
Protein Profiling in Human Papillomavirus-Associated Cervical Carcinogenesis: Cornulin as a Biomarker for Disease Progression
 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Tissue Samples
2.2. Peptide Extraction from FFPE Tissue and Mass Spectrometry Analysis
2.3. Protein Identification
2.4. Protein Quantification
2.5. Bioinformatics Analysis
2.6. Immunohistochemistry of Annexin A2 (ANXA2) and Cornulin (CRNN) Proteins
2.7. Statistical Analysis
3. Results
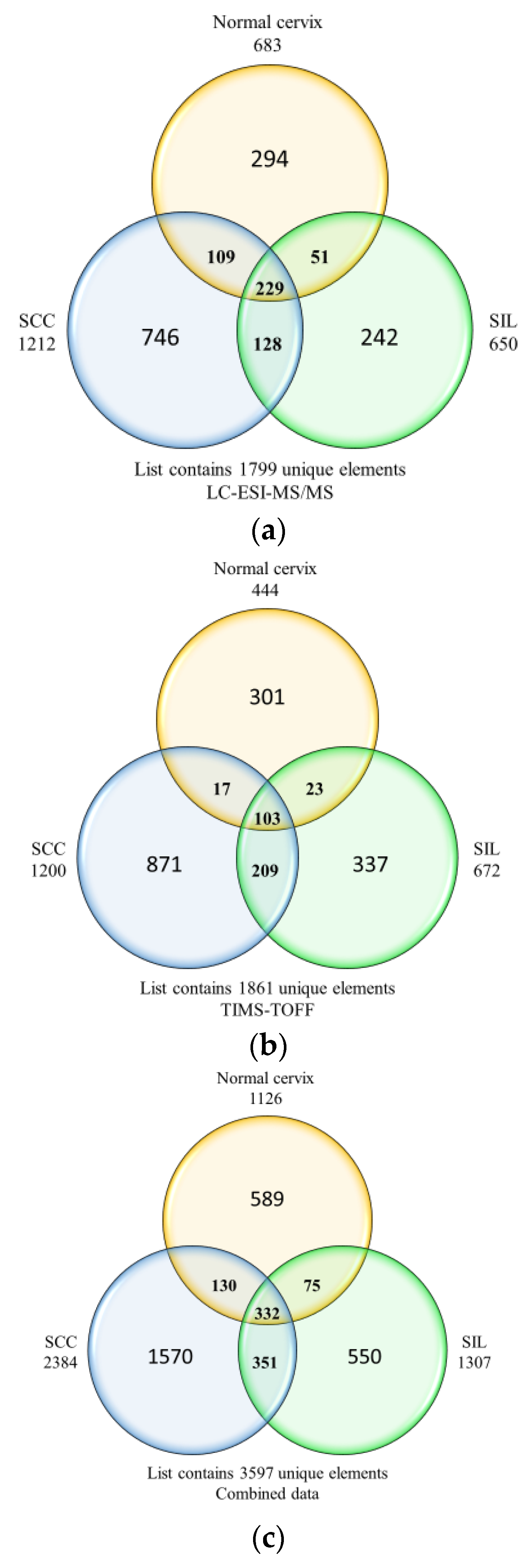
3.1. Protein Identification
3.2. Differentially Expressed Proteins
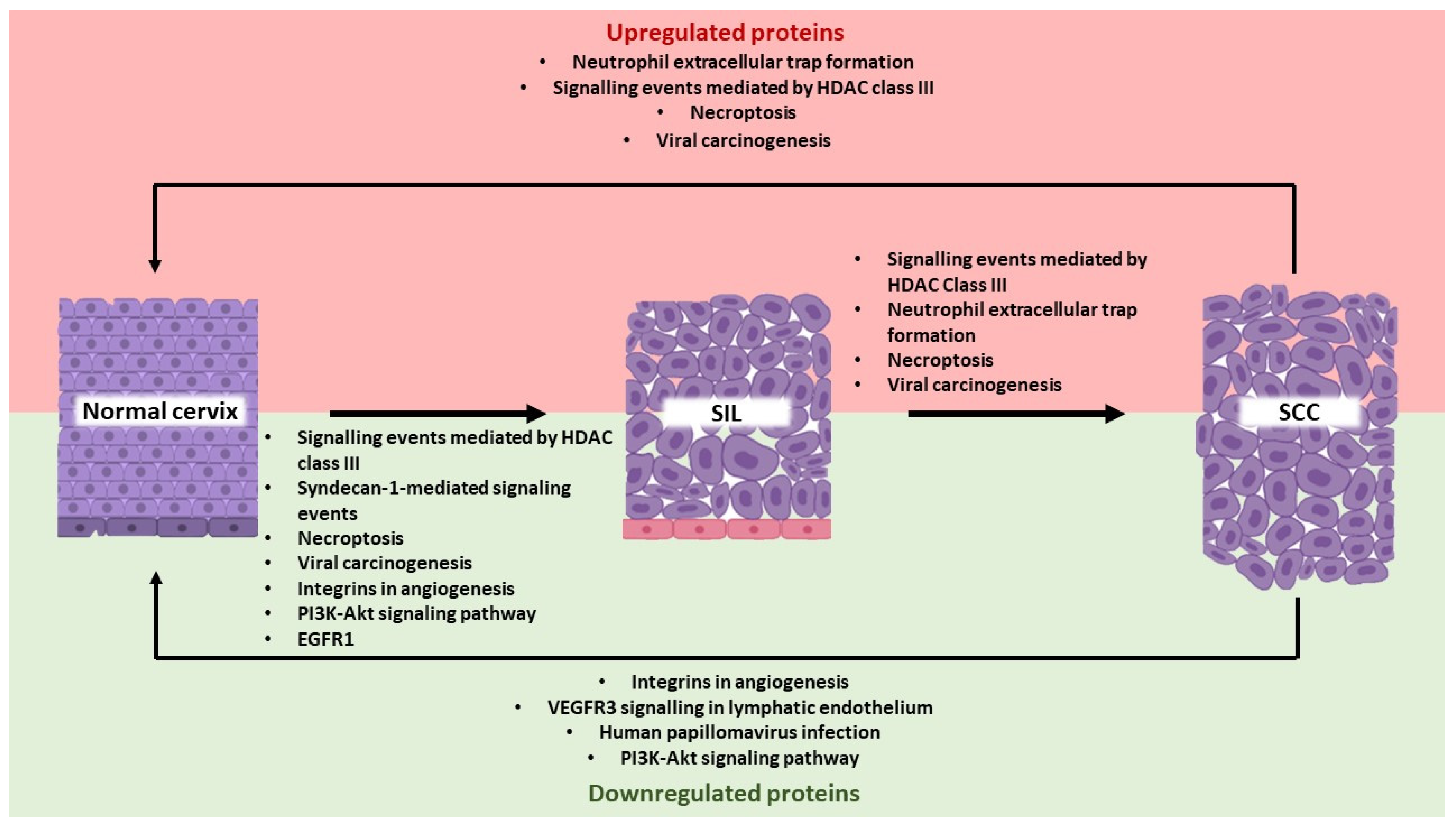
3.3. Gene Ontology, Pathways, and Protein-Protein Interaction Networks in Cervical Cancer Development
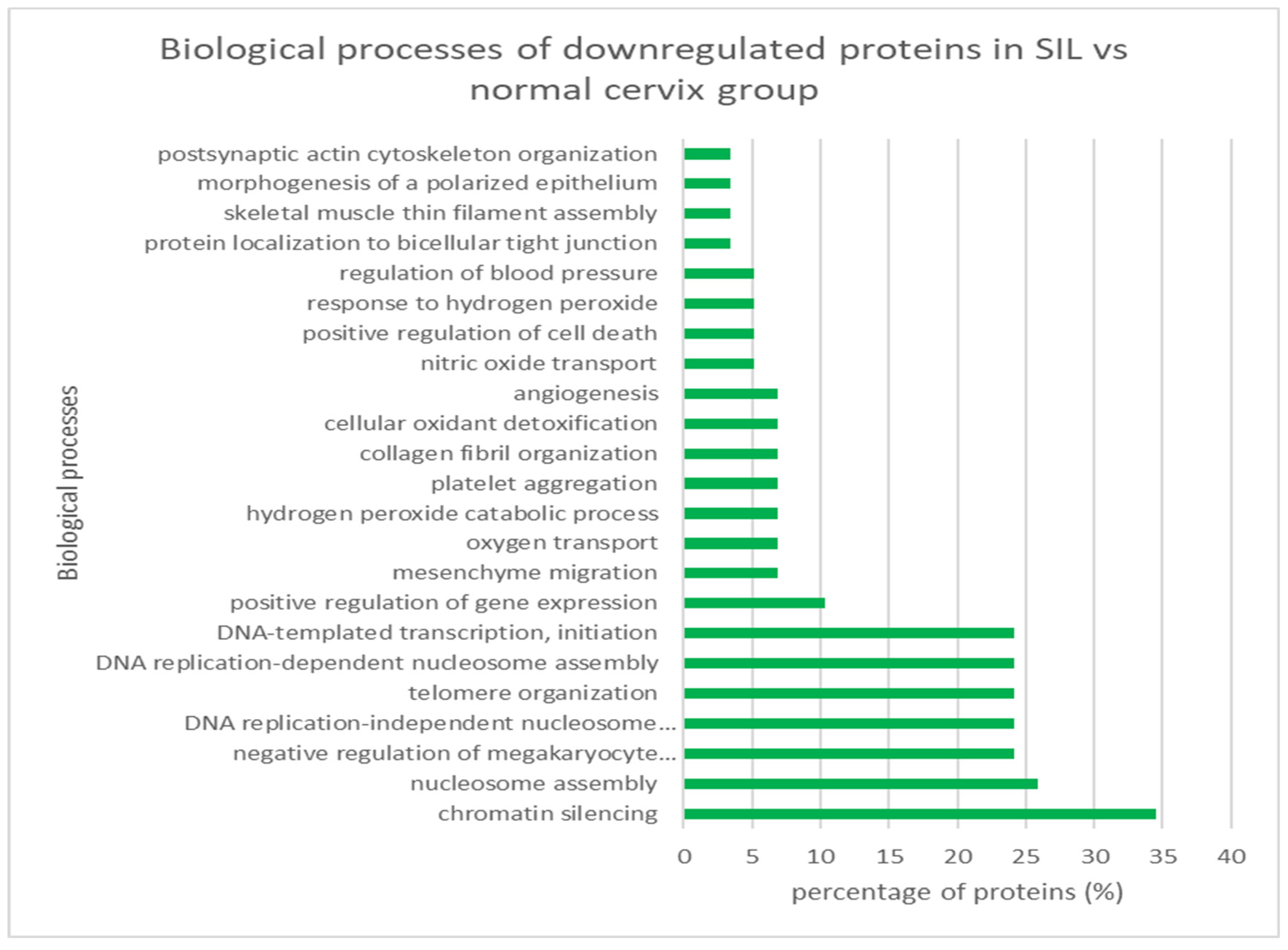
3.3.1. SIL vs. Normal Cervix
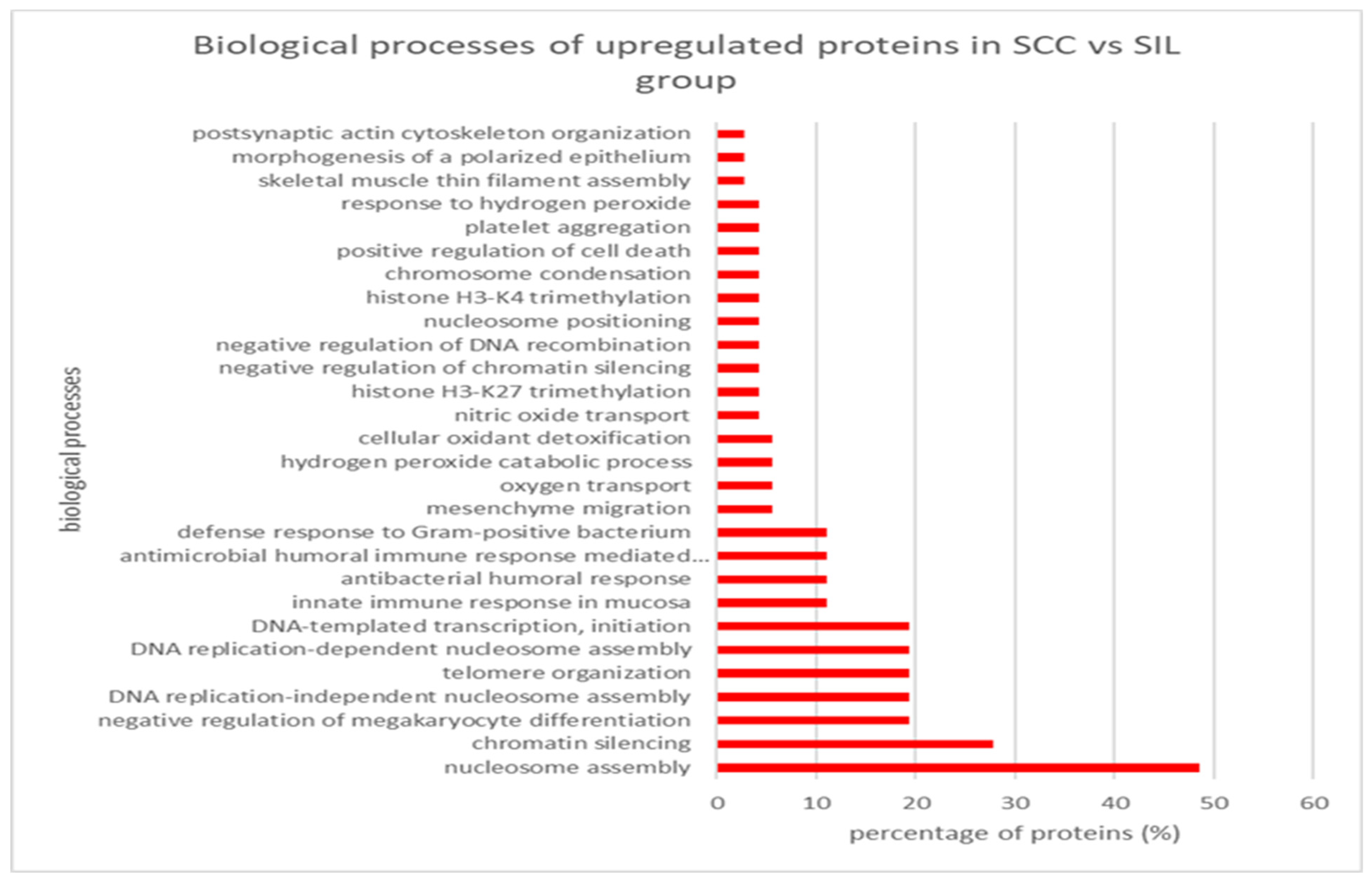
3.3.2. SCC vs. SIL
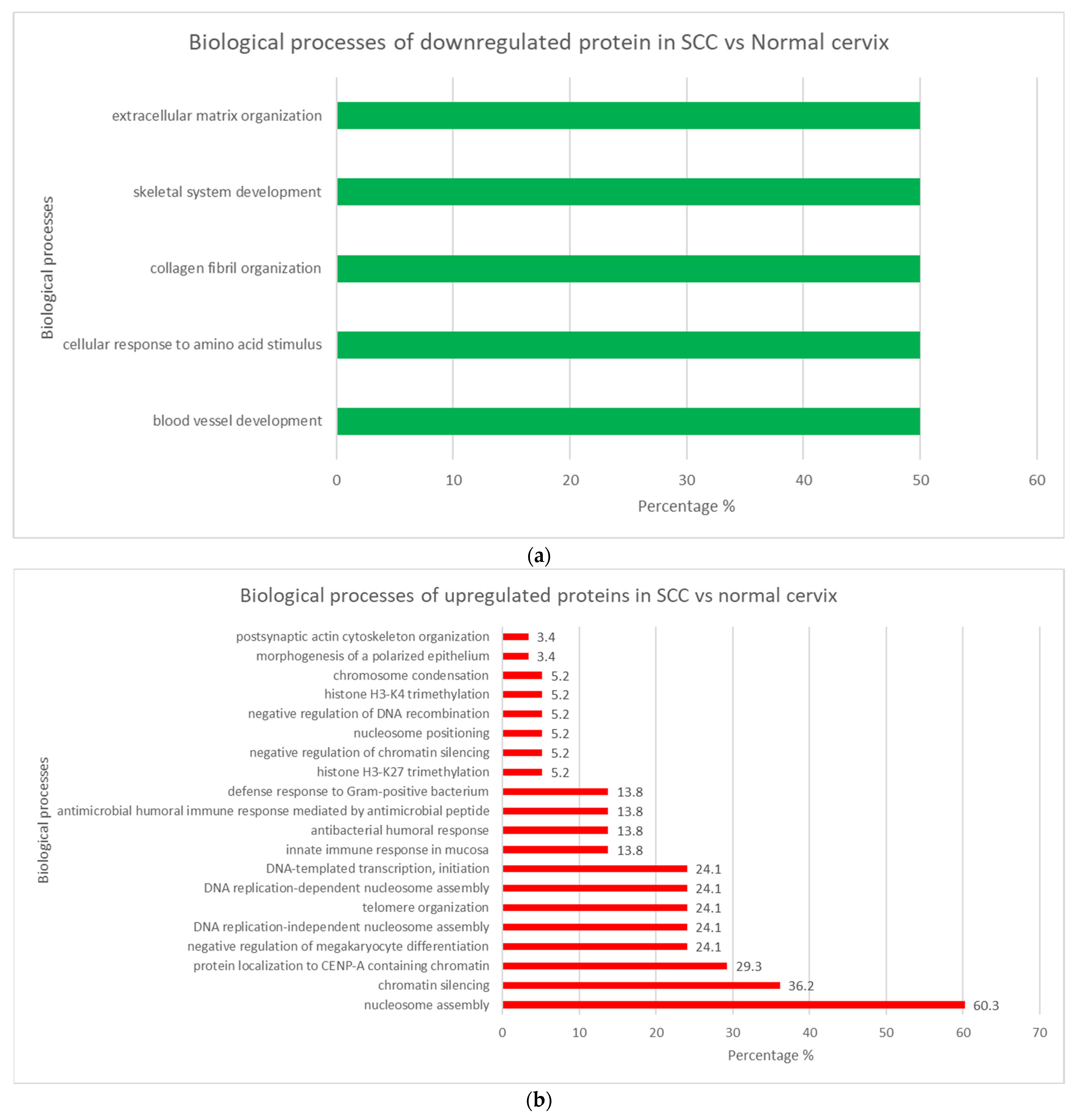
3.3.3. SCC vs. Normal Cervix
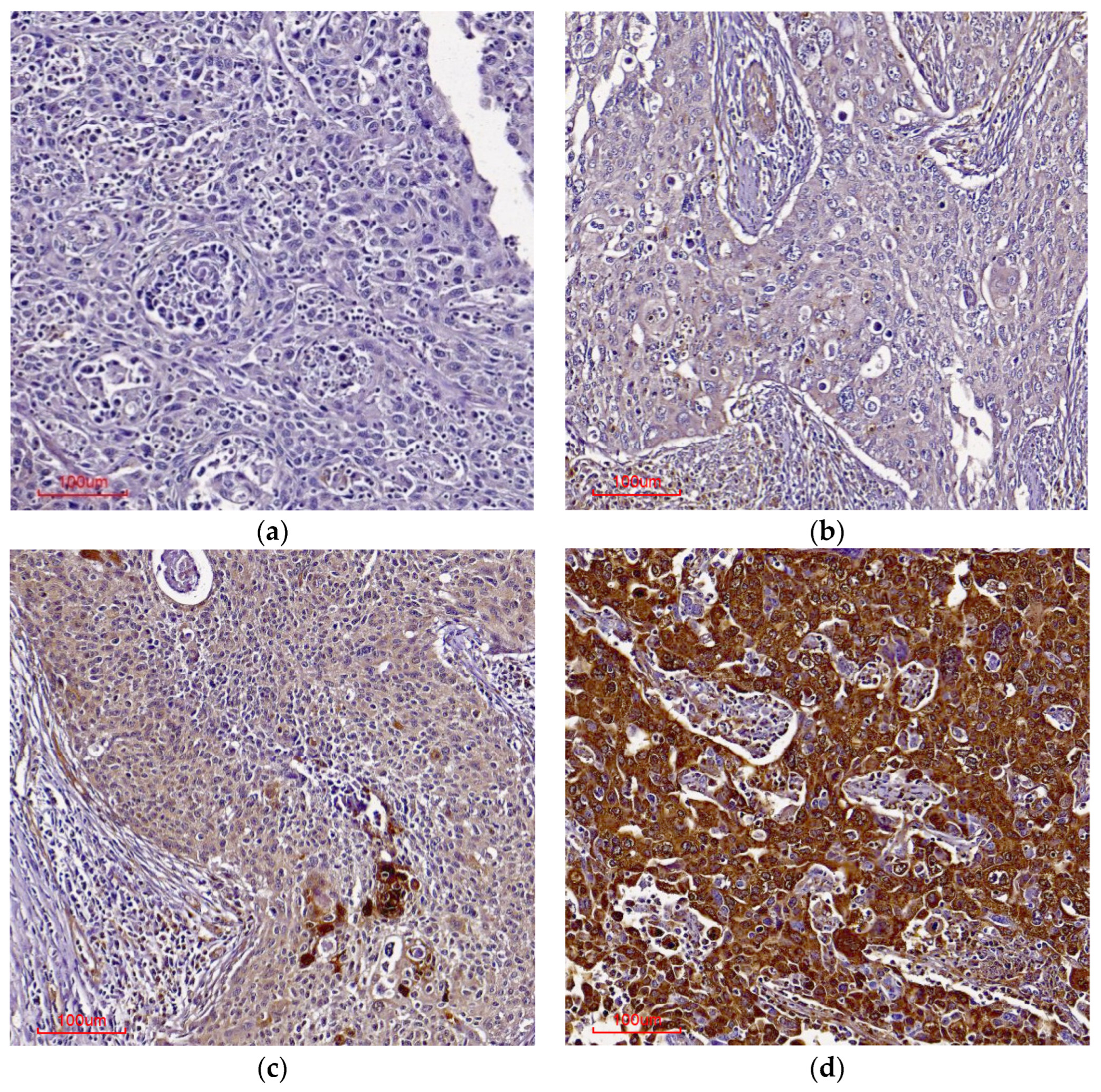
3.4. Annexin A2 (ANXA2) and Cornulin (CRNN) Immunohistochemistry
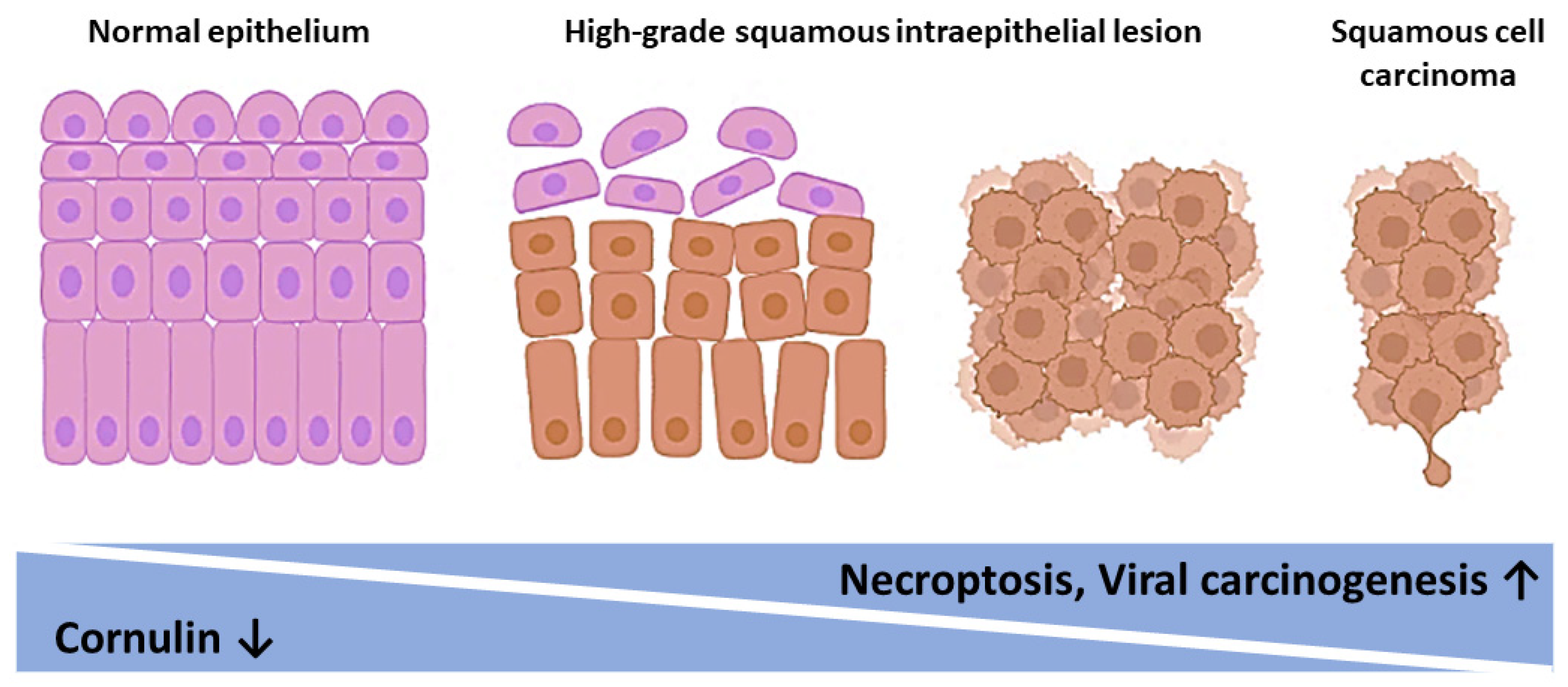
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Arbyn, M.; Weiderpass, E.; Bruni, L.; de Sanjosé, S.; Saraiya, M.; Ferlay, J.; Bray, F. Estimates of Incidence and Mortality of Cervical Cancer in 2018: A Worldwide Analysis. Lancet Glob. Health 2020, 8, e191–e203. [Google Scholar] [CrossRef] [PubMed]
- Zur Hausen, H. Papillomaviruses and Cancer: From Basic Studies to Clinical Application. Nat. Rev. Cancer 2002, 2, 342–350. [Google Scholar] [CrossRef] [PubMed]
- De Sanjose, S.; Quint, W.G.V.; Alemany, L.; Geraets, D.T.; Klaustermeier, J.E.; Lloveras, B.; Tous, S.; Felix, A.; Bravo, L.E.; Shin, H.-R.R.; et al. Human Papillomavirus Genotype Attribution in Invasive Cervical Cancer: A Retrospective Cross-Sectional Worldwide Study. Lancet Oncol. 2010, 11, 1048–1056. [Google Scholar] [CrossRef] [PubMed]
- Rogovskaya, S.I.; Shabalova, I.P.; Mikheeva, I.V.; Minkina, G.N.; Podzolkova, N.M.; Shipulina, O.Y.; Sultanov, S.N.; Kosenko, I.A.; Brotons, M.; Buttmann, N.; et al. Human Papillomavirus Prevalence and Type-Distribution, Cervical Cancer Screening Practices and Current Status of Vaccination Implementation in Russian Federation, the Western Countries of the Former Soviet Union, Caucasus Region and Central Asia. Vaccine 2013, 31, H46–H58. [Google Scholar] [CrossRef] [PubMed]
- Woodman, C.B.J.; Collins, S.I.; Young, L.S. The Natural History of Cervical HPV Infection: Unresolved Issues. Nat. Rev. Cancer 2007, 7, 11–22. [Google Scholar] [CrossRef]
- Muñoz, N.; Bosch, F.X.; de Sanjosé, S.; Herrero, R.; Castellsagué, X.; Shah, K.V.; Snijders, P.J.F.; Meijer, C.J.L.M. Epidemiologic Classification of Human Papillomavirus Types Associated with Cervical Cancer. N. Engl. J. Med. 2003, 348, 518–527. [Google Scholar] [CrossRef] [PubMed]
- Chan, C.K.; Aimagambetova, G.; Ukybassova, T.; Kongrtay, K.; Azizan, A. Human Papillomavirus Infection and Cervical Cancer: Epidemiology, Screening, and Vaccination-Review of Current Perspectives. J. Oncol. 2019, 2019, 3257939. [Google Scholar] [CrossRef]
- Oyervides-Muñoz, M.A.; Pérez-Maya, A.A.; Rodríguez-Gutiérrez, H.F.; Gómez-Macias, G.S.; Fajardo-Ramírez, O.R.; Treviño, V.; Barrera-Saldaña, H.A.; Garza-Rodríguez, M.L. Understanding the HPV Integration and Its Progression to Cervical Cancer. Infect. Genet. Evol. 2018, 61, 134–144. [Google Scholar] [CrossRef]
- zur Hausen, H. Papillomaviruses in the Causation of Human Cancers-a Brief Historical Account. Virology 2009, 384, 260–265. [Google Scholar] [CrossRef] [PubMed]
- McBride, A.A.; Warburton, A. The Role of Integration in Oncogenic Progression of HPV-Associated Cancers. PLoS Pathog. 2017, 13, 1006211. [Google Scholar] [CrossRef]
- Rusan, M.; Li, Y.Y.; Hammerman, P.S. Genomic Landscape of Human Papillomavirus-Associated Cancers. Clin. Cancer Res. 2015, 21, 2009–2019. [Google Scholar] [CrossRef] [PubMed]
- Scarth, J.A.; Patterson, M.R.; Morgan, E.L.; Macdonald, A. The Human Papillomavirus Oncoproteins: A Review of the Host Pathways Targeted on the Road to Transformation. J. Gen. Virol. 2021, 102, 001540. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, A.; Ansari, I.A. A Comprehensive Review on Cross-Talk of Human Papilloma Virus Oncoproteins and Developmental/Self-Renewal Pathways During the Pathogenesis of Uterine Cervical Cancer. Curr. Mol. Med. 2021, 21, 402–416. [Google Scholar] [CrossRef] [PubMed]
- Aslam, B.; Basit, M.; Nisar, M.A.; Khurshid, M.; Rasool, M.H. Proteomics: Technologies and Their Applications. J. Chromatogr. Sci. 2017, 55, 182–196. [Google Scholar] [CrossRef]
- Dasari, S.; Wudayagiri, R.; Valluru, L. Cervical Cancer: Biomarkers for Diagnosis and Treatment. Clin. Chim. Acta 2015, 445, 7–11. [Google Scholar] [CrossRef]
- Domon, B.; Aebersold, R. Mass Spectrometry and Protein Analysis. Science 2006, 312, 212–217. [Google Scholar] [CrossRef]
- Thompson, S.M.; Craven, R.A.; Nirmalan, N.J.; Harnden, P.; Selby, P.J.; Banks, R.E. Impact of Pre-Analytical Factors on the Proteomic Analysis of Formalin-Fixed Paraffin-Embedded Tissue. Proteom. Clin. Appl. 2013, 7, 241–251. [Google Scholar] [CrossRef]
- Hussen, B.M.; Abdullah, S.T.; Salihi, A.; Sabir, D.K.; Sidiq, K.R.; Rasul, M.F.; Hidayat, H.J.; Ghafouri-Fard, S.; Taheri, M.; Jamali, E. The Emerging Roles of NGS in Clinical Oncology and Personalized Medicine. Pathol. Pract. 2022, 230, 153760. [Google Scholar] [CrossRef]
- Rosai, J.; Ackerman, L. V The Pathology of Tumors, Part III: Grading, Staging & Classification. CA Cancer J. Clin. 1979, 29, 66–77. [Google Scholar]
- Grillo, F.; Bruzzone, M.; Pigozzi, S.; Prosapio, S.; Migliora, P.; Fiocca, R.; Mastracci, L. Immunohistochemistry on Old Archival Paraffin Blocks: Is There an Expiry Date? J. Clin. Pathol. 2017, 70, 988–993. [Google Scholar] [CrossRef]
- Catenacci, D.V.T.; Liao, W.L.; Zhao, L.; Whitcomb, E.; Henderson, L.; O’Day, E.; Xu, P.; Thyparambil, S.; Krizman, D.; Bengali, K.; et al. Mass-Spectrometry-Based Quantitation of Her2 in Gastroesophageal Tumor Tissue: Comparison to IHC and FISH. Gastric Cancer 2016, 19, 1066–1079. [Google Scholar] [CrossRef] [PubMed]
- Weißer, J.; Lai, Z.W.; Bronsert, P.; Kuehs, M.; Drendel, V.; Timme, S.; Kuesters, S.; Jilg, C.A.; Wellner, U.F.; Lassmann, S.; et al. Quantitative Proteomic Analysis of Formalin-Fixed, Paraffin-Embedded Clear Cell Renal Cell Carcinoma Tissue Using Stable Isotopic Dimethylation of Primary Amines. BMC Genom. 2015, 16, 559. [Google Scholar] [CrossRef] [PubMed]
- Arnouk, H.; Merkley, M.A.; Podolsky, R.H.; Stöppler, H.; Santos, C.; Álvarez, M.; Mariategui, J.; Ferris, D.; Lee, J.R.; Dynan, W.S. Characterization of Molecular Markers Indicative of Cervical Cancer Progression. Proteom. Clin. Appl. 2009, 3, 516–527. [Google Scholar] [CrossRef]
- Qiu, F.; Chen, F.; Liu, D.; Xu, J.; He, J.; Xiao, J.; Cao, L.; Huang, X. LC-MS/MS-based screening of new protein biomarkers for cervical precancerous lesions and cervical cancer. Nan Fang Yi Ke Da Xue Xue Bao 2019, 39, 13–22. [Google Scholar] [CrossRef]
- Taylor, J.R.; Skeate, J.G.; Martin Kast, W. Annexin A2 in Virus Infection. Front. Microbiol. 2018, 9, 2954. [Google Scholar] [CrossRef] [PubMed]
- Hajjar, K.A. The Biology of Annexin A2: From Vascular Fibrinolysis to Innate Immunity. Trans. Am. Clin. Climatol. Assoc. 2015, 126, 144–155. [Google Scholar]
- Schloer, S.; Pajonczyk, D.; Rescher, U. Annexins in Translational Research: Hidden Treasures to Be Found. Int. J. Mol. Sci. 2018, 19, 1781. [Google Scholar] [CrossRef]
- Hitchcock, J.K.; Katz, A.A.; Schäfer, G. Dynamic Reciprocity: The Role of Annexin A2 in Tissue Integrity. J. Cell Commun. Signal. 2014, 8, 125–133. [Google Scholar] [CrossRef]
- Bharadwaj, A.; Bydoun, M.; Holloway, R.; Waisman, D. Annexin A2 Heterotetramer: Structure and Function. Int. J. Mol. Sci. 2013, 14, 6259–6305. [Google Scholar] [CrossRef]
- Contzler, R.; Favre, B.; Huber, M.; Hohl, D. Cornulin, a New Member of the “Fused Gene” Family, Is Expressed during Epidermal Differentiation. J. Investig. Dermatol. 2005, 124, 990–997. [Google Scholar] [CrossRef]
- Kaur, R.; Chauhan, A.; Bhat, S.A.; Chatterjee, D.; Ghoshal, S.; Pal, A. Gene of the Month: Cornulin. J. Clin. Pathol. 2022, 75, 289–291. [Google Scholar] [CrossRef]
- Liu, Y.; Li, H.; Ban, Z.; Nai, M.; Yang, L.; Chen, Y.; Xu, Y. Annexin A2 Inhibition Suppresses Ovarian Cancer Progression via Regulating β-Catenin/EMT. Oncol. Rep. 2017, 37, 3643–3650. [Google Scholar] [CrossRef]
- Deng, Y.; Chen, C.; Hua, M.; Xi, Q.; Liu, R.; Yang, S.; Liu, J.; Zhong, J.; Tang, M.; Lu, S.; et al. Annexin A2 Plays a Critical Role in Epithelial Ovarian Cancer. Arch. Gynecol. Obstet. 2015, 292, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Pawar, H.; Maharudraiah, J.; Kashyap, M.K.; Sharma, J.; Srikanth, S.M.; Choudhary, R.; Chavan, S.; Sathe, G.; Manju, H.C.; Kumar, K.V.V.; et al. Downregulation of Cornulin in Esophageal Squamous Cell Carcinoma. Acta Histochem. 2013, 115, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Merkley, M.A.; Weinberger, P.M.; Jackson, L.L.; Podolsky, R.H.; Lee, J.R.; Dynan, W.S. 2D-DIGE Proteomic Characterization of Head and Neck Squamous Cell Carcinoma. Otolaryngol. Head Neck Surg. 2009, 141, 626–632. [Google Scholar] [CrossRef]
- Olusola, P.; Banerjee, H.N.; Philley, J.V.; Dasgupta, S. Human Papilloma Virus-Associated Cervical Cancer and Health Disparities. Cells 2019, 8, 622. [Google Scholar] [CrossRef] [PubMed]
- Whittaker, K.; Burgess, R.; Jones, V.; Yang, Y.; Zhou, W.; Luo, S.; Wilson, J.; Huang, R.P. Quantitative Proteomic Analyses in Blood: A Window to Human Health and Disease. J. Leukoc. Biol. 2019, 106, 759–775. [Google Scholar] [CrossRef] [PubMed]
- Giusti, L.; Angeloni, C.; Lucacchini, A. Update on Proteomic Studies of Formalin-Fixed Paraffin-Embedded Tissues. Expert Rev. Proteomics 2019, 16, 513–520. [Google Scholar] [CrossRef] [PubMed]
- Bannister, A.J.; Kouzarides, T. Regulation of Chromatin by Histone Modifications. Cell Res. 2011, 21, 381–395. [Google Scholar] [CrossRef]
- Li, X.; Tian, R.; Gao, H.; Yang, Y.; Williams, B.R.G.; Gantier, M.P.; McMillan, N.A.J.; Xu, D.; Hu, Y.; Gao, Y. Identification of a Histone Family Gene Signature for Predicting the Prognosis of Cervical Cancer Patients. Sci. Rep. 2017, 7, 16495. [Google Scholar] [CrossRef]
- Peng, X.; Wang, J.; Peng, W.; Wu, F.X.; Pan, Y. Protein-Protein Interactions: Detection, Reliability Assessment and Applications. Brief. Bioinform. 2017, 18, 798–819. [Google Scholar] [CrossRef] [PubMed]
- Audia, J.E.; Campbell, R.M. Histone Modifications and Cancer. Cold Spring Harb. Perspect. Biol. 2016, 8, 019521. [Google Scholar] [CrossRef]
- Stoll, G.; Ma, Y.; Yang, H.; Kepp, O.; Zitvogel, L.; Kroemer, G. Pro-Necrotic Molecules Impact Local Immunosurveillance in Human Breast Cancer. Oncoimmunology 2017, 6, 1299302. [Google Scholar] [CrossRef]
- Seehawer, M.; Heinzmann, F.; D’Artista, L.; Harbig, J.; Roux, P.F.; Hoenicke, L.; Dang, H.; Klotz, S.; Robinson, L.; Doré, G.; et al. Necroptosis Microenvironment Directs Lineage Commitment in Liver Cancer. Nature 2018, 562, 69–75. [Google Scholar] [CrossRef]
- Galluzzi, L.; Vitale, I.; Aaronson, S.A.; Abrams, J.M.; Adam, D.; Agostinis, P.; Alnemri, E.S.; Altucci, L.; Amelio, I.; Andrews, D.W.; et al. Molecular Mechanisms of Cell Death: Recommendations of the Nomenclature Committee on Cell Death 2018. Cell Death Differ. 2018, 25, 486–541. [Google Scholar] [CrossRef]
- Vanden Berghe, T.; Kaiser, W.J.; Bertrand, M.J.M.; Vandenabeele, P. Molecular Crosstalk between Apoptosis, Necroptosis, and Survival Signaling. Mol. Cell. Oncol. 2015, 2, 975093. [Google Scholar] [CrossRef]
- Fontecha, N.; Basaras, M.; Hernáez, S.; Andía, D.; Cisterna, R. Assessment of Human Papillomavirus E6/E7 Oncogene Expression as Cervical Disease Biomarker. BMC Cancer 2016, 16, 852. [Google Scholar] [CrossRef] [PubMed]
- Dziduszko, A.; Ozbun, M.A. Annexin A2 and S100A10 Regulate Human Papillomavirus Type 16 Entry and Intracellular Trafficking in Human Keratinocytes. J. Virol. 2013, 87, 7502–7515. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Liao, G.; Li, G. Regulatory Effects of COL1A1 on Apoptosis Induced by Radiation in Cervical Cancer Cells. Cancer Cell Int. 2017, 17, 1–9. [Google Scholar] [CrossRef]
- Bahrami, A.; Hasanzadeh, M.; Hassanian, S.M.; ShahidSales, S.; Ghayour-Mobarhan, M.; Ferns, G.A.; Avan, A. The Potential Value of the PI3K/Akt/MTOR Signaling Pathway for Assessing Prognosis in Cervical Cancer and as a Target for Therapy. J. Cell. Biochem. 2017, 118, 4163–4169. [Google Scholar] [CrossRef]
- Jiang, N.; Dai, Q.; Su, X.; Fu, J.; Feng, X.; Peng, J. Role of PI3K/AKT Pathway in Cancer: The Framework of Malignant Behavior. Mol. Biol. Rep. 2020, 47, 4587–4629. [Google Scholar] [CrossRef]
- Liu, R.; Chen, Y.; Liu, G.; Li, C.; Song, Y.; Cao, Z.; Li, W.; Hu, J.; Lu, C.; Liu, Y. PI3K/AKT Pathway as a Key Link Modulates the Multidrug Resistance of Cancers. Cell Death Dis. 2020, 11, 797. [Google Scholar] [CrossRef] [PubMed]
- Manning, B.D.; Cantley, L.C. AKT/PKB Signaling: Navigating Downstream. Cell 2007, 129, 1261–1274. [Google Scholar] [CrossRef]
- Murakami, I.; Iwata, T.; Morisada, T.; Tanaka, K.; Aoki, D. Nucleosome Positioning on Episomal Human Papillomavirus Dna in Cultured Cells. Pathogens 2021, 10, 772. [Google Scholar] [CrossRef] [PubMed]
- Jorch, S.K.; Kubes, P. An Emerging Role for Neutrophil Extracellular Traps in Noninfectious Disease. Nat. Med. 2017, 23, 279–287. [Google Scholar] [CrossRef]
- Metzler, K.D.; Goosmann, C.; Lubojemska, A.; Zychlinsky, A.; Papayannopoulos, V. Myeloperoxidase-Containing Complex Regulates Neutrophil Elastase Release and Actin Dynamics during NETosis. Cell Rep. 2014, 8, 883–896. [Google Scholar] [CrossRef]
- Park, J.; Wysocki, R.W.; Amoozgar, Z.; Maiorino, L.; Fein, M.R.; Jorns, J.; Schott, A.F.; Kinugasa-Katayama, Y.; Lee, Y.; Won, N.H.; et al. Cancer Cells Induce Metastasis-Supporting Neutrophil Extracellular DNA Traps. Sci. Transl. Med. 2016, 8, 1711. [Google Scholar] [CrossRef]
- Snoderly, H.T.; Boone, B.A.; Bennewitz, M.F. Neutrophil Extracellular Traps in Breast Cancer and beyond: Current Perspectives on NET Stimuli, Thrombosis and Metastasis, and Clinical Utility for Diagnosis and Treatment. Breast Cancer Res. 2019, 21, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Yan, B.; Dai, X.; Ma, Q.; Wu, X. Stromal Neutrophil Extracellular Trap Density Is an Independent Prognostic Factor for Cervical Cancer Recurrence. Front. Oncol. 2021, 11, 659445. [Google Scholar] [CrossRef]
- Dobbs, S.; Hewett, P.; Johnson, I.; Carmichael, J.; Murray, C. Angiogenesis Is Associated with Vascular Endothelial Growth Factor Expression in Cervical Intraepithelial Neoplasia. Br. J. Radiol. 1998, 71, 106–107. [Google Scholar] [CrossRef]
- Hammes, L.S.; Tekmal, R.R.; Naud, P.; Edelweiss, M.I.; Kirma, N.; Valente, P.T.; Syrjänen, K.J.; Cunha-Filho, J.S. Up-Regulation of VEGF, c-Fms and COX-2 Expression Correlates with Severity of Cervical Cancer Precursor (CIN) Lesions and Invasive Disease. Gynecol. Oncol. 2008, 110, 445–451. [Google Scholar] [CrossRef] [PubMed]
- Sideras, K.; Bots, S.J.; Biermann, K.; Sprengers, D.; Polak, W.G.; Ijzermans, J.N.M.; De Man, R.A.; Pan, Q.; Sleijfer, S.; Bruno, M.J.; et al. Tumour Antigen Expression in Hepatocellular Carcinoma in a Low-Endemic Western Area. Br. J. Cancer 2015, 112, 1911–1920. [Google Scholar] [CrossRef] [PubMed]
- Kondo, T.; Oka, T.; Sato, H.; Shinnou, Y.; Washio, K. Accumulation of Aberrant CpG Hypermethylation by Helicobacter Pylori Infection Promotes Development. Int. J. Oncol. 2009, 35, 547–557. [Google Scholar] [CrossRef] [PubMed]
- Zhao, P.; Zhang, W.; Tang, J.; Ma, X.K.; Dai, J.Y.; Li, Y.; Jiang, J.L.; Zhang, S.H.; Chen, Z.N. Annexin II Promotes Invasion and Migration of Human Hepatocellular Carcinoma Cells in Vitro via Its Interaction with HAb18G/CD147. Cancer Sci. 2010, 101, 387–395. [Google Scholar] [CrossRef] [PubMed]
- Lokman, N.A.; Elder, A.S.F.; Ween, M.P.; Pyragius, C.E.; Hoffmann, P.; Oehler, M.K.; Ricciardelli, C. Annexin A2 Is Regulated by Ovarian Cancer-Peritoneal Cell Interactions and Promotes Metastasis. Oncotarget 2013, 4, 1199–1211. [Google Scholar] [CrossRef]
- Choi, C.H.; Chung, J.Y.; Chung, E.J.; Sears, J.D.; Lee, J.W.; Bae, D.S.; Hewitt, S.M. Prognostic Significance of Annexin A2 and Annexin A4 Expression in Patients with Cervical Cancer. BMC Cancer 2016, 16, 448. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Jiang, C.; Pang, L.; Jia, W.; Wang, C.; Gao, X.; Zhang, X.; Dang, H.; Ren, Y. ANXA2 Is a Potential Marker for the Diagnosis of Human Cervical Cancer. Biomark. Med. 2021, 15, 57–67. [Google Scholar] [CrossRef] [PubMed]
- Balasubramaniam, S.D.; Wong, K.K.; Oon, C.E.; Balakrishnan, V.; Kaur, G. Comparative Transcriptomic Profiling in HPV-Associated Cervical Carcinogenesis: Implication of MHC Class II and Immunoglobulin Heavy Chain Genes. Life Sci. 2020, 256, 118026. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.A.; Shim, J.H.; Kho, C.W.; Park, S.G.; Park, B.C.; Kim, J.W.; Lim, J.S.; Choe, Y.K.; Paik, S.G.; Yoon, D.Y. Protein Profiling and Identification of Modulators Regulated by the E7 Oncogene in the C33A Cell Line by Proteomics and Genomics. Proteomics 2004, 4, 839–848. [Google Scholar] [CrossRef]
- Li, C.; Xiao, L.; Jia, J.; Li, F.; Wang, X.; Duan, Q.; Jing, H.; Yang, P.; Chen, C.; Wang, Q.; et al. Cornulin Is Induced in Psoriasis Lesions and Promotes Keratinocyte Proliferation via Phosphoinositide 3-Kinase/Akt Pathways. J. Investig. Dermatol. 2019, 139, 71–80. [Google Scholar] [CrossRef]
- Sahu, N.; Grandis, J.R. New Advances in Molecular Approaches to Head and Neck Squamous Cell Carcinoma. Anticancer. Drugs 2011, 22, 656–664. [Google Scholar] [CrossRef] [PubMed]
- Imai, F.L.; Uzawa, K.; Nimura, Y.; Moriya, T.; Imai, M.A.; Shiiba, M.; Bukawa, H.; Yokoe, H.; Tanzawa, H. Chromosome 1 Open Reading Frame 10 (C1orf10) Gene Is Frequently down-Regulated and Inhibits Cell Proliferation in Oral Squamous Cell Carcinoma. Int. J. Biochem. Cell Biol. 2005, 37, 1641–1655. [Google Scholar] [CrossRef]
- Xiao, H.; Langerman, A.; Zhang, Y.; Khalid, O.; Hu, S.; Cao, C.X.; Lingen, M.W.; Wong, D.T.W. Quantitative Proteomic Analysis of Microdissected Oral Epithelium for Cancer Biomarker Discovery. Oral Oncol. 2015, 51, 1011–1019. [Google Scholar] [CrossRef] [PubMed]
- Santosh, N.; McNamara, K.K.; Beck, F.M.; Kalmar, J.R. Expression of Cornulin in Oral Premalignant Lesions. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. 2019, 127, 526–534. [Google Scholar] [CrossRef] [PubMed]
- Balasubramaniam, S.D.; Balakrishnan, V.; Oon, C.E.; Kaur, G. Gene Expression Profiling of HPV-Associated Cervical Carcinogenesis in Formalin-Fixed Paraffin-Embedded (FFPE) Tissues Using the NanoString NCounterTM Platform. Gene 2022, 825, 146385. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Histological Groups | Total Number of Differentially Expressed Proteins | Number of Upregulated Proteins | Number of Downregulated Proteins |
|---|---|---|---|
| SIL vs. normal cervix | 39 | 0 | 39 |
| SCC vs. SIL | 51 | 51 | 0 |
| SCC vs. normal cervix | 53 | 49 | 4 |
| Pathways | Percentage of Proteins (%) | p-Value | Pathway Source |
|---|---|---|---|
| Signaling events mediated by HDAC class III | 35.90% | 1.48 × 10−24 | PID |
| Hemoglobins chaperone | 23.10% | 2.00 × 10−5 | BioCarta |
| Neutrophil extracellular trap formation | 19.50% | 8.31 × 10−56 | KEGG |
| Necroptosis | 13.20% | 1.27 × 10−26 | KEGG |
| Cell to cell adhesion signaling | 12.50% | 0.0334 | BioCarta |
| Syndecan-1-mediated signaling events | 9.30% | 3.16 × 10−5 | PID |
| Viral carcinogenesis | 7.40% | 3.86 × 10−15 | KEGG |
| Integrins in angiogenesis | 6.30% | 0.000143 | PID |
| Beta1 integrin cell surface interactions | 6.10% | 0.000171 | PID |
| Protein digestion and absorption | 4.90% | 7.27 × 10−5 | KEGG |
| Aurora B signaling | 4.90% | 0.013 | PID |
| ECM-receptor interaction | 4.50% | 0.000518 | KEGG |
| Focal adhesion | 3.50% | 2.10 × 10−5 | KEGG |
| Caspase cascade in apoptosis | 3.50% | 0.0242 | PID |
| Viral myocarditis | 3.30% | 0.0267 | KEGG |
| AP-1 transcription factor network | 2.90% | 0.0354 | PID |
| Prolactin | 2.90% | 0.0354 | NetPath |
| Adherens junction | 2.80% | 0.0364 | KEGG |
| Alpha6Beta4Integrin | 2.80% | 0.0364 | NetPath |
| Bacterial invasion of epithelial cells | 2.60% | 0.0421 | KEGG |
| Proteoglycans in cancer | 2.40% | 0.00172 | KEGG |
| Platelet activation | 2.40% | 0.0156 | KEGG |
| Hippo signaling pathway | 1.90% | 0.0288 | KEGG |
| PI3K-Akt signaling pathway | 1.40% | 0.0168 | KEGG |
| EGFR1 | 1.10% | 0.043 | NetPath |
| Pathway | Percentage of Proteins (%) | p-Value | Pathway Source |
|---|---|---|---|
| Signaling events mediated by HDAC Class III | 38.50% | 4.05 × 10−25 | PID |
| Neutrophil extracellular trap formation | 28.90% | 3.72 × 10−90 | KEGG |
| Hemoglobins chaperone | 23.10% | 4.04 × 10−5 | BioCarta |
| Viral carcinogenesis | 16.20% | 1.31 × 10−41 | KEGG |
| Necroptosis | 13.20% | 3.96 × 10−24 | KEGG |
| Cell to cell adhesion signaling | 12.50% | 0.042 | BioCarta |
| Viral myocarditis | 3.30% | 0.0409 | KEGG |
| Upregulated Proteins | |||
|---|---|---|---|
| Pathway | Percentage of Proteins (%) | p-Value | Pathway Source |
| Neutrophil extracellular trap formation | 28.9% | 9.73 × 10−102 | KEGG |
| Viral carcinogenesis | 15.7% | 7.71 × 10−44 | KEGG |
| Signaling events mediated by HDAC Class III | 13.2% | 1.97 × 10−26 | KEGG |
| Necroptosis | 4.0% | 0.0196 | KEGG |
| Viral myocarditis | 2.8% | 0.0375 | KEGG |
| Adherens junction | 2.6% | 0.0425 | KEGG |
| Bacterial invasion of epithelial cells | 2.6% | 0.0435 | KEGG |
| Downregulated proteins | |||
| VEGFR3 signaling in lymphatic endothelium | 8.00% | 9.93 × 10−6 | PID |
| Syndecan-1-mediated signaling events | 7.00% | 3.04 × 10−8 | PID |
| Integrins in angiogenesis | 4.80% | 9.78 × 10−8 | PID |
| Beta1 integrin cell surface interactions | 4.50% | 1.13 × 10−7 | PID |
| Beta3 integrin cell surface interactions | 4.50% | 3.13 × 10−5 | PID |
| ECM-receptor interaction | 3.40% | 2.70 × 10−7 | KEGG |
| IL4-mediated signaling events | 3.10% | 6.87 × 10−5 | PID |
| Protein digestion and absorption | 2.90% | 4.36 × 10−7 | KEGG |
| Platelet activation | 1.60% | 0.00025 | KEGG |
| Relaxin signaling pathway | 1.60% | 0.00027 | KEGG |
| Focal adhesion | 1.50% | 3.28 × 10−6 | KEGG |
| Proteoglycans in cancer | 1.00% | 0.00069 | KEGG |
| Human papillomavirus infection | 0.90% | 1.47 × 10−5 | KEGG |
| PI3K-Akt signaling pathway | 0.80% | 1.81 × 10−5 | KEGG |
| ANXA2 Histoscore | CRNN Histoscore | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Parameters | Negative (n) | Low (n) | High (n) | Total (n) | p-Value | Parameters | Negative (n) | Low (n) | High (n) | Total (n) | p-Value |
| Age (years) | Age (years) | ||||||||||
| <50 | 3 | 4 | 64 | 71 | 0.043 | <50 | 14 | 28 | 30 | 72 | 0.552 |
| ≥50 | 9 | 1 | 44 | 54 | ≥50 | 7 | 25 | 22 | 54 | ||
| SIL grade | SIL grade | ||||||||||
| 1 | 0 | 0 | 10 | 10 | 0.110 | 1 | 1 | 1 | 8 | 10 | 0.010 |
| 2 | 0 | 0 | 4 | 4 | 2 | 1 | 0 | 3 | 4 | ||
| 3 | 0 | 2 | 5 | 7 | 3 | 2 | 5 | 0 | 7 | ||
| SCC stage | SCC stage | ||||||||||
| Stage I | 1 | 2 | 27 | 30 | 0.444 | Stage I | 6 | 13 | 11 | 30 | 0.095 |
| Stage IA | 0 | 0 | 1 | 1 | Stage IA | 0 | 0 | 1 | 1 | ||
| Stage IB | 3 | 0 | 19 | 22 | Stage IB | 4 | 9 | 9 | 22 | ||
| Stage IC | 0 | 0 | 1 | 1 | Stage IC | 1 | 0 | 0 | 1 | ||
| Stage II | 0 | 0 | 6 | 6 | Stage II | 0 | 5 | 1 | 6 | ||
| Stage IIA | 0 | 0 | 19 | 19 | Stage IIA | 0 | 13 | 6 | 19 | ||
| Stage IIB | 0 | 0 | 11 | 11 | Stage IIB | 3 | 6 | 3 | 12 | ||
| Stage III | 0 | 0 | 1 | 1 | Stage III | 1 | 0 | 0 | 1 | ||
| Stage IIIB | 0 | 1 | 3 | 4 | Stage IIIB | 2 | 1 | 1 | 4 | ||
| Histological Group | Negative n | Histoscore Low n | High n | Total n | Comparison between all Histological Groups p-Value | Histological Group vs. Normal Cervix p-Value | SCC vs. SIL p-Value |
|---|---|---|---|---|---|---|---|
| ANXA2 protein | |||||||
| Normal cervix | 1 | 0 | 7 | 8 | |||
| SIL | 0 | 2 | 19 | 21 | 0.351 | 0.183 | |
| SCC | 4 | 3 | 89 | 96 | 0.512 | 0.281 | |
| Total | 5 | 5 | 115 | 125 | |||
| CRNN protein | |||||||
| Normal cervix | 0 | 0 | 8 | 8 | |||
| SIL | 4 | 6 | 11 | 21 | 0.004 | 0.055 | |
| SCC | 18 | 47 | 33 | 98 | 0.001 | 0.212 | |
| Total | 22 | 53 | 52 | 127 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kumarasamy, G.; Ismail, M.N.; Tuan Sharif, S.E.; Desire, C.; Mittal, P.; Hoffmann, P.; Kaur, G. Protein Profiling in Human Papillomavirus-Associated Cervical Carcinogenesis: Cornulin as a Biomarker for Disease Progression. Curr. Issues Mol. Biol. 2023, 45, 3603-3627. https://doi.org/10.3390/cimb45040235
Kumarasamy G, Ismail MN, Tuan Sharif SE, Desire C, Mittal P, Hoffmann P, Kaur G. Protein Profiling in Human Papillomavirus-Associated Cervical Carcinogenesis: Cornulin as a Biomarker for Disease Progression. Current Issues in Molecular Biology. 2023; 45(4):3603-3627. https://doi.org/10.3390/cimb45040235
Chicago/Turabian StyleKumarasamy, Gaayathri, Mohd Nazri Ismail, Sharifah Emilia Tuan Sharif, Christopher Desire, Parul Mittal, Peter Hoffmann, and Gurjeet Kaur. 2023. "Protein Profiling in Human Papillomavirus-Associated Cervical Carcinogenesis: Cornulin as a Biomarker for Disease Progression" Current Issues in Molecular Biology 45, no. 4: 3603-3627. https://doi.org/10.3390/cimb45040235