
Potential Effects of High Temperature and Heat Wave on Nanorana pleskei Based on Transcriptomic Analysis
 , , ,
, , , 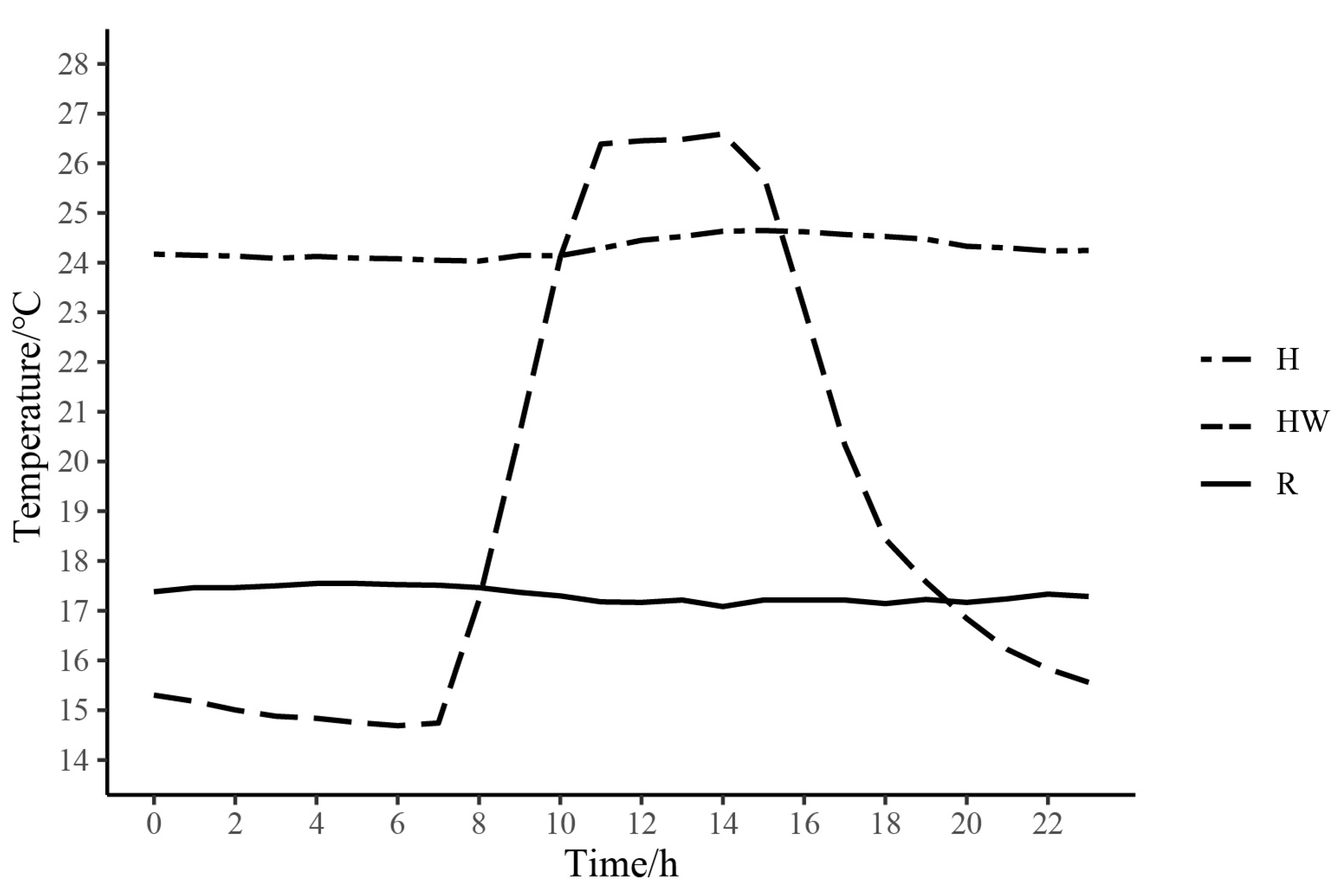{kind=link}
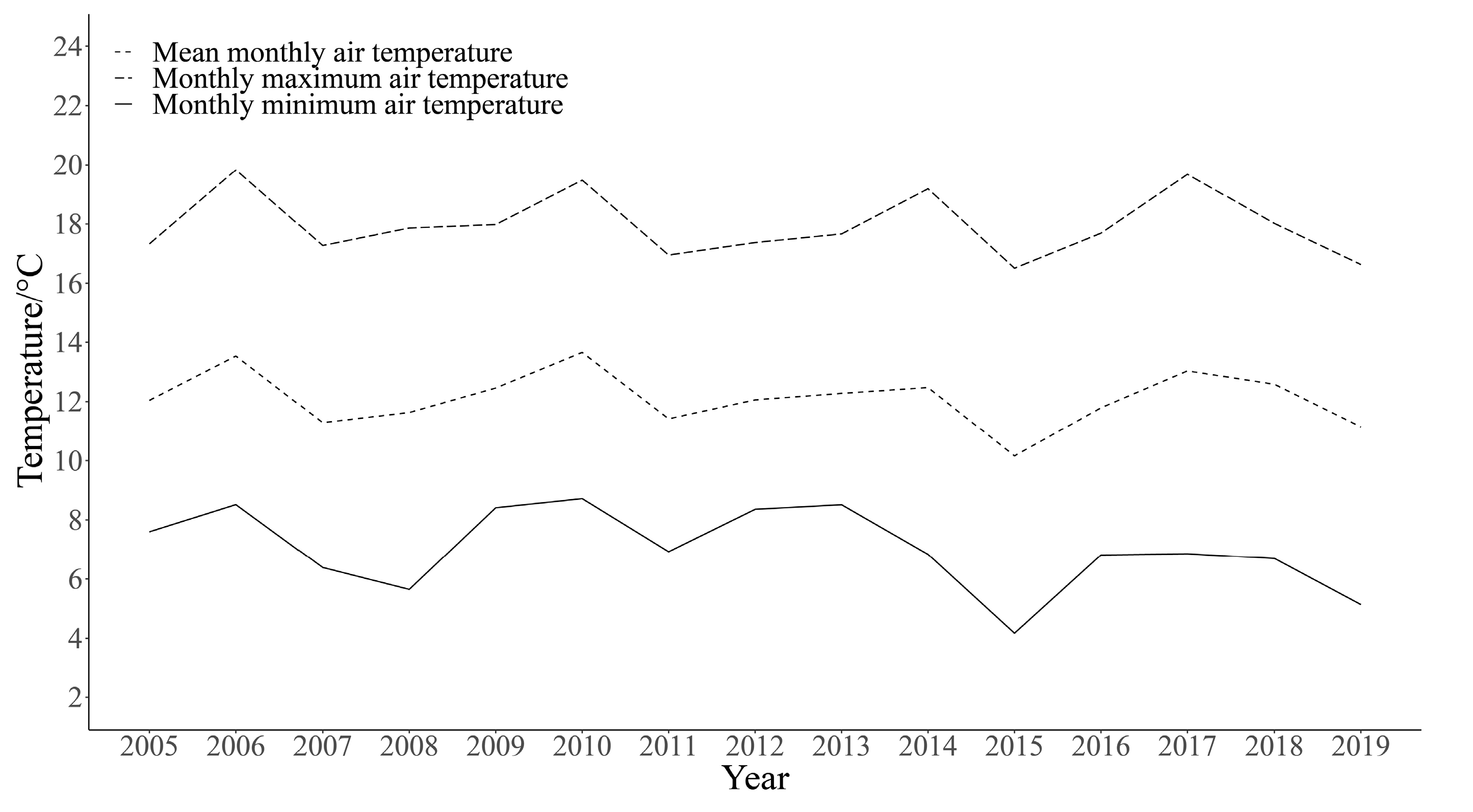{kind=link}
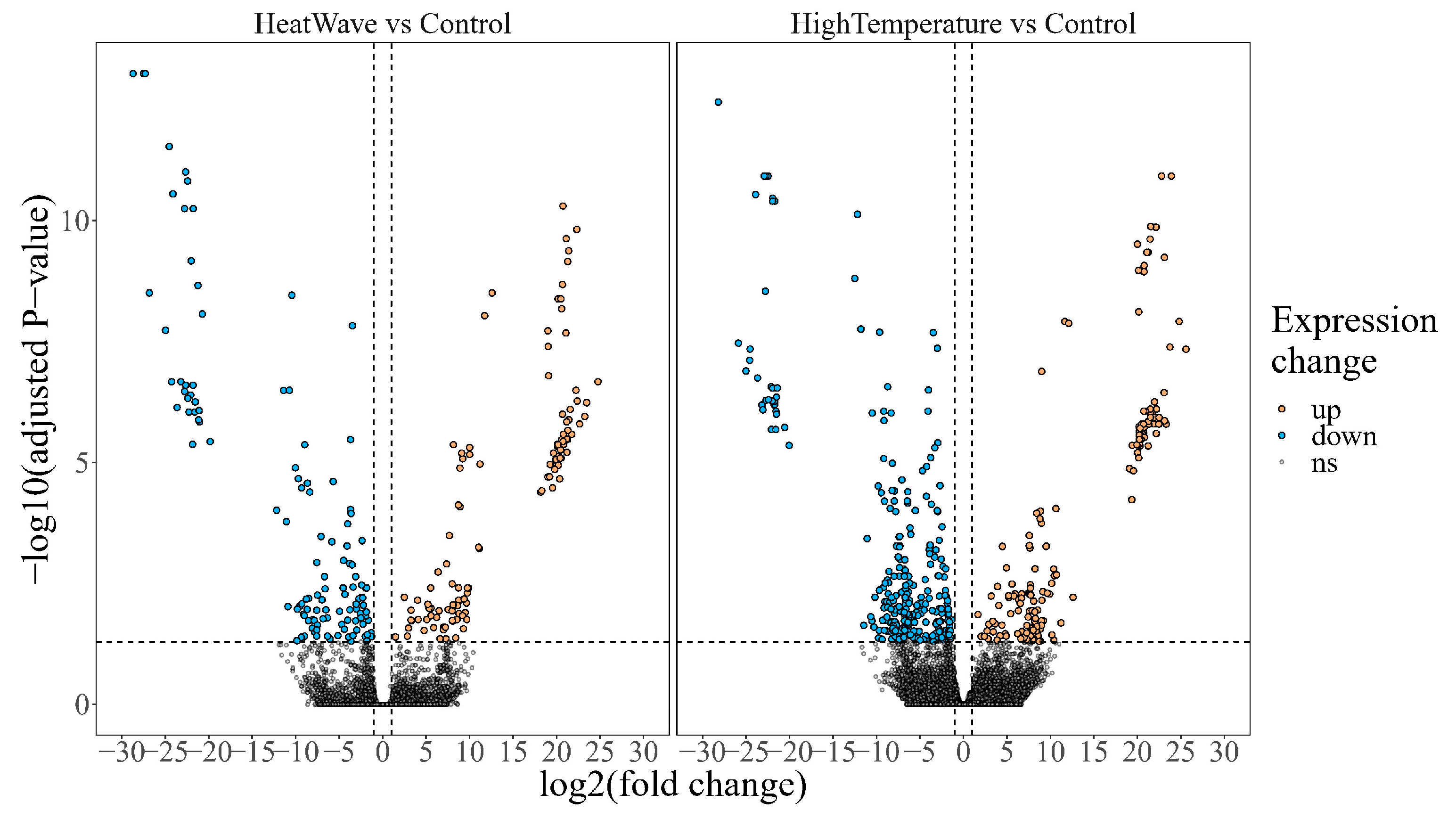{kind=link}
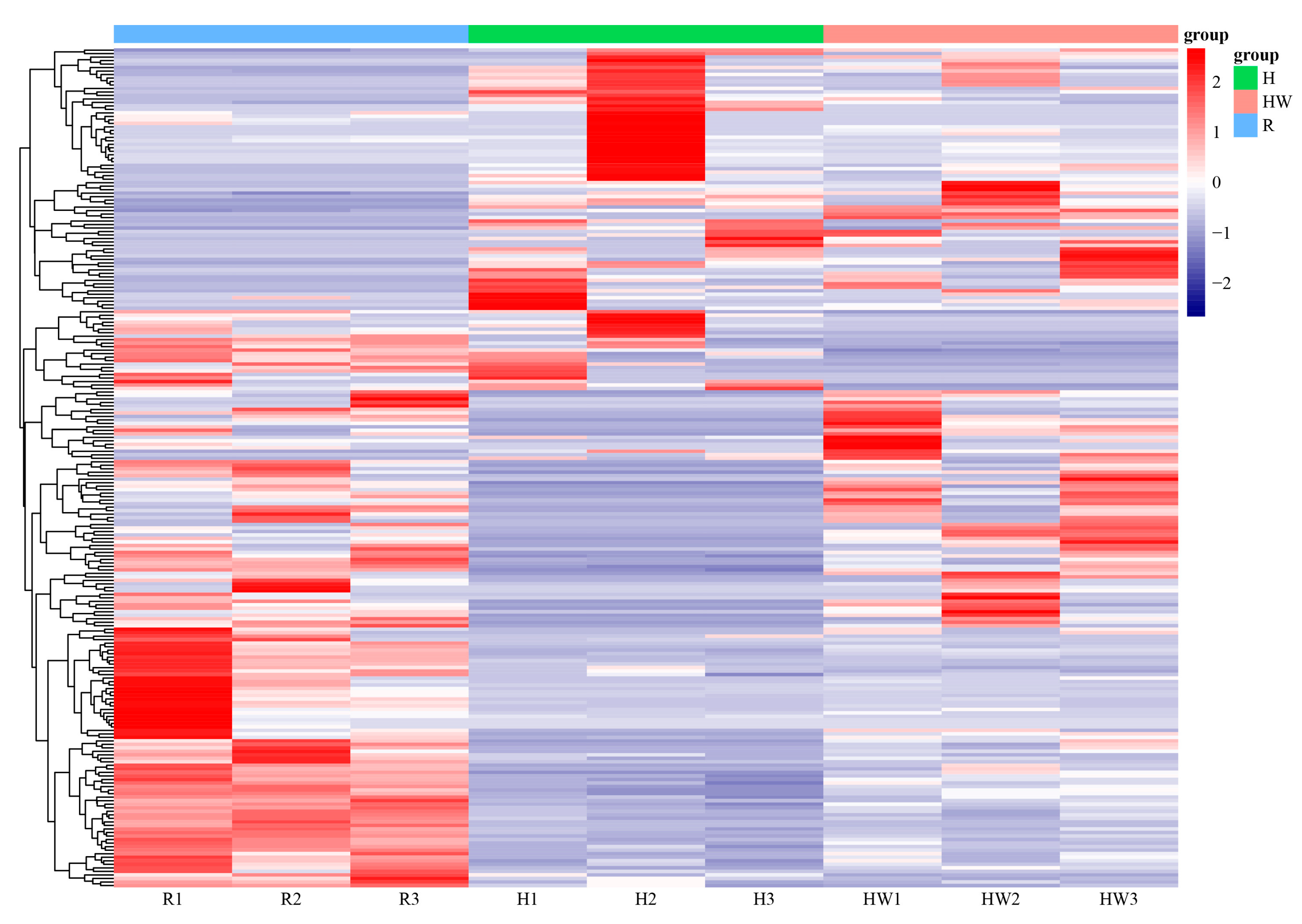{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Acclimation
2.2. Assembly and Evaluation
2.3. Function Annotation
2.4. Analysis of Differentially Expressed Genes and Functional Enrichment
3. Results
3.1. De Novo Assembly and Annotation
3.2. Differential Gene Expression and Pathway Enrichment
4. Discussion
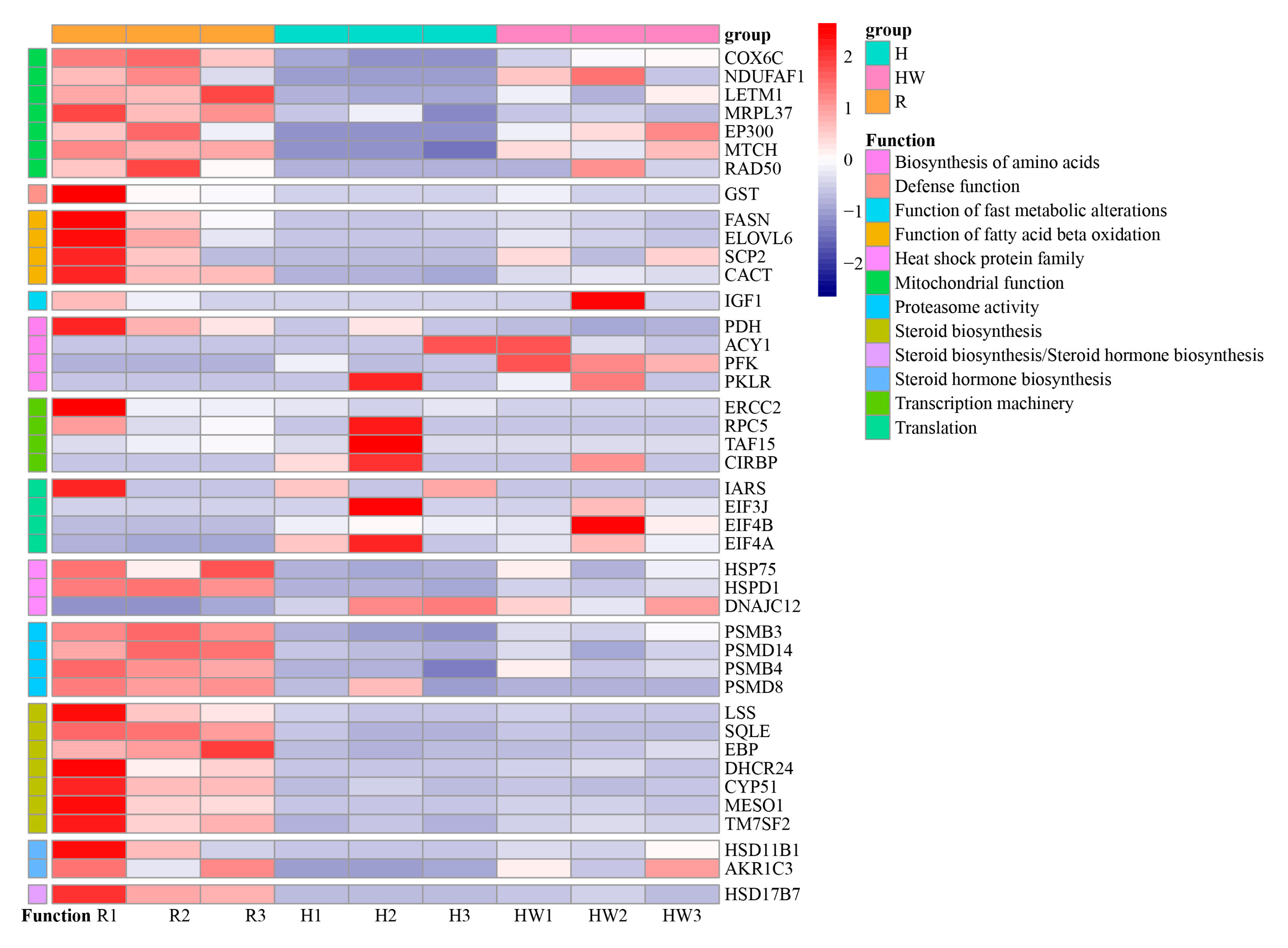
4.1. Treatment-Specific Genes and KEGG Pathways
4.2. Shared Genes and KEGG Pathways
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Crump, M.L. Anuran Reproductive Modes: Evolving Perspectives. J. Herpetol. 2015, 49, 1–16. [Google Scholar] [CrossRef]
- Nunes-de-Almeida, C.H.L.; Haddad, C.F.B.; Toledo, L.F. A revised classification of the amphibian reproductive modes. Salamandra 2021, 57, 413–427. [Google Scholar]
- Stuart, S.N.; Chanson, J.S.; Cox, N.A.; Young, B.E.; Rodrigues, A.S.L.; Fischman, D.L.; Waller, R.W. Status and Trends of Amphibian Declines and Extinctions Worldwide. Science 2004, 306, 1783–1786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rohr, J.R.; Palmer, B.D. Climate change, multiple stressors, and the decline of ectotherms. Conserv. Biol. 2013, 27, 741–751. [Google Scholar] [CrossRef]
- Solomon, S.; Manning, M.; Marquis, M.; Qin, D. Climate Change 2007—The Physical Science Basis: Working Group I Contribution to the Fourth Assessment Report of the IPCC; Cambridge University Press: Cambridge, UK, 2007; Volume 4. [Google Scholar]
- Chapman, S.; Mustin, K.; Renwick, A.R.; Segan, D.B.; Hole, D.G.; Pearson, R.G.; Watson, J.E.M. Publishing trends on climate change vulnerability in the conservation literature reveal a predominant focus on direct impacts and long time-scales. Divers. Distrib. 2014, 20, 1221–1228. [Google Scholar] [CrossRef]
- Barry, J.P.; Baxter, C.H.; Sagarin, R.D.; Gilman, S.E. Climate-related, long-term faunal changes in a California rocky intertidal community. Science 1995, 267, 672–675. [Google Scholar] [CrossRef]
- Easterling, D.R.; Meehl, G.A.; Parmesan, C.; Changnon, S.A.; Karl, T.R.; Mearns, L.O. Climate extremes: Observations, modeling, and impacts. Science 2000, 289, 2068–2074. [Google Scholar] [CrossRef] [Green Version]
- Perry, A.L.; Low, P.J.; Ellis, J.R.; Reynolds, J.D. Climate change and distribution shifts in marine fishes. Science 2005, 308, 1912–1915. [Google Scholar] [CrossRef]
- Thomas, C.D. Climate, climate change and range boundaries. Divers. Distrib. 2010, 16, 488–495. [Google Scholar] [CrossRef]
- Sheridan, J.A.; Bickford, D. Shrinking body size as an ecological response to climate change. Nat. Clim. Change 2011, 1, 401–406. [Google Scholar] [CrossRef]
- Goodman, R.E.; Lebuhn, G.; Seavy, N.E.; Gardali, T.; Bluso-Demers, J.D. Avian body size changes and climate change: Warming or increasing variability? Glob. Change Biol. 2012, 18, 63–73. [Google Scholar] [CrossRef]
- Duan, R.-Y.; Kong, X.-Q.; Huang, M.-Y.; Varela, S.; Ji, X. The potential effects of climate change on amphibian distribution, range fragmentation and turnover in China. PeerJ 2016, 4, e2185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urban, M.C.; Richardson, J.L.; Freidenfelds, N.A. Plasticity and genetic adaptation mediate amphibian and reptile responses to climate change. Evol. Appl. 2014, 7, 88–103. [Google Scholar] [CrossRef] [PubMed]
- Frich, P.; Alexander, L.V.; Della-Marta, P.; Gleason, B.; Haylock, M.; Tank, A.K.; Peterson, T. Observed coherent changes in climatic extremes during the second half of the twentieth century. Clim. Res. 2002, 19, 193–212. [Google Scholar] [CrossRef] [Green Version]
- You, Q.; Jiang, Z.; Kong, L.; Wu, Z.; Bao, Y.; Kang, S.; Pepin, N. A comparison of heat wave climatologies and trends in China based on multiple definitions. Clim. Dyn. 2017, 48, 3975–3989. [Google Scholar] [CrossRef] [Green Version]
- Alvarez, M.; Schrey, A.W.; Richards, C.L. Ten years of transcriptomics in wild populations: What have we learned about their ecology and evolution? Mol. Ecol. 2015, 24, 710–725. [Google Scholar] [CrossRef]
- Todd, E.V.; Black, M.A.; Gemmell, N.J. The power and promise of RNA-seq in ecology and evolution. Mol. Ecol. 2016, 25, 1224–1241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Logan, M.L.; Cox, C.L. Genetic Constraints, Transcriptome Plasticity, and the Evolutionary Response to Climate Change. Front. Genet. 2020, 11, 538226. [Google Scholar] [CrossRef]
- McCartney-Melstad, E.; Shaffer, H.B. Amphibian molecular ecology and how it has informed conservation. Mol. Ecol. 2015, 24, 5084–5109. [Google Scholar] [CrossRef]
- Song, J.; McDowell, J.R. Comparative transcriptomics of spotted seatrout (Cynoscion nebulosus) populations to cold and heat stress. Ecol. Evol. 2021, 11, 1352–1367. [Google Scholar] [CrossRef]
- Fan, W.; Huang, M.; Xu, J.; Zhang, M.; Jiang, Y.; Cai, M.; Sun, H.; Li, X. Transcriptome analysis reveals the effects of temperature on growth in tadpoles of spiny-bellied frog (Quasipaa boulengeri). Aquac. Int. 2021, 29, 925–939. [Google Scholar] [CrossRef]
- Komaki, S.; Matsunami, M.; Lin, J.-W.; Lee, K.-H.; Lin, Y.-P.; Lee, Y.; Lin, S.-M.; Igawa, T. Transcriptomic Changes in Hot Spring Frog Tadpoles (Buergeria otai) in Response to Heat Stress. Front. Ecol. Evol. 2021, 9, 658. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, B.; Nie, X.; Liu, Q.; Xie, F.; Shang, D. Transcriptome Analysis and Identification of Genes Related to Immune Function in Skin of the Chinese Brown Frog. Zool. Sci. 2009, 26, 80–86. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Liu, Z.; Quan, J.; Li, L.; Zhao, G.; Lu, J. RNA-seq Analysis Reveals Alternative Splicing Under Heat Stress in Rainbow Trout (Oncorhynchus mykiss). Mar. Biotechnol. 2022, 24, 5–17. [Google Scholar] [CrossRef]
- Wu, X.; Zhang, C.; Griffin, J.N.; Sun, S. The brown-world role of insectivores: Frogs reduce plant growth by suppressing detritivores in an alpine meadow. Basic Appl. Ecol. 2014, 15, 66–74. [Google Scholar] [CrossRef]
- Zhang, J.; Li, Y.; Dai, Q.; Wang, Y. Effects of Grazing on Diet of Anurans in Zoige Wetlands, Sichuan, China. Chin. J. Appl. Environ. Biol. 2010, 16, 683–687. [Google Scholar] [CrossRef]
- Fei, L.; Hu, S.; Ye, C.; Huang, Y. Fauna Sinica. Amphibia; Volume 2. Anura; Chinese Academy of Science Press: Beijing, China, 2009. [Google Scholar]
- Dai, J.-H.; Dai, Q.; Zhang, M.; Zhang, J.-D.; Li, C.; Liu, B.; Liu, Z.-J.; Wang, Y.-Z. Habitat selection of three amphibians (Rana kukunoris, Nanorana pleskei, Bufo minshanicus) during summer-autumn in Zoige Wetland National Nature Reserve. Zool. Res. 2005, 26, 263–271. [Google Scholar]
- Wang, G.; Gong, Y.; Han, J.; Li, C.; Xie, F. Oviposition Site Selection of the Plateau Frog (Nanorana pleskei) in the Zoige Wetland, China. Asian Herpetol. Res. 2017, 8, 269–274. [Google Scholar] [CrossRef]
- Yang, W.; Lu, B.; Fu, J. Molecular Convergent Evolution of the MYBPC2 Gene Among Three High-Elevation Amphibian Species. J. Mol. Evol. 2017, 84, 139–143. [Google Scholar] [CrossRef]
- Zhang, X.; Niu, Y.; Zhang, H.; Xu, T.; Zeng, Q.; Storey, K.B.; Chen, Q. The effect of long-term cold acclimation on redox state and antioxidant defense in the high-altitude frog, Nanorana pleskei. J. Therm. Biol. 2021, 99, 103008. [Google Scholar] [CrossRef]
- He, J.; Tang, X.; Pu, P.; Zhang, T.; Niu, Z.; Meng, F.; Xi, L.; Ma, M.; Wu, J.; Ma, M.; et al. Influence of High Temperatures and Heat Wave on Thermal Biology, Locomotor Performance, and Antioxidant System of High-Altitude Frog Nanorana pleskei Endemic to Qinghai-Tibet Plateau. Front. Ecol. Evol. 2021, 9, 831. [Google Scholar] [CrossRef]
- Heikkila, J.J. Heat shock protein gene expression and function in amphibian model systems. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2010, 156, 19–33. [Google Scholar] [CrossRef] [PubMed]
- Clay, T.A.; Steffen, M.A.; Treglia, M.L.; Torres, C.D.; Trujano-Alvarez, A.L.; Bonett, R.M. Multiple stressors produce differential transcriptomic patterns in a stream-dwelling salamander. BMC Genom. 2019, 20, 482. [Google Scholar] [CrossRef] [PubMed]
- Holson, R.R. Euthanasia by decapitation: Evidence that this technique produces prompt, painless unconsciousness in laboratory rodents. Neurotoxicol. Teratol. 1992, 14, 253–257. [Google Scholar] [CrossRef] [PubMed]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haas, B.J.; Papanicolaou, A.; Yassour, M.; Grabherr, M.; Blood, P.D.; Bowden, J.; Couger, M.B.; Eccles, D.; Li, B.; Lieber, M.; et al. De novo transcript sequence reconstruction from RNA-seq using the Trinity platform for reference generation and analysis. Nat. Protoc. 2013, 8, 1494–1512. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.; Niu, B.; Zhu, Z.; Wu, S.; Li, W. CD-HIT: Accelerated for clustering the next-generation sequencing data. Bioinformatics 2012, 28, 3150–3152. [Google Scholar] [CrossRef] [PubMed]
- Smith-Unna, R.; Boursnell, C.; Patro, R.; Hibberd, J.M.; Kelly, S. TransRate: Reference-free quality assessment of de novo transcriptome assemblies. Genome Res. 2016, 26, 1134–1144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Consortium, T.U. UniProt: A worldwide hub of protein knowledge. Nucleic Acids Res. 2018, 47, D506–D515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, G.; Wang, L.-G.; Han, Y.; He, Q.-Y. clusterProfiler: An R package for comparing biological themes among gene clusters. Omics J. Integr. Biol. 2012, 16, 284–287. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Hu, E.; Xu, S.; Chen, M.; Guo, P.; Dai, Z.; Feng, T.; Zhou, L.; Tang, W.; Zhan, L.; et al. clusterProfiler 4.0: A universal enrichment tool for interpreting omics data. Innovation 2021, 2, 100141. [Google Scholar] [CrossRef]
- Pörtner, H.O. Climate variations and the physiological basis of temperature dependent biogeography: Systemic to molecular hierarchy of thermal tolerance in animals. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2002, 132, 739–761. [Google Scholar] [CrossRef]
- Storey, K.B.; Storey, J.M. Molecular Physiology of Freeze Tolerance in Vertebrates. Physiol. Rev. 2017, 97, 623–665. [Google Scholar] [CrossRef] [Green Version]
- Scarpulla, R.C.; Vega, R.B.; Kelly, D.P. Transcriptional integration of mitochondrial biogenesis. Trends Endocrinol. Metab. 2012, 23, 459–466. [Google Scholar] [CrossRef] [Green Version]
- Popov, L.-D. Mitochondrial biogenesis: An update. J. Cell. Mol. Med. 2020, 24, 4892–4899. [Google Scholar] [CrossRef] [Green Version]
- Strange, R.C.; Spiteri, M.A.; Ramachandran, S.; Fryer, A.A. Glutathione-S-transferase family of enzymes. Mutat. Res./Fundam. Mol. Mech. Mutagen. 2001, 482, 21–26. [Google Scholar] [CrossRef]
- Leiers, B.; Kampkötter, A.; Grevelding, C.G.; Link, C.D.; Johnson, T.E.; Henkle-Dührsen, K. A stress-responsive glutathione S-transferase confers resistance to oxidative stress in Caenorhabditis elegans. Free. Radic. Biol. Med. 2003, 34, 1405–1415. [Google Scholar] [CrossRef]
- Li, B.; Ma, Y.; Zhang, Y.H. Oxidative stress and hepatotoxicity in the frog, Rana chensinensis, when exposed to low doses of trichlorfon. J. Environ. Sci. Health Part B 2017, 52, 476–482. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.; Chen, Q.; Storey, K.B.; Teng, L.; Li, X.; Xu, T.; Zhang, H. Physiological Ecology of Winter Hibernation by the High-Altitude Frog Nanorana parkeri. Physiol. Biochem. Zool. 2022, 95, 201–211. [Google Scholar] [CrossRef] [PubMed]
- Poli, G.; Albano, E.; Dianzani, M.U. The role of lipid peroxidation in liver damage. Chem. Phys. Lipids 1987, 45, 117–142. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B.; Chirico, S. Lipid peroxidation: Its mechanism, measurement, and significance. Am. J. Clin. Nutr. 1993, 57, 715S–725S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Dell, S.D.; Day, I.N.M. Molecules in focus Insulin-like growth factor II (IGF-II). Int. J. Biochem. Cell Biol. 1998, 30, 767–771. [Google Scholar] [CrossRef] [PubMed]
- Patel, M.; Korotchkina, L. Regulation of the pyruvate dehydrogenase complex. Biochem. Soc. Trans. 2006, 34, 217–222. [Google Scholar] [CrossRef]
- Liao, Y.; Tong, L.; Tang, L.; Wu, S. The role of cold-inducible RNA binding protein in cell stress response. Int. J. Cancer 2017, 141, 2164–2173. [Google Scholar] [CrossRef] [Green Version]
- Ibba, M.; Söll, D. Aminoacyl-tRNA synthesis. Annu. Rev. Biochem. 2000, 69, 617–650. [Google Scholar] [CrossRef]
- Zhan, K.; Narasimhan, J.; Wek, R.C. Differential Activation of eIF2 Kinases in Response to Cellular Stresses in Schizosaccharomyces pombe. Genetics 2004, 168, 1867–1875. [Google Scholar] [CrossRef] [Green Version]
- Rupik, W.; Jasik, K.; Bembenek, J.; Widłak, W. The expression patterns of heat shock genes and proteins and their role during vertebrate’s development. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2011, 159, 349–366. [Google Scholar] [CrossRef]
- Ketola-Pirie, C.A.; Atkinson, B.G. Cold- and heat-shock induction of new gene expression in cultured amphibian cells. Can. J. Biochem. Cell Biol. 1983, 61, 462–471. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.-P.; Gu, W.-B.; Tu, D.-D.; Zhu, Q.-H.; Zhou, Y.-L.; Wang, C.; Wang, L.-Z.; Shu, M.-A. Effects of both cold and heat stress on the liver of the giant spiny frog (Quasipaa spinosa): Stress response and histological changes. J. Exp. Biol. 2018, 221, jeb186379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gormally, B.M.G.; Romero, L.M. What are you actually measuring? A review of techniques that integrate the stress response on distinct time-scales. Funct. Ecol. 2020, 34, 2030–2044. [Google Scholar] [CrossRef]
- Tanaka, K. The proteasome: Overview of structure and functions. Proc. Jpn. Acad. Ser. B 2009, 85, 12–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Regnault, C.; Usal, M.; Veyrenc, S.; Couturier, K.; Batandier, C.; Bulteau, A.-L.; Lejon, D.; Sapin, A.; Combourieu, B.; Chetiveaux, M.; et al. Unexpected metabolic disorders induced by endocrine disruptors in Xenopus tropicalis provide new lead for understanding amphibian decline. Proc. Natl. Acad. Sci. USA 2018, 115, E4416–E4425. [Google Scholar] [CrossRef] [Green Version]
- Nagorny, P.; Cichowicz, N. Chapter 9—New Strategy Based on Sequential Michael/Aldol Reactions for the Asymmetric Synthesis of Cardenolides. In Strategies and Tactics in Organic Synthesis; Harmata, M., Ed.; Academic Press: Cambridge, MA, USA, 2016; Volume 12, pp. 237–267. [Google Scholar]
- Hinther, A.; Bromba, C.M.; Wulff, J.E.; Helbing, C.C. Effects of Triclocarban, Triclosan, and Methyl Triclosan on Thyroid Hormone Action and Stress in Frog and Mammalian Culture Systems. Environ. Sci. Technol. 2011, 45, 5395–5402. [Google Scholar] [CrossRef]
- Wilczynski, W.; Burmeister, S.S. Effects of steroid hormones on hearing and communication in frogs. In Hearing and Hormones; Springer: Berlin/Heidelberg, Germany, 2016; pp. 53–75. [Google Scholar]
- Hayes, T.; Chan, R.; Licht, P. Interactions of temperature and steroids on larval growth, development, and metamorphosis in a toad (Bufo boreas). J. Exp. Zool. 1993, 266, 206–215. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, T.; Niu, Z.; He, J.; Pu, P.; Meng, F.; Xi, L.; Tang, X.; Ding, L.; Ma, M.; Chen, Q. Potential Effects of High Temperature and Heat Wave on Nanorana pleskei Based on Transcriptomic Analysis. Curr. Issues Mol. Biol. 2023, 45, 2937-2949. https://doi.org/10.3390/cimb45040192
Zhang T, Niu Z, He J, Pu P, Meng F, Xi L, Tang X, Ding L, Ma M, Chen Q. Potential Effects of High Temperature and Heat Wave on Nanorana pleskei Based on Transcriptomic Analysis. Current Issues in Molecular Biology. 2023; 45(4):2937-2949. https://doi.org/10.3390/cimb45040192
Chicago/Turabian StyleZhang, Tao, Zhiyi Niu, Jie He, Peng Pu, Fei Meng, Lu Xi, Xiaolong Tang, Li Ding, Miaojun Ma, and Qiang Chen. 2023. "Potential Effects of High Temperature and Heat Wave on Nanorana pleskei Based on Transcriptomic Analysis" Current Issues in Molecular Biology 45, no. 4: 2937-2949. https://doi.org/10.3390/cimb45040192