
A Comparative Study on the Carcass and Meat Chemical Composition, and Lipid-Metabolism-Related Gene Expression in Korean Hanwoo and Brindle Chikso Cattle
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Carcass Traits and Meat Yield
2.3. Chemical Composition
2.4. Fatty Acid Composition
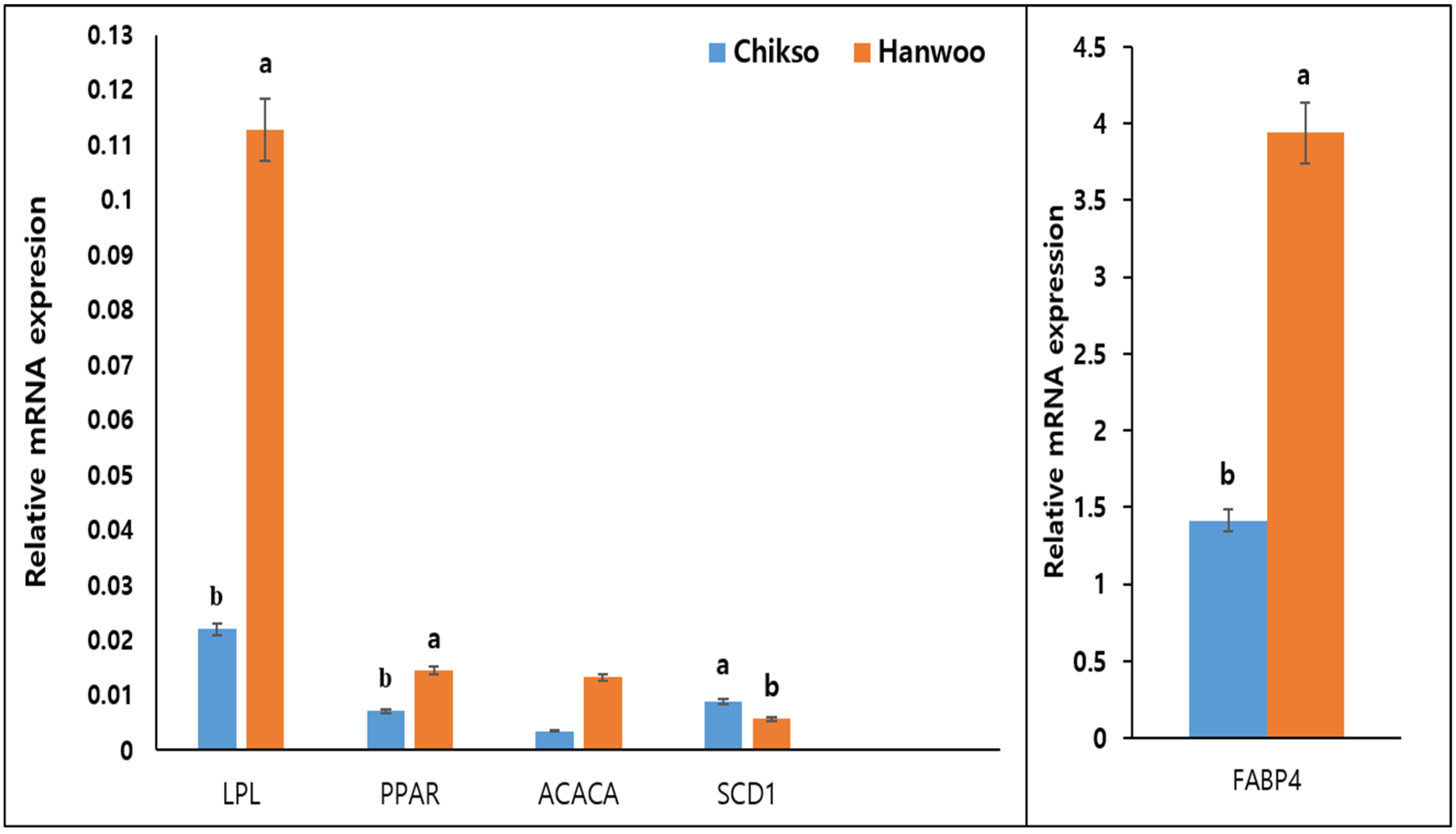
2.5. Adipogenesis-Related Genes Expression
2.6. Statistical Analysis
3. Results and Discussion
3.1. Carcass Traits and Composition
3.2. Meat Yield
3.3. Proximate Composition
3.4. Fatty Acid Profiles
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chung, K.Y.; Lee, S.H.; Cho, S.H.; Kwon, E.G.; Lee, J.H. Current situation and future prospects for beef production in South Korea—A review. Anim. Biosci. 2018, 31, 951–960. [Google Scholar] [CrossRef] [PubMed]
- Gotoh, T.; Nishimura, T.; Kuchida, K.; Mannen, H. The Japanese Wagyu beef industry: Current situation and future prospects—A review. Asian-Australas. J. Anim. Sci. 2018, 31, 933–950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.J.; Koh, Y.J.; Kim, Y.K.; Lee, S.H.; Lee, J.H.; Seo, D.W. MSENet: Marbling score estimation network for automated assessment of Korean beef. Meat Sci. 2022, 188, 108785. [Google Scholar] [CrossRef]
- Hunt, M.; Garmyn, A.; O'Quinn, T.; Corbin, C.; Legako, J.; Rathmann, R.; Brooks, J.; Miller, M. Consumer assessment of beef palatability from four beef muscles from USDA Choice and Select graded carcasses. Meat Sci. 2014, 98, 1–8. [Google Scholar] [CrossRef]
- Schumacher, M.; DelCurto-Wyffels, H.; Thomson, J.; Boles, J. Fat deposition and fat effects on meat quality—A Review. Animals 2022, 12, 1550. [Google Scholar] [CrossRef] [PubMed]
- Joo, S.T.; Hwang, Y.H.; Frank, D. Characteristics of Hanwoo cattle and health implications of consuming highly marbled Hanwoo beef. A review. Meat Sci. 2017, 132, 45–51. [Google Scholar] [CrossRef]
- Vargas-Bello-Pérez, E.; Larraín, R.E. Impacts of fat from ruminants’ meat on cardiovascular health and possible strategies to alter its lipid composition. J. Sci. Food Agric. 2017, 97, 1969–1978. [Google Scholar] [CrossRef]
- Dinh, T.T.N.; To, K.V.; Schilling, M.W. Fatty acid composition of meat animals as flavor precursors. Meat Muscle Biol. 2021, 5, 1–16. [Google Scholar] [CrossRef]
- Listrat, A.; Gagaoua, M.; Andueza, D.; Gruffat, D.; Normand, J.; Mairessec, G.; Picard, B.; Hocquettea, J.F. What are the drivers of beef sensory quality using metadata of intramuscular connective tissue, fatty acids and muscle fiber characteristics? Livest. Sci. 2020, 240, 104209. [Google Scholar] [CrossRef]
- Hwang, Y.H.; Joo, S.T. Fatty acid profiles, meat quality, and sensory palatability of grain-fed and grass-fed beef from Hanwoo, American, and Australian crossbred cattle. Korean J. Food Sci. Anim. Resour. 2017, 37, 153–161. [Google Scholar] [CrossRef] [Green Version]
- Vázquez-Mosquera, J.M.; Fernandez-Novo, A.; de Mercado, E.; Vázquez-Gómez, M.; Gardon, J.C.; Pesántez-Pacheco, J.L.; Revilla-Ruiz, Á.; Patrón-Collantes, R.; Pérez-Solana, M.L.; Villagrá, A.; et al. Beef nutritional characteristics, fat profile and blood metabolic markers from purebred Wagyu, crossbred Wagyu and crossbred European steers raised on a fattening farm in Spain. Animals 2023, 13, 864. [Google Scholar] [CrossRef]
- Liu, L.; Cao, P.; Zhang, L.; Qi, M.; Wang, L.; Li, Z.; Shao, G.; Ding, L.; Zhao, X.; Zhao, X.; et al. Comparisons of adipogenesis- and lipid metabolism-related gene expression levels in muscle, adipose tissue and liver from Wagyucross and Holstein steers. PLoS ONE 2021, 16, e0247559. [Google Scholar] [CrossRef]
- Baik, M.; Vu, T.T.T.; Piao, M.Y.; Kang, H.J. Association of DNA methylation levels with tissue-specific expression of adipogenic and lipogenic genes in longissimus dorsi muscle of Korean cattle. Asian-Australas. J. Anim. Sci. 2014, 27, 1493–1498. [Google Scholar] [CrossRef]
- Cho, S.H.; Kang, S.M.; Seong, P.N.; Kang, G.H.; Kim, Y.; Kim, J.H.; Chang, S.S.; Park, B.Y. Effect of aging and freezing conditions on meat quality and storage stability of 1++ grade Hanwoo steers: Implication for shelf-life. Korea J. Food Sci. Anim. Resour. 2017, 37, 440–448. [Google Scholar] [CrossRef] [Green Version]
- Utama, D.T.; Lee, C.W.; Park, Y.S.; Jang, A.; Lee, S.K. Comparison of meat quality, fatty acid composition and aroma volatiles of Chikso and Hanwoo beef. Asian-Australas. J. Anim. Sci. 2018, 31, 1500–1506. [Google Scholar] [CrossRef] [Green Version]
- Park, B.; Choi, T.J.; Park, M.N.; Oh, S.H. Estimation of environmental effects and genetic parameters of carcass traits on Chikso (Korean brindle cattle). Asian-Australas. J. Anim. Sci. 2020, 33, 525–530. [Google Scholar] [CrossRef] [Green Version]
- Park, M.N.; Choi, T.J.; Park, B.; Lee, S.S.; Choi, J.G.; Cho, K.H.; Yang, C.B.; Lee, J.G.; Choo, H.J.; Mahboob, A.; et al. Distribution of Chikso (Korean Brindle Cattle) in south Korea and their coat color expression. Korean J. Int. Agric. 2016, 28, 407–413. [Google Scholar] [CrossRef]
- Lim, D.; Strucken, E.M.; Choi, B.H.; Chai, H.H.; Cho, Y.M.; Jang, G.W.; Kim, T.H.; Gondro, C.; Lee, S.H. Genomic footprints in selected and unselected beef cattle breeds in Korea. PLoS ONE 2016, 11, e0151324. [Google Scholar] [CrossRef] [Green Version]
- Song, J.S.; Seong, H.S.; Choi, B.H.; Lee, C.W.; Hwang, N.H.; Lim, D.; Lee, J.H.; Kim, J.S.; Kim, J.D.; Park, Y.S.; et al. Genome-wide analysis of Hanwoo and Chikso populations using the BovineSNP50 genotyping array. Genes Genom. 2018, 40, 1373–1382. [Google Scholar] [CrossRef]
- Korean Hanwoo Beef Cutting Specification; National Institute of Animal Science, Rural Development Administration: Wanju-gun, Republic of Korea, 2018; pp. 11–43.
- Anderson, S.; Aldana, S.; Beggs, M.; Birkey, J.; Conquest, A.; Conway, R.; Hemminger, T.; Herrick, J.; Hurley, C.; Ionita, C.; et al. Determination of fat, moisture, and protein in meat and meat products by using the FOSS FoodScan™ near-infrared spectrophotometer with FOSS artificial neural network calibration model and associated database: Collaborative study. J. AOAC Int. 2007, 90, 1073–1083. [Google Scholar] [CrossRef]
- Hoa, V.B.; Seol, K.H.; Seo, H.W.; Seong, P.N.; Kang, S.M.; Kim, Y.S.; Moon, S.S.; Kim, J.H.; Cho, S.H. Meat quality characteristics of pork bellies in relation to fat level. Anim. Biosci. 2021, 34, 1663–1673. [Google Scholar] [CrossRef] [PubMed]
- Thomas, S.; Kenneth, D.; Livak, J. Analyzing real-time PCR data by comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Gajaweera, C.; Chung, K.Y.; Kwon, E.G.; Hwang, I.H.; Cho, S.H.; Lee, S.H. Evaluation of sensory attributes of Hanwoo Longissimus dorsi muscle and its relationships with intramuscular fat, shear force, and environmental factors. Can. J. Anim. Sci. 2018, 98, 443–452. [Google Scholar] [CrossRef] [Green Version]
- Korea Institute of Animal Products Quality Evaluation (KAPE). The Beef Carcass Grading. Public Notion No. 2017.3.13. 2017. Available online: http://www.ekapepia.or.kr/view/eng/system/beef.asp (accessed on 10 October 2022).
- Gotoh, T.; Albrecht, E.; Teuscher, F.; Kawabata, K.; Sakashita, K.; Iwamoto, H.; Wegner, J. Differences in muscle and fat accretion in Japanese Black and European cattle. Meat Sci. 2009, 82, 300–308. [Google Scholar] [CrossRef] [PubMed]
- Jo, C.; Cho, S.H.; Chang, J.; Nam, K.C. Keys to production and processing of Hanwoo beef: A perspective of tradition and science. Anim. Front. 2012, 2, 32–38. [Google Scholar] [CrossRef]
- Gotoh, T.; Joo, S.T. Characteristics and health benefit of highly marbled Wagyu and Hanwoo beef. Korean J. Food Sci. Anim. Resour. 2016, 36, 709–718. [Google Scholar] [CrossRef] [Green Version]
- Konarska, M.; Kuchida, K.; Tarr, G.; Polkinghorne, R.J. Relationships between marbling measures across principal muscles. Meat Sci. 2017, 123, 67–78. [Google Scholar] [CrossRef]
- Nguyen, D.V.; Nguyen, O.C.; Malau-Aduli, A.E.O. Main regulatory factors of marbling level in beef cattle. Vet. Anim. Sci. 2021, 14, 100219. [Google Scholar] [CrossRef]
- Ministry of Food and Drug Safety. Method for Labeling and Division of Beef and Pork Meat; Notice No. 2014-116; Ministry of Food and Drug Safety: Cheongju, Republic of Korea, 2014.
- Clinquart, A.; Ellies-Oury, M.P.; Hocquette, J.F.; Guillier, L.; Sante-Lhoutellier, V.; Prache, D. On-farm and processing factors affecting bovine carcass and meat quality. Review. Animals 2022, 16, 100426. [Google Scholar] [CrossRef]
- Hoa, V.B.; Oliveros, C.M.; Park, K.M.; Dashdorj, D.; Hwang, I.H. Effect of marbling and chilled ageing on meat-quality traits, volatile compounds and sensory characteristics of beef longissimus dorsi muscle. Anim. Prod. Sci. 2017, 57, 981–992. [Google Scholar]
- Hwang, Y.H.; Joo, S.T. Fatty acid profiles of ten muscles from high and low marbled (quality grade 1++ and 2) Hanwoo beef. Korean J. Food Sci. Anim. Resour. 2016, 36, 679–688. [Google Scholar] [CrossRef] [Green Version]
- Okumura, T.; Saito, K.; Nade, T.; Misumi, S.; Masuda, Y.; Nakayama, H.S.S.; Fujita, K.; Kawamura, T. Effects of Intramuscular Fat on the Sensory Characteristics of M. longissimus dorsi in Japanese Black Steers as Judged by a Trained Analytical Panel. Asian-Australas. J. Anim. Sci. 2007, 20, 577–581. [Google Scholar] [CrossRef]
- Lee, S.H.; Park, B.H.; Sharma, A.; Dang, C.G.; Lee, S.S.; Choi, T.J.; Choy, Y.H.; Kim, H.C.; Jeon, K.J.; Kim, S.D.; et al. Hanwoo cattle: Origin, domestication, breeding strategies and genomic selection. J. Anim. Sci. Technol. 2014, 56, 2. [Google Scholar] [CrossRef] [Green Version]
- Malgwi, I.H.; Halas, V.; Grünvald, P.; Schiavon, S.; Jócsák, I. Genes Related to Fat Metabolism in Pigs and Intramuscular Fat Content of Pork: A Focus on Nutrigenetics and Nutrigenomics. Animals 2022, 12, 150. [Google Scholar] [CrossRef]
- Gao, Y.Y.; Cheng, G.; Cheng, Z.X.; Bao, C.; Yamada, T.; Cao, G.F.; Bao, S.Q.; Schreurs, N.M.; Zan, L.S.; Tong, B. Association of variants in FABP4, FASN, SCD, SREBP1 and TCAP genes with intramuscular fat, carcass traits and body size in Chinese Qinchuan cattle. Meat Sci. 2022, 192, 108882. [Google Scholar] [CrossRef]
- Liu, R.; Liu, X.; Bai, X.; Xiao, C.; Dong, Y. Different expression of lipid metabolism-related genes in Shandong black cattle and Luxi cattle based on transcriptome analysis. Sci. Rep. 2020, 10, 21915. [Google Scholar] [CrossRef]
- Poklukar, K.; Candek-Potokar, M.; Lukac, N.B.; Tomažin, U.; Škrlep, M. Lipid Deposition and Metabolism in local and modern pig breeds: A Review. Animals 2020, 10, 424. [Google Scholar] [CrossRef] [Green Version]
- Wei, X.; Han, S.; Wang, S.; Zheng, Q.; Li, X.; Du, J.; Zhao, J.; Li, F.; Ma, Y. ANGPTL8 regulates adipocytes differentiation and adipogenesis in bovie. Gene 2019, 707, 93–99. [Google Scholar] [CrossRef]
- Lee, S.H.; van der Werf, J.H.J.; Lee, S.H.; Park, E.W.; Oh, S.J.; Gibson, J.P.; Thompson, J.M. Genetic polymorphisms of the bovine fatty acid binding protein 4 gene are significantly associated with marbling and carcass weight in Hanwoo (Korean Cattle). Anim. Genet. 2010, 41, 442–444. [Google Scholar]
- Wang, H.; Wang, J.; Yang, D.D.; Liu, Z.L.; Zeng, Y.Q.; Chen, W. Expression of lipid metabolism genes provides new insights into intrmuscular fat deposition in Laiwu pigs. Asian-Australas. J. Anim. Sci. 2020, 33, 390–397. [Google Scholar] [CrossRef]
- Cho, S.H.; Lee, W.Y.; Seol, K.H.; Kim, Y.S.; Kang, S.M.; Seo, H.W.; Jung, Y.B.; Kim, J.H.; Hoa, V.B. Comparison of storage stability, volatile compounds and sensory properties between coarsely-and finely-marbled 1+ grade Hanwoo beef loins. Food Sci. Anim. Resour. 2020, 40, 497–511. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.H.; Seol, K.H.; Kang, S.M.; Kim, Y.S.; Seo, H.W.; Lee, W.; Kim, J.H.; Hoa, V.B. Comparison of tastes-related components and eating quality between hanwoo steer and cow longissimus thoracis muscles. Food Sci. Anim. Resour. 2020, 40, 908–923. [Google Scholar] [CrossRef] [PubMed]
- Burlingame, B.; Nishida, C.; Uauy, R.; Weisell, R. Fats and fatty acids in human nutrition (Report of a joint FAO/WHO Export Consultation, November, 2008). Ann. Nutr. Metab. 2009, 55, 1–380. [Google Scholar]
- Coleman, L.W.; Hickson, R.E.; Schreurs, N.M.; Martin, N.P.; Kenyon, P.R.; Lopez-Villalobos, N.; Morris, S.T. Carcass characteristics and meat quality of Hereford sired steers born to beef-cross-dairy and Angus breeding cows. Meat Sci. 2016, 121, 403–408. [Google Scholar] [CrossRef]


{kind=link}
| Gene Name | Primer Sequence (5′–3′) * | Accession No. | Product Size (bp) |
|---|---|---|---|
| Lipoprotein lipase (LPL) | Forward: AGCTCCAAGTCGCCTTTCTC | BC118091 | 181 |
| Reverse: TGCAATCACACGGAGAGCTT | |||
| Peroxisome proliferator activated receptor gamma (PPARG) | Forward: ACTTTGGGATCAGCTCCGTG | NM_181024 | 115 |
| Reverse: TCCTCATAGTGCGGAGTGGA | |||
| Acetyl-CoA-carboxylase alpha (ACACA) | Forward: ACGGCTGACTGGAGTTGAAG | AJ132890 | 336 |
| Reverse: AATCGGGAGTGCTGGTTCAG | |||
| Stearoyl CoA desaturase 1 (SCD1) | Forward: TCACATTGATCCCCACCTGC | AF188710 | 962 |
| Reverse: TCACATTGATCCCCACCTGC | |||
| Fatty acid binding protein 4 (FABP4) | Forward: GCATGGCCAAACCCACTTTG | NM_174314 | 107 |
| Reverse: TTCCTGGCCCAATTTGAAGGA | |||
| Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) | Forward: GGTCACCAGGGCTGCTTTTA | NM_001034034 | 222 |
| Reverse: CCAGCATCACCCCACTTGAT |
| Traits | Chikso (n = 6) | Hanwoo (n = 6) |
|---|---|---|
| Slaughter weight (kg) | 679.92 ± 76.04 | 745.83 ± 78.70 |
| Hot carcass weight (kg) | 423.58 ± 50.28 | 466.29 ± 60.31 |
| Cold carcass weight (kg) | 417.30 ± 50.02 | 459.83 ± 60.30 |
| Dressing rate (%) | 62.30 ± 5.12 | 62.60 ± 5.61 |
| Back-fat thickness (mm) at 12th rib | 14.50 ± 6.66 | 12.00 ± 4.73 |
| Marbling score (1–9) * | 2.83 ± 1.83 b | 6.33 ± 2.07 a |
| Ribeye area (cm2) | 87.00 ± 9.44 | 88.83 ± 4.22 |
| Trimmed fat (kg) | 120.59 ± 32.80 | 137.40 ± 31.07 |
| Trimmed fat (%) | 28.53 ± 5.21 | 29.61 ± 3.21 |
| Bone (kg) | 67.29 ± 6.60 | 73.05 ± 6.42 |
| Bone (%) | 16.27 ± 2.21 | 15.97 ± 0.92 |
| Sub-Primal Cuts | Meat Yield in Weight (kg) | Meat Yield in Percentage (%) | ||
|---|---|---|---|---|
| Chikso | Hanwoo | Chikso | Hanwoo | |
| Tenderloin | 7.05 ± 0.48 | 7.40 ± 0.65 | 1.70 ± 0.13 | 1.62 ± 0.13 |
| Upper loin (1st–5th) | 15.74 ± 1.84 | 16.62 ± 0.88 | 3.79 ± 0.39 | 3.65 ± 0.39 |
| Middle loin (6th–9th) | 10.75 ± 1.36 | 11.96 ± 1.70 | 2.58 ± 0.16 | 2.60 ± 0.19 |
| Lower loin (10th–13th) | 7.54 b ± 0.65 | 8.52 a ± 0.51 | 1.82 ± 0.23 | 1.89 ± 0.32 |
| Chuck flap | 4.12 b ± 0.57 | 4.89 a ± 0.46 | 1.00 ± 0.17 | 1.08 ± 0.16 |
| Striploin | 9.57 b ± 0.62 | 11.11 a ± 1.39 | 2.31 ± 0.22 | 2.43 ± 0.32 |
| Chuck roll | 20.24 ± 1.67 | 19.08 ± 3.13 | 4.87 a ± 0.29 | 4.14 b ± 0.19 |
| Chuck tender | 3.37 ± 0.29 | 3.53 ± 0.46 | 0.81 ± 0.10 | 0.77 ± 0.05 |
| Oyster blade | 4.40 ± 0.41 | 5.01 ± 0.65 | 1.06 ± 0.07 | 1.09 ± 0.09 |
| Bolar blade | 13.89 ± 1.00 | 14.33 ± 1.29 | 3.36 ± 0.35 | 3.14 ± 0.23 |
| Rib blade | 1.97 ± 0.23 | 1.93 ± 0.42 | 0.48 ± 0.05 | 0.42 ± 0.09 |
| Upper oyster blade | 2.68 ± 0.28 | 2.64 ± 0.27 | 0.65 ± 0.10 | 0.58 ± 0.03 |
| Top round | 19.94 ± 1.52 | 19.74 ± 1.48 | 4.82 ± 0.56 | 4.33 ± 0.38 |
| Eye of round | 4.56 ± 0.37 | 4.61 ± 0.61 | 1.10 a ± 0.06 | 1.00 b ± 0.07 |
| Rump | 11.09 ± 0.73 | 11.71 ± 0.83 | 2.67 ± 0.22 | 2.56 ± 0.18 |
| Outside round | 13.93 ± 1.06 | 14.55 ± 1.28 | 3.36 ± 0.36 | 3.19 ± 0.25 |
| Outside round head | 4.68 ± 0.46 | 4.20 ± 0.42 | 1.13 a ± 0.16 | 0.92 b ± 0.09 |
| Knuckle | 11.14 b ± 1.18 | 12.83 a ± 1.15 | 2.69 ± 0.29 | 2.81 ± 0.29 |
| Tri-tip | 2.71 ± 0.15 | 2.95 ± 0.24 | 0.65 ± 0.05 | 0.65 ± 0.09 |
| Brisket point end | 22.41 ± 1.92 | 24.81 ± 4.59 | 5.39 ± 0.28 | 5.36 ± 0.35 |
| Brisket point end-deckle off | 5.36 ± 1.18 | 6.30 ± 1.13 | 1.28 ± 0.22 | 1.36 ± 0.09 |
| Short blade | 8.56 ± 0.97 | 8.02 ± 1.41 | 2.06 a ± 0.22 | 1.74 b ± 0.18 |
| Inside skirt | 1.64 a ± 0.18 | 1.21 b ± 0.07 | 0.40 a ± 0.08 | 0.27 b ± 0.03 |
| Thin flank | 11.94 ± 1.87 | 14.59 ± 2.30 | 2.85 ± 0.17 | 3.17 ± 0.31 |
| Internal flank plate | 3.75 ± 0.48 | 3.98 ± 0.62 | 0.90 ± 0.08 | 0.86 ± 0.05 |
| Flank steak | 2.80 ± 0.82 | 3.56 ± 0.67 | 0.66 ± 0.13 | 0.78 ± 0.12 |
| Fore shank | 8.44 ± 0.81 | 8.62 ± 1.09 | 2.04 ± 0.28 | 1.88 ± 0.12 |
| Hind shank | 8.88 ± 0.84 | 9.60 ± 0.99 | 2.14 ± 0.23 | 2.10 ± 0.12 |
| Heel meat | 4.35 ± 0.38 | 4.39 ± 0.41 | 1.05 ± 0.12 | 0.96 ± 0.09 |
| Heel meat center | 0.75 ± 0.06 | 0.82 ± 0.12 | 0.18 ± 0.02 | 0.18 ± 0.02 |
| Conical meat | 1.35 ± 0.16 | 1.46 ± 0.21 | 0.33 ± 0.06 | 0.32 ± 0.02 |
| Chuck short rib(1st–5th) | 6.94 ± 0.61 | 7.69 ± 0.99 | 1.67 ± 0.11 | 1.67 ± 0.07 |
| Short rib (6th–8th) | 11.60 ± 0.60 | 12.74 ± 1.60 | 2.80 ± 0.20 | 2.77 ± 0.03 |
| Short rib (9th–13th) | 8.97 ± 0.81 | 9.97 ± 1.35 | 2.16 ± 0.21 | 2.17 ± 0.14 |
| Rib finger | 18.96 ± 1.51 | 20.89 ± 2.51 | 4.56 ± 0.19 | 4.55 ± 0.14 |
| Hanging tender | 1.04 ± 0.11 | 1.16 ± 0.15 | 0.25 ± 0.03 | 0.25 ± 0.02 |
| Outside skirt | 1.56 ± 0.19 | 1.75 ± 0.27 | 0.37 ± 0.02 | 0.38 ± 0.03 |
| Neck chain | 0.91 ± 0.05 | 0.92 ± 0.13 | 0.22 ± 0.03 | 0.20 ± 0.02 |
| Total meat yield | 237.50 ± 19.54 | 247.44 ± 26.22 | 57.31 ± 5.72 | 53.99 ± 2.45 |
| Composition | M. Longissimus Lumborum | M. Semimembranosus | ||
|---|---|---|---|---|
| Chikso | Hanwoo | Chikso | Hanwoo | |
| Protein (%) | 17.19 ± 1.03 | 17.18 ± 0.52 | 17.21 b ± 0.49 | 20.43 a ± 0.10 |
| Moisture (%) | 67.74 a ± 1.29 | 56.61 b ± 1.83 | 70.02 a ± 0.87 | 67.49 b ± 0.45 |
| Intramuscular fat (%) | 8.83 b ± 1.92 | 23.96 a ± 2.58 | 5.44 b ± 0.92 | 8.34 a ± 0.89 |
| Collagen (%) | 1.25 b ± 0.20 | 2.21 a ± 0.13 | 1.25 b ± 0.14 | 1.78 a ± 0.15 |
| Ash (%) | 3.42 a ± 0.27 | 1.97 b ± 0.07 | 3.58 a ± 0.20 | 2.35 b ± 0.11 |
| Fatty Acids | M. Longissimus Lumborum | M. Semimembranosus | ||
|---|---|---|---|---|
| Chikso | Hanwoo | Chikso | Hanwoo | |
| Myristic acid (C14:0) (%) | 3.43 ± 0.28 | 4.30 ± 0.28 | 2.79 b ± 0.20 | 3.55 a ± 0.23 |
| Palmitic acid (C16:0) (%) | 28.18 b ± 0.52 | 31.18 a ± 0.56 | 27.22 b ± 0.52 | 32.51 a ± 0.51 |
| Palmitoleic acid (C16:1n7) (%) | 4.13 ± 0.49 | 4.96 ± 0.17 | 4.07 ± 0.51 | 3.49 ± 0.36 |
| Stearic acid (C18:0) (%) | 11.51 ± 0.61 | 10.87 ± 0.43 | 10.71 ± 0.68 | 11.08 ± 1.02 |
| Cis-Vaccenic acid (C18:1n7) (%) | 0.46 a ± 0.07 | 0.30 b ± 0.03 | 0.49 a ± 0.08 | 0.27 b ± 0.04 |
| Oleic acid (C18:1n9) (%) | 49.33 ± 0.80 | 46.65 ± 0.89 | 50.51 a ± 1.00 | 46.90 b ± 0.49 |
| Linoleic acid (C18:2n6) (%) | 2.37 a ± 0.38 | 1.37 b ± 0.14 | 3.06 ± 0.74 | 1.75 ± 0.17 |
| Linolenic acid (C18:3n3) (%) | 0.09 ± 0.01 | 0.06 ± 0.01 | 0.10 a ± 0.01 | 0.06 b ± 0.01 |
| Gamma linoleic acid (C18:3n6) (%) | 0.02 a ± 0.00 | 0.001 b ± 0.00 | 0.02 a ± 0.00 | 0.01 b ± 0.00 |
| Eicosenoic acid (C20:1n9) (%) | 0.16 ± 0.02 | 0.22 ± 0.02 | 0.20 ± 0.03 | 0.20 ± 0.03 |
| Arachidonic acid (C20:4n6) (%) | 0.27 ± 0.11 | 0.05 ± 0.00 | 0.72 ± 0.29 | 0.15 ± 0.03 |
| Eicosapetaenoic acid (C20:5n3) (%) | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Adrenic acid (C22:4n6) (%) | 0.05 ± 0.01 | 0.04 ± 0.01 | 0.11 ± 0.03 | 0.05 ± 0.01 |
| Docosahexanoic acid (C22:6n3) (%) | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| SFA (%) | 43.12 b ± 0.70 | 46.35 a ± 0.89 | 40.73 b ± 0.69 | 47.13 a ± 0.60 |
| UFA (%) | 56.88 A ± 0.70 | 53.66 B ± 0.89 | 59.27 A ± 0.69 | 52.87 B ± 0.60 |
| MUFA (%) | 54.08 ± 0.92 | 52.14 ± 0.97 | 55.27 a ± 1.34 | 50.85 b ± 0.62 |
| PUFA (%) | 2.80 a ± 0.51 | 1.52 b ± 0.15 | 4.01 ± 1.06 a | 2.02 ± 0.20 b |
| n3(%) | 0.09 ± 0.01 | 0.06 ± 0.01 | 0.10 a ± 0.01 | 0.06 b ± 0.01 |
| n6 (%) | 2.72 a ± 0.50 | 1.46 b ± 0.14 | 3.91 ± 1.05 | 1.96 ± 0.19 |
| n6/n3 | 32.40 ± 4.47 | 25.87 ± 1.87 | 37.58 ± 8.03 | 32.25 ± 2.14 |
| MUFA/SFA | 1.26 ± 0.04 | 1.13 ± 0.04 | 1.36 A ± 0.05 | 1.08 B ± 0.03 |
| PUFA/SFA | 0.07 a ± 0.01 | 0.03 b ± 0.00 | 0.10 ± 0.03 | 0.04 ± 0.00 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hoa, V.-B.; Song, D.-H.; Seol, K.-H.; Kang, S.-M.; Kim, H.-W.; Bae, I.-S.; Kim, E.-S.; Park, Y.-S.; Cho, S.-H. A Comparative Study on the Carcass and Meat Chemical Composition, and Lipid-Metabolism-Related Gene Expression in Korean Hanwoo and Brindle Chikso Cattle. Curr. Issues Mol. Biol. 2023, 45, 3279-3290. https://doi.org/10.3390/cimb45040214
Hoa V-B, Song D-H, Seol K-H, Kang S-M, Kim H-W, Bae I-S, Kim E-S, Park Y-S, Cho S-H. A Comparative Study on the Carcass and Meat Chemical Composition, and Lipid-Metabolism-Related Gene Expression in Korean Hanwoo and Brindle Chikso Cattle. Current Issues in Molecular Biology. 2023; 45(4):3279-3290. https://doi.org/10.3390/cimb45040214
Chicago/Turabian StyleHoa, Van-Ba, Dong-Heon Song, Kuk-Hwan Seol, Sun-Moon Kang, Hyun-Wook Kim, In-Seon Bae, Eun-Sung Kim, Yeon-Soo Park, and Soo-Hyun Cho. 2023. "A Comparative Study on the Carcass and Meat Chemical Composition, and Lipid-Metabolism-Related Gene Expression in Korean Hanwoo and Brindle Chikso Cattle" Current Issues in Molecular Biology 45, no. 4: 3279-3290. https://doi.org/10.3390/cimb45040214