

Molecular Evaluation of the Impact of Polymorphic Variants in Apoptotic (Bcl-2/Bax) and Proinflammatory Cytokine (TNF-α/IL-8) Genes on the Susceptibility and Progression of Myeloproliferative Neoplasms: A Case-Control Biomarker Study
 , and
, and
Abstract
:1. Introduction
2. Experimental Methods
2.1. Study Population
2.2. Inclusion and Exclusion Criteria
2.3. Peripheral Blood Collection from Controls and Patients Cohorts
2.4. Selection Criteria of Healthy Controls
2.5. Genomic DNA Extraction
2.6. Genotyping of Bcl-2-938 C>A, Bax G>A, TNF-α G>A and IL-8 T>A
2.7. PCR Cocktail and Optimization Procedure by a Gradient PCR
2.8. Thermocycling Conditions and Gel Electrophoresis
2.9. Statistical Analysis
3. Results
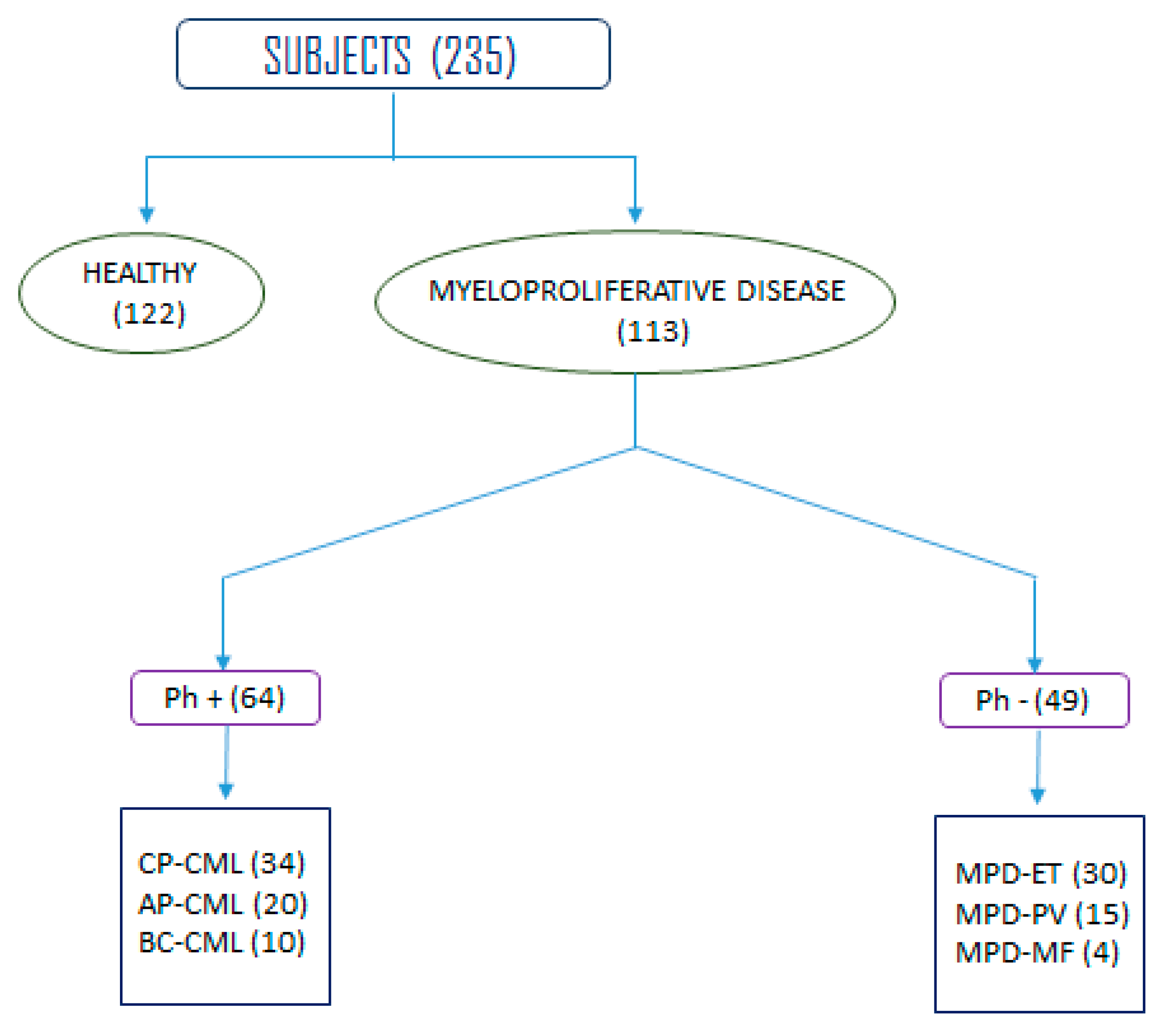
3.1. Demographic Characteristics of Study Population
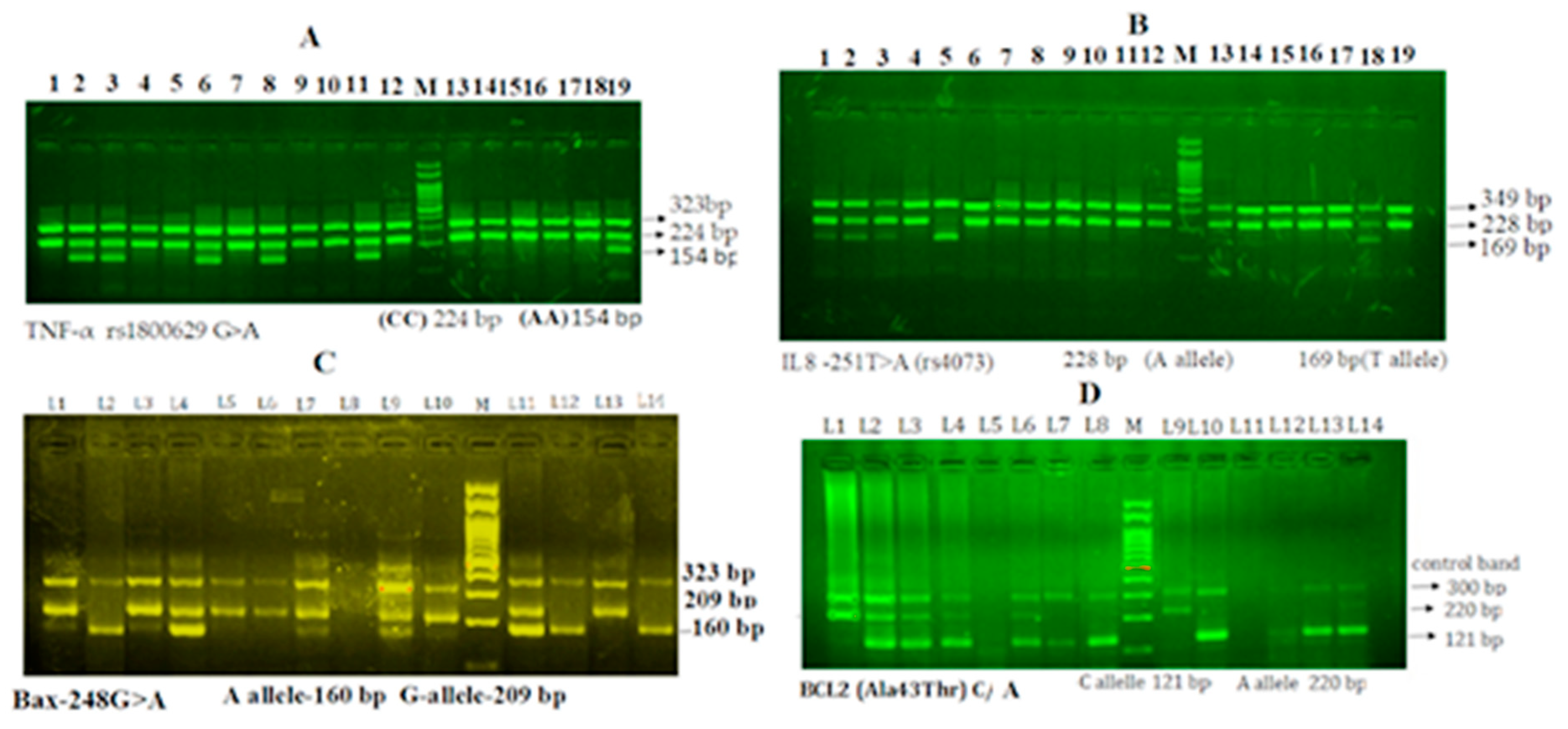
3.2. Genotyping and Gel Electrophoresis for the Identification of Alleles
3.3. Gel Electrophoresis of TNF-α rs1800629 G>A Gene Amplification
3.4. Gel Electrophoresis of Interleukin-8 rs4073 T>A Gene Amplification
3.5. Gel Electrophoresis of Bax G (−248) A Gene Amplification
3.6. Gel Electrophoresis of Bcl-2-938 C>A Gene Amplification
3.7. Comparative Analysis of the Bcl-2-938 C>A rs2279115, Bax-248G>A, TNF-α rs1800629 G>A and Interleukin-8 rs4073 T>A (IL-8-251T>A) Genotypes in Myeloproliferative Leukemia Patients and Healthy Controls
3.8. Logistic Regression Analysis of Bcl-2-938 C>A in Leukemia Patients
3.9. Logistic Regression Analysis of Bax-248 G>A in Leukemia Patients
3.10. Logistic Regression Analysis of IL-8 rs4073 T>A Genotypes to Predict the Risk of Leukemia
3.11. Logistic Regression Analysis of TNF-α rs1800629 G>A Genotypes to Predict the Risk of Leukemia
3.12. Genotypes Association with Clinico-Pathological Features of the Leukemia Patients
4. Discussion
5. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Correction Statement
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- IARC. Global Cancer Observatory: Cancer Today. 2020. Available online: http://gco.iarc.fr/today/home (accessed on 21 November 2021).
- Autret, A.; Martin, S.J. Emerging Role for Members of the Bcl-2 Family in Mitochondrial Morphogenesis. Mol. Cell 2009, 36, 355–363. [Google Scholar] [CrossRef] [PubMed]
- Basu, A. The interplay between apoptosis and cellular senescence: Bcl-2 family proteins as targets for cancer therapy. Pharmacol. Ther. 2021, 230, 107943. [Google Scholar] [CrossRef] [PubMed]
- Cory, S.; Adams, J.M. The Bcl2 family: Regulators of the cellular life-or-death switch. Nat. Rev. Cancer 2002, 2, 647–656. [Google Scholar] [CrossRef] [PubMed]
- Kaur, D.; Deshmukh, R. Chapter 2—Physiology of cellular demise: Apoptosis, necrosis, and autophagy. In Clinical Perspectives and Targeted Therapies in Apoptosis; Sodhi, R.K., Madan, J., Eds.; Academic Press: Cambridge, MA, USA, 2021; pp. 23–78. [Google Scholar]
- Llambi, F.; Moldoveanu, T.; Tait, S.W.; Bouchier-Hayes, L.; Temirov, J.; McCormick, L.L.; Dillon, C.P.; Green, D.R. A Unified Model of Mammalian BCL-2 Protein Family Interactions at the Mitochondria. Mol. Cell 2011, 44, 517–531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strasser, A.; Cory, S.; Adams, J.M. Deciphering the rules of programmed cell death to improve therapy of cancer and other diseases. EMBO J. 2011, 30, 3667–3683. [Google Scholar] [CrossRef] [PubMed]
- Hengartner, M.O. The biochemistry of apoptosis. Nature 2000, 407, 770–776. [Google Scholar] [CrossRef]
- Handa, H.; Hegde, U.P.; Kotelnikov, V.M.; Mundle, S.D.; Dong, L.M.; Burke, P.; Rose, S.; Gaskin, F.; Raza, A.; Preisler, H.D. Bcl-2 and c-myc expression, cell cycle kinetics and apoptosis during the progression of chronic myelogenous leukemia from diagnosis to blastic phase. Leuk. Res. 1997, 21, 479–489. [Google Scholar] [CrossRef]
- Bachmann, H.S.; Otterbach, F.; Callies, R.; Nückel, H.; Bau, M.; Schmid, K.W.; Siffert, W.; Kimmig, R. The AA Genotype of the Regulatory BCL2 Promoter Polymorphism (−938C>A) Is Associated with a Favorable Outcome in Lymph Node–Negative Invasive Breast Cancer Patients. Clin. Cancer Res. 2007, 13, 5790–5797. [Google Scholar] [CrossRef] [Green Version]
- Heubner, M.; Wimberger, P.; Otterbach, F.; Kasimir-Bauer, S.; Siffert, W.; Kimmig, R.; Nuckel, H. Association of the AA genotype of the Bcl-2 (-938C > A) promoter polymorphism with better survival in ovarian cancer. Int. J. Biol. Markers 2009, 24, 223–229. [Google Scholar] [CrossRef]
- Hirata, H.; Hinoda, Y.; Nakajima, K.; Kikuno, N.; Suehiro, Y.; Tabatabai, Z.L.; Ishii, N.; Dahiya, R. The bcl2 −938CC Genotype Has Poor Prognosis and Lower Survival in Renal Cancer. J. Urol. 2009, 182, 721–727. [Google Scholar] [CrossRef] [PubMed]
- Künkele, A.; Grosse-Lordemann, A.; Schramm, A.; Eggert, A.; Schulte, J.H.; Bachmann, H.S. The BCL2-938 C > A promoter polymorphism is associated with risk group classification in children with acute lymphoblastic leukemia. BMC Cancer 2013, 13, 452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moon, J.H.; Sohn, S.K.; Lee, M.-H.; Jang, J.H.; Kim, K.; Jung, C.W.; Kim, D.H. BCL2 gene polymorphism could predict the treatment outcomes in acute myeloid leukemia patients. Leuk. Res. 2010, 34, 166–172. [Google Scholar] [CrossRef]
- El Hindy, N.; Bachmann, H.S.; Lambertz, N.; Adamzik, M.; Nückel, H.; Worm, K.; Zhu, Y.; Sure, U.; Siffert, W.; Sandalcioglu, I.E. Association of the CC genotype of the regulatory BCL2 promoter polymorphism (−938C>A) with better 2-year survival in patients with glioblastoma multiforme. J. Neurosurg. 2011, 114, 1631–1639. [Google Scholar] [CrossRef] [PubMed]
- Masago, K.; Togashi, Y.; Fujita, S.; Nagai, H.; Sakamori, Y.; Okuda, C.; Kim, Y.H.; Mishima, M. Effect of the BCL2 Gene Polymorphism on Survival in Advanced-Stage Non-Small Cell Lung Cancer Patients Who Received Chemotherapy. Oncology 2013, 84, 214–218. [Google Scholar] [CrossRef]
- Chou, D.; Miyashita, T.; Mohrenweiser, H.W.; Ueki, K.; Kastury, K.; Druck, T.; von Deimling, A.; Huebner, K.; Reed, J.C.; Louis, D.N. The Bax gene maps to the glioma candidate region at 19q13.3, but is not altered in human gliomas. Cancer Genet. Cytogenet. 1996, 88, 136–140. [Google Scholar] [CrossRef]
- Sahu, S.K.; Choudhuri, T. Lack of Association between Bax Promoter (-248G>A) Single Nucleotide Polymorphism and Susceptibility towards Cancer: Evidence from a Meta-Analysis. PLoS ONE 2013, 8, e77534. [Google Scholar] [CrossRef]
- Bukholm, I.K.; Nesland, J.M. Protein expression of p53, p21 (WAF1/CIP1), Bcl-2, Bax, cyclin D1 and pRb in human colon carcinomas. Virchows Arch.-Int. J. Pathol. 2000, 436, 224–228. [Google Scholar] [CrossRef]
- Linjawi, A.; Kontogiannea, M.; Halwani, F.; Edwardes, M.; Meterissian, S. Prognostic significance of p53, Bcl-2, and Bax expression in early breast cancer. J. Am. Coll. Surg. 2004, 198, 83–90. [Google Scholar] [CrossRef]
- Dorjgochoo, T.; Xiang, Y.-B.; Long, J.; Shi, J.; Deming, S.; Xu, W.-H.; Cai, H.; Cheng, J.; Cai, Q.; Zheng, W.; et al. Association of Genetic Markers in the BCL-2 Family of Apoptosis-Related Genes with Endometrial Cancer Risk in a Chinese Population. PLoS ONE 2013, 8, e60915. [Google Scholar] [CrossRef]
- Li, W.; Qian, C.; Wang, L.; Teng, H.; Zhang, L. Association of BCL2-938C>A genetic polymorphism with glioma risk in Chinese Han population. Tumor Biol. 2014, 35, 2259–2264. [Google Scholar] [CrossRef]
- Starczynski, J.; Pepper, C.; Pratt, G.; Hooper, L.; Thomas, A.; Milligan, D.; Bentley, P.; Fegan, C. Common Polymorphism G(-248)A in the Promoter Region of the bax Gene Results in Significantly Shorter Survival in Patients With Chronic Lymphocytic Leukemia Once Treatment Is Initiated. J. Clin. Oncol. 2005, 23, 1514–1521. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Li, X.; Tao, K.; Jiang, L.; Ma, T.; Yan, S.; Yuan, C.; Moran, M.S.; Liang, F.; Haffty, B.G.; et al. BCL-2 (-938C > A) polymorphism is associated with breast cancer susceptibility. BMC Med. Genet. 2011, 12, 48. [Google Scholar] [CrossRef] [Green Version]
- Singh, N.; Baby, D.; Rajguru, J.P.; Patil, P.B.; Thakkannavar, S.S.; Pujari, V.B. Inflammation and cancer. Ann. Afr. Med. 2019, 18, 121–126. [Google Scholar] [CrossRef]
- Lan, T.; Chen, L.; Wei, X. Inflammatory Cytokines in Cancer: Comprehensive Understanding and Clinical Progress in Gene Therapy. Cells 2021, 10, 100. [Google Scholar] [CrossRef]
- Grunnet, L.G.; Aikin, R.; Tonnesen, M.F.; Paraskevas, S.; Blaabjerg, L.; Størling, J.; Rosenberg, L.; Billestrup, N.; Maysinger, D.; Mandrup-Poulsen, T. Proinflammatory cytokines activate the intrinsic apoptotic pathway in beta-cells. Diabetes 2009, 58, 1807–1815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Place, D.E.; Kanneganti, T.-D. Cell death–mediated cytokine release and its therapeutic implications. J. Exp. Med. 2019, 216, 1474–1486. [Google Scholar] [CrossRef]
- Dettmer, R.; Niwolik, I.; Cirksena, K.; Yoshimoto, T.; Tang, Y.; Mehmeti, I.; Gurgul-Convey, E.; Naujok, O. Proinflammatory cytokines induce rapid, NO-independent apoptosis, ex-pression of chemotactic mediators and interleukin-32 secretion in human pluripotent stem cell-derived beta cells. Diabetologia 2022, 65, 829–843. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; O’Malley, J.; Chaudhary, A.K.; Inigo, J.R.; Yadav, N.; Kumar, R.; Chandra, D. Hsp60 and IL-8 axis promotes apoptosis resistance in cancer. Br. J. Cancer 2019, 121, 934–943. [Google Scholar] [CrossRef] [PubMed]
- Landskron, G.; De la Fuente, M.; Thuwajit, P.; Thuwajit, C.; Hermoso, M.A. Chronic Inflammation and Cytokines in the Tumor Microenvironment. J. Immunol. Res. 2014, 2014, 149185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amin, M.N.; Siddiqui, S.A.; Ibrahim, M.; Hakim, M.L.; Ahammed, M.S.; Kabir, A.; Sultana, F. Inflammatory cytokines in the pathogenesis of cardiovascular disease and cancer. SAGE Open Med. 2020, 8, 2050312120965752. [Google Scholar] [CrossRef]
- Montfort, A.; Colacios, C.; Levade, T.; Andrieu-Abadie, N.; Meyer, N.; Ségui, B. The TNF Paradox in Cancer Progression and Im-munotherapy. Front. Immunol. 2019, 10, 1818. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Li, Z.; Zhou, J. Tumor necrosis factor α in the onset and progression of leukemia. Exp. Hematol. 2017, 45, 17–26. [Google Scholar] [CrossRef]
- Min, L.; Chen, D.; Qu, L.; Shou, C. Tumor Necrosis Factor-A Polymorphisms and Colorectal Cancer Risk: A Meta-Analysis. PLoS ONE 2014, 9, e85187. [Google Scholar] [CrossRef] [PubMed]
- Bernhard, S.; Hug, S.; Stratmann, A.E.P.; Erber, M.; Vidoni, L.; Knapp, C.L.; Thomaß, B.D.; Fauler, M.; Nilsson, B.; Ekdahl, K.N.; et al. Interleukin 8 Elicits Rapid Physiological Changes in Neutrophils That Are Altered by Inflammatory Conditions. J. Innate Immun. 2021, 13, 225–241. [Google Scholar] [CrossRef] [PubMed]
- Kuett, A.; Rieger, C.; Perathoner, D.; Herold, T.; Wagner, M.; Sironi, S.; Sotlar, K.; Horny, H.-P.; Deniffel, C.; Drolle, H.; et al. IL-8 as mediator in the microenvironment-leukaemia network in acute myeloid leukaemia. Sci. Rep. 2015, 5, 18411. [Google Scholar] [CrossRef] [Green Version]
- Tobler, A.; Moser, B.; Dewald, B.; Geiser, T.; Studer, H.; Baggiolini, M.; Fey, M.F. Constitutive Expression of Interleukin-8 and its Re-ceptor in Human Myeloid and Lymphoid Leukemia. Blood 1993, 82, 2517–2525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allegra, A.; Musolino, C.; Tonacci, A.; Pioggia, G.; Casciaro, M.; Gangemi, S. Clinico-Biological Implications of Modified Levels of Cytokines in Chronic Lymphocytic Leukemia: A Possible Therapeutic Role. Cancers 2020, 12, 524. [Google Scholar] [CrossRef] [Green Version]
- Lello, L.; Raben, T.G.; Yong, S.Y.; Tellier, L.C.; Hsu, S.D. Genomic Prediction of 16 Complex Disease Risks Including Heart Attack, Diabetes, Breast and Prostate Cancer. Sci. Rep. 2019, 9, 15286. [Google Scholar] [CrossRef] [Green Version]
- Butt, H.; Khalid, A.; Yaqoob, M. Polymorphisms of BCL2 Gene in Acute Lymphoblastic Leukemia Patients in Pa-kistan and Screening of Phytochemicals to Overcome its Expression. Biol. Med. 2017, 10, 424. [Google Scholar] [CrossRef]
- Matheson, M.C.; Ellis, J.A.; Raven, J.; Walters, E.H.; Abramson, M.J. Association of IL8, CXCR2 and TNF-alpha polymorphisms and airway disease. J. Hum. Genet. 2006, 51, 196–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jurlander, J. Hematological Malignancies, Leukemias and Lymphomas. In Encyclopedia of Cancer; Schwab, M., Ed.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 1640–1644. [Google Scholar] [CrossRef]
- Senapati, J.; Jabbour, E.; Kantarjian, H.; Short, N.J. Pathogenesis and management of accelerated and blast phases of chronic myeloid leukemia. Leukemia 2023, 37, 5–17. [Google Scholar] [CrossRef] [PubMed]
- Popp, H.D.; Kohl, V.; Naumann, N.; Flach, J.; Brendel, S.; Kleiner, H.; Weiss, C.; Seifarth, W.; Saussele, S.; Hofmann, W.K.; et al. DNA damage and dna damage response in chronic myeloid leukemia. Int. J. Mol. Sci. 2020, 21, 1177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdulmawjood, B.; Costa, B.; Roma-Rodrigues, C.; Baptista, P.V.; Fernandes, A.R. Genetic Biomarkers in Chronic Myeloid Leukemia: What Have We Learned So Far? Int. J. Mol. Sci. 2021, 22, 12516. [Google Scholar] [CrossRef]
- Provan, D.; Gribben, J. Molecular Hematology; John Wiley & Sons: New York, NY, USA, 2010. [Google Scholar]
- Spivak, J.L. The chronic myeloproliferative disorders: Clonality and clinical heterogeneity. Semin. Hematol. 2004, 41, 1–5. [Google Scholar] [CrossRef]
- Bonzheim, I.; Geissinger, E.; Chuang, W.Y.; Roth, S.; Strobel, P.; Marx, A.; Reimer, P.; Wilhelm, M.; Puppe, B.; Rosenwald, A.; et al. Analysis of single nucleotide polymorphisms in the FAS and CTLA-4 genes of pe-ripheral T-cell lymphomas. J. Hematop. 2008, 1, 11–21. [Google Scholar] [CrossRef] [Green Version]
- Knoefel, L.F.; Werle-Schneider, G.; Dally, H.; Müller, P.J.; Edler, L.; Bartsch, H.; Tuengerthal, S.; Heussel, C.P.; Reinmuth, N.; Thomas, M.; et al. Polymorphisms in the Apoptotic Pathway Gene BCL-2 and Survival in Small Cell Lung Cancer. J. Thorac. Oncol. 2011, 6, 183–189. [Google Scholar] [CrossRef] [Green Version]
- Bachmann, H.S.; Heukamp, L.C.; Schmitz, K.J.; Hilburn, C.F.; Kahl, P.; Buettner, R.; Nückel, H.; Eisenhardt, A.; Rübben, H.; Schmid, K.W.; et al. Regulatory BCL2 promoter polymorphism (−938C>A) is associated with adverse outcome in patients with prostate carcinoma. Int. J. Cancer 2011, 129, 2390–2399. [Google Scholar] [CrossRef]
- Kim, D.H.; Xu, W.; Ma, C.; Liu, X.; Siminovitch, K.; Messner, H.A.; Lipton, J.H. Genetic variants in the candidate genes of the apoptosis pathway and susceptibility to chronic myeloid leukemia. Blood 2009, 113, 2517–2525. [Google Scholar] [CrossRef] [Green Version]
- Pirocanac, E.C.; Nassirpour, R.; Yang, M.; Wang, J.; Nardin, S.R.; Gu, J.; Fang, B.; Moossa, A.; Hoffman, R.M.; Bouvet, M. Bax-Induction Gene Therapy of Pancreatic Cancer. J. Surg. Res. 2002, 106, 346–351. [Google Scholar] [CrossRef] [Green Version]
- Sturm, I.; Kohne, C.H.; Wolff, G.; Petrowsky, H.; Hillebrand, T.; Hauptmann, S.; Lorenz, M.; Dorken, B.; Daniel, P.T. Analysis of the p53/Bax pathway in colorectal cancer: Low Bax is a negative prognostic factor in patients with resected liver metastases. J. Clin. Oncol. 1999, 17, 1364–1374. [Google Scholar] [CrossRef] [PubMed]
- Miquel, C.; Borrini, F.; Grandjouan, S.; Aupérin, A.; Viguier, J.; Velasco, V.; Duvillard, P.; Praz, F.; Sabourin, J.-C. Role of bax Mutations in Apoptosis in Colorectal Cancers With Microsatellite Instability. Am. J. Clin. Pathol. 2005, 123, 562–570. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.Y.; Han, J.H.; Lee, K.J.; Choi, J.-H.; Park, J.I.; Kim, H.I.; Lee, H.-W.; Jang, J.H.; Park, J.S.; Kim, H.C.; et al. Low Expression of Bax Predicts Poor Prognosis in Patients with Locally Advanced Esophageal Cancer Treated with Definitive Chemoradiotherapy. Clin. Cancer Res. 2007, 13, 4146–4153. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Wei, L.Y.; Wei, L.; Li, D.M. Correlation between Bax gene polymorphisms and esophagus cancer. Oncol. Lett. 2018, 16, 7097–7101. [Google Scholar] [CrossRef] [Green Version]
- Porebska, I.; Wyrodek, E.; Kosacka, M.; Adamiak, J.; Jankowska, R.; Harłozińska-Szmyrka, A. Apoptotic markers p53, Bcl-2 and Bax in primary lung cancer. Vivo 2006, 20, 599–604. [Google Scholar]
- Jeong, S.H.; Lee, H.W.; Han, J.H.; Kang, S.Y.; Choi, J.-H.; Jung, Y.M.; Choi, H.; Oh, Y.T.; Park, K.J.; Hwang, S.C.; et al. Low Expression of Bax Predicts Poor Prognosis in Resected Non-small Cell Lung Cancer Patients with Non-squamous Histology. Jpn. J. Clin. Oncol. 2008, 38, 661–669. [Google Scholar] [CrossRef]
- Chen, K.X.; Hu, Z.B.; Wang, L.E.; Sturgis, E.M.; El-Naggar, A.K.; Zhang, W.; Wei, Q.Y. Single-nucleotide poly-morphisms at the TP53-binding or responsive promoter regions of Bax and Bcl-2 genes and risk of squamous cell carcinoma of the head and neck. Carcinogenesis 2007, 28, 2008–2012. [Google Scholar] [CrossRef]
- Anvari, K.; Toussi, M.S.; Kalantari, M.; Naseri, S.; Shahri, M.K.; Ahmadnia, H.; Katebi, M.; Pashaki, A.S.; Dayani, M.; Broumand, M. Expression of Bcl-2 and Bax in Advanced or Metastatic Prostate Carcinoma. Urol. J. 2012, 9, 381–388. [Google Scholar] [CrossRef]
- Yigit, S.; Demir, L.; Tarhan, M.O.; Cabuk, F.K.; Ellidokuz, H.; Erten, C.; Somali, I.; Dirican, A.; Cakalagaoglu, F.; Çakalaǧaoǧlu, F. The clinicopathological significance of Bax and Bcl-2 protein expression with tumor infiltrating lymphocytes in ovarian carcinoma. Neoplasma 2012, 59, 475–485. [Google Scholar] [CrossRef] [PubMed]
- Lopes-Costa, P.V.; dos Santos, A.R.; da Silva, B.B. The effect of raloxifene on Bax protein expression in breast carcinomas of postmenopausal women. Diagn. Cytopathol. 2012, 40, 570–574. [Google Scholar] [CrossRef] [PubMed]
- Knudson, C.M.; Tung, K.S.K.; Tourtellotte, W.G.; Brown, G.A.J.; Korsmeyer, S.J. Bax-Deficient Mice with Lymphoid Hyperplasia and Male Germ Cell Death. Science 1995, 270, 96–99. [Google Scholar] [CrossRef]
- Yin, C.; Knudson, C.M.; Korsmeyer, S.J.; Van Dyke, T. Bax suppresses tumorigenesis and stimulates apoptosis in vivo. Nature 1997, 385, 637–640. [Google Scholar] [CrossRef]
- Peng, H.; Aiello, A.; Packham, G.; Isaacson, P.G.; Pan, L. Infrequent bax gene mutations in B-cell lymphomas. J. Pathol. 1998, 186, 378–382. [Google Scholar] [CrossRef]
- Torrealba, N.; Rodríguez-Berriguete, G.; Vera, R.; Fraile, B.; Olmedilla, G.; Martínez-Onsurbe, P.; Sánchez-Chapado, M.; Paniagua, R.; Royuela, M. Homeostasis: Apoptosis and cell cycle in normal and pathological prostate. Aging Male 2021, 23, 335–345. [Google Scholar] [CrossRef]
- Alam, M.; Kashyap, T.; Mishra, P.; Panda, A.K.; Nagini, S.; Mishra, R. Role and regulation of pro-apoptotic Bax in oral squamous cell carcinoma and drug resistance. Head Neck-J. Sci. Spec. Head Neck 2019, 41, 185–197. [Google Scholar]
- Feng, Y.; Chen, X.L.; Zheng, Y.; Liu, Q.; Chen, H.W.; Cai, Y.H.; Cao, L.X.; Lai, X.L.; Pan, L.L.; Li, Y.; et al. Prognostic value and susceptibility of Bax rs4645878 polymorphism in cancer A systematic review and meta-analysis. Medicine 2018, 97, 8. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-C.; Wang, Z.-H.; Yen, J.-H.; Shen, Y.-C.; Shen, T.-C.; Chang, W.-S.; Su, C.-H.; Chen, K.-Y.; Yen, C.-M.; Lee, H.-T.; et al. The Contribution of Interleukin-8 Rs4073 Genotypes to Triple Negative Breast Cancer Risk in Taiwan. Anticancer Res. 2022, 42, 3799–3806. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Liu, J.-B.; Liu, X.; Qian, L. Association of interleukin 10 rs1800896 polymorphism with susceptibility to breast cancer: A meta-analysis. J. Int. Med. Res. 2020, 48, 0300060520904863. [Google Scholar] [CrossRef] [PubMed]
- Abdalhabib, E.K.; Alzahrani, B.; Saboor, M.; Hamza, A.; Elfaki, E.M.; Alanazi, F.; Alenazy, O.F.; Algarni, A.; Ibrahim, I.K.; Mohamed, A.H.; et al. IL-10 rs1800896 Polymorphism: A Risk Factor for Adult Acute Lymphoblastic Leukemia. Pharm. Pers. Med. 2022, 15, 809–815. [Google Scholar] [CrossRef]
- Singh, J.K.; Simões, B.M.; Howell, S.J.; Farnie, G.; Clarke, R. Recent advances reveal IL-8 signaling as a potential key to targeting breast cancer stem cells. Breast Cancer Res. BCR 2013, 15, 210. [Google Scholar] [CrossRef] [Green Version]
- Moghimi, M.; Dastgheib, S.A.; Heiranizadeh, N.; Zare, M.; Sheikhpour, E.; Neamatzadeh, H. Association of il-8 -251t>a (rs4073) polymorphism with susceptibility to gastric cancer: A systematic review and meta-analysis based on 33 case-control studies. Arq. Gastroenterol. 2020, 57, 91–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernando, R.I.; Castillo, M.D.; Litzinger, M.; Hamilton, D.H.; Palena, C. IL-8 signaling plays a critical role in the epitheli-al-mesenchymal transition of human carcinoma cells. Cancer Res. 2011, 71, 5296–5306. [Google Scholar] [CrossRef] [Green Version]
- Moreno-Guerrero, S.S.; Ramírez-Pacheco, A.; Rocha-Ramírez, L.M.; Hernández-Pliego, G.; Eguía-Aguilar, P.; Escobar-Sánchez, M.A.; Reyes-López, A.; Juárez-Villegas, L.E.; Sienra-Monge, J.J.L. Association of Genetic Polymorphisms and Serum Levels of IL-6 and IL-8 with the Prognosis in Children with Neuroblastoma. Cancers 2021, 13, 529. [Google Scholar] [CrossRef] [PubMed]
- Carswell, A.E.; Old, L.J.; Kassel, R.L.; Green, S.; Fiore, N.; Williamson, B. An endotoxin-induced serum factor that causes necrosis of tumors. Proc. Natl. Acad. Sci. USA 1975, 72, 3666–3670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tartaglia, L.A.; Ayres, T.; Wong, G.H.; Goeddel, D.V. A novel domain within the 55 kd TNF receptor signals cell death. Cell 1993, 74, 845–853. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Vincent, A.; Cates, J.; Brantley-Sieders, D.M.; Polk, D.B.; Young, P.P. Low Levels of Tumor Necrosis Factor α Increase Tumor Growth by Inducing an Endothelial Phenotype of Monocytes Recruited to the Tumor Site. Cancer Res. 2009, 69, 338–348. [Google Scholar] [CrossRef] [Green Version]
- Mehta, A.; Kim, Y.J.; Robert, L.; Tsoi, J.; Comin-Anduix, B.; Berent-Maoz, B.; Cochran, A.J.; Economou, J.S.; Tumeh, P.C.; Puig-Saus, C.; et al. Immunotherapy Resistance by Inflammation-Induced Dedifferentiation. Cancer Discov. 2018, 8, 935–943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.W.; Xia, W.; Huo, L.; Lim, S.O.; Wu, Y.; Hsu, J.L.; Chao, C.-H.; Yamaguchi, H.; Yang, N.-K.; Ding, Q.; et al. Epithelial-mesenchymal transition induced by TNF-α requires NF-κB-mediated transcriptional upregulation of Twist1. Cancer Res. 2012, 72, 1290–1300. [Google Scholar] [CrossRef] [Green Version]
- Liao, S.-J.; Luo, J.; Li, D.; Zhou, Y.-H.; Yan, B.; Wei, J.-J.; Tu, J.-C.; Li, Y.-R.; Zhang, G.-M.; Feng, Z.-H. TGF-β1 and TNF-α synergistically induce epithelial to mesenchymal transition of breast cancer cells by enhancing TAK1 activation. J. Cell Commun. Signal. 2019, 13, 369–380. [Google Scholar] [CrossRef]
- Verma, S.; Singh, A.; Yadav, G.; Kushwaha, R.; Ali, W.; Verma, S.P.; Singh, U. Serum Tumor Necrosis Factor-Alpha Levels in Acute Leukemia and Its Prognostic Significance. Cureus 2022, 14, e24835. [Google Scholar] [CrossRef]
- Abdalhabib, E.K.; Algarni, A.; Saboor, M.; Alanazi, F.; Ibrahim, I.K.; Alfeel, A.H.; Alanazi, A.M.; Alanazi, A.M.; Alruwaili, A.M.; Alanazi, M.H.; et al. Association of TNF-α rs1800629 with Adult Acute B-Cell Lymphoblastic Leukemia. Genes 2022, 13, 1237. [Google Scholar] [CrossRef] [PubMed]
- Bănescu, C.; Tripon, F.; Trifa, A.P.; Crauciuc, A.G.; Moldovan, V.G.; Bogliş, A.; Benedek, I.; Dima, D.; Cândea, M.; Duicu, C.; et al. Cytokine rs361525, rs1800750, rs1800629, rs1800896, rs1800872, rs1800795, rs1800470, and rs2430561 SNPs in relation with prognostic factors in acute myeloid leukemia. Cancer Med. 2019, 8, 5492–5506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abd El-Aziz, S.M.; Alkhiary, W.; Mokhtar, N.; Talaab, M. Tumor necrosis factor-α (TNF-α) − 308 G/A and lymphotoxin-α (LT-α) +252 A/G genetic polymorphisms in Egyptian acute lymphoblastic leukemia. Comp. Clin. Pathol. 2018, 27, 363–369. [Google Scholar] [CrossRef]
- Liu, K.; Hu, J.; Wang, X.; Li, L. Chronic myeloid leukemia blast crisis presented with AML of t(9;22) and t(3;14) mimicking acute lymphocytic leukemia. J. Clin. Lab. Anal. 2019, 33, e22961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ochi, Y.; Yoshida, K.; Huang, Y.J.; Kuo, M.C.; Nannya, Y.; Sasaki, K.; Mitani, K.; Hosoya, N.; Hiramoto, N.; Ishikawa, T.; et al. Clonal evolution and clinical implications of genetic abnormalities in blastic transfor-mation of chronic myeloid leukaemia. Nat. Commun. 2021, 12, 2833. [Google Scholar] [CrossRef]
- Alarcón-Payer, C.; Sánchez Suárez, M.D.M.; Martín Roldán, A.; Puerta Puerta, J.M.; Jiménez Morales, A. Impact of Genetic Polymorphisms and Biomarkers on the Effectiveness and Toxicity of Treatment of Chronic Myeloid Leukemia and Acute Myeloid Leukemia. J. Pers. Med. 2022, 12, 1607. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Direction | ARMS Primer Sequence | Annealing Tempt | Product Size |
|---|---|---|---|
| Bax G(−248) A rs4645878 gene polymorphism | |||
| Bax FO | 5′-CCT GGA AGC ATG CTA TTT TGGGCCT-3′ | 65 °C | 323 bp |
| Bax RO | 5′-ACG TGA GAG CCC CGC TGA ACG T-3′ | ||
| Bax FI-G | 5′-GGC ATT AGA GCT GCG ATT GGA CTG G-3′ | 209 bp | |
| Bax RI-A | 5′-AGTGGCGCCGTCCAACAGCAGT-3′ | 160 bp | |
| Bcl-2-938 C>A rs2279115 gene polymorphism | |||
| Bcl-2-FO | 5′-CCG GCT CCT TCA TCG TCT CC-3′ | 58 °C | 300 bp |
| Bcl-2-RO | 5′-CCC AGG AGA GAG ACA GGG GAA AT-3′ | ||
| Bcl-2-FI-A | 5′-AATAAAACCCTCCCCCACCACCT-3′ | 220 bp | |
| Bcl-2-RI-C | 5′-CCCTTCTCGGCAATTTACACGC-3′ | 121 bp | |
| TNF-α rs1800629 G>A gene polymorphism | |||
| TNF-αF0 | 5′-ACC CAA ACA CAG GCC TCA GGACTCAACA-3′ | 62 °C | 323 bp |
| TNF-αR0 | 5′-TGGAGGCAATAGCTTTTGAGGGGCAGGA-3′ | ||
| TNF-αFI-A | 5′-AGTTGGGGACACGCAAGCATGAAGGATA-3′ | 154 bp | |
| TNF-α RI-C | 5′-TAGGACCCTGGAGGCTAGACCCCGTACC-3′ | 224 bp | |
| IL8-251T>A (rs4073) gene polymorphism | |||
| IL8-Fo | 5′-CAT GAT AGC ATC TGT AAT TAA CTG-3′ | 58 °C | 349 bp |
| IL8-Ro | 5′-CACAATTTGGTGAATTATCAAA-3′ | ||
| IL8-FI-A | 5′-GTTATCTAGAAATAAAAAAGCATACAA-3′ | 228 bp | |
| IL8-RI-T | 5′-CTCATCTTTTCATTATGTCAGA-3′ | 169 bp | |
| Patients (113) | Controls (122) | |
|---|---|---|
| Age | N = % | N = % |
| >40 | 76 (67.21%) | 80 (65.57%) |
| <40 | 37 (32.78%) | 42 (34.42%) |
| Gender | N = % | N = % |
| Males | 82 (72.56%) | 74 (60.65%) |
| Females | 31 (27.44%) | 48 (39.34%) |
| Frequency of different stages of CML (Ph+) | ||
| N = 64 | ||
| Chronic phase CML | 34 (53.12%) | |
| Accelerated phase CML | 20 (31.25%) | |
| Blast crisis CML | 10 (15.62%) | |
| Frequency of other MPDs (Ph−) | ||
| MPD TYPE | N = 49 | |
| Essential thrombocytopenia | 30 (61.22%) | |
| Polycythemia verra | 15 (30.61%) | |
| Myelofibrosis | 04 (8.16%) | |
| Bcl-2-938 C>A rs2279115 | |||||||||
| Study Population | N= | CC % | CA % | AA % | C allele | A allele | DF | X2 | p Value |
| Patients | 100 | 34 (34.0%) | 44 (44.0%) | 22 (22.0%) | 0.56 | 0.44 | 2 | 7.32 | 0.025 |
| Healthy controls | 100 | 49 (49.0%) | 41 (41.0%) | 10 (10.0%) | 0.67 | 0.31 | |||
| Bax-248G>A | |||||||||
| N= | GG % | GA % | AA % | G allele | A allele | DF | X2 | p value | |
| Patients | 108 | 56 (51.85%) | 45 (41.66%) | 07 (6.48%) | 0.73 | 0.27 | 2 | 6.23 | 0.048 |
| Healthy controls | 110 | 75 (68.50%) | 30 (27.27%) | 5 (4.54%) | 0.82 | 0.18 | |||
| IL-8 rs4073 T>A | |||||||||
| Subjects | N= | TT | AT | AA | T | A | Df | X2 | p value |
| Patients | 113 | 19 (16.81%) | 41 (36.28%) | 53 (56.90%) | 0.25 | 0.75 | 2 | 20.24 | 0.0001 |
| Healthy controls | 122 | 48 (39.34%) | 46 (37.70%) | 28 (22.95%) | 0.39 | 61 | |||
| TNF-α rs1800629 G>A | |||||||||
| Subjects | N= | GG | GA | AA | G | A | Df | X2 | p value |
| Patients | 113 | 94 (83.18%) | 10 (8.84%) | 09 (7.96%) | 0.87 | 0.13 | 2 | 7.64 | 0.021 |
| Healthy controls | 122 | 100 (69.67%) | 20 (28.68%) | 02 (1.63%) | 0.84 | 0.16 | |||
| Controls | Patients | ||||
|---|---|---|---|---|---|
| Genotypes | (N = 100) | (N = 100) | Odd Ratio (95% CI) | Risk Ratio (95% CI) | p Value |
| Codominant | |||||
| Bcl-2 (CC) | 49 (49.0%) | 34 (34.0%) | (ref.) | (ref.) | |
| Bcl-2 (CA) | 41 (41.0%) | 44 (44.0%) | 1.540 (0.840–2.840) | 1.220 (0.920–1.620) | 0.160 |
| Bcl-2 (AA) | 10 (10.0%) | 22 (49.0%) | 3.170 (1.330–7.530) | 1.880 (1.090–3.250) | 0.009 |
| Dominant | |||||
| Bcl-2 (CC) | 49 (49.0%) | 34 (34.0%) | (ref.) | (ref.) | |
| Bcl-2 (CA + AA) | 51 (51.0%) | 66 (66.0%) | 1.860 (1.050–3.290) | 1.350 (1.030–1.770) | 0.030 |
| Recessive | |||||
| Bcl-2 (CC + CA) | 90 (90.0%) | 78 (78.0%) | (ref.) | (ref.) | |
| Bcl-2 (AA) | 10 (10.0%) | 22 (22.0%) | 2.530 (1.130–5.680) | 1.710 (1.006–2.920) | 0.020 |
| Allele | |||||
| Bcl-2 (C) | 139 (69.50%) | 112 (56.0%) | (ref.) | (ref.) | |
| Bcl-2 (A) | 61 (30.50%) | 88 (44.0%) | 1.790 (1.180–2.690) | 1.350 (1.08–1.68) | 0.005 |
| Genotypes | Controls | Patients | Odd Ratio (95% CI) | Risk Ratio (95% CI) | p Value | ||
|---|---|---|---|---|---|---|---|
| Inheritance Model | (N = 110) | % | (N = 108) | % | |||
| Codominant | |||||||
| Bax (GG) | 75 | 68.18% | 56 | 51.85% | (ref.) | (ref.) | |
| Bax (GA) | 30 | 27.27% | 45 | 41.66% | 2.0 (1.12–3.57) | 1.43 (1.045–1.95) | 0.01 |
| Bax (AA) | 05 | 4.54% | 07 | 4.68% | 1.87 (0.56–6.21) | 1.37 (0.69–2.72) | 0.30 |
| Dominant | |||||||
| Bax (GG) | 75 | 68.18 | 56 | 51.85 % | (ref.) | (ref.) | |
| Bax (GA + AA) | 35 | 31.82 | 52 | 48.15 % | 1.98 (1.14–3.45) | 1.43 (1.05–1.91) | 0. 01 |
| Recessive | |||||||
| Bax (GG + GA) | 105 | 95.45 | 101 | 93.52% | (ref.) | (ref.) | |
| Bax (AA) | 05 | 4.55 | 07 | 6.48 % | 1.45 (0.44–4.73) | 1.23 (0.61–2.42) | 0.53 |
| Allele | |||||||
| Bax (G) | 180 | 81.82 | 157 | 73.36% | (ref.) | (ref.) | |
| Bax (A) | 40 | 18.18 | 57 | 26.63% | 1.63 (1.03–2.58) | 1.29 (1.00–1.67) | 0.035 |
| Genotypes Inheritance Model | Controls (N = 122) | Patients (N = 113) | Odd Ratio (95% CI) | Risk Ratio (95% CI) | p Value |
|---|---|---|---|---|---|
| Codominant | |||||
| IL-8 (TT) | 48 | 19 | (ref.) | (ref.) | |
| IL-8 (TA) | 46 | 41 | 2.25 (1.1433 to 4.434) | 1.35 (1.0562 to 1.738) | 0.018 |
| IL-8 (AA) | 28 | 53 | 4.7 (2.3714 to 9.642) | 2.07 (1.4820 to 2.898) | 0.0001 |
| Dominant | |||||
| IL-8 (TT) | 48 | 19 | (ref.) | (ref.) | |
| IL-8-(TA + AA) | 74 | 94 | 3.20 (1.7395 to 5.920) | 1.62 (1.2956 to 2.041) | 0.0002 |
| Recessive | |||||
| IL-8 (TT + TA) | 94 | 60 | (ref.) | (ref.) | |
| IL-8- (AA) | 28 | 53 | 2.96 (1.6953 to 5.112) | 1.76 (1.2706 to 2.447) | 0.0001 |
| Allele | |||||
| IL-8 (T) | 142 | 79 | (ref.) | (ref.) | |
| IL-8-A | 102 | 147 | 2.59 (1.78057 to 3.7657) | 1.56 (1.314 to 1.871) | 0.0001 |
| Over Dominant | |||||
| IL-8 (TT + AA) | 76 | 72 | (ref.) | (ref.) | |
| IL-8 (TA) | 46 | 41 | 0.94 (0.5536 to 1.5924) | 0.97 (0.758 to 1.2723) | 0.82 |
| Genotypes | Controls | Patients | Odd Ratio (95% CI) | Risk Ratio (95% CI) | p Value |
|---|---|---|---|---|---|
| Inheritance Model | (N = 122) | (N = 113) | |||
| Codominant | |||||
| TNF-α (GG) | 100 | 94 | (ref.) | (ref.) | |
| TNF-α (GA) | 20 | 11 | 0.58 (0.2661 to 1.286) | 0.79 (0.5951 to 1.072) | 0.182 |
| TNF-α (AA) | 02 | 08 | 4.78 (1.0081 to 22.732) | 2.83 (0.8034 to 10.005) | 0.048 |
| Dominant | |||||
| TNF-α (GG) | 100 | 94 | (ref.) | (ref.) | |
| TNF-α (GA+AA) | 22 | 19 | 0.96 (0.4960 to 1.885) | 0.98 (0.7153 to 1.353) | 0.92 |
| Recessive | |||||
| TNF-α (GA+GG) | 120 | 105 | (ref.) | (ref.) | |
| TNF-α (AA) | 02 | 09 | 5.14 (1.0867 to 24.337) | 2.93 (0.8324 to 10.336) | 0.038 |
| Allele | |||||
| TNF-α (G) | 220 | 199 | (ref.) | (ref.) | |
| TNF-α (A) | 24 | 29 | 1.33 (0.7526 to 2.371) | 1.15 (0.8507 to 1.580) | 0.32 |
| Over dominant | |||||
| TNF-α-(GG+AA) | 102 | 105 | (ref.) | (ref.) | |
| TNF-α (GA) | 20 | 08 | 0.53 (0.2438 to 1.170) | 0.76 (0.5684 to 1.026) | 0.11 |
| Bcl-2-938 C>A genotypes | ||||||||
| Gender | N = 100 | CC % | CA % | (AA) % | Df | X2 | p Value | |
| Age | >40 | 60 | 20 (33.33%) | 30 (50%) | 10 (16.66%) | 2 | 3.19 | 0.20 |
| <40 | 40 | 14 (35%) | 14 (35%) | 12 (30%) | ||||
| Gender | Males | 78 | 28 (35.89%) | 36 (46.15%) | 14 (17.95%) | 2 | 4.89 | 0.08 |
| Females | 22 | 06 (27.27%) | 08 (36.36%) | 08 (36.36%) | ||||
| Stage | CP-CML | 25 | 5 (20%) | 15 (60%) | 5 (20%) | 2 | 5.43 | 0.04 |
| AP-CML | 20 | 10 (50%) | 5 (25%) | 5 (25%) | ||||
| CP-CML | 25 | 5 (20%) | 15 (60%) | 5 (20%) | 2 | 1.5 | 0.47 | |
| MPD types | BC-CML | 13 | 5 (38.46%) | 6 (46.15%) | 2 (15.38%) | |||
| MPD ET | 15 | 6 (0.4%) | 5 (33.33%) | 4 (26.67%) | 0.64 | 4 | 0.95 | |
| MPD PV | 12 | 5 (41.67%) | 5 (41.67%) | 2 (16.67%) | ||||
| MPD MF | 06 | 3 (0.5%) | 2 (33.33%) | 1 (16.67%) | ||||
| Bax-248G>A genotypes | ||||||||
| N = 108 | GG (56) | GA (45) | AA (07) | Df | X2 | p value | ||
| Age | >40 | 63 | 26 (41.26%) | 35 (55.55%) | 02 (3.17%) | 2 | 12.82 | 0.001 |
| <40 | 45 | 30 (66.66%) | 10 (22.22%) | 05 (11.11%) | ||||
| Gender | Males | 73 | 36 (49.32%) | 33 (45.21%) | 03 (4.11%) | 2 | 2.83 | 0.24 |
| Females | 35 | 20 (57.14%) | 12 (34.29%) | 04 (11.43%) | ||||
| Stage | CP-CML | 30 | 14 (46.67%) | 13 (43.33%) | 03 (1%) | 2 | 0.31 | 0.98 |
| AP-CML | 14 | 06 (42.86%) | 07 (0.5%) | 01 (7.14%) | ||||
| CP-CML | 30 | 14 (46.67%) | 13 (43.33%) | 03 (1%) | 2 | 0.15 | 0.92 | |
| BC-CML | 10 | 04 (40%) | 05 (50%) | 01 (10%) | ||||
| MPD types | MPD ET | 30 | 18 (6%) | 11 (36.67%) | 1 (3.33%) | 4 | 0.80 | 0.93 |
| MPD PV | 16 | 10 (62.5%) | 05 (31.25%) | 1 (6.25%) | ||||
| MPD MF | 04 | 02 (0.5%) | 2 (0.5%) | 0 (0%) | ||||
| TNF-α rs1800629 G>A genotypes | ||||||||
| N = 113 | GG94 | GA10 | AA09 | Df | X2 | p value | ||
| Age | >40 | 76 | 70 (92.10%) | 02 (3.94%) | 04 (5.26%) | 14.49 | 2 | 0.0007 |
| <40 | 37 | 24 (64.86%) | 8 (21.62%) | 05 (13.51%) | ||||
| Gender | Males | 82 | 72 (87.80%) | 08 (9.75%) | 02 (3.65%) | 12.5 | 2 | 0.0019 |
| Females | 31 | 22 (70.96%) | 02 (9.67%) | 07 (19.35%) | ||||
| Stage | CP-CML | 34 | 27 (79.41%) | 04 (11.76%) | 03 (8.82%) | 1.04 | 2 | 0.59 |
| AP-CML | 20 | 18 (90%) | 01 (5%) | 01 (5%) | ||||
| CP-CML | 34 | 27 (79.41%) | 04 (11.76%) | 03 (8.82%) | 7.32 | 2 | 0.025 | |
| BC-CML | 10 | 5 (50%) | 05 (50%) | 00 (0%) | ||||
| MPD types | MPD ET | 30 | 24 (80%) | 3 (10%) | 03 (10%) | 4.56 | 4 | 0.332 |
| MPD PV | 15 | 11 (73.33%) | 3 (20%) | 1 (6.66%) | ||||
| MPD MF | 04 | 2 (50%) | 2 (50%) | 0 (0%) | ||||
| IL-8 rs4073 T>A genotypes | ||||||||
| N = 113 | TT19 | TA41 | AA53 | Df | X2 | p value | ||
| Age | >40 | 76 | 08 (10.52%) | 31 (40.78%) | 37 (48.68%) | 6.91 | 2 | 0.031 |
| <40 | 37 | 11 (29.72%) | 10 (27%) | 16 (43.24%) | ||||
| Gender | Males | 82 | 8 (9.75%) | 33 (40.24%) | 41 (50%) | 10.76 | 2 | 0.004 |
| Females | 31 | 11 (35.48%) | 8 (25.80%) | 12 (38.70%) | ||||
| Stage | CP-CML | 34 | 06 (17.64%) | 12 (35.29%) | 16 (47.05%) | 2 | 0.32 | 0.85 |
| AP-CML | 20 | 03 (15%) | 06 (30%) | 11 (55%) | ||||
| CP-CML | 34 | 06 (17.64%) | 12 (35.29%) | 16 (47.05%) | 2 | 4.32 | 0.11 | |
| BC-CML | 10 | 05 (50%) | 02 (20%) | 03 (30%) | ||||
| MPD types | MPD ET | 30 | 02 (6.66%) | 16 (53.33%) | 12 (40%) | 4.21 | 4 | 0.37 |
| MPD PV | 15 | 02 (13.33%) | 04 (26.66%) | 9 (60%) | ||||
| MPD MF | 04 | 01 (25%) | 01 (25%) | 02 (50%) | ||||
| Association of Bcl-2-938 gene polymorphism | |||||||
| N = 91 | CC(34) | CA (38) | AA (19) | Df | X2 | p value | |
| Ph (+) patients | 58 | 20 | 26 | 12 | 2 | 0.28 | 0.86 |
| Ph (−) patients | 33 | 14 | 12 | 07 | |||
| Association of Bax-248G>A gene polymorphism | |||||||
| N = 94 | GG(44) | GA(43) | AA(07) | Df | X2 | p value | |
| Ph (+) patients | 54 | 24 | 25 | 05 | 2 | 0.72 | 0.69 |
| Ph (−) patients | 40 | 20 | 18 | 02 | |||
| Association of TNF-α rs1800629 G>A polymorphism | |||||||
| N = 113 | GG87 | GA18 | AA08 | Df | X2 | p value | |
| Ph (+) patients | 64 | 50 | 10 | 04 | 2 | 0.18 | 0.91 |
| Ph (−) patients | 49 | 37 | 08 | 04 | |||
| Association of IL-8 rs4073 T>A gene polymorphism | |||||||
| N = 112 | TT19 | TA41 | AA53 | Df | X2 | p value | |
| Ph (+) patients | 63 | 13 | 20 | 30 | 2 | 2.8 | 0.24 |
| Ph (−) patients | 49 | 05 | 21 | 23 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moawadh, M.S.; Mir, R.; Tayeb, F.J.; Asim, O.; Ullah, M.F. Molecular Evaluation of the Impact of Polymorphic Variants in Apoptotic (Bcl-2/Bax) and Proinflammatory Cytokine (TNF-α/IL-8) Genes on the Susceptibility and Progression of Myeloproliferative Neoplasms: A Case-Control Biomarker Study. Curr. Issues Mol. Biol. 2023, 45, 3933-3952. https://doi.org/10.3390/cimb45050251
Moawadh MS, Mir R, Tayeb FJ, Asim O, Ullah MF. Molecular Evaluation of the Impact of Polymorphic Variants in Apoptotic (Bcl-2/Bax) and Proinflammatory Cytokine (TNF-α/IL-8) Genes on the Susceptibility and Progression of Myeloproliferative Neoplasms: A Case-Control Biomarker Study. Current Issues in Molecular Biology. 2023; 45(5):3933-3952. https://doi.org/10.3390/cimb45050251
Chicago/Turabian StyleMoawadh, Mamdoh S., Rashid Mir, Faris J. Tayeb, Orooba Asim, and Mohammad Fahad Ullah. 2023. "Molecular Evaluation of the Impact of Polymorphic Variants in Apoptotic (Bcl-2/Bax) and Proinflammatory Cytokine (TNF-α/IL-8) Genes on the Susceptibility and Progression of Myeloproliferative Neoplasms: A Case-Control Biomarker Study" Current Issues in Molecular Biology 45, no. 5: 3933-3952. https://doi.org/10.3390/cimb45050251