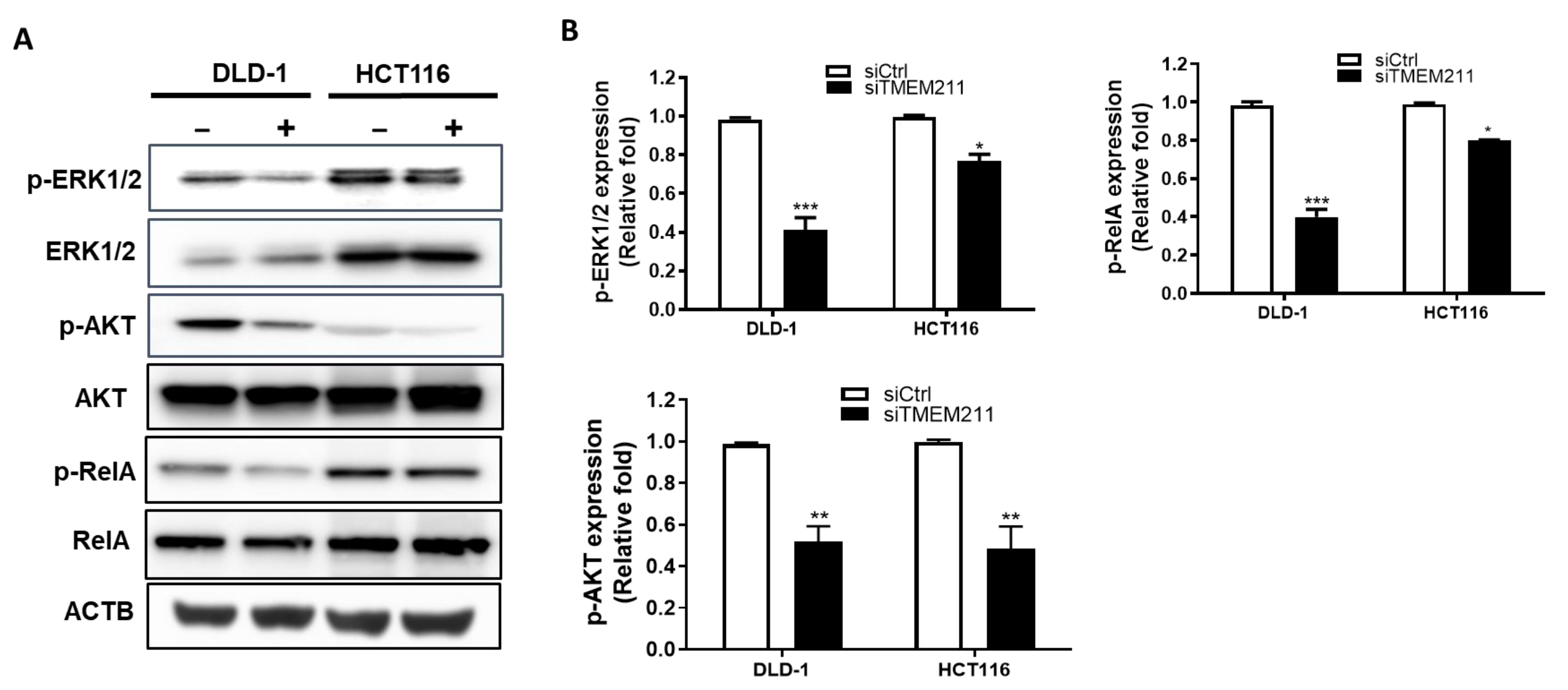
3.4. The Regulation of ERK, AKT and NF-κB Signaling Pathways by TMEM211 in Colon Cancer Cells
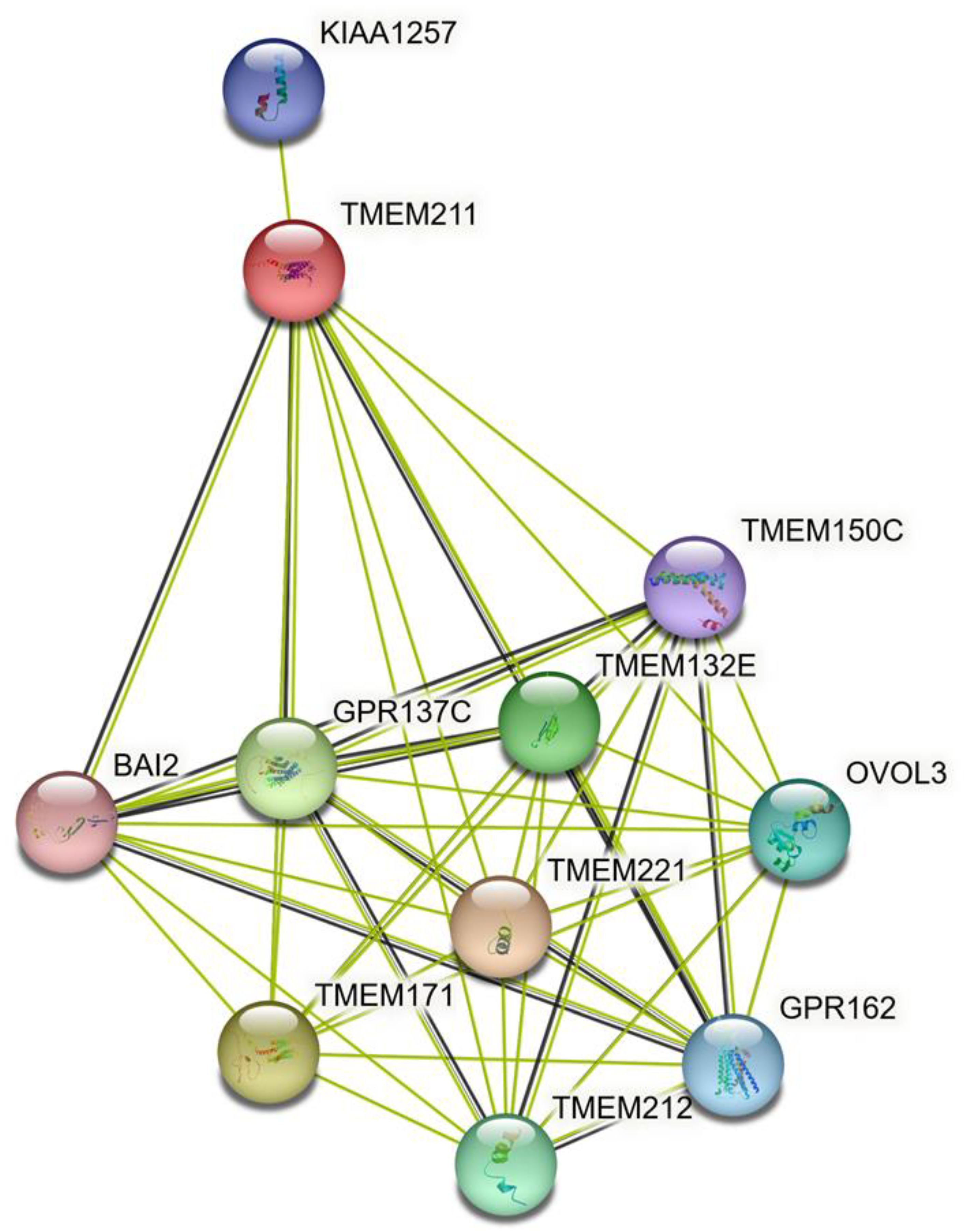
3.5. The Predicted Interaction of TMEM211 and Its Associated Proteins
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer statistics, 2022. CA Cancer J. Clin. 2022, 72, 7–33. [Google Scholar] [CrossRef]
- Siegel, R.L.; Wagle, N.S.; Cercek, A.; Smith, R.A.; Jemal, A. Colorectal cancer statistics, 2023. CA Cancer J. Clin. 2023, 73, 233–254. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Leowattana, W.; Leowattana, P.; Leowattana, T. Systemic treatment for metastatic colorectal cancer. World J. Gastroenterol. 2023, 29, 1569–1588. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.; Bhatt, L.K.; Johnston, T.P.; Prabhavalkar, K.S. Colon cancer stem cells: Potential target for the treatment of colorectal cancer. Cancer Biol. Ther. 2019, 20, 1068–1082. [Google Scholar] [CrossRef] [PubMed]
- Schmit, K.; Michiels, C. TMEM Proteins in Cancer: A Review. Front. Pharmacol. 2018, 9, 1345. [Google Scholar] [CrossRef]
- Marx, S.; Dal Maso, T.; Chen, J.W.; Bury, M.; Wouters, J.; Michiels, C.; Le Calvé, B. Transmembrane (TMEM) protein family members: Poorly characterized even if essential for the metastatic process. Semin. Cancer Biol. 2020, 60, 96–106. [Google Scholar] [CrossRef]
- Gregersen, N.O.; Buttenschøn, H.N.; Hedemand, A.; Dahl, H.A.; Kristensen, A.S.; Clementsen, B.; Woldbye, D.P.; Koefoed, P.; Erhardt, A.; Kruse, T.A.; et al. Are TMEM genes potential candidate genes for panic disorder? Psychiatr. Genet. 2014, 24, 37–41. [Google Scholar] [CrossRef]
- Perneel, J.; Neumann, M.; Heeman, B.; Cheung, S.; Van den Broeck, M.; Wynants, S.; Baker, M.; Vicente, C.T.; Faura, J.; Rademakers, R.; et al. Accumulation of TMEM106B C-terminal fragments in neurodegenerative disease and aging. Acta Neuropathol. 2023, 145, 285–302. [Google Scholar] [CrossRef]
- Fu, K.; Miyamoto, Y.; Sumi, K.; Saika, E.; Muramatsu, S.I.; Uno, K.; Nitta, A. Overexpression of transmembrane protein 168 in the mouse nucleus accumbens induces anxiety and sensorimotor gating deficit. PLoS ONE 2017, 12, e0189006. [Google Scholar] [CrossRef]
- Segelcke, D.; Fischer, H.K.; Hütte, M.; Dennerlein, S.; Benseler, F.; Brose, N.; Pogatzki-Zahn, E.M.; Schmidt, M. Tmem160 contributes to the establishment of discrete nerve injury-induced pain behaviors in male mice. Cell Rep. 2021, 37, 110152. [Google Scholar] [CrossRef] [PubMed]
- Negro, A.; Graiani, G.; Nicoli, D.; Farnetti, E.; Casali, B.; Verzicco, I.; Tedeschi, S.; Ghirarduzzi, A.; Cannone, V.; Marco, L.D.E.; et al. Concurrent heterozygous Von-Hippel-Lindau and transmembrane-protein-127 gene mutation causing an erythropoietin-secreting pheochromocytoma in a normotensive patient with severe erythrocytosis. J. Hypertens. 2020, 38, 340–346. [Google Scholar] [CrossRef]
- Sui, Y.; Sun, M.; Wu, F.; Yang, L.; Di, W.; Zhang, G.; Zhong, L.; Ma, Z.; Zheng, J.; Fang, X.; et al. Inhibition of TMEM16A expression suppresses growth and invasion in human colorectal cancer cells. PLoS ONE 2014, 9, e115443. [Google Scholar] [CrossRef] [PubMed]
- Manawapat-Klopfer, A.; Thomsen, L.T.; Martus, P.; Munk, C.; Russ, R.; Gmuender, H.; Frederiksen, K.; Haedicke-Jarboui, J.; Stubenrauch, F.; Kjaer, S.K.; et al. TMEM45A, SERPINB5 and p16INK4A transcript levels are predictive for development of high-grade cervical lesions. Am. J. Cancer Res. 2016, 6, 1524–1536. [Google Scholar]
- Wu, Y.; Wan, X.; Jia, G.; Xu, Z.; Tao, Y.; Song, Z.; Du, T. Aberrantly Methylated and Expressed Genes as Prognostic Epigenetic Biomarkers for Colon Cancer. DNA Cell Biol. 2020, 39, 1961–1969. [Google Scholar] [CrossRef] [PubMed]
- Ohnami, S.; Maruyama, K.; Chen, K.; Takahashi, Y.; Hatakeyama, K.; Ohshima, K.; Shimoda, Y.; Sakai, A.; Kamada, F.; Nakatani, S.; et al. BMP4 and PHLDA1 are plausible drug-targetable candidate genes for KRAS G12A-, G12D-, and G12V-driven colorectal cancer. Mol. Cell Biochem. 2021, 476, 3469–3482. [Google Scholar] [CrossRef] [PubMed]
- Majidpoor, J.; Mortezaee, K. Steps in metastasis: An updated review. Med. Oncol. 2021, 38, 3. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, M.; Christofori, G. EMT, the cytoskeleton, and cancer cell invasion. Cancer Metastasis Rev. 2009, 28, 15–33. [Google Scholar] [CrossRef]
- Thiery, J.P.; Acloque, H.; Huang, R.Y.; Nieto, M.A. Epithelial-mesenchymal transitions in development and disease. Cell 2009, 139, 871–890. [Google Scholar] [CrossRef]
- Dongre, A.; Weinberg, R.A. New insights into the mechanisms of epithelial-mesenchymal transition and implications for cancer. Nat. Rev. Mol. Cell Biol. 2019, 20, 69–84. [Google Scholar] [CrossRef]
- Singh, M.; Yelle, N.; Venugopal, C.; Singh, S.K. EMT: Mechanisms and therapeutic implications. Pharmacol. Ther. 2018, 182, 80–94. [Google Scholar] [CrossRef] [PubMed]
- Kaszak, I.; Witkowska-Piłaszewicz, O.; Niewiadomska, Z.; Dworecka-Kaszak, B.; Ngosa Toka, F.; Jurka, P. Role of Cadherins in Cancer-A Review. Int. J. Mol. Sci. 2020, 21, 7624. [Google Scholar] [CrossRef] [PubMed]
- Odero-Marah, V.; Hawsawi, O.; Henderson, V.; Sweeney, J. Epithelial-Mesenchymal Transition (EMT) and Prostate Cancer. Adv. Exp. Med. Biol. 2018, 1095, 101–110. [Google Scholar] [PubMed]
- Pal, M.; Bhattacharya, S.; Kalyan, G.; Hazra, S. Cadherin profiling for therapeutic interventions in Epithelial Mesenchymal Transition (EMT) and tumorigenesis. Exp. Cell Res. 2018, 368, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Radice, G.L. N-cadherin-mediated adhesion and signaling from development to disease: Lessons from mice. Prog. Mol. Biol. Transl. Sci. 2013, 116, 263–289. [Google Scholar]
- Andreiana, B.C.; Stepan, A.E.; Taisescu, O.; Al Khatib, A.M.; Florescu, M.M.; Simionescu, C.E.; Crişan, A.E. Immunoexpression of Snail, Twist1 and Slug in clear cell renal cell carcinoma. Rom. J. Morphol. Embryol. 2019, 60, 463–468. [Google Scholar]
- Sterneck, E.; Poria, D.K.; Balamurugan, K. Slug and E-Cadherin: Stealth Accomplices? Front. Mol. Biosci. 2020, 7, 138. [Google Scholar] [CrossRef]
- Paul, M.C.; Schneeweis, C.; Falcomatà, C.; Shan, C.; Rossmeisl, D.; Koutsouli, S.; Klement, C.; Zukowska, M.; Widholz, S.A.; Jesinghaus, M.; et al. Non-canonical functions of SNAIL drive context-specific cancer progression. Nat. Commun. 2023, 14, 1201. [Google Scholar] [CrossRef]
- de Morais, E.F.; de Farias Morais, H.G.; de Moura Santos, E.; Barboza, C.A.G.; Téo, F.H.; Salo, T.; Coletta, R.D.; de Almeida Freitas, R. TWIST1 regulates proliferation, migration, and invasion and is a prognostic marker for oral tongue squamous cell carcinoma. J. Oral Pathol. Med. 2023, 52, 127–135. [Google Scholar] [CrossRef]
- Liu, F.; Cao, Q.H.; Lu, D.J.; Luo, B.; Lu, X.F.; Luo, R.C.; Wang, X.G. TMEM16A overexpression contributes to tumor invasion and poor prognosis of human gastric cancer through TGF-β signaling. Oncotarget 2015, 6, 11585–11599. [Google Scholar] [CrossRef]
- Zhang, X.; He, Y.; Jiang, Y.; Bao, Y.; Chen, Q.; Xie, D.; Yu, H.; Wang, X. TMEM229A suppresses non-small cell lung cancer progression via inactivating the ERK pathway. Oncol. Rep. 2021, 46, 176. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Song, K.; Zhang, Y.; Xu, H.; Zhang, X.; Wang, L.; Fan, C.; Jiang, G.; Wang, E. TMEM17 promotes malignant progression of breast cancer via AKT/GSK3β signaling. Cancer Manag. Res. 2018, 10, 2419–2428. [Google Scholar] [CrossRef] [PubMed]
- Anjum, J.; Mitra, S.; Das, R.; Alam, R.; Mojumder, A.; Emran, T.B.; Islam, F.; Rauf, A.; Hossain, M.J.; Aljohani, A.S.M.; et al. A renewed concept on the MAPK signaling pathway in cancers: Polyphenols as a choice of therapeutics. Pharmacol. Res. 2022, 184, 106398. [Google Scholar] [CrossRef]
- Miricescu, D.; Totan, A.; Stanescu, S., II; Badoiu, S.C.; Stefani, C.; Greabu, M. PI3K/AKT/mTOR Signaling Pathway in Breast Cancer: From Molecular Landscape to Clinical Aspects. Int. J. Mol. Sci. 2020, 22, 173. [Google Scholar] [CrossRef] [PubMed]
- Eisenreich, A.; Orphal, M.; Böhme, K.; Kreutz, R. Tmem63c is a potential pro-survival factor in angiotensin II-treated human podocytes. Life Sci. 2020, 258, 118175. [Google Scholar] [CrossRef] [PubMed]
- Pires, B.R.; Mencalha, A.L.; Ferreira, G.M.; de Souza, W.F.; Morgado-Díaz, J.A.; Maia, A.M.; Corrêa, S.; Abdelhay, E.S. NF-kappaB Is Involved in the Regulation of EMT Genes in Breast Cancer Cells. PLoS ONE 2017, 12, e0169622. [Google Scholar] [CrossRef]
- Jiang, C.; Zhu, Y.; Zhou, Z.; Gumin, J.; Bengtsson, L.; Wu, W.; Songyang, Z.; Lang, F.F.; Lin, X. TMEM43/LUMA is a key signaling component mediating EGFR-induced NF-κB activation and tumor progression. Oncogene 2017, 36, 2813–2823. [Google Scholar] [CrossRef]
- Sun, T.; Bi, F.; Liu, Z.; Yang, Q. TMEM119 facilitates ovarian cancer cell proliferation, invasion, and migration via the PDGFRB/PI3K/AKT signaling pathway. J. Transl. Med. 2021, 19, 111. [Google Scholar] [CrossRef]
- Liu, P.F.; Shu, C.W.; Lee, C.H.; Sie, H.C.; Liou, H.H.; Cheng, J.T.; Ger, L.P.; Chen, C.L.; Chen, C.C.; Chen, C.F. Clinical Significance and the Role of Guanylate-Binding Protein 5 in Oral Squamous Cell Carcinoma. Cancers 2021, 13, 4043. [Google Scholar] [CrossRef]
- Tseng, Y.K.; Chen, C.F.; Shu, C.W.; Lee, C.H.; Chou, Y.T.; Li, Y.J.; Liou, H.H.; Cheng, J.T.; Chen, C.L.; Ger, L.P.; et al. Effect of EGFR on SQSTM1 Expression in Malignancy and Tumor Progression of Oral Squamous Cell Carcinoma. Int. J. Mol. Sci. 2021, 22, 12226. [Google Scholar] [CrossRef]
- Mlecnik, B.; Lugli, A.; Bindea, G.; Marliot, F.; Bifulco, C.; Lee, J.J.; Zlobec, I.; Rau, T.T.; Berger, M.D.; Nagtegaal, I.D.; et al. Multicenter International Study of the Consensus Immunoscore for the Prediction of Relapse and Survival in Early-Stage Colon Cancer. Cancers 2023, 15, 418. [Google Scholar] [CrossRef] [PubMed]
- Dolcet, X.; Llobet, D.; Pallares, J.; Matias-Guiu, X. NF-kB in development and progression of human cancer. Virchows Arch. 2005, 446, 475–482. [Google Scholar] [CrossRef] [PubMed]
- Shiraishi, T.; Ikeda, K.; Tsukada, Y.; Nishizawa, Y.; Sasaki, T.; Ito, M.; Kojima, M.; Ishii, G.; Tsumura, R.; Saijou, S.; et al. High expression of TMEM180, a novel tumour marker, is associated with poor survival in stage III colorectal cancer. BMC Cancer 2021, 21, 302. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Wang, X.; Zhang, X.; Zhan, J.; Zhang, B.; Jia, J.; Chen, J. TMEM14A aggravates the progression of human ovarian cancer cells by enhancing the activity of glycolysis. Exp. Ther. Med. 2022, 24, 614. [Google Scholar] [CrossRef]
- Duan, J.; Qian, Y.; Fu, X.; Chen, M.; Liu, K.; Liu, H.; Yang, J.; Liu, C.; Chang, Y. TMEM106C contributes to the malignant characteristics and poor prognosis of hepatocellular carcinoma. Aging 2021, 13, 5585–5606. [Google Scholar] [CrossRef]
- Saberbaghi, T.; Wong, R.; Rutka, J.T.; Wang, G.L.; Feng, Z.P.; Sun, H.S. Role of Cl− channels in primary brain tumour. Cell Calcium 2019, 81, 1–11. [Google Scholar] [CrossRef]
- Li, J.; Song, Y.; Zhang, C.; Wang, R.; Hua, L.; Guo, Y.; Gan, D.; Zhu, L.; Li, S.; Ma, P.; et al. TMEM43 promotes pancreatic cancer progression by stabilizing PRPF3 and regulating RAP2B/ERK axis. Cell. Mol. Biol. Lett. 2022, 27, 24. [Google Scholar] [CrossRef]
- Guo, Y.; Cao, F.; Hu, S.; Liu, S.; Sun, H.; Li, A.; Li, F. TMEM176A acts as a tumor suppressor gene in pancreatic cancer by inhibiting ERK signaling. Discov. Med. 2020, 30, 145–153. [Google Scholar]
- Zhang, S.; Dai, H.; Li, W.; Wang, R.; Wu, H.; Shen, M.; Hu, Y.; Xie, L.; Xing, Y. TMEM116 is required for lung cancer cell motility and metastasis through PDK1 signaling pathway. Cell Death Dis. 2021, 12, 1086. [Google Scholar] [CrossRef]
- Ebrahimi, N.; Afshinpour, M.; Fakhr, S.S.; Kalkhoran, P.G.; Shadman-Manesh, V.; Adelian, S.; Beiranvand, S.; Rezaei-Tazangi, F.; Khorram, R.; Hamblin, M.R.; et al. Cancer stem cells in colorectal cancer: Signaling pathways involved in stemness and therapy resistance. Crit. Rev. Oncol. Hematol. 2023, 182, 103920. [Google Scholar] [CrossRef]
- Chen, J.; Wang, D.; Chen, H.; Gu, J.; Jiang, X.; Han, F.; Cao, J.; Liu, W.; Liu, J. TMEM196 inhibits lung cancer metastasis by regulating the Wnt/beta-catenin signaling pathway. J. Cancer Res. Clin. Oncol. 2023, 149, 653–667. [Google Scholar] [CrossRef] [PubMed]
- Koteluk, O.; Bielicka, A.; Lemańska, Ż.; Jóźwiak, K.; Klawiter, W.; Mackiewicz, A.; Kazimierczak, U.; Kolenda, T. The Landscape of Transmembrane Protein Family Members in Head and Neck Cancers: Their Biological Role and Diagnostic Utility. Cancers 2021, 13, 4737. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TMEM211 Expression | Normal (n = 41) | Tumor (n = 286) | p Value * | ||
|---|---|---|---|---|---|
| Mean ± SD | Median | Mean ± SD | MEDIAN | ||
| 1.43b ± 0.98 | 1.3842 | 3.82 ± 2.05 | 4.0342 | <0.001 | |
| TMEM211 Expression | Pair No. | Tumor Adjacent Normal | Tumor | p Value * | ||
|---|---|---|---|---|---|---|
| Mean ± SD | Median | Mean ± SD | Median | |||
| 26 | 1.34 ± 1.02 | 1.1866 | 3.82 ± 1.86 | 3.9703 | <0.001 | |
| Variable | ROC | No. (%) | CHR (95% CI) | p Value * | AHR (95% CI) | p Value † |
|---|---|---|---|---|---|---|
| Overall survival | ||||||
| TMEM211 | Low | 213 (78.6) | 1.00 | 1.00 | ||
| High | 58 (21.4) | 1.44 (0.84–2.46) | 0.187 | 1.69 (0.98–2.91) | 0.059 | |
| Progression-free interval survival | ||||||
| TMEM211 | Low | 89 (32.8) | 1.00 | 1.00 | ||
| High | 182 (67.2) | 1.72 (1.00–2.95) | 0.047 | 1.91 (1.11–3.27) | 0.019 | |
| Disease-specific survival | ||||||
| TMEM211 | Low | 139 (54.3) | 1.00 | 1.00 | ||
| High | 117 (45.7) | 1.81 (0.91–3.59) | 0.090 | 2.30 (1.15–4.60) | 0.019 | |
| Disease-free interval survival | ||||||
| TMEM211 | Low | 27 (28.4) | 1.00 | 1.00 | ||
| High | 68 (71.6) | 6.19 (0.81–47.41) | 0.079 | 6.31 (0.82–48.47) | 0.076 | |
| Variable | ROC | No. (%) | CHR (95% CI) | p Value * | AHR (95% CI) | p Value † |
|---|---|---|---|---|---|---|
| Sex | ||||||
| Female | Low | 69 (59.0) | 1 | 1 | ||
| High | 48 (41.0) | 0.72 (0.22–2.39) | 0.591 | 0.75 (0.23–2.51) | 0.645 a | |
| Male | Low | 70 (50.4) | 1 | |||
| High | 69 (49.6) | 3.70 (1.34–10.24) | 0.012 | 5.51 (1.89–16.09) | 0.002 a | |
| Age, yrs | ||||||
| ≤70 | Low | 84 (53.8) | 1 | 1 | ||
| High | 72 (46.2) | 0.96 (0.40–2.31) | 0.920 | 1.32 (0.54–3.23) | 0.546 a | |
| >70 | Low | 55 (55.0) | 1 | 1 | ||
| High | 45 (45.0) | 4.25 (1.18–15.23) | 0.026 | 4.95 (1.35–18.19) | 0.016 a | |
| AJCC pathological stage | ||||||
| I, II | Low | 76 (52.8) | 1 | |||
| High | 68 (47.2) | 2.16 (0.40–11.83) | 0.374 | ND | ND | |
| III, IV | Low | 63 (56.3) | 1 | |||
| High | 49 (43.8) | 2.20 (1.03–4.69) | 0.041 | ND | ND | |
| T classification | ||||||
| T1, T2 | Low | 24 (52.2) | 1 | 1 | ||
| High | 22 (47.8) | 0.013 (0.00–135,544.68) | 0.598 | 0.01 (0.00–139,945.54) | 0.598 b | |
| T3, T4 | Low | 115 (54.8) | 1 | 1 | ||
| High | 95 (45.2) | 2.08 (1.03–4.20) | 0.04 | 2.39 (1.18–4.85) | 0.016 b | |
| N classification | ||||||
| N0 | Low | 79 (53.0) | 1 | 1 | ||
| High | 70 (47.0) | 3.31 (0.67–16.41) | 0.143 | 3.31 (0.67–16.41) | 0.143 c | |
| N1, N2 | Low | 60 (56.1) | 1 | 1 | ||
| High | 47 (43.9) | 1.90 (0.87–4.14) | 0.109 | 1.91 (0.88–4.18) | 0.104 c |
| Variable | ROC | No. (%) | CHR (95% CI) | p Value * | AHR (95% CI) | p Value † |
|---|---|---|---|---|---|---|
| TMEM211 | Low | 139 (54.3) | 1.00 | 1.00 | ||
| High | 117 (45.7) | 1.81 (0.91–3.59) | 0.09 | 2.30 (1.15–4.60) | 0.019 a | |
| TWIST1 | Low | 144 (56.3) | 1.00 | 1.00 | ||
| High | 112(43.8) | 1.86(0.94–3.67) | 0.075 | 1.69 (0.86–3.35) | 0.130 a | |
| TMEM211 (L), Twist1 (L) | 83 (32.4) | 1.00 | 1.00 | |||
| TMEM211 (H), Twist1 (L) | 61 (23.8) | 1.14 (0.53–2.45) | 0.732 | 2.23 (0.79–6.28) | 0.129 b | |
| TMEM211 (L), Twist1 (H) | 56 (21.9) | 1.20 (0.54–2.65) | 0.657 | 2.33 (0.81–6.75) | 0.119 b | |
| TMEM211 (H), Twist1 (H) | 56 (21.9) | 1.89 (0.92–3.90) | 0.083 | 3.25 (1.19–8.85) | 0.021 b | |
| Snail | Low | 91 (35.5) | 1.00 | 1.00 | ||
| High | 165 (64.5) | 3.50 (1.36–9.05) | 0.010 | 2.97 (1.15–7.68) | 0.025 a | |
| TMEM211 (L), Snail (L) | 44 (17.2) | 1.00 | 1.00 | |||
| TMEM211 (H), Snail (L) | 47 (18.4) | 0.55 (0.19–1.56) | 0.258 | 3.68 (0.41–32.98) | 0.244 b | |
| TMEM211 (L), Snail (H) | 95 (37.1) | 1.02 (0.51–2.05) | 0.951 | 6.20 (0.81–47.40) | 0.079 b | |
| TMEM211 (H), Snail (H) | 70 (27.3) | 2.76 (1.40–5.46) | 0.003 | 11.84 (1.57–89.41) | 0.017 b | |
| Slug | Low | 108 (42.2) | 1.00 | 1.00 | ||
| High | 148 (57.8) | 2.39 (1.11–5.13) | 0.025 | 2.40 (1.12–5.14) | 0.025 a | |
| TMEM211 (L), Slug (L) | 57 (22.3) | 1.00 | 1.00 | |||
| TMEM211 (H), Slug (L) | 51 (19.9) | 0.81 (0.34–1.97) | 0.646 | 2.44 (0.61–9.77) | 0.208 b | |
| TMEM211 (L), Slug (H) | 82 (32.0) | 1.09 (0.53–2.24) | 0.812 | 3.06 (0.85–10.98) | 0.087 b | |
| TMEM211 (H), Slug (H) | 66 (25.8) | 2.32 (1.17–4.61) | 0.017 | 5.11 (1.47–17.86) | 0.011 b | |
| N-cad | Low | 119 (46.5) | 1.00 | 1.00 | ||
| High | 137 (53.5) | 2.67 (1.27–5.60) | 0.010 | 2.37 (1.12–4.99) | 0.024 a | |
| TMEM211 (L), N-cad (L) | 62 (24.2) | 1.00 | 1.00 | |||
| TMEM211 (H), N-cad (L) | 57 (22.3) | 0.87 (0.38–1.99) | 0.736 | 3.02 (0.78–11.72) | 0.110 b | |
| TMEM211 (L), N-cad (H) | 77 (30.1) | 1.33 (0.65–2.73) | 0.438 | 4.16 (1.15–14.96) | 0.029 b | |
| TMEM211 (H), N-cad (H) | 60 (23.4) | 2.32 (1.16–4.65) | 0.018 | 6.17 (1.75–21.80) | 0.005 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, Y.-F.; Wang, H.-H.; Shu, C.-W.; Tsai, W.-L.; Lee, C.-H.; Chen, C.-L.; Liu, P.-F. TMEM211 Promotes Tumor Progression and Metastasis in Colon Cancer. Curr. Issues Mol. Biol. 2023, 45, 4529-4543. https://doi.org/10.3390/cimb45060287
Chang Y-F, Wang H-H, Shu C-W, Tsai W-L, Lee C-H, Chen C-L, Liu P-F. TMEM211 Promotes Tumor Progression and Metastasis in Colon Cancer. Current Issues in Molecular Biology. 2023; 45(6):4529-4543. https://doi.org/10.3390/cimb45060287
Chicago/Turabian StyleChang, Yung-Fu, Hsing-Hsang Wang, Chih-Wen Shu, Wei-Lun Tsai, Cheng-Hsin Lee, Chun-Lin Chen, and Pei-Feng Liu. 2023. "TMEM211 Promotes Tumor Progression and Metastasis in Colon Cancer" Current Issues in Molecular Biology 45, no. 6: 4529-4543. https://doi.org/10.3390/cimb45060287