
The Effects of Ionizing Radiation on Gut Microbiota: What Can Animal Models Tell Us?—A Systematic Review

Abstract
:1. Introduction
2. Materials and Methods
2.1. Search Strategy and Selection Criteria
2.2. Inclusion and Exclusion Criteria
2.3. Study Selection and Data Extraction
2.4. Risk of Bias in Individual Studies

{kind=link}
Author, Year / Study Design | Participants / N Irradiated | Microbiome Assessment Method / Type of Sample / Number of Samples | Main Findings |
|---|---|---|---|
| Type of Radiation | |||
| Li Y, 2020 [17] / Interventional | Mice C57BL/6J Male and female 8 weeks N = 5 | 16s rRNA V4 region / Illumina Miseq / Fecal N = 3 Before irradiation, and 6 days and 12 days after irradiation | Diversity/richness
|
| γ-ray TAI Single dose of 12 Gy or 15 Gy TBI Single dose of 4 or 7 Gy | |||
| Yamanouchi K, 2018 [18] / Interventional | Mice C57BL/6Njcl Female 8 weeks N = 6 | DNA Primer PCR / NucleoSpin® DNA Stool / Fecal N = 8 before irradiation, at 1, 2, 6, 12, 24, 48 and 72 h after irradiation | Composition
|
| X-ray TBI Single dose of 2 Gy and 4 Gy | |||
| Goudarzi M, 2016 [19] / Interventional | Mice C57BL/6J Male 8 weeks N = 14 7 control | 16s rRNA V4 region / Illumina HiSeq 2500 QIIME version 1.8.0 Phyloseq packages / Fecal N =3 0 and 5 Gy groups (1 day before and 3 and 30 days post-irradiation) N = 2 12 Gy group (1 day before and 3 days post-irradiation) | Diversity/richness
|
| X-rays TBI Single dose 5 or 12 Gy | |||
| Cui M, 2017 [20] / Interventional | Mice C57BL/6 Male and Female 6-8 weeks / N = 4 | 16s rRNA V4 region / Illumina Hiseq Uparse / Fecal N = 2 Days 5 and 10 postirradiation | Diversity
|
| γ-rays TBI Single dose 6.5 Gy | |||
| Sittipo P, 2020 [21] / Interventional | Mice C57BL/6L Male 8 weeks / N = 10 | 16s rRNA V4-V5 regions / Qubit 2.0 Fluorometer and 2100 Bioanalyzer Ion Torrent PGM platform QIIME v1.9.1 and Microbiome Helper package / Fecal N = 4: -1 day before irradiation -3 (D1, D3 and D10) | Diversity/richness
|
| γ-rays TBI Single dose 6 Gy | |||
| Gerassy-Vainberg S, 2018 [22] / Interventional | Mice C57BL/6J Female 6-8 weeks / N = 23 Control n = 22 | 16S rRNA / QIIME V 1.8.0 Illumina Miseq platform Roche 454 Pyrosequencing / Fecal N = 3 1 week before 2 and 6 weeks post radiation | Composition
|
| γ-rays 4 fractions of 550 cGy Localized internal rectal radiation | |||
| Liu X, 2019 [23] / Interventional | Mice BALB/c / Male 8 weeks / 4 groups: Control; low-dose 6 mice sampled at each timepoint N = 24 | 16s rRNA V4 / Qiagen Mini Kit Qubit 2.0 fluorometer Illumina Hiseq / Fecal N = 4 Before 7, 21 and 35 Postirradiation | Diversity/richness
|
| γ-rays TBI group 1–0.5 Gy ×1 dose group 5–0.1 Gy per dose ×5 doses group 10–0.0 Gy per dose ×10 doses | |||
| Johnson LB, 2004 [24] / Interventional | Mice C57/Bl6 / N = 30 6/per time point controls | Anaerobic vs. Anaerobic / Viable counts / Tissue samples from the irradiated small intestine N = 5 2, 6, 16, 24 and 48 h | Composition
|
| X-rays / Single dose 19 Gy | |||
| Lu L, 2019 [25] / Interventional | Mice C57BL/6 Female 4 to 5 week old / N = 18 6/groups RT only; Control; RT + PC | 16s rRNA V3 and V4 Illumina Hiseq platform / Fecal N = 1 24 h after | Diversity/richness
|
| X-rays TAI Single dose 12 Gy | |||
| Casero D, 2017 [26] / Interventional | Mice C57BL/6 Male 6 months / N = 30 10 controls | 16S rRNA V4 region / Illumina HiSeq 2500 QIIME / Fecal N = 2 after 10 and 30 days | Diversity/richness
|
| TBI 16O (600 MeV/n) at 0.1, 0.25, and 1 Gy | |||
| Kim YS, 2015 [27] / Interventional | Mice C57BL/6 Male 8–10 week old / N = 3 Control = 2 | 16S rRNA UltraClean® Fecal DNA Isolation Kit / Illumina MiSeq // Fecal (small and large intestine) / N = 1 3 days after irradiation. | Composition
|
| γ-rays TBI Single dose 8 Gy | |||
| Wang M, 2020 [28] / Interventional | Mice C57BL/6J Male 8–10 weeks / N = 70 Intestinal group/Survival group Hematopoietic experiments | 16s rRNA V3-V4 / Illumina MiSeq QIIME / Feces removed from the rectum (Small intestines were taken out after three days of irradiation) N = 1 3 days after IR | Diversity/richness
|
| γ-ray TBI 9.0 Gy—intestinal group 10.0 Gy—survival group 4.0 Gy—hematopoietic group | |||
| Zhao Z, 2020 [29] / Interventional | Mice C57BL/6 Male 8–10 weeks / N = 4/3 4 pre radiation 3 post radiation | 16S rRNA V4 region / QIIME (v 1.8) PANDAseq (version 2.9) / Fecal Terminal ileum and cecum 3 months after | Diversity/richness
|
| γ-rays TAI Single dose of 10 Gy | |||
| Wang W, 2020 [30] / Interventional | Mice / N = 18 Controls = 6 | 16s rRNA / Illumina MiSeq qRT-PCR / Fecal samples directly collected from the lower segment of the colon N = 2 Days 7 and 30 after irradiation | Diversity/richness
|
| TBI | |||
| Zhao Y, 2019 [31] / Interventional | Mice C57BL/6J Male 8–12 weeks / N = 5 | 16s rRNA / Illumina Hiseq / Fecal Fresh from rectum 10 month after | Diversity/richness
|
| γ-rays TBI Single dose 8 Gy | |||
| Li Yiyi, 2020 [32] / Interventional | Mice C57BL/6J Male 6- to 8-week-old | 16s rDNA / Fecal N = 2 1 week 6 weeks | Diversity/richness
|
| X-rays Single dose 18 Gy 500 cGy/min for abdominal colorectal localized external radiation | |||
| Raber J, 2020 [33] / Interventional | Mice C57BL/6 F1 4–6 months / N = 99 | 16S rRNA V4 region / Illumina Miseq / Fecal N = 1 2 months post-radiation | Diversity/richness
|
| Protons, 4He, 16O, 28Si, 48Ti and 56Fe ions | |||
| Tong JY, 2022 [34] / Interventional | Mice C57BL/6J Female 3 weeks / N = 24 Controls = 6 | 16S rRNA V4 region / Illumina MiSeq / Fecal | Diversity/richness
|
| X-rays TBI 5 groups: Test; 0.05, 0.10, 0.15 and 0.20 Gy | |||
| Cheema AK, 2021 [35] / Interventional | Mice CD2F1 Male 6/7 weeks / N = 16/group | 16S rRNA V3/V4 region / Illumina MiSeq SILVA / Fecal N = 5 7 and 1 days before irradiation and 3, 14 and 30 post-irradiation | Composition
|
| γ-rays Single dose 9.2 Gy |
| Animals | Author, Year / Interventional | Participants / N Irradiated | Microbiome Assessment Method / Type of Sample / Number of Samples | Main Findings |
|---|---|---|---|---|
| Type of Radiation | ||||
| Rats | Rentea RM, 2016 [36] / Interventional | Rats WAG/RijCmer Male 5 weeks / N = 15 5—Nonirradiated; 5—irradiated; 5—intestinal alkaline phosphatase (RT + IAP) | 16s rRNA / Real-time PCR / Fecal N = 2 D0 and 4 days after irraiation | Composition
|
| X-rays 13 Gy—single dose / Intestinal lower hemibody radiation | ||||
| Lam V, 2012 [37] / Interventional | Rats WAG/RijCmcr (Wistar) Male 5 weeks / N = 10 (n = 5/group) | qPCR and 16S rRNA / Second Genome Inc. G3 PhyloChipe 16S rRNA microarray-based assay / Fecal N = 4 D0 and days 4, 11, and 21 post-irradiation | Composition
| |
| X-rays TBI Single dose 10.0 Gy Multiple-fraction 18.0 Gy | ||||
| Wild rodent: Bank Vole Myodes glareolus | Lavrinienko A, 2018 [38] / Observational | Wild rodent: Bank Vole Myodes glareolus / N = 137 | 16S rRNA V4 / Illumina MiSeq platform at BGI / Fecal | Diversity/richness
|
| 3 study areas of environmental radiation: (1) high (CH) and (2) low (CL and KL) | ||||
| Lavrinienko, 2020 [39] / Observational | Wild rodent: Bank Vole Myodes glareolus / 28 individuals provided fecal (CL1 n = 3, CL2 n = 13; CH1 n = 8, CH2 n = 4). (84–43 Recapture) | 16s rRNA V4 / Illumina MiSeq platform at BGI / Fecal N = 1 | Diversity/richness
Second capture CL:
| |
| Ambient radiation Chernobyl High Radiation (CH) and Chernobyl Low radiation (CL) | ||||
| Göttingen minipigs and Chinese rhesus macaques | Carbonero F, 2018 [40] / Interventional | Göttingen Minipigs and Chinese rhesus Macaques 8 Minipigs 8 Macaques | 16s rRNA / Illumina MiSeq / Fecal N = 2 -2/3 days before -3 days after | Minipigs Diversity/richness
Diversity/richness
|
| 6 MV linear accelerator (LINAC) 80 ± 2.5 Gy/min 1.8 Gy Minipigs 6.8 Gy Macaques | ||||
| Carbonero F, 2018 [41] / Interventional | Göttingen minipigs Chinese rhesus macaques / N = 74 male Chinese rhesus macaques 50 Minipigs | 16s rRNA / Illumina MiSeq QIAGEN / Fecal / Minipigs: collected on days 0 and 3 Macaque fecal samples were collected 24 h before irradiation, between 1–3 h postirradiation and on days 3 and 14 postirradiation | Macaques Diversity/richness
Minipigs Richness
| |
| Macaques 5.9 Gy (n = 12); 6.3 Gy (n = 14); 6.8 Gy (n = 16); 7.2 Gy (n = 16); and 7.7 Gy (n = 16) Minipigs 1.65 Gy (n = 9); 1.80 Gy (n = 10); 1.95 Gy (n = 11); 2.10 Gy (n = 13); and 2.25 Gy (n = 7) | ||||
|
Chinese rhesus macaques,
Macaca mulatta | Kalkeri R, 2021 [42] / Interventional | Chinese rhesus macaques, Macaca Mulatta / N = 19 | Fecal samples / N = 3 1 day prior and 1 and 4 days after exposure | Diversity/richness
|
| Gamma-rays 7.4 Gy | ||||
| Flies | Cai Z, 2018 [43] / Interventional | Flies Males Bactrocera dorsalis 3000 pupae irradiated 15 guts irradiated 15 guts control | 16s rRNA V4 / Illumina MiSeq QIIME v1.8 / Gut / Irradiation 48h before eclosion Day1 Day7 Day14 Post eclosion | Diversity/richness
|
| 100Gy gamma ray Gammacell 220 60Co With an activity of 9435 × 1015 Bq Central dose of 8Gy/min at the beginning of the test | ||||
| Ben Ami, 2020 [44] / Interventional | Flies Vienna 8 Wild C capitata pupae / 150 bacterial colonies from non irradiated 150 colonies from 5-day-old irradiated flies and 100 colonies from field flies | 16s rRNA / PCR-DGGE | Diversity
Non-irradiated vs. irradiated vs. irradiated mass 5 day-read
| |
| Delta irradiation | ||||
| Woruba DN [45] / Interventional | Flies Queensland fruit fly, Bactrocera Tryoni 54 = (3 × 18) | 16S rRNA V3 and V4 regions QIIME / Intact gut dissections / N = 2 1 and 14 days after irradiation | Diversity/richness
| |
| Delta irradiation |
3. Results
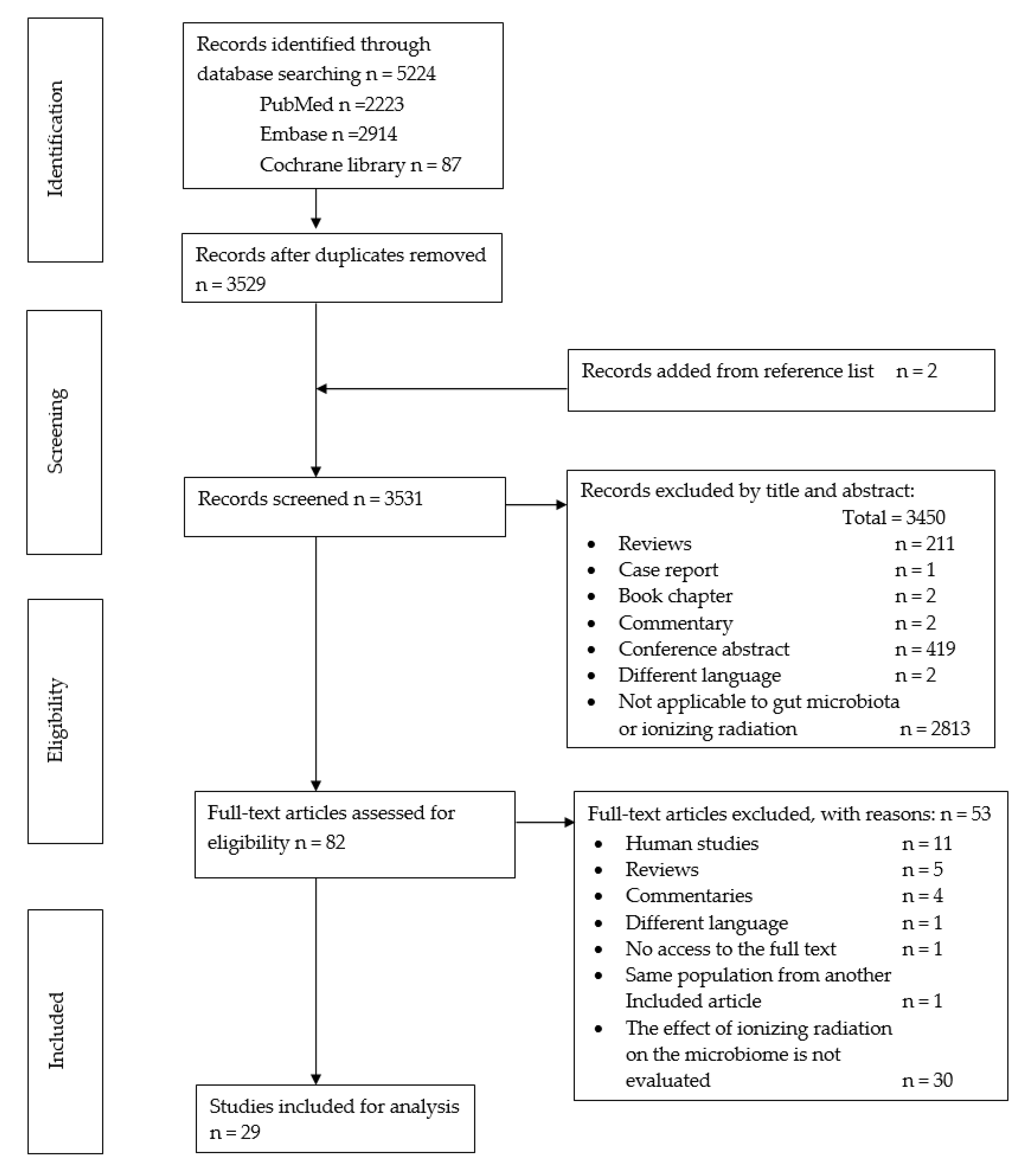
3.1. Search Results
3.2. Study Characteristics
3.2.1. Animal Models
3.2.2. Radiation Exposure Characteristics
3.2.3. Sampling and Microbiota Analysis
3.3. Quality Assessment
3.4. Findings
3.4.1. Diversity and Richness Analysis
3.4.2. Gut Microbial Composition
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kumagai, T.; Rahman, F.; Smith, A.M. The Microbiome and Radiation Induced-Bowel Injury: Evidence for Potential Mechanistic Role in Disease Pathogenesis. Nutrients 2018, 10, 1405. [Google Scholar] [CrossRef] [PubMed]
- Lynch, S.V.; Pedersen, O. The Human Intestinal Microbiome in Health and Disease. N. Engl. J. Med. 2016, 375, 2369–2379. [Google Scholar] [CrossRef] [PubMed]
- Marchesi, J.R.; Adams, D.H.; Fava, F.; Hermes, G.D.; Hirschfield, G.M.; Hold, G.; Quraishi, M.N.; Kinross, J.; Smidt, H.; Tuohy, K.M.; et al. The gut microbiota and host health: A new clinical frontier. Gut 2016, 65, 330–339. [Google Scholar] [CrossRef] [PubMed]
- Nam, Y.D.; Kim, H.J.; Seo, J.G.; Kang, S.W.; Bae, J.W. Impact of pelvic radiotherapy on gut microbiota of gynecological cancer patients revealed by massive pyrosequencing. PLoS ONE 2013, 8, e82659. [Google Scholar] [CrossRef]
- Crawford, P.A.; Gordon, J.I. Microbial regulation of intestinal radiosensitivity. Proc. Natl. Acad. Sci. USA 2005, 102, 13254–13259. [Google Scholar] [CrossRef]
- Wang, A.; Ling, Z.; Yang, Z.; Kiela, P.R.; Wang, T.; Wang, C.; Cao, L.; Geng, F.; Shen, M.; Ran, X.; et al. Gut microbial dysbiosis may predict diarrhea and fatigue in patients undergoing pelvic cancer radiotherapy: A pilot study. PLoS ONE 2015, 10, e0126312. [Google Scholar] [CrossRef]
- Desouky, O.; Ding, N.; Zhou, G. Targeted and non-targeted effects of ionizing radiation. J. Radiat. Res. Appl. Sci. 2015, 8, 247–254. [Google Scholar] [CrossRef]
- Molla, M.; Panes, J. Radiation-induced intestinal inflammation. World J. Gastroenterol. 2007, 13, 3043–3046. [Google Scholar] [CrossRef]
- Leibowitz, B.J.; Wei, L.; Zhang, L.; Ping, X.; Epperly, M.; Greenberger, J.; Cheng, T.; Yu, J. Ionizing irradiation induces acute haematopoietic syndrome and gastrointestinal syndrome independently in mice. Nat. Commun. 2014, 5, 3494. [Google Scholar] [CrossRef]
- Booth, C.; Tudor, G.; Tudor, J.; Katz, B.P.; MacVittie, T.J. Acute gastrointestinal syndrome in high-dose irradiated mice. Health Phys. 2012, 103, 383–399. [Google Scholar] [CrossRef]
- Andreyev, J. Gastrointestinal complications of pelvic radiotherapy: Are they of any importance? Gut 2005, 54, 1051–1054. [Google Scholar] [CrossRef]
- Hauer-Jensen, M.; Wang, J.; Boerma, M.; Fu, Q.; Denham, J.W. Radiation damage to the gastrointestinal tract: Mechanisms, diagnosis, and management. Curr. Opin. Support. Palliat. Care 2007, 1, 23–29. [Google Scholar] [CrossRef]
- Nguyen, T.L.; Vieira-Silva, S.; Liston, A.; Raes, J. How informative is the mouse for human gut microbiota research? Dis. Model. Mech. 2015, 8, 1–16. [Google Scholar] [CrossRef]
- Fernandes, A.; Oliveira, A.; Soares, R.; Barata, P. The Effects of Ionizing Radiation on Gut Microbiota, a Systematic Review. Nutrients 2021, 13, 3025. [Google Scholar] [CrossRef]
- Higgins, J.; Thomas, J.; Chandler, J.; Cumpston, M.; Li, T.; Page, M.J.; Welch, V.A. Cochrane Handbook for Systematic Reviews of Interventions version 6.2 (updated February 2021). Cochrane. 2021. Available online: www.training.cochrane.org/handbook (accessed on 30 October 2022).
- Hooijmans, C.R.; Rovers, M.M.; de Vries, R.B.M.; Leenaars, M.; Ritskes-Hoitinga, M.; Langendam, M.W. SYRCLE’s risk of bias tool for animal studies. BMC Med. Res. Methodol. 2014, 14, 43. [Google Scholar] [CrossRef]
- Li, Y.; Dong, J.; Xiao, H.; Zhang, S.; Wang, B.; Cui, M.; Fan, S. Gut commensal derived-valeric acid protects against radiation injuries. Gut Microbes 2020, 11, 789–806. [Google Scholar] [CrossRef]
- Yamanouchi, K.; Tsujiguchi, T.; Sakamoto, Y.; Ito, K. Short-term follow-up of intestinal flora in radiation-exposed mice. J. Radiat. Res. 2019, 60, 328–332. [Google Scholar] [CrossRef]
- Goudarzi, M.; Mak, T.D.; Jacobs, J.P.; Moon, B.H.; Strawn, S.J.; Braun, J.; Brenner, D.J.; Fornace, A.J., Jr.; Li, H.H. An Integrated Multi-Omic Approach to Assess Radiation Injury on the Host-Microbiome Axis. Radiat. Res. 2016, 186, 219–234. [Google Scholar] [CrossRef]
- Cui, M.; Xiao, H.; Li, Y.; Zhou, L.; Zhao, S.; Luo, D.; Zheng, Q.; Dong, J.; Zhao, Y.; Zhang, X.; et al. Faecal microbiota transplantation protects against radiation-induced toxicity. EMBO Mol. Med. 2017, 9, 448–461. [Google Scholar] [CrossRef]
- Sittipo, P.; Pham, H.Q.; Park, C.E.; Kang, G.U.; Zhi, Y.; Ji, H.J.; Jang, A.; Seo, H.S.; Shin, J.H.; Lee, Y.K. Irradiation-Induced Intestinal Damage Is Recovered by the Indigenous Gut Bacteria Lactobacillus acidophilus. Front. Cell Infect. Microbiol. 2020, 10, 415. [Google Scholar] [CrossRef]
- Gerassy-Vainberg, S.; Blatt, A.; Danin-Poleg, Y.; Gershovich, K.; Sabo, E.; Nevelsky, A.; Daniel, S.; Dahan, A.; Ziv, O.; Dheer, R.; et al. Radiation induces proinflammatory dysbiosis: Transmission of inflammatory susceptibility by host cytokine induction. Gut 2018, 67, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhou, Y.; Wang, S.; Guan, H.; Hu, S.; Huang, R.; Zhou, P. Impact of Low-dose Ionising Radiation on the Composition of the Gut Microbiota of Mice. Toxicol. Sci. 2019, 171, 258–268. [Google Scholar] [CrossRef] [PubMed]
- Johnson, L.B.; Riaz, A.A.; Adawi, D.; Wittgren, L.; Back, S.; Thornberg, C.; Osman, N.; Gadaleanu, V.; Thorlacius, H.; Jeppsson, B. Radiation enteropathy and leucocyte-endothelial cell reactions in a refined small bowel model. BMC Surg. 2004, 4, 10. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Li, W.; Sun, C.; Kang, S.; Li, J.; Luo, X.; Su, Q.; Liu, B.; Qin, S. Phycocyanin Ameliorates Radiation-Induced Acute Intestinal Toxicity by Regulating the Effect of the Gut Microbiota on the TLR4/Myd88/NF-κB Pathway. JPEN J. Parenter Enteral. Nutr. 2020, 44, 1308–1317. [Google Scholar] [CrossRef]
- Casero, D.; Gill, K.; Sridharan, V.; Koturbash, I.; Nelson, G.; Hauer-Jensen, M.; Boerma, M.; Braun, J.; Cheema, A.K. Space-type radiation induces multimodal responses in the mouse gut microbiome and metabolome. Microbiome 2017, 5, 105. [Google Scholar] [CrossRef]
- Kim, Y.S.; Kim, J.; Park, S.J. High-throughput 16S rRNA gene sequencing reveals alterations of mouse intestinal microbiota after radiotherapy. Anaerobe 2015, 33, 1–7. [Google Scholar] [CrossRef]
- Wang, M.; Dong, Y.; Wu, J.; Li, H.; Zhang, Y.; Fan, S.; Li, D. Baicalein ameliorates ionizing radiation-induced injuries by rebalancing gut microbiota and inhibiting apoptosis. Life Sci. 2020, 261, 118463. [Google Scholar] [CrossRef]
- Zhao, Z.; Cheng, W.; Qu, W.; Shao, G.; Liu, S. Antibiotic Alleviates Radiation-Induced Intestinal Injury by Remodeling Microbiota, Reducing Inflammation, and Inhibiting Fibrosis. ACS Omega 2020, 5, 2967–2977. [Google Scholar] [CrossRef]
- Wang, W.; Hu, L.; Chang, S.; Ma, L.; Li, X.; Yang, Z.; Du, C.; Qu, X.; Zhang, C.; Wang, S. Total body irradiation-induced colon damage is prevented by nitrate-mediated suppression of oxidative stress and homeostasis of the gut microbiome. Nitric Oxide 2020, 102, 1–11. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhang, J.; Han, X.; Fan, S. Total body irradiation induced mouse small intestine senescence as a late effect. J. Radiat. Res. 2019, 60, 442–450. [Google Scholar] [CrossRef]
- Li, Y.; Yan, H.; Zhang, Y.; Li, Q.; Yu, L.; Li, Q.; Liu, C.; Xie, Y.; Chen, K.; Ye, F.; et al. Alterations of the Gut Microbiome Composition and Lipid Metabolic Profile in Radiation Enteritis. Front. Cell Infect. Microbiol. 2020, 10, 541178. [Google Scholar] [CrossRef]
- Raber, J.; Fuentes Anaya, A.; Torres, E.R.S.; Lee, J.; Boutros, S.; Grygoryev, D.; Hammer, A.; Kasschau, K.D.; Sharpton, T.J.; Turker, M.S.; et al. Effects of Six Sequential Charged Particle Beams on Behavioral and Cognitive Performance in B6D2F1 Female and Male Mice. Front. Physiol. 2020, 11, 959. [Google Scholar] [CrossRef] [PubMed]
- Tong, J.Y.; Jiang, W.; Yu, X.Q.; Wang, R.; Lu, G.H.; Gao, D.W.; Lv, Z.W.; Li, D. Effect of low-dose radiation on thyroid function and the gut microbiota. World J. Gastroenterol. 2022, 28, 5557–5572. [Google Scholar] [CrossRef] [PubMed]
- Cheema, A.K.; Li, Y.; Singh, J.; Johnson, R.; Girgis, M.; Wise, S.Y.; Fatanmi, O.O.; Kaytor, M.D.; Singh, V.K. Microbiome study in irradiated mice treated with BIO 300, a promising radiation countermeasure. Anim. Microbiome 2021, 3, 71. [Google Scholar] [CrossRef]
- Rentea, R.M.; Lam, V.; Biesterveld, B.; Fredrich, K.M.; Callison, J.; Fish, B.L.; Baker, J.E.; Komorowski, R.; Gourlay, D.M.; Otterson, M.F. Radiation-induced changes in intestinal and tissue-nonspecific alkaline phosphatase: Implications for recovery after radiation therapy. Am. J. Surg. 2016, 212, 602–608. [Google Scholar] [CrossRef] [PubMed]
- Lam, V.; Moulder, J.E.; Salzman, N.H.; Dubinsky, E.A.; Andersen, G.L.; Baker, J.E. Intestinal microbiota as novel biomarkers of prior radiation exposure. Radiat. Res. 2012, 177, 573–583. [Google Scholar] [CrossRef]
- Lavrinienko, A.; Mappes, T.; Tukalenko, E.; Mousseau, T.A.; Møller, A.P.; Knight, R.; Morton, J.T.; Thompson, L.R.; Watts, P.C. Environmental radiation alters the gut microbiome of the bank vole Myodes glareolus. ISME J. 2018, 12, 2801–2806. [Google Scholar] [CrossRef]
- Lavrinienko, A.; Tukalenko, E.; Kesäniemi, J.; Kivisaari, K.; Masiuk, S.; Boratyński, Z.; Mousseau, T.A.; Milinevsky, G.; Mappes, T.; Watts, P.C. Applying the Anna Karenina principle for wild animal gut microbiota: Temporal stability of the bank vole gut microbiota in a disturbed environment. J. Anim. Ecol. 2020, 89, 2617–2630. [Google Scholar] [CrossRef]
- Carbonero, F.; Mayta-Apaza, A.C.; Yu, J.Z.; Lindeblad, M.; Lyubimov, A.; Neri, F.; Szilagyi, E.; Bartholomew, A. A comparative analysis of gut microbiota disturbances in the Gottingen minipig and rhesus macaque models of acute radiation syndrome following bioequivalent radiation exposures. Radiat. Environ. Biophys. 2018, 57, 419–426. [Google Scholar] [CrossRef]
- Carbonero, F.; Mayta, A.; Bolea, M.; Yu, J.Z.; Lindeblad, M.; Lyubimov, A.; Neri, F.; Szilagyi, E.; Smith, B.; Halliday, L.; et al. Specific Members of the Gut Microbiota are Reliable Biomarkers of Irradiation Intensity and Lethality in Large Animal Models of Human Health. Radiat. Res. 2019, 191, 107–121. [Google Scholar] [CrossRef]
- Kalkeri, R.; Walters, K.; Van Der Pol, W.; McFarland, B.C.; Fisher, N.; Koide, F.; Morrow, C.D.; Singh, V.K. Changes in the gut microbiome community of nonhuman primates following radiation injury. BMC Microbiol. 2021, 21, 93. [Google Scholar] [CrossRef]
- Cai, Z.; Yao, Z.; Li, Y.; Xi, Z.; Bourtzis, K.; Zhao, Z.; Bai, S.; Zhang, H. Intestinal probiotics restore the ecological fitness decline of Bactrocera dorsalis by irradiation. Evol. Appl. 2018, 11, 1946–1963. [Google Scholar] [CrossRef]
- Ben Ami, E.; Yuval, B.; Jurkevitch, E. Manipulation of the microbiota of mass-reared Mediterranean fruit flies Ceratitis capitata (Diptera: Tephritidae) improves sterile male sexual performance. ISME J. 2010, 4, 28–37. [Google Scholar] [CrossRef]
- Woruba, D.N.; Morrow, J.L.; Reynolds, O.L.; Chapman, T.A.; Collins, D.P.; Riegler, M. Diet and irradiation effects on the bacterial community composition and structure in the gut of domesticated teneral and mature Queensland fruit fly, Bactrocera tryoni (Diptera: Tephritidae). BMC Microbiol. 2019, 19, 281. [Google Scholar] [CrossRef]
- Bryda, E.C. The Mighty Mouse: The impact of rodents on advances in biomedical research. Mo. Med. 2013, 110, 207–211. [Google Scholar]
- Bassols, A.; Costa, C.; Eckersall, P.D.; Osada, J.; Sabrià, J.; Tibau, J. The pig as an animal model for human pathologies: A proteomics perspective. Proteom. Clin. Appl. 2014, 8, 715–731. [Google Scholar] [CrossRef]
- Singh, V.K.; Olabisi, A.O. Nonhuman primates as models for the discovery and development of radiation countermeasures. Expert Opin. Drug Discov. 2017, 12, 695–709. [Google Scholar] [CrossRef]
- Raber, J.; Yamazaki, J.; Torres, E.R.S.; Kirchoff, N.; Stagaman, K.; Sharpton, T.; Turker, M.S.; Kronenberg, A. Combined Effects of Three High-Energy Charged Particle Beams Important for Space Flight on Brain, Behavioral and Cognitive Endpoints in B6D2F1 Female and Male Mice. Front. Physiol. 2019, 10, 179. [Google Scholar] [CrossRef]
- Claesson, M.J.; Clooney, A.G.; O’Toole, P.W. A clinician’s guide to microbiome analysis. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 585–595. [Google Scholar] [CrossRef]
- Touchefeu, Y.; Montassier, E.; Nieman, K.; Gastinne, T.; Potel, G.; Bruley des Varannes, S.; Le Vacon, F.; de La Cochetière, M.F. Systematic review: The role of the gut microbiota in chemotherapy- or radiation-induced gastrointestinal mucositis—Current evidence and potential clinical applications. Aliment. Pharmacol. Ther. 2014, 40, 409–421. [Google Scholar] [CrossRef]
- Ley, R.E.; Bäckhed, F.; Turnbaugh, P.; Lozupone, C.A.; Knight, R.D.; Gordon, J.I. Obesity alters gut microbial ecology. Proc. Natl. Acad. Sci. USA 2005, 102, 11070–11075. [Google Scholar] [CrossRef] [PubMed]
- Hugenholtz, F.; de Vos, W.M. Mouse models for human intestinal microbiota research: A critical evaluation. Cell Mol. Life Sci. 2018, 75, 149–160. [Google Scholar] [CrossRef] [PubMed]
- Imaoka, A.; Setoyama, H.; Takagi, A.; Matsumoto, S.; Umesaki, Y. Improvement of human faecal flora-associated mouse model for evaluation of the functional foods. J. Appl. Microbiol. 2004, 96, 656–663. [Google Scholar] [CrossRef] [PubMed]
- Mosca, A.; Leclerc, M.; Hugot, J.P. Gut Microbiota Diversity and Human Diseases: Should We Reintroduce Key Predators in Our Ecosystem? Front. Microbiol. 2016, 7, 455. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.T.; Amos, G.C.A.; Murphy, A.R.J.; Murch, S.; Wellington, E.M.H.; Arasaradnam, R.P. Microbial imbalance in inflammatory bowel disease patients at different taxonomic levels. Gut Pathog. 2020, 12, 1. [Google Scholar] [CrossRef] [PubMed]
- Manichanh, C.; Rigottier-Gois, L.; Bonnaud, E.; Gloux, K.; Pelletier, E.; Frangeul, L.; Nalin, R.; Jarrin, C.; Chardon, P.; Marteau, P.; et al. Reduced diversity of faecal microbiota in Crohn’s disease revealed by a metagenomic approach. Gut 2006, 55, 205–211. [Google Scholar] [CrossRef]
- Han, H.; Li, Y.; Fang, J.; Liu, G.; Yin, J.; Li, T.; Yin, Y. Gut Microbiota and Type 1 Diabetes. Int. J. Mol. Sci. 2018, 19, 995. [Google Scholar] [CrossRef]
- Lee, C.J.; Sears, C.L.; Maruthur, N. Gut microbiome and its role in obesity and insulin resistance. Ann. N. Y. Acad. Sci. 2020, 1461, 37–52. [Google Scholar] [CrossRef]
- Chu, M.; Zhang, X. Bacterial Atlas of Mouse Gut Microbiota. Cell. Microbiol. 2022, 2022, 5968814. [Google Scholar] [CrossRef]
- Magne, F.; Gotteland, M.; Gauthier, L.; Zazueta, A.; Pesoa, S.; Navarrete, P.; Balamurugan, R. The Firmicutes/Bacteroidetes Ratio: A Relevant Marker of Gut Dysbiosis in Obese Patients? Nutrients 2020, 12, 1474. [Google Scholar] [CrossRef]
- Tseng, C.H.; Wu, C.Y. The gut microbiome in obesity. J. Formos. Med. Assoc. 2019, 118 (Suppl. S1), S3–S9. [Google Scholar] [CrossRef]
- Zhang, T.; Li, Q.; Cheng, L.; Buch, H.; Zhang, F. Akkermansia muciniphila is a promising probiotic. Microb. Biotechnol. 2019, 12, 1109–1125. [Google Scholar] [CrossRef]
- Derrien, M.; Belzer, C.; de Vos, W.M. Akkermansia muciniphila and its role in regulating host functions. Microb. Pathog. 2017, 106, 171–181. [Google Scholar] [CrossRef]
- Wang, L.; Christophersen, C.T.; Sorich, M.J.; Gerber, J.P.; Angley, M.T.; Conlon, M.A. Low relative abundances of the mucolytic bacterium Akkermansia muciniphila and Bifidobacterium spp. in feces of children with autism. Appl. Environ. Microbiol. 2011, 77, 6718–6721. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, Q.; Wang, X.; Zhu, L.; Chen, J.; Zhang, B.; Chen, Y.; Yuan, Z. Gut microbial dysbiosis is associated with development and progression of radiation enteritis during pelvic radiotherapy. J. Cell Mol. Med. 2019, 23, 3747–3756. [Google Scholar] [CrossRef]
- Sheikh Sajjadieh, M.R.; Kuznetsova, L.V.; Bojenko, V.B. Dysbiosis in ukrainian children with irritable bowel syndrome affected by natural radiation. Iran J. Pediatr. 2012, 22, 364–368. [Google Scholar]
- García-Peris, P.; Velasco, C.; Lozano, M.A.; Moreno, Y.; Paron, L.; de la Cuerda, C.; Bretón, I.; Camblor, M.; García-Hernández, J.; Guarner, F.; et al. Effect of a mixture of inulin and fructo-oligosaccharide on Lactobacillus and Bifidobacterium intestinal microbiota of patients receiving radiotherapy: A randomised, double-blind, placebo-controlled trial. Nutr. Hosp. 2012, 27, 1908–1915. [Google Scholar] [CrossRef]
- Rosli, D.; Shahar, S.; Manaf, Z.A.; Lau, H.J.; Yusof, N.Y.M.; Haron, M.R.; Majid, H.A. Randomized controlled trial on the effect of partially hydrolyzed guar gum supplementation on diarrhea frequency and gut microbiome count among pelvic radiation patients. JPEN J Parenter Enteral Nutr. 2021;45(2):277–286. J. Parenter. Enter. Nutr. 2021, 46, 475. [Google Scholar] [CrossRef]
- Parker, B.J.; Wearsch, P.A.; Veloo, A.C.M.; Rodriguez-Palacios, A. The Genus Alistipes: Gut Bacteria with Emerging Implications to Inflammation, Cancer, and Mental Health. Front. Immunol. 2020, 11, 906. [Google Scholar] [CrossRef]
- Dziarski, R.; Park, S.Y.; Kashyap, D.R.; Dowd, S.E.; Gupta, D. Pglyrp-Regulated Gut Microflora Prevotella falsenii, Parabacteroides distasonis and Bacteroides eggerthii Enhance and Alistipes finegoldii Attenuates Colitis in Mice. PLoS ONE 2016, 11, e0146162. [Google Scholar] [CrossRef]
- Feng, Q.; Liang, S.; Jia, H.; Stadlmayr, A.; Tang, L.; Lan, Z.; Zhang, D.; Xia, H.; Xu, X.; Jie, Z.; et al. Gut microbiome development along the colorectal adenoma-carcinoma sequence. Nat. Commun. 2015, 6, 6528. [Google Scholar] [CrossRef] [PubMed]
- Villa, J.K.; Han, R.; Tsai, C.-H.; Chen, A.; Sweet, P.; Franco, G.; Vaezian, R.; Tkavc, R.; Daly, M.J.; Contreras, L.M. A small RNA regulates pprM, a modulator of pleiotropic proteins promoting DNA repair, in Deinococcus radiodurans under ionizing radiation. Sci. Rep. 2021, 11, 12949. [Google Scholar] [CrossRef] [PubMed]
- Šiková, M.; Janoušková, M.; Ramaniuk, O.; Páleníková, P.; Pospíšil, J.; Bartl, P.; Suder, A.; Pajer, P.; Kubičková, P.; Pavliš, O.; et al. Ms1 RNA increases the amount of RNA polymerase in Mycobacterium smegmatis. Mol. Microbiol. 2019, 111, 354–372. [Google Scholar] [CrossRef]
- Winter, S.E.; Winter, M.G.; Xavier, M.N.; Thiennimitr, P.; Poon, V.; Keestra, A.M.; Laughlin, R.C.; Gomez, G.; Wu, J.; Lawhon, S.D.; et al. Host-derived nitrate boosts growth of E. coli in the inflamed gut. Science 2014, 339, 708–711. [Google Scholar] [CrossRef] [PubMed]
- Morgan, X.C.; Tickle, T.; Sokol, H.; Gevers, D.; Devaney, K.L.; Ward, D.V.; A Reyes, J.; A Shah, S.; Leleiko, N.; Snapper, S.B.; et al. Dysfunction of the intestinal microbiome in inflammatory bowel disease and treatment. Genome Biol. 2012, 13, R79. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Shin, Y.-C.; Kim, T.-Y.; Kim, Y.; Lee, Y.-S.; Lee, S.-H.; Kim, M.-N.; O, E.; Kim, K.S.; Kweon, M.-N. Mucin degrader Akkermansia muciniphila accelerates intestinal stem cell-mediated epithelial development. Gut Microbes 2021, 13, 1–20. [Google Scholar] [CrossRef]
- Sankar, S.A.; Lagier, J.C.; Pontarotti, P.; Raoult, D.; Fournier, P.E. The human gut microbiome, a taxonomic conundrum. Syst. Appl. Microbiol. 2015, 38, 276–286. [Google Scholar] [CrossRef]

| Database | Search Number | Search Terms |
|---|---|---|
| PubMed | Search #1 | “microbiota” OR “gastrointestinal microbiome” OR “microbiome” OR “16s rRNA” |
| Search #2 | “radiation” OR “radiotherapy” | |
| Search #3 | Search #1 AND Search #2 | |
| EMBASE (via OVID) | Search #1 | “microbiota” OR “gastrointestinal microbiome” OR “microbiome” OR “16s rRNA” OR “microflora” |
| Search #2 | “radiation” OR “radiotherapy” | |
| Search #3 | English OR Spanish OR Portuguese | |
| Search #4 | Search #1 AND Search #2 AND Search #3 | |
| Cochrane Library | Search #1 | “microbiota” OR “gastrointestinal microbiome” OR “microbiome” OR “16s rRNA” OR “microflora” |
| Search #2 | “radiation” OR “radiotherapy” | |
| Search #3 | Search #1 AND Search #2 |
| Key Findings from the Studies | |
|---|---|
| Diversity | |
| |
| ACE index | |
| α diversity | |
| |
| Shannon index |
|
| Simpson diversity index | |
| Chao1 index |
|
| Beta diversity | |
| Richness | |
| Number of OTUs/Taxa number |
|
| Altered composition/Dysbiosis | |
| |
| Anaerobic counts | |
| |
| Aerobic counts | |
| |
| Phylum | |
| Ratio Firmicutes/Bacteroidetes | |
| Actinobacteria | |
| Bacteroidetes |
|
| Epsilonbacteraeota |
|
| Firmicutes |
|
| Proteobacteria |
|
| Verrucomicrobia |
|
| Spirochaetes |
|
| Class | |
| Clostridia | |
| Bacteroida | |
| Betaproteobacteria |
|
| Unidentified_Saccharibacteria |
|
| Epsilonproteobacteria |
|
| Deltaproteobacteria |
|
| Erysipelotrichia |
|
| Order | |
| Clostridiales | |
| Bifidobacteriales |
|
| Coriobacteriales |
|
| Verrucomicrobiales |
|
| Lactobacillales |
|
| Bacteroidales |
|
| Family | |
| Desulfovibrionaceae | |
| Staphylococcaceae |
|
| Lactobacillacea | |
| Prevotellacea |
|
| Clostridiaceae | |
| Lachnospiracea |
|
| Moraxellaceae |
|
| Ruminococcaceae | |
| Porphyromonadaceae |
|
| Rikenellaceae |
|
| Eggerthellaceae |
|
| Enterobacteriaceae | |
| Flavobacteriaceae |
|
| Muribaculaceae S24-7 family |
|
| Bacillaceae |
|
| Xanthomonadaceae |
|
| Sphingobacteriaceae |
|
| Aeromonadacea |
|
| Peptostreptococcaceae |
|
| Veillonellaceae |
|
| Genus | |
| Acinetobacter |
|
| Aerococcus |
|
| Actinobacillus |
|
| Actinobacteria major genera |
|
| Akkermansia | |
| Alloprevotella | |
| Alistipes |
|
| Anaerotruncus |
|
| Bacteroides |
|
| Barnesiella | |
| Betaproteobacteria members (Desulfovibrio and Bilophila) |
|
| Bacillus spp. | |
| Bifidobacterium | |
| Butyricimonas |
|
| Blautia | |
| Citrobacter sp. |
|
| Collinsella |
|
| Coprococcus_1 |
|
| Corynebacterium |
|
| Clostridium | |
| Clostridium cluster IV, XIVa and XIVb | |
| Dubosiella |
|
| Elusimicrobium |
|
| Enterobacter sp. |
|
| Enterococcus |
|
| Escherichia-Shigella | |
| Eubacterium_coprostanoligenes_group |
|
| Faecalibacterium |
|
| Helicobacter | |
| Klebsiella sp. | Decreased (Ben Ami, 2020) [44] |
| Lactobacillus |
|
| Mucispirilum |
|
| Olsenella |
|
| Oscillibacter | |
| Parabacteroides | |
| Paraprevotella | |
| Pseudomonas sp. |
|
| Pseudoflavonifractor |
|
| Prevotella | |
| Providencia sp. |
|
| Quinella |
|
| Ralstonia sp. |
|
| Roseburia | |
| Ruminococcus | |
| Slackia |
|
| Streptococcus | |
| Suterella spp. |
|
| Treponema | |
| Veillonella |
|
| Species | |
| Adlercreutzia unclassified |
|
| Akkermansia muciniphila |
|
| Clostridiaceae species |
|
| Eubacterium biforme |
|
| Mollicutes species (Tenericutes phylum) |
|
| Prevotellaceae_UCG-001 | |
| Ruminococcaceae_UCG-014 | |
| Ruminococcus gnavus | |
| S24–7 unclassified species |
|
| Unclassified Lactobacillus |
|
| uncultured_bacterium_g_Acinetobacter, |
|
| uncultured_bacterium_o_, Mollicutes_RF39, |
|
| uncultured_bacterium_g_Citrobacter, |
|
| uncultured_bacterium_g_Lactococcus—decreased |
|
| Streptococcus_gallolyticus |
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernandes, A.; Oliveira, A.; Soares, R.; Barata, P. The Effects of Ionizing Radiation on Gut Microbiota: What Can Animal Models Tell Us?—A Systematic Review. Curr. Issues Mol. Biol. 2023, 45, 3877-3910. https://doi.org/10.3390/cimb45050249
Fernandes A, Oliveira A, Soares R, Barata P. The Effects of Ionizing Radiation on Gut Microbiota: What Can Animal Models Tell Us?—A Systematic Review. Current Issues in Molecular Biology. 2023; 45(5):3877-3910. https://doi.org/10.3390/cimb45050249
Chicago/Turabian StyleFernandes, Ana, Ana Oliveira, Raquel Soares, and Pedro Barata. 2023. "The Effects of Ionizing Radiation on Gut Microbiota: What Can Animal Models Tell Us?—A Systematic Review" Current Issues in Molecular Biology 45, no. 5: 3877-3910. https://doi.org/10.3390/cimb45050249