
Brassica napus Haploid and Double Haploid Production and Its Latest Applications

Abstract
:1. Introduction
2. Double Haploid
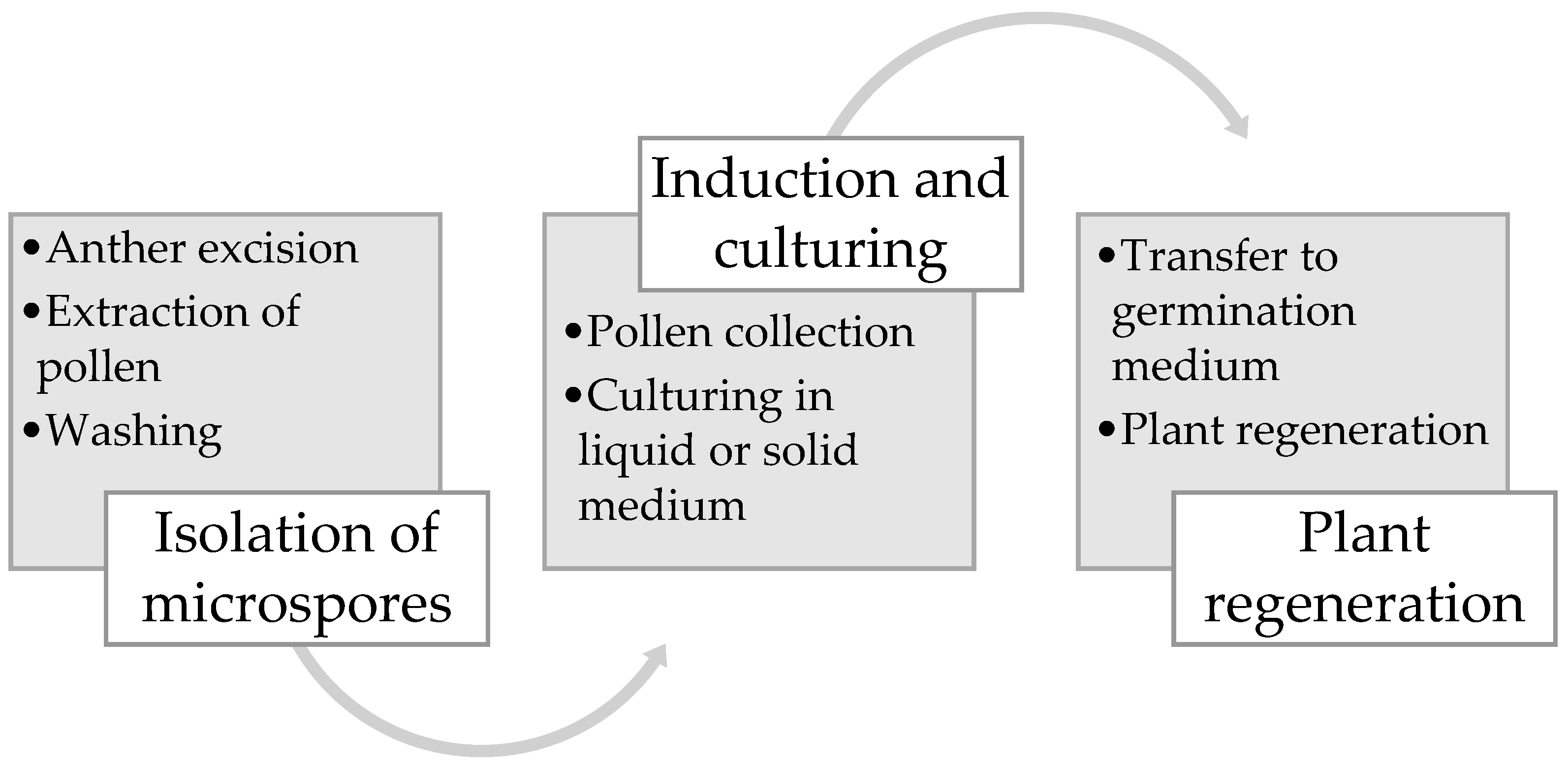
3. Haploid/Double Haploid Induction Methods
4. Brassica napus Microspore Embryogenesis
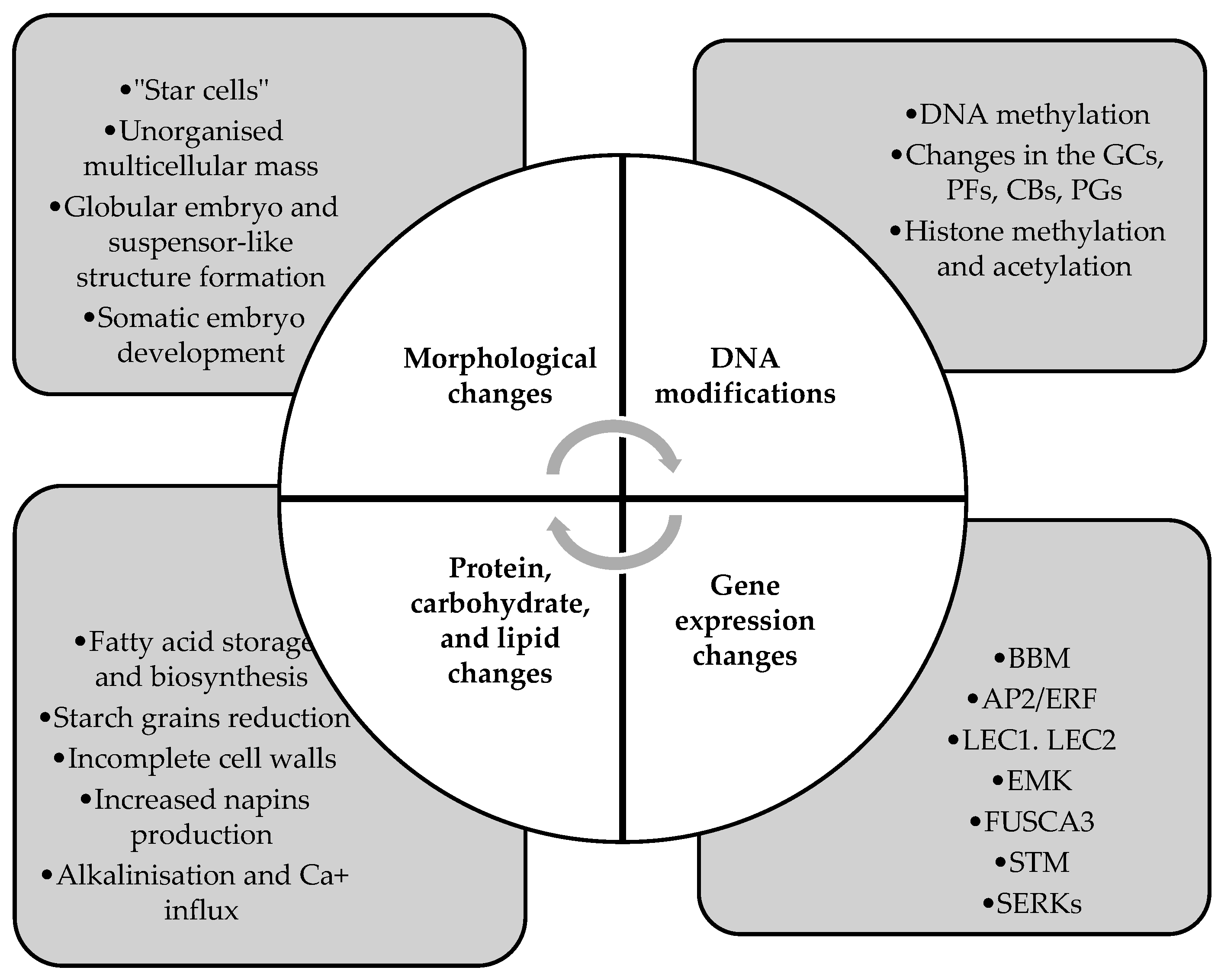
4.1. Mechanisms of Microspore Reprogramming to Embryogenesis in Brassica napus
4.1.1. Morphological Changes
4.1.2. Protein, Carbohydrate, and Lipid Changes during Microspore Embryogenesis
4.1.3. DNA Modifications
4.1.4. Gene Expression Changes
5. Chromosome Doubling
6. Brassica napus Haploid Production via Interploidy Hybridization
7. Brassica napus Haploid Induction Lines
8. Recent Findings Contributing to Brassica napus Double Haploid Plants
8.1. Plant Structure
8.2. Number of Inflorescence, Seeds, and Pod-Shattering Tendencies
8.3. Flowering Time
8.4. Seed Traits
8.5. Resistance to Diseases
8.5.1. Sclerotinia Stem Rot
8.5.2. Clubroot
8.5.3. Blackleg
9. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Iniguez-Luy, F.L.; Federico, M.L. The Genetics of Brassica napus. In Genetics and Genomics of the Brassicaceae. Plant Genetics and Genomics: Crops and Models; Schmidt, R., Bancroft, I., Eds.; Springer: New York, NY, USA, 2011; Volume 9, pp. 291–322. ISBN 978-1-4419-7118-0. [Google Scholar]
- Shahbandeh, M. Production of Major Vegetable Oils Worldwide from 2012/13 to 2021/2022, by Type. Available online: https://www.statista.com/statistics/263933/production-of-vegetable-oils-worldwide-since-2000/ (accessed on 28 March 2023).
- Rudko, T. Kierunki Hodowli Rzepaku. In Uprawa Rzepaku Ozimego; Wydawnictwo Instytutu Agrofizyki im. Bohdana Dobrzańskiego PAN: Lublin, Poland, 2011; ISBN 9788389969910. [Google Scholar]
- Hallauer, A.R. Evolution of Plant Breeding. Crop. Breed. Appl. Biotechnol. 2011, 11, 197–206. [Google Scholar] [CrossRef]
- Szała, L.; Sosnowska, K.; Cegielska-Taras, T. Induced Chromosome Doubling in Microspores and Regenerated Haploid Plants of Brassica napus. Acta Biol. Crac. Ser. Bot. 2020, 62, 23–31. [Google Scholar] [CrossRef]
- Rahman, M.; Michalak, M.; Michalak De Jiménez, M. Behind the Scenes of Microspore-Based Double Haploid Development in Brassica napus: A Review. J. Plant Sci. Mol. Breed. 2016, 5, 51. [Google Scholar] [CrossRef]
- Shen, K.; Qu, M.; Zhao, P.; Shen, K.; Qu, M.; Zhao, P. The Roads to Haploid Embryogenesis. Plants 2023, 12, 243. [Google Scholar] [CrossRef] [PubMed]
- Alan, A.R.; Celebi-Toprak, F.; Lachin, A.; Yildiz, D.; Gozen, V.; Besirli, G. Doubled Haploid Broccoli (Brassica olearacea var. italica) Plants from Anther Culture. Methods Mol. Biol. 2021, 2288, 201–216. [Google Scholar] [CrossRef]
- Shariatpanahi, M.E.; Niazian, M.; Ahmadi, B. Methods for Chromosome Doubling. Methods Mol. Biol. 2021, 2287, 127–148. [Google Scholar] [CrossRef]
- Su, Y.H.; Tang, L.P.; Zhao, X.Y.; Zhang, X.S. Plant Cell Totipotency: Insights into Cellular Reprogramming. J. Integr. Plant Biol. 2021, 63, 228–243. [Google Scholar] [CrossRef]
- Jacquier, N.M.A.; Gilles, L.M.; Martinant, J.P.; Rogowsky, P.M.; Widiez, T. Maize In planta Haploid Inducer Lines: A Cornerstone for Doubled Haploid Technology. Methods Mol. Biol. 2021, 2288, 25–48. [Google Scholar] [CrossRef]
- Seguí-Simarro, J.M.; Nuez, F. Pathways to Doubled Haploidy: Chromosome Doubling during Androgenesis. Cytogenet. Genome Res. 2008, 120, 358–369. [Google Scholar] [CrossRef]
- Niazian, M.; Shariatpanahi, M.E. In vitro-Based Doubled Haploid Production: Recent Improvements. Euphytica 2020, 216, 69. [Google Scholar] [CrossRef]
- Devaux, P. The Hordeum bulbosum (L.) Method. In Doubled Haploid Production in Crop Plants; Springer: Dordrecht, The Netherlands, 2003; pp. 15–19. [Google Scholar] [CrossRef]
- Chmielowiec, M.; Kuta, E.; Przywara, L.; Rog, L. An Attempt to Induce Gynogenesis in Brassica napus L. Acta Biol. Cracoviensia. Ser. Bot. Et. Zool. Suppl. 1997, 39, 1–49. [Google Scholar]
- Rojek, J. Autonomous Endosperm Development in Unpollinated Pistils of Brassica napus L. cv. Topas Cultured in vitro Inducing Autonomous Endosperm Development in Sexual Brassicaceae. Acta Biol. Crac. Ser. Bot. 2002, 44, 195–202. [Google Scholar]
- Siebel, J.; Pauls, K.P. A Comparison of Anther and Microspore Culture as a Breeding Tool in Brassica napus. Theor. Appl. Genet. 1989, 78, 473–479. [Google Scholar] [CrossRef] [PubMed]
- Mathias, R.; Röbbelen, G. Effective Diploidization of Microspore-Derived Haploids of Rape (Brassica napus L.) by In vitro Colchicine Treatment. Plant Breed. 1991, 106, 82–84. [Google Scholar] [CrossRef]
- Mohammadi, P.P.; Moieni, A.; Ebrahimi, A.; Javidfar, F. Doubled Haploid Plants Following Colchicine Treatment of Microspore-Derived Embryos of Oilseed Rape (Brassica napus L.). Plant Cell Tissue Organ. Cult. 2012, 108, 251–256. [Google Scholar] [CrossRef]
- Zhong, Y.; Liu, C.; Qi, X.; Jiao, Y.; Wang, D.; Wang, Y.; Liu, Z.; Chen, C.; Chen, B.; Tian, X.; et al. Mutation of ZmDMP Enhances Haploid Induction in Maize. Nat. Plants 2019, 5, 575–580. [Google Scholar] [CrossRef]
- Zhong, Y.; Chen, B.; Wang, D.; Zhu, X.; Wang, Y.; Li, Y.; Liu, J.; Zhang, J.; Chen, M.; Wang, M.; et al. A Genotype Independent DMP-HI System in Dicot Crops. bioRxiv 2021. [Google Scholar] [CrossRef]
- Fu, S.; Yin, L.; Xu, M.; Li, Y.; Wang, M.; Yang, J.; Fu, T.; Wang, J.; Shen, J.; Ali, A.; et al. Maternal Doubled Haploid Production in Interploidy Hybridization between Brassica napus and Brassica Allooctaploids. Planta 2018, 247, 113–125. [Google Scholar] [CrossRef]
- Chen, H.F.; Wang, H.; Li, Z.Y. Production and Genetic Analysis of Partial Hybrids in Intertribal Crosses between Brassica Species (B. Rapa, B. Napus) and Capsella Bursa-Pastoris. Plant Cell Rep. 2007, 26, 1791–1800. [Google Scholar] [CrossRef]
- Thomas, E.; Wenzel, G. Embryogenesis from Microspores of Brassica napus. Z. Fuer Pflanz. 1975, 74, 77–81. [Google Scholar]
- Lichter, R. Induction of Haploid Plants from Isolated Pollen of Brassica napus. Z. Für Pflanzenphysiol. 1982, 105, 427–434. [Google Scholar] [CrossRef]
- Touraev, A.; Pfosser, M.; Heberle-Bors, E. The Microspore: A Haploid Multipurpose Cell. Adv. Bot. Res. 2001, 35, 53–109. [Google Scholar] [CrossRef]
- Berenguer, E.; Minina, E.A.; Carneros, E.; Barany, I.; Bozhkov, P.V.; Testillano, P.S. Suppression of Metascapase- and Autophagy-Dependent Cell Death Improves Stress-Induced Microspore Embryogenesis in Brassica napus. Plant Cell Physiol. 2020, 61, 2097–2110. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Sanz, H.; Moreno-Romero, J.; Solís, M.T.; Köhler, C.; Risueño, M.C.; Testillano, P.S. Changes in Histone Methylation and Acetylation during Microspore Reprogramming to Embryogenesis Occur Concomitantly with BnHKMT and BnHAT Expression and Are Associated with Cell Totipotency, Proliferation, and Differentiation in Brassica napus. Cytogenet. Genome Res. 2014, 143, 209–218. [Google Scholar] [CrossRef]
- Corral-Martínez, P.; Siemons, C.; Horstman, A.; Angenent, G.C.; de Ruijter, N.; Boutilier, K. Live Imaging of Embryogenic Structures in Brassica napus Microspore Embryo Cultures Highlights the Developmental Plasticity of Induced Totipotent Cells. Plant Reprod. 2020, 33, 143–158. [Google Scholar] [CrossRef] [PubMed]
- Weigt, D.; Niemann, J.; Siatkowski, I.; Zyprych-Walczak, J.; Olejnik, P.; Kurasiak-Popowska, D. Effect of Zearalenone and Hormone Regulators on Microspore Embryogenesis in Anther Culture of Wheat. Plants 2019, 8, 487. [Google Scholar] [CrossRef]
- Gu, H.H.; Hagberg, P.; Zhou, W.J. Cold Pretreatment Enhances Microspore Embryogenesis in Oilseed Rape (Brassica napus L.). Plant Growth Regul. 2004, 42, 137–143. [Google Scholar] [CrossRef]
- Prem, D.; Solís, M.T.; Bárány, I.; Rodríguez-Sanz, H.; Risueño, M.C.; Testillano, P.S. A New Microspore Embryogenesis System under Low Temperature Which Mimics Zygotic Embryogenesis Initials, Expresses Auxin and Efficiently Regenerates Doubled-Haploid Plants in Brassica napus. BMC Plant Biol. 2012, 12, 127. [Google Scholar] [CrossRef]
- Ahmadi, B.; Shariatpanahi, M.E.; Teixeira da Silva, J.A. Efficient Induction of Microspore Embryogenesis Using Abscisic Acid, Jasmonic Acid and Salicylic Acid in Brassica napus L. Plant Cell Tissue Organ. Cult. 2014, 116, 343–351. [Google Scholar] [CrossRef]
- Corral-Martínez, P.; Camacho-Fernández, C.; Mir, R.; Seguí-Simarro, J.M. Doubled Haploid Production in High- and Low-Response Genotypes of Rapeseed (Brassica napus) Through Isolated Microspore Culture. Methods Mol. Biol. 2021, 2288, 129–144. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures. Physiol. Plant 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Gamborg, O.L.; Murashige, T.; Thorpe, T.A.; Vasil, I.K. Plant Tissue Culture Media. Soc. Vitr. Biol. 1976, 12, 473–478. [Google Scholar] [CrossRef] [PubMed]
- Shmykova, N.A.; Shumilina, D.V.; Suprunova, T.P. Doubled Haploid Production in Brassica L. Vavilov J. Genet. Breed. 2015, 19, 111. [Google Scholar] [CrossRef]
- Hosp, J.; Maraschin, S.D.F.; Touraev, A.; Boutilier, K. Functional Genomics of Microspore Embryogenesis. Euphytica 2007, 158, 275–285. [Google Scholar] [CrossRef]
- Pauls, K.P.; Chan, J.; Woronuk, G.; Schulze, D.; Brazolot, J. When Microspores Decide to Become Embryos-Cellular and Molecular Changes. Can. J. Bot. 2006, 84, 668–678. [Google Scholar] [CrossRef]
- Soriano, M.; Li, H.; Jacquard, C.; Angenent, G.C.; Krochko, J.; Offringa, R.; Boutilier, K. Plasticity in Cell Division Patterns and Auxin Transport Dependency during in Vitro Embryogenesis in Brassica napus C W. Plant Cell 2014, 26, 2568–2581. [Google Scholar] [CrossRef]
- Hause, B.; van Veenendaal, W.L.H.; Hause, G.; van Lammeren, A.A.M. Expression of Polarity during Early Development of Microspore-Derived and Zygotic Embryos of Brassica napus L. Cv. Topas. Bot. Acta 1994, 107, 407–415. [Google Scholar] [CrossRef]
- Custers, J.B.M.; Cordewener, J.H.G.; Nöllen, Y.; Dons, H.J.M.; van Lockeren Campagne, M.M. Temperature Controls Both Gametophytic and Sporophytic Development in Microspore Cultures of Brassica napus. Plant Cell. Rep. 1994, 13, 267–271. [Google Scholar] [CrossRef]
- Joosen, R.; Cordewener, J.; Supena, E.D.J.; Vorst, O.; Lammers, M.; Maliepaard, C.; Zeilmaker, T.; Miki, B.; America, T.; Custers, J.; et al. Combined Transcriptome and Proteome Analysis Identifies Pathways and Markers Associated with the Establishment of Rapeseed Microspore-Derived Embryo Development. Plant Physiol. 2007, 144, 155–172. [Google Scholar] [CrossRef]
- Telmer, C.A.; Newcomb, W.; Simmonds, D.H. Microspore Development in Brassica napus and the Effect of High Temperature on Division in vivo and in vitro. Protoplasma 1993, 172, 154–165. [Google Scholar] [CrossRef]
- Ilić-Grubor, K.; Attree, S.M.; Fowke, L.C. Comparative Morphological Study of Zygotic and Microspore-Derived Embryos of Brassica napus L. as Revealed by Scanning Electron Microscopy. Ann. Bot. 1998, 82, 157–165. [Google Scholar] [CrossRef]
- Kott, L.S.; Beversdorf, W.D. Enhanced Plant Regeneration from Microspore-Derived Embryos of Brassica napus by Chilling, Partial Desiccation and Age Selection. Plant Cell Tissue Organ. Cult. 1990, 23, 187–192. [Google Scholar] [CrossRef]
- Senaratna, T.; Kott, L.; Beversdorf, W.D.; McKersie, B.D. Desiccation of Microspore Derived Embryos of Oilseed Rape (Brassica napus L.). Plant Cell Rep. 1991, 10, 342–344. [Google Scholar] [CrossRef]
- Cegielska-Taras, T.; Tykarska, T.; Szała, L.; Kuraś, L.; Krzymański, J. Direct Plant Development from Microspore-Derived Embryos of Winter Oilseed Rape Brassica napus L. ssp. oleifera (DC.) Metzger. Euphytica 2002, 124, 341–347. [Google Scholar] [CrossRef]
- Zhang, G.Q.; Zhang, D.Q.; Tang, G.X.; He, Y.; Zhou, W.J. Plant Development from Microspore-Derived Embryos in Oilseed Rape as Affected by Chilling, Desiccation and Cotyledon Excision. Biol. Plant 2006, 50, 180–186. [Google Scholar] [CrossRef]
- Taylor, D.C.; Weber, N.; Underhill, E.W.; Pomeroy, M.K.; Keller, W.A.; Scowcroft, W.R.; Wilen, R.W.; Moloney, M.M.; Holbrook, L.A. Storage-Protein Regulation and Lipid Accumulation in Microspore Embryos of Brassica napus L. Planta 1990, 181, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Pomeroy, M.K.; Kramer, J.K.G.; Hunt, D.J.; Keller, W.A. Fatty Acid Changes during Development of Zygotic and Microspore-Derived Embryos of Brassica napus. Physiol. Plant 1991, 81, 447–454. [Google Scholar] [CrossRef]
- Wiberg, E.; Råhlen, L.; Hellman, M.; Tillberg, E.; Glimelius, K.; Stymne, S. The Microspore-Derived Embryo of Brassica napus L. as a Tool for Studying Embryo-Specific Lipid Biogenesis and Regulation of Oil Quality. Theor. Appl. Genet. 1991, 82, 515–520. [Google Scholar] [CrossRef]
- Piffanelli, P.; Ross, J.H.E.; Murphy, D.J. Intra- and Extracellular Lipid Composition and Associated Gene Expression Patterns during Pollen Development in Brassica napus. Plant J. 1997, 11, 549–562. [Google Scholar] [CrossRef]
- Satpute, G.K.; Long, H.; Seguui-Simarro, J.M.; Risueno, M.C.; Testillano, P.S. Cell Architecture during Gametophytic and Embryogenic Microspore Development in Brassica napus L. Acta Physiol. Plant 2005, 27, 665–674. [Google Scholar] [CrossRef]
- Telmer, C.A.; Newcomb, W.; Simmonds, D.H. Cellular Changes during Heat Shock Induction and Embryo Development of Cultured Microspores of Brassica napus Cv. Topas. Protoplasma 1995, 185, 106–112. [Google Scholar] [CrossRef]
- Parra-Vega, V.; Corral-Martínez, P.; Rivas-Sendra, A.; Seguí-Simarro, J.M. Induction of Embryogenesis in Brassica napus Microspores Produces a Callosic Subintinal Layer and Abnormal Cell Walls with Altered Levels of Callose and Cellulose. Front. Plant Sci. 2015, 6, 1018. [Google Scholar] [CrossRef] [PubMed]
- Corral-Martínez, P.; Driouich, A.; Seguí-Simarro, J.M. Dynamic Changes in Arabinogalactan-Protein, Pectin, Xyloglucan and Xylan Composition of the Cell Wall during Microspore Embryogenesis in Brassica napus. Front. Plant Sci. 2019, 10, 332. [Google Scholar] [CrossRef]
- Parra-Vega, V.; Corral-Martínez, P.; Rivas-Sendra, A.; Seguí-Simarro, J.M. Formation and Excretion of Autophagic Plastids (Plastolysomes) in Brassica napus Embryogenic Microspores. Front. Plant Sci. 2015, 6, 94. [Google Scholar] [CrossRef] [PubMed]
- Perera, S.P.; McIntosh, T.C.; Wanasundara, J.P.D. Structural Properties of Cruciferin and Napin of Brassica napus (Canola) Show Distinct Responses to Changes in pH and Temperature. Plants 2016, 5, 36. [Google Scholar] [CrossRef]
- Boutilier, K.A.; Ginés, M.J.; DeMoor, J.M.; Huang, B.; Baszczynski, C.L.; Iyer, V.N.; Miki, B.L. Expression of the BnmNAP Subfamily of Napin Genes Coincides with the Induction of Brassica Microspore Embryogenesis. Plant Mol. Biol. 1994, 26, 1711–1723. [Google Scholar] [CrossRef]
- Hays, D.B.; Wilen, R.W.; Sheng, C.; Moloney, M.M.; Pharis, R.P. Embryo-Specific Gene Expression in Microspore-Derived Embryos Of Brassica napus. An Interaction between Abscisic Acid and Jasmonic Acid1,2. Plant Physiol. 1999, 119, 1065–1072. [Google Scholar] [CrossRef]
- Solís, M.-T.; Berenguer, E.; Risueño, M.C.; Testillano, P.S. BnPME Is Progressively Induced after Microspore Reprogramming to Embryogenesis, Correlating with Pectin de-Esterification and Cell Differentiation in Brassica Napus. BMC Plant Biol. 2016, 16, 176. [Google Scholar] [CrossRef]
- Rodríguez-Sanz, H.; Solís, M.T.; Lopez, M.F.; Gómez-Cadenas, A.; Risueño, M.C.; Testillano, P.S. Auxin Biosynthesis, Accumulation, Action and Transport Are Involved in Stress-Induced Microspore Embryogenesis Initiation and Progression in Brassica napus. Plant Cell. Physiol. 2015, 56, 1401–1417. [Google Scholar] [CrossRef]
- Solís, M.T.; Rodríguez-Serrano, M.; Meijón, M.; Cañal, M.J.; Cifuentes, A.; Risueño, M.C.; Testillano, P.S. DNA Methylation Dynamics and MET1a-like Gene Expression Changes during Stress-Induced Pollen Reprogramming to Embryogenesis. J. Exp. Bot. 2012, 63, 6431–6444. [Google Scholar] [CrossRef]
- Li, J.; Huang, Q.; Sun, M.; Zhang, T.; Li, H.; Chen, B.; Xu, K.; Gao, G.; Li, F.; Yan, G.; et al. Global DNA Methylation Variations after Short-Term Heat Shock Treatment in Cultured Microspores of Brassica napus cv. Topas. Sci. Rep. 2016, 6, 38401. [Google Scholar] [CrossRef] [PubMed]
- Seguí-Simarro, J.M.; Corral-Martínez, P.; Corredor, E.; Raska, I.; Testillano, P.S.; Risueño, M.C. A Change of Developmental Program Induces the Remodeling of the Interchromatin Domain during Microspore Embryogenesis in Brassica napus L. J. Plant Physiol. 2011, 168, 746–757. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Soriano, M.; Cordewener, J.; Muiño, J.M.; Riksen, T.; Fukuok, H.; Angenent, G.C.; Boutilier, K. The Histone Deacetylase Inhibitor Trichostatin a Promotes Totipotency in the Male Gametophyte. Plant Cell 2014, 26, 195–209. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Wrobel-Marek, J.; Heidmann, I.; Horstman, A.; Chen, B.; Reis, R.; Angenent, G.C.; Boutilier, K. Auxin Biosynthesis Maintains Embryo Identity and Growth during BABY BOOM-Induced Somatic Embryogenesis Performed Experiments and Analyzed the Data. Plant Physiol. 2022, 188, 1095–1110. [Google Scholar] [CrossRef]
- Jha, P.; Kumar, V. BABY BOOM (BBM): A Candidate Transcription Factor Gene in Plant Biotechnology. Biotechnol. Lett. 2018, 40, 1467–1475. [Google Scholar] [CrossRef] [PubMed]
- Boutilier, K.; Offringa, R.; Sharma, V.K.; Kieft, H.; Ouellet, T.; Zhang, L.; Hattori, J.; Liu, C.M.; van Lammeren, A.A.M.; Miki, B.L.A.; et al. Ectopic Expression of BABY BOOM Triggers a Conversion from Vegetative to Embryonic Growth. Plant Cell 2002, 14, 1737–1749. [Google Scholar] [CrossRef]
- Passarinho, P.; Ketelaar, T.; Xing, M.; van Arkel, J.; Maliepaard, C.; Hendriks, M.W.; Joosen, R.; Lammers, M.; Herdies, L.; den Boer, B.; et al. BABY BOOM Target Genes Provide Diverse Entry Points into Cell Proliferation and Cell Growth Pathways. Plant Mol. Biol. 2008, 68, 225–237. [Google Scholar] [CrossRef]
- Tsuwamoto, R.; Yokoi, S.; Takahata, Y. Arabidopsis EMBRYOMAKER Encoding an AP2 Domain Transcription Factor Plays a Key Role in Developmental Change from Vegetative to Embryonic Phase. Plant Mol. Biol. 2010, 73, 481–492. [Google Scholar] [CrossRef]
- Belide, S.; Zhou, X.R.; Kennedy, Y.; Lester, G.; Shrestha, P.; Petrie, J.R.; Singh, S.P. Rapid Expression and Validation of Seed-Specific Constructs in Transgenic LEC2 Induced Somatic Embryos of Brassica napus. Plant Cell Tissue Organ. Cult. 2013, 113, 543–553. [Google Scholar] [CrossRef]
- Elahi, N.; Duncan, R.W.; Stasolla, C. Effects of Altered Expression of LEAFY COTYLEDON1 and FUSCA3 on Microspore-Derived Embryogenesis of Brassica napus L. J. Genet. Eng. Biotechnol. 2016, 14, 19–30. [Google Scholar] [CrossRef]
- Takahata, Y.; Takahashi, Y.; Tsuwamoto, R. Microspore Culture and Doubled Haploid Technology. In Biotechnology of Crucifers; Gupta, S.K., Ed.; Springer: New York, NY, USA, 2013; Volume 1, pp. 45–62. ISBN 78-1-4614-7795-2. [Google Scholar]
- Malik, M.R.; Wang, F.; Dirpaul, J.M.; Zhou, N.; Polowick, P.L.; Ferrie, A.M.R.; Krochko, J.E. Transcript Profiling and Identification of Molecular Markers for Early Microspore Embryogenesis in Brassica napus. Plant Physiol. 2020, 144, 134–154. [Google Scholar] [CrossRef] [PubMed]
- Tsuwamoto, R.; Fukuoka, H.; Takahata, Y. Identification and Characterization of Genes Expressed in Early Embryogenesis from Microspores of Brassica napus. Planta 2007, 225, 641–652. [Google Scholar] [CrossRef] [PubMed]
- Tsuwamoto, R.; Takahata, Y. Identification of Genes Specifically Expressed in Androgenesis-Derived Embryo in Rapeseed (Brassica napus L.). Breed. Sci. 2008, 58, 251–259. [Google Scholar] [CrossRef]
- Ahmadi, B.; Masoomi-Aladizgeh, F.; Shariatpanahi, M.E.; Azadi, P.; Keshavarz-Alizadeh, M. Molecular Characterization and Expression Analysis of SERK1 and SERK2 in Brassica napus L.: Implication for Microspore Embryogenesis and Plant Regeneration. Plant Cell Rep. 2016, 35, 185–193. [Google Scholar] [CrossRef]
- Möllers, C.; Iqbal, M.C.M.; Röbbelen, G. Efficient Production of Doubled Haploid Brassica napus Plants by Colchicine Treatment of Microspores. Euphytica 1994, 75, 95–104. [Google Scholar] [CrossRef]
- Zhou, W.J.; Tang, G.X.; Hagberg, P. Efficient Production of Doubled Haploid Plants by Immediate Colchicine Treatment of Isolated Microspores in Winter Brassica napus. Plant Growth Regul. 2002, 37, 185–192. [Google Scholar] [CrossRef]
- Chen, Z.Z.; Snyder, S.; Fan, Z.G.; Loh, W.H. Efficient Production of Doubled Haploid Plants through Chromosome Doubling of Isolated Microspores in Brassica napus. Plant Breed. 1994, 113, 217–221. [Google Scholar] [CrossRef]
- Zhao, J.; Simmonds, D.H. Application of Trifluralin to Embryogenic Microspore Cultures to Generate Doubled Haploid Plants in Brassica napus. Physiol. Plant 1995, 95, 304–309. [Google Scholar] [CrossRef]
- Klíma, M.; Vyvadilová, M.; Kučera, V. Chromosome Doubling Effects of Selected Antimitotic Agents in Brassica napus Microspore Culture. Czech J. Genet. Plant Breed. 2008, 44, 30–36. [Google Scholar] [CrossRef]
- Zhao, X.; Xu, X.; Xie, H.; Chen, S.; Jin, W. Fertilization and Uniparental Chromosome Elimination during Crosses with Maize Haploid Inducers. Plant Physiol. 2013, 163, 721–731. [Google Scholar] [CrossRef]
- Zhang, W.; Ma, Y.; Zhu, Z.; Huang, L.; Ali, A.; Luo, X.; Zhou, Y.; Li, Y.; Xu, P.; Yang, J.; et al. Maternal Karyogene and Cytoplasmic Genotype Affect the Induction Efficiency of Doubled Haploid Inducer in Brassica napus. BMC Plant Biol. 2021, 21, 207. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Yang, J.; Zhu, Z.; Huang, L.; Ali, A.; Javed, H.H.; Zhang, W.; Zhou, Y.; Yin, L.; Xu, P.; et al. Genetic Characteristics and Ploidy Trigger the High Inducibility of Double Haploid (DH) Inducer in Brassica napus. BMC Plant Biol. 2021, 21, 538. [Google Scholar] [CrossRef]
- Zhang, W.; Shi, H.; Zhou, Y.; Liang, X.; Luo, X.; Xiao, C.; Li, Y.; Xu, P.; Wang, J.; Gong, W.; et al. Rapid and Synchronous Breeding of Cytoplasmic Male Sterile and Maintainer Line Through Mitochondrial DNA Rearrangement Using Doubled Haploid Inducer in Brassica napus. Front. Plant Sci. 2022, 13, 871006. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Yang, M.; Zhao, S.; Shi, H.; Li, Y.; Gong, W.; Yang, J.; Wang, J.; Zou, Q.; Tao, L.; et al. Rapid Creation of Interspecific Hybrid Progeny to Broaden Genetic Distance through Double Haploid (DH) Inducer in Brassica napus. Plants 2022, 11, 695. [Google Scholar] [CrossRef]
- Zhong, Y.; Chen, B.; Li, M.; Wang, D.; Jiao, Y.; Qi, X.; Wang, M.; Liu, Z.; Chen, C.; Wang, Y.; et al. A DMP-Triggered in vivo Maternal Haploid Induction System in the Dicotyledonous Arabidopsis. Nat. Plants 2020, 6, 466–472. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Xia, X.; Jiang, T.; Li, L.; Zhang, P.; Niu, L.; Cheng, H.; Wang, K.; Lin, H. In planta Haploid Induction by Genome Editing of DMP in the Model Legume Medicago truncatula. Plant Biotechnol. J. 2022, 20, 22–24. [Google Scholar] [CrossRef]
- Zhong, Y.; Chen, B.; Wang, D.; Zhu, X.; Li, M.; Zhang, J.; Chen, M.; Wang, M.; Riksen, T.; Liu, J.; et al. In vivo Maternal Haploid Induction in Tomato. Plant Biotechnol. J. 2021, 20, 25–252. [Google Scholar] [CrossRef]
- Zhong, Y.; Wang, Y.; Chen, B.; Liu, J.; Wang, D.; Li, M.; Qi, X.; Liu, C.; Boutilier, K.; Chen, S. Establishment of a dmp Based Maternal Haploid Induction System for Polyploid Brassica napus and Nicotiana tabacum. J. Integr. Plant Biol. 2022, 64, 1281–1294. [Google Scholar] [CrossRef]
- Li, Y.; Li, D.; Xiao, Q.; Wang, H.; Wen, J.; Tu, J.; Shen, J.; Fu, T.; Yi, B. An in planta Haploid Induction System in Brassica napus. J. Integr. Plant Biol. 2022, 64, 1140–1144. [Google Scholar] [CrossRef]
- Wang, X.; Wang, H.; Long, Y.; Liu, L.; Zhao, Y.; Tian, J.; Zhao, W.; Li, B.; Chen, L.; Chao, H.; et al. Dynamic and Comparative QTL Analysis for Plant Height in Different Developmental Stages of Brassica napus L. Theor. Appl. Genet. 2015, 128, 1175–1192. [Google Scholar] [CrossRef]
- Cai, G.; Yang, Q.; Yi, B.; Fan, C.; Zhang, C.; Edwards, D.; Batley, J.; Zhou, Y. A Bi-Filtering Method for Processing Single Nucleotide Polymorphism Array Data Improves the Quality of Genetic Map and Accuracy of Quantitative Trait Locus Mapping in Doubled Haploid Populations of Polyploid Brassica napus. BMC Genom. 2015, 16, 409. [Google Scholar] [CrossRef]
- Yan, S.; He, J.; Tang, M.; Ming, B.; Li, H.; Fan, S.; Xiong, Y.; Chao, H.; Zhang, L.; Wang, A.; et al. Dissecting the Meiotic Recombination Patterns in a Brassica napus Double Haploid Population Using 60K SNP Array. Int. J. Mol. Sci. 2023, 24, 4469. [Google Scholar] [CrossRef] [PubMed]
- Raman, R.; Diffey, S.; Carling, J.; Cowley, R.B.; Kilian, A.; Luckett, D.J.; Raman, H.; Raman, R.; Diffey, S.; Carling, J.; et al. Quantitative Genetic Analysis of Grain Yield in an Australian Brassica napus Doubled-Haploid Population. Crop. Pasture Sci. 2016, 67, 298–307. [Google Scholar] [CrossRef]
- Li, C.; Sang, S.; Sun, M.D.; Yang, J.; Shi, Y.Q.; Hu, X.Z.; Li, Y.; Hao, M.Y.; Chu, W.; Zhang, H.Y.; et al. Direct Modification of Multiple Gene Homoeologs in Brassica oleracea and Brassica napus Using Doubled Haploid Inducer-mediated Genome-editing System. Plant Biotechnol. J. 2021, 19, 1889. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Paillard, S.; Fopa-Fomeju, B.; Falentin, C.; Deniot, G.; Baron, C.; Vallée, P.; Manzanares-Dauleux, M.J.; Delourme, R. Multi-Year Linkage and Association Mapping Confirm the High Number of Genomic Regions Involved in Oilseed Rape Quantitative Resistance to Blackleg. Theor. Appl. Genet. 2018, 131, 1627–1643. [Google Scholar] [CrossRef] [PubMed]
- Raman, H.; Raman, R.; Diffey, S.; Qiu, Y.; McVittie, B.; Barbulescu, D.M.; Salisbury, P.A.; Marcroft, S.; Delourme, R. Stable Quantitative Resistance Loci to Blackleg Disease in Canola (Brassica napus L.) over Continents. Front. Plant Sci. 2018, 871, 1622. [Google Scholar] [CrossRef]
- Huang, Y.J.; Paillard, S.; Kumar, V.; King, G.J.; Fitt, B.D.L.; Delourme, R. Oilseed Rape (Brassica napus) Resistance to Growth of Leptosphaeria maculans in Leaves of Young Plants Contributes to Quantitative Resistance in Stems of Adult Plants. PLoS ONE 2019, 14, e0222540. [Google Scholar] [CrossRef]
- Raman, R.; Taylor, B.; Marcroft, S.; Stiller, J.; Eckermann, P.; Coombes, N.; Rehman, A.; Lindbeck, K.; Luckett, D.; Wratten, N.; et al. Molecular Mapping of Qualitative and Quantitative Loci for Resistance to Leptosphaeria maculans Causing Blackleg Disease in Canola (Brassica napus L.). Theor. Appl. Genet. 2012, 125, 405–418. [Google Scholar] [CrossRef]
- Larkan, N.J.; Raman, H.; Lydiate, D.J.; Robinson, S.J.; Yu, F.; Barbulescu, D.M.; Raman, R.; Luckett, D.J.; Burton, W.; Wratten, N.; et al. Multi-Environment QTL Studies Suggest a Role for Cysteine-Rich Protein Kinase Genes in Quantitative Resistance to Blackleg Disease in Brassica napus. BMC Plant Biol. 2016, 16, 1–16. [Google Scholar] [CrossRef]
- Raman, R.; Diffey, S.; Barbulescu, D.M.; Coombes, N.; Luckett, D.; Salisbury, P.; Cowley, R.; Marcroft, S.; Raman, H. Genetic and Physical Mapping of Loci for Resistance to Blackleg Disease in Canola (Brassica napus L.). Sci. Rep. 2020, 10, 183. [Google Scholar] [CrossRef]
- Yu, F.; Zhang, Y.; Wang, J.; Chen, Q.; Karim, M.M.; Gossen, B.D.; Peng, G. Identification of Two Major QTLs in Brassica napus Lines with Introgressed Clubroot Resistance from Turnip Cultivar ECD01. Front. Plant Sci. 2022, 12, 3092. [Google Scholar] [CrossRef]
- Wang, Z.; Megha, S.; Kebede, B.; Kav, N.N.V.; Rahman, H. Genetic and Molecular Analysis Reveals That Two Major Loci and Their Interaction Confer Clubroot Resistance in Canola Introgressed from Rutabaga. Plant Genome 2022, 15, e20241. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.; Zhang, X.; Peng, G.; Falk, K.C.; Strelkov, S.E.; Gossen, B.D. Genotyping-by-Sequencing Reveals Three QTL for Clubroot Resistance to Six Pathotypes of Plasmodiophora brassicae in Brassica rapa. Sci. Rep. 2017, 7, 4516. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.R.; Oh, S.H.; Chhapekar, S.S.; Dhandapani, V.; Lee, C.Y.; Rameneni, J.J.; Ma, Y.; Choi, G.J.; Lee, S.S.; Lim, Y.P. Quantitative Trait Locus Mapping of Clubroot Resistance and Plasmodiophora brassicae Pathotype Banglim-Specific Marker Development in Brassica rapa. Int. J. Mol. Sci. 2020, 21, 4157. [Google Scholar] [CrossRef]
- Fredua-Agyeman, R.; Rahman, H. Mapping of the Clubroot Disease Resistance in Spring Brassica napus Canola Introgressed from European Winter Canola cv. ‘Mendel.’ Euphytica 2016, 211, 201–213. [Google Scholar] [CrossRef]
- Behla, R.; Hirani, A.H.; Zelmer, C.D.; Yu, F.; Fernando, W.G.D.; McVetty, P.; Li, G. Identification of Common QTL for Resistance to Sclerotinia sclerotiorum in Three Doubled Haploid Populations of Brassica napus (L.). Euphytica 2017, 213, 260. [Google Scholar] [CrossRef]
- Wu, J.; Chen, P.; Zhao, Q.; Cai, G.; Hu, Y.; Xiang, Y.; Yang, Q.; Wang, Y.; Zhou, Y. Co-Location of QTL for Sclerotinia Stem Rot Resistance and Flowering Time in Brassica napus. Crop. J. 2019, 7, 227–237. [Google Scholar] [CrossRef]
- Wu, J.; Cai, G.; Tu, J.; Li, L.; Liu, S.; Luo, X.; Zhou, L.; Fan, C.; Zhou, Y. Identification of QTLs for Resistance to Sclerotinia Stem Rot and BnaC.IGMT5.a as a Candidate Gene of the Major Resistant QTL SRC6 in Brassica napus. PLoS ONE 2013, 8, e67740. [Google Scholar] [CrossRef]
- Song, J.; Li, B.; Cui, Y.; Zhuo, C.; Gu, Y.; Hu, K.; Wen, J.; Yi, B.; Shen, J.; Ma, C.; et al. QTL Mapping and Diurnal Transcriptome Analysis Identify Candidate Genes Regulating Brassica napus Flowering Time. Int. J. Mol. Sci. 2021, 22, 7559. [Google Scholar] [CrossRef]
- Fletcher, R.S.; Mullen, J.L.; Heiliger, A.; McKay, J.K. QTL Analysis of Root Morphology, Flowering Time, and Yield Reveals Trade-Offs in Response to Drought in Brassica napus. J. Exp. Bot. 2015, 66, 245–256. [Google Scholar] [CrossRef]
- Javed, N.; Geng, J.; Tahir, M.; McVetty, P.B.E.; Li, G.; Duncan, R.W. Identification of QTL Influencing Seed Oil Content, Fatty Acid Profile and Days to Flowering in Brassica napus L. Euphytica 2016, 207, 191–211. [Google Scholar] [CrossRef]
- Li, B.; Zhao, W.; Li, D.; Chao, H.; Zhao, X.; Ta, N.; Li, Y.; Guan, Z.; Guo, L.; Zhang, L.; et al. Genetic Dissection of the Mechanism of Flowering Time Based on an Environmentally Stable and Specific QTL in Brassica napus. Plant Sci. 2018, 277, 296–310. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Chao, H.; Zhang, L.; Ta, N.; Zhao, Y.; Li, B.; Zhang, K.; Guan, Z.; Hou, D.; Chen, K.; et al. Integration of QTL Mapping and Gene Fishing Techniques to Dissect the Multi-Main Stem Trait in Rapeseed (Brassica napus L.). Front. Plant Sci. 2019, 10, 1152. [Google Scholar] [CrossRef]
- Chen, L.; Lei, W.; He, W.; Wang, Y.; Tian, J.; Gong, J.; Hao, B.; Cheng, X.; Shu, Y.; Fan, Z. Mapping of Two Major QTLs Controlling Flowering Time in Brassica napus Using a High-Density Genetic Map. Plants 2022, 11, 2635. [Google Scholar] [CrossRef] [PubMed]
- Xing, X.; Liu, H.; Ye, J.; Yao, Y.; Li, K.; Li, Y.; Du, D. QTL Analysis and Candidate Gene Prediction for Seed Density per Silique by QTL-Seq and RNA-Seq in Spring Brassica napus L. PLoS ONE 2023, 18, e0281875. [Google Scholar] [CrossRef] [PubMed]
- Jian, H.; Zhang, A.; Ma, J.; Wang, T.; Yang, B.; Shuang, L.S.; Liu, M.; Li, J.; Xu, X.; Paterson, A.H.; et al. Joint QTL Mapping and Transcriptome Sequencing Analysis Reveal Candidate Flowering Time Genes in Brassica napus L. BMC Genom. 2019, 20, 21. [Google Scholar] [CrossRef]
- Ganal, M.W.; Altmann, T.; Röder, M.S. SNP Identification in Crop Plants. Curr. Opin. Plant Biol. 2009, 12, 211–217. [Google Scholar] [CrossRef]
- Ding, C.; Jin, S. High-Throughput Methods for SNP Genotyping. In Single Nucleotide Polymorphism; Humana Press: Totowa, NJ, USA, 2009; Volume 578, pp. 245–254. [Google Scholar]
- Ganal, M.W.; Polley, A.; Graner, E.M.; Plieske, J.; Wieseke, R.; Luerssen, H.; Durstewitz, G. Large SNP Arrays for Genotyping in Crop Plants. J. Biosci. 2012, 37, 821–828. [Google Scholar] [CrossRef]
- Zhao, B.; Wang, B.; Li, Z.; Guo, T.; Zhao, J.; Guan, Z.; Liu, K. Identification and Characterization of a New Dwarf Locus DS-4 Encoding an Aux/IAA7 Protein in Brassica napus. Theor. Appl. Genet. 2019, 132, 1435–1449. [Google Scholar] [CrossRef]
- Wei, T.; Zhang, L.; Zhu, R.; Jiang, X.; Yue, C.; Su, Y.; Ren, H.; Wang, M. A Gain-of-Function Mutant of Iaa7 Inhibits Stem Elongation by Transcriptional Repression of Expa5 Genes in Brassica napus. Int. J. Mol. Sci. 2021, 22, 9018. [Google Scholar] [CrossRef]
- Chai, L.; Li, H.; Zhang, J.; Wu, L.; Zheng, B.; Cui, C.; Jiang, J.; Zuo, S.; Jiang, L. Rapid Identification of a Genomic Region Conferring Dwarfism in Rapeseed (Brassica napus L.) YA2016-12. Agronomy 2019, 9, 129. [Google Scholar] [CrossRef]
- Ye, S.; Yan, L.; Ma, X.; Chen, Y.; Wu, L.; Ma, T.; Zhao, L.; Yi, B.; Ma, C.; Tu, J.; et al. Combined BSA-Seq Based Mapping and RNA-Seq Profiling Reveal Candidate Genes Associated with Plant Architecture in Brassica napus. Int. J. Mol. Sci. 2022, 23, 2472. [Google Scholar] [CrossRef] [PubMed]
- Xin, S.; Dong, H.; Cui, Y.; Liu, Y.; Tian, G.; Deng, N.; Wan, H.; Liu, Z.; Li, X.; Qian, W. Identification of a Candidate QTG for Seed Number per Silique by Integrating QTL Mapping and RNA-Seq in Brassica napus L. Crop. J. 2023, 11, 189–197. [Google Scholar] [CrossRef]
- Samizadeh, H.; Yazdi-Samadi, B.Y.; Bihamta, M.R.; Taleii, A.; Stringam, G.R. Study of Pod Length Trait in Doubled Haploid Brassica napus Population by Molecular Markers. J. Agric. Sci. Technol. 2007, 9, 129–136. [Google Scholar]
- Chu, W.; Liu, J.; Cheng, H.; Li, C.; Fu, L.; Wang, W.; Wang, H.; Hao, M.; Mei, D.; Liu, K.; et al. A Lignified-Layer Bridge Controlled by a Single Recessive Gene Is Associated with High Pod-Shatter Resistance in Brassica napus L. Crop. J. 2022, 10, 638–646. [Google Scholar] [CrossRef]
- Fr´, F.; Fréd´, F.; Bouché, F.; Bouché, B.; Lobet, G.; Tocquin, P.; Erilleux, C.P. ´ FLOR-ID: An Interactive Database of Flowering-Time Gene Networks in Arabidopsis thaliana. Nucleic Acids Res. 2016, 44, D1167–D1171. [Google Scholar] [CrossRef]
- Fang, C.; Wang, Z.; Wang, P.; Song, Y.; Ahmad, A.; Dong, F.; Hong, D.; Yang, G. Heterosis Derived from Nonadditive Effects of the BnFLC Homologs Coordinates Early Flowering and High Yield in Rapeseed (Brassica napus L.). Front. Plant Sci. 2022, 12, 3351. [Google Scholar] [CrossRef]
- Rahman, H.; Bennett, R.A.; Kebede, B. Molecular Mapping of QTL Alleles of Brassica oleracea Affecting Days to Flowering and Photosensitivity in Spring Brassica napus. PLoS ONE 2018, 13, e0189723. [Google Scholar] [CrossRef]
- Shah, S.; Weinholdt, C.; Jedrusik, N.; Molina, C.; Zou, J.; Große, I.; Schiessl, S.; Jung, C.; Emrani, N. Whole-Transcriptome Analysis Reveals Genetic Factors Underlying Flowering Time Regulation in Rapeseed (Brassica napus L.). Plant Cell Environ. 2018, 41, 1935–1947. [Google Scholar] [CrossRef]
- Rahman, H.; Bennett, R.A.; Kebede, B. Mapping of Days to Flower and Seed Yield in Spring Oilseed Brassica napus Carrying Genome Content Introgressed from Brassica oleracea. Mol. Breed. 2017, 37, 5. [Google Scholar] [CrossRef]
- Cai, D.; Zhang, S.; Wang, J.; He, J.; Yi, L.; Zhu, J. Fine Mapping of a Minor QTL on Chromosome 2 Controlling Flowering Time in Brassica napus L. Genet. Resour. Crop. Evol. 2022, 69, 1137–1146. [Google Scholar] [CrossRef]
- Chao, H.; Wang, H.; Wang, X.; Guo, L.; Gu, J.; Zhao, W.; Li, B.; Chen, D.; Raboanatahiry, N.; Li, M. Genetic Dissection of Seed Oil and Protein Content and Identification of Networks Associated with Oil Content in Brassica napus. Sci. Rep. 2017, 7, srep46295. [Google Scholar] [CrossRef] [PubMed]
- Chao, H.; Guo, L.; Zhao, W.; Li, H.; Li, M. A Major Yellow-Seed QTL on Chromosome A09 Significantly Increases the Oil Content and Reduces the Fiber Content of Seed in Brassica napus. Theor. Appl. Genet. 2022, 135, 1293–1305. [Google Scholar] [CrossRef] [PubMed]
- Holzenkamp, K.; Link, W.; Möllers, C. Effect of a Low Seed Lignin Content on Seed Yield and Quality in Winter Oilseed Rape (Brassica napus). Plant Breed. 2022, 141, 801–806. [Google Scholar] [CrossRef]
- Widiarsih, S.; Nagel, M.; Börner, A.; Feussner, K.; Feussner, I.; Möllers, C. Inheritance of Seed Quality and Seed Germination in Two Doubled Haploid Populations of Oilseed Rape Segregating for Acid Detergent Lignin (ADL) Content. Euphytica 2021, 217, 161. [Google Scholar] [CrossRef]
- Geng, X.; Jiang, C.; Yang, J.; Wang, L.; Wu, X.; Wei, W. Rapid Identification of Candidate Genes for Seed Weight Using the SLAF-Seq Method in Brassica napus. PLoS ONE 2016, 11, e0147580. [Google Scholar] [CrossRef]
- Wang, H.; Yan, M.; Xiong, M.; Wang, P.; Liu, Y.; Xin, Q.; Wan, L.; Yang, G.; Hong, D. Genetic Dissection of Thousand-Seed Weight and Fine Mapping of CqSW.A03-2 via Linkage and Association Analysis in Rapeseed (Brassica napus L.). Theor. Appl. Genet. 2020, 133, 1321–1335. [Google Scholar] [CrossRef]
- Zhang, X.; Huang, Q.; Wang, P.; Liu, F.; Luo, M.; Li, X.; Wang, Z.; Wan, L.; Yang, G.; Hong, D. A 24,482-Bp Deletion Is Associated with Increased Seed Weight in Brassica napus L. Theor. Appl. Genet. 2021, 134, 2653–2669. [Google Scholar] [CrossRef]
- Qasim, M.U.; Zhao, Q.; Shahid, M.; Samad, R.A.; Ahmar, S.; Wu, J.; Fan, C.; Zhou, Y. Identification of QTLs Containing Resistance Genes for Sclerotinia Stem Rot in Brassica napus Using Comparative Transcriptomic Studies. Front. Plant Sci. 2020, 11, 776. [Google Scholar] [CrossRef]
- Dakouri, A.; Zhang, X.; Peng, G.; Falk, K.C.; Gossen, B.D.; Strelkov, S.E.; Yu, F. Analysis of Genome-Wide Variants through Bulked Segregant RNA Sequencing Reveals a Major Gene for Resistance to Plasmodiophora brassicae in Brassica oleracea. Sci. Rep. 2018, 8, 17657. [Google Scholar] [CrossRef]
- Botero-Ramírez, A.; Laperche, A.; Guichard, S.; Jubault, M.; Gravot, A.; Strelkov, S.E.; Manzanares-Dauleux, M.J. Clubroot Symptoms and Resting Spore Production in a Doubled Haploid Population of Oilseed Rape (Brassica napus) Are Controlled by Four Main QTLs. Front. Plant Sci. 2020, 11, 1998. [Google Scholar] [CrossRef]
- Shaikh, R.; Farid, M.; Rahman, H. Inheritance of Resistance to the Newly Identified Plasmodiophora brassicae Pathotypes in Brassica napus L. Can. J. Plant Pathol. 2020, 43, 256–266. [Google Scholar] [CrossRef]
- Gwiazdowski, R.; Gwiazdowska, D. Oddziaływanie Mieszanych Kultur Bakterii Fermentacji Propionowej i Mlekowej Na Grzyby Patogeniczne. Prog. Plant Prot. 2008, 48, 719–723. [Google Scholar]
- Howlett, B.J. Current Knowledge of the Interaction between Brassica napus and Leptosphaeria maculans. Can. J. Plant Pathol. 2010, 26, 245–252. [Google Scholar] [CrossRef]
- Balesdent, M.-H.; Brun, H.; Hayden, H.L.; Pinochet, X. Special Issue on Clubroot and Blackleg Diseases of Brassicas View Project Transcriptional and Chromatin-Based Control of Gene Expression and Pathogenicity in the Plant Pathogenic Fungus Leptosphaeria maculans View Project. Eur. J. Plant Pathol. 2006, 114, 33–40. [Google Scholar] [CrossRef]
- Sivasithamparam, K.; Barbetti, M.J.; Li, H. Recurring Challenges from a Necrotrophic Fungal Plant Pathogen: A Case Study with Leptosphaeria maculans (Causal Agent of Blackleg Disease in Brassicas) in Western Australia. Ann. Bot. 2005, 96, 363–377. [Google Scholar] [CrossRef] [PubMed]
- Raman, R.; Taylor, B.; Lindbeck, K.; Coombes, N.; Barbulescu, D.; Salisbury, P.; Raman, H.; Raman, R.; Taylor, B.; Lindbeck, K.; et al. Molecular Mapping and Validation of Rlm1 Gene for Resistance to Leptosphaeria maculans in Canola (Brassica napus L.). Crop. Pasture Sci. 2012, 63, 1007–1017. [Google Scholar] [CrossRef]
- Fredua-Agyeman, R.; Coriton, O.; Huteau, V.; Parkin, I.A.P.; Chèvre, A.M.; Rahman, H. Molecular Cytogenetic Identification of B Genome Chromosomes Linked to Blackleg Disease Resistance in Brassica napus × B. carinata Interspecific Hybrids. Theor. Appl. Genet. 2014, 127, 1305–1318. [Google Scholar] [CrossRef]
- Mayerhofer, R.; Wilde, K.; Mayerhofer, M.; Lydiate, D.; Bansal, V.K.; Good, A.G.; Parkin, I.A.P. Complexities of Chromosome Landing in a Highly Duplicated Genome: Toward Map-Based Cloning of a Gene Controlling Blackleg Resistance in Brassica napus. Genetics 2005, 171, 1977–1988. [Google Scholar] [CrossRef]
- Chèvre, A.M.; Barret, P.; Eber, F.; Dupuy, P.; Brun, H.; Tanguy, X.; Renard, M. Selection of Stable Brassica napus-B. juncea Recombinant Lines Resistant to Blackleg (Leptosphaeria maculans). 1. Identification of Molecular Markers, Chromosomal and Genomic Origin of the Introgression. Theor. Appl. Genet. 1997, 95, 1104–1111. [Google Scholar] [CrossRef]
- Long, Y.; Wang, Z.; Sun, Z.; Fernando, D.W.G.; McVetty, P.B.E.; Li, G. Identification of Two Blackleg Resistance Genes and Fine Mapping of One of These Two Genes in a Brassica napus Canola Cultivar “Surpass 400”. Theor. Appl. Genet. 2011, 122, 1223–1231. [Google Scholar] [CrossRef]
- Kaur, S.; Cogan, N.O.I.; Ye, G.; Baillie, R.C.; Hand, M.L.; Ling, A.E.; McGearey, A.K.; Kaur, J.; Hopkins, C.J.; Todorovic, M.; et al. Genetic Map Construction and QTL Mapping of Resistance to Blackleg (Leptosphaeria maculans) Disease in Australian Canola (Brassica napus L.) Cultivars. Theor. Appl. Genet. 2009, 120, 71–83. [Google Scholar] [CrossRef] [PubMed]
- Raman, H.; McVittie, B.; Pirathiban, R.; Raman, R.; Zhang, Y.; Barbulescu, D.M.; Qiu, Y.; Liu, S.; Cullis, B. Genome-Wide Association Mapping Identifies Novel Loci for Quantitative Resistance to Blackleg Disease in Canola. Front. Plant Sci. 2020, 11, 1184. [Google Scholar] [CrossRef] [PubMed]
- Amas, J.; Anderson, R.; Edwards, D.; Cowling, W.; Batley, J. Status and Advances in Mining for Blackleg (Leptosphaeria maculans) Quantitative Resistance (QR) in Oilseed Rape (Brassica napus). Theor. Appl. Genet. 2021, 134, 3123–3145. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| In Vitro | Semi-In Vitro/In Vivo | In Vivo | |||
|---|---|---|---|---|---|
| Gynogenesis | Androgenesis | Haploid inducer lines | Interploidy hybridization | Intraspecific hybridization Interspecific hybridization | |
| Flower buds Placenta Ovule Ovary | Anther Microspore | - | - | Spontaneous occurrence | |
| Yield | Haploid/Double haploid | Haploid/Double haploid | Amphihaploid | Double haploid | Haploid/Double haploid |
| Success rate in Brassica napus H/DH production | Low | Low/Medium | High | High | Low |
| Genotype dependent | Yes | Yes | No | Yes | Yes |
| Reliant on pathway conversion ability (saprophytic → sporophytic) | Yes | Yes | No | No | No |
| Chromosome elimination | No | No | Yes | Yes | Yes |
| Dependent on environmental conditions (i.e., medium composition, light, temperature, humidity) | Yes | Yes | No | Yes | Yes |
| Pretreatment | Yes (temperature, chemical pretreatment, irradiation etc.) | Yes (temperature, chemical pretreatment, irradiation etc.) | Yes (genetic modification) | No | No |
| References | [15,16] | [15,16,17,18,19] | [20,21] | [22] | [23] |
| Rapeseed Population | Trait | Experimental Conditions | Used Marker Type | Number of Genes/QTLs | Author |
|---|---|---|---|---|---|
| Darmor-bzh × Yudal Darmor × Samourai | Blackleg resistance | Field | SNP | 16 4 | [100] |
| Darmor-bzh × Yudal | Blackleg resistance | Field and greenhouse | DArT | 27 | [101] |
| Darmor-bzh × Yudal | Blackleg resistance | Greenhouse | SNP | 8 | [102] |
| Skipton × AgSpectrum | Blackleg resistance | Field and greenhouse | SSR, SRAP, SCAR | 8 | [103] |
| Topas × AGCastle Topas × AVSapphire | Blackleg resistance | Field | SSR, DArT | 22 21 | [104] |
| RP04 × Ag-Outback | Blackleg resistance | Field and greenhouse | DArT | 21 | [105] |
| ECD01 (B. rapa) × DH16516 (B. napus) | Clubroot | Greenhouse | SNP, | 2 | [106] |
| 1CA1446.476-A1296 × Hi-Q A04-73NA × Hi-Q | Clubroot | Greenhouse | SNP | 2 | [107] |
| T19 × ACDC | Clubroot | Greenhouse | SNP | 3 | [108] |
| 09CR500 × 09CR501 | Clubroot | Greenhouse | SNP, SSR | 2 | [109] |
| Mendel × A07-26NR | Clubroot | Greenhouse | SSR, | 5 | [110] |
| Zhongyou 821 × DHBao604 (H1), Zhongyou 821 × DH6576 (H2), Zhongyou 821 × Westar | Sclerotinia stem rot | Greenhouse | SNP | H1: 4–6 H2: 3–6 H3: 2–6 | [111] |
| ZP1 × D12 (B. napus inberd lines) | Sclerotinia stem rot | Field | SNP | 4 | [112] |
| Huashuang 5 × J7005 | Sclerotinia stem rot | Field | SSR | 13 | [113] |
| Bing 409 × Zhongshuang 8 | Flowering time | Field | SNP | 5 | [114] |
| IMC106RR × Wichita | Root morphology Flowering time Drought resistance | Field and greenhouse | SNP | 20 | [115] |
| Polo × Topas | Flowering time Fatty acid profile Oil content | Field | SSR | 14 131 14 | [116] |
| KenC-8 × N53-2 | Flowering time | Field | SNP | 55 (12 environment-stable, 43 environment-specific) | [117] |
| KenC-8 × N53-2 | Multi-main stem trait | Field | SNP, SSR, STS, SRAP, IFLP | 43 | [118] |
| SGDH284 × 158A (derived from Sollux and Zhoungyou9988 rapeseed cultivars) | Flowering time | Field | SNP | 56 | [119] |
| Low SD line No. 935 and high SD line No. 3641 | Seed density per silique | Field | SNP | 28 | [120] |
| GH06 × P174 (late flowering × early flowering | Flowering time | Field | SNP | 27 | [121] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Starosta, E.; Szwarc, J.; Niemann, J.; Szewczyk, K.; Weigt, D. Brassica napus Haploid and Double Haploid Production and Its Latest Applications. Curr. Issues Mol. Biol. 2023, 45, 4431-4450. https://doi.org/10.3390/cimb45050282
Starosta E, Szwarc J, Niemann J, Szewczyk K, Weigt D. Brassica napus Haploid and Double Haploid Production and Its Latest Applications. Current Issues in Molecular Biology. 2023; 45(5):4431-4450. https://doi.org/10.3390/cimb45050282
Chicago/Turabian StyleStarosta, Ewa, Justyna Szwarc, Janetta Niemann, Katarzyna Szewczyk, and Dorota Weigt. 2023. "Brassica napus Haploid and Double Haploid Production and Its Latest Applications" Current Issues in Molecular Biology 45, no. 5: 4431-4450. https://doi.org/10.3390/cimb45050282